characterization of seed producing pearl millet x elephantgrass hexaploid hybrids
TRANSCRIPT

Euphytica 67: 143-149, 1993. © 1993 Kluwer Academic Publishers. Printed in the Netherlands.
Characterization of seed producing pearl millet x elephantgrass hexaploid hybrids
D.A. Diz & S.C. Schank Department of Agronomy, University of Florida, 2183 McCarty Hall, Gainesville, Florida 32608, USA
Received 26 November 1992; accepted 9 April 1993
Key words: elephantgrass, interspecific hybrid, hexaploid, pearl millet, Pennisetum purpureum, Pennisetum glaucum, seed
Summary
Successful widespread use of a forage or biomass plant depends largely on its ease of establishment. Elephant- grass (Pennisetum purpureum Schum.) is used for both forage and biomass production, due to its high dry matter production, aggressiveness, perenniality, and forage quality. However, lack of high quality seed has limited the use of elephantgrass and its hybrids to pearl millet (P. glaucum L. R.Br.). Development of a seed-propagated pearl millet x elephantgrass hexaploid cultivar, which would combine desirable character- istics from both of these species, would be highly desirable. The objectives of this study were to characterize morphological and seed-related traits from the selfed progeny of seven hexaploid hybrids, and to determine whether seed size had any influence on these traits. Traits studied included number of tillers, height, leaf length and width, panicles per plant, days to flowering, panicle length, seed set, seed production, and weight of 100 seeds. Genetic differences were found among the progeny of the seven hybrids. Differences were also found between plants derived from large- and small-seed lots within families. Plants descended from larger seed had better growth and seed-related characteristics, which resulted in more desirable plants. These hex- aploid hybrids showed potential for direct seeding into the field for biomass or forage production.
Introduction
The successful widespread use of a forage or bio- mass species is dependent upon many factors. One crucial factor is its reproductive behavior, which in- fluences the methodology of propagation used for that species. Elephantgrass (Pennisetum purpu- reum Schum.) is one of the highest dry matter pro- ducing forage species in tropical and subtropical en- vironments. Mislevy et al. (1986) reported dry mat- ter yields of 52.2 Mg ha -~ yr -1 for cv. Kinggrass (PI 300086) at Ona, Florida. Elephantgrass has consid- erable potential as a forage or biomass plant due to its high dry matter production, aggressiveness, pe-
renniality, and forage quality (Schank et al., 1993). However, even a very persistent, high quality culti- var released in Florida, Mott, which produced aver- age cattle gains of 0.97 Kg d -~ during a three yr study of summer grazing (Sollenberger & Jones, 1989), is not being used commercially in the U.S. because of propagation limitations. Elephantgrass is vegeta- tively propagated due to low natural seed produc- tion and poor seed quality. Planting typically con- sists of placing whole stems horizontally in shallow furrows (Sollenberger et al., 1990). Although this species is widely distributed throughout tropical and subtropical regions of the world as a forage grass, its method of propagation has restricted a

144
more widespread use, especially in developed coun- tries where labour is more expensive (Schank & Diz, 1991).
Problems with seed propagation may potentially be solved through hybridization with pearl millet (Pennisetum glaucum L. R.Br.) and further selec- tion. Pearl millet is an annual, high quality, diploid grass (2n = 2x = 14) which is grown as a grain and forage crop. Elephantgrass is a tetraploid (2n = 4x = 28). The F 1 hybrids (triploids: 2n = 3x = 21) are com- pletely sterile due to their unbalanced chromosome number, which causes irregularities during meiosis (Khan & Rahman, 1963; Muldoon & Pearson, 1979). Powell & Burton (1966) suggested a commer- cial method of producing triploid hybrid seed by seeding a cytoplasmic male-sterile pearl millet line between rows of perennial elephantgrass (pollen source). However, as Powell & Burton mention, this would only be possible in areas which are frost free until late December (U.S.A.), due to the late flowering of elephantgrass. Also, elephantgrass rows would still have to be planted vegetatively. Another approach involves doubling the chromo- some number of the triploid hybrid, which then re- stores fertility (Hanna, 1981). The amphiploids (hexaploids: 2n = 6x = 42) obtained usually showed a high degree of regular meiosis, and their progeny had a wide range of pollen and seed fertility. Through selection, a fertile, superior forage or bio- mass hexaploid which would produce adequate amounts of high quality seed may potentially be ob- tained.
An important characteristic commonly evaluat- ed in seed-producing crops is seed size. Small seed size has generally been associated with poor per- formance in a vast number of crops (Lawan et al., 1985; Singh, 1987). Evaluating the effect of seed size on numerous morphological and seed-related char- acteristics would be important for obtaining future seed-propagated cultivars. Fertility parameters in pearl millet × elephantgrass (PMxE) amphiploids have been published (Gonzalez & Hanna, 1984; Jauhar, 1981), and seed and seedling characteristics of seven hexaploid genotypes have been described by Diz & Schank (1990). No other reports on PMxE amphiploid seed characteristics or seed production were found in the literature. This paper will report
the field observations and characterization of the selfed progeny of parental plants which have been described previously (Diz & Schank, 1990).
The objectives of this study were: 1) to character- ize morphological and seed-related traits from the selfed progeny of seven hexaploid hybrids under field conditions; and 2) to determine whether seed size influenced these traits.
Materials and methods
Experimental site
This experiment was conducted at the Dairy Re- search Unit of the University of Florida, Gaines- ville (29 ° 60' N lat.), during 1990. The soil of the ex- perimental site is classified as a Sparr fine sand; a sandy, siliceous, hyperthermic Grossarenic Paleu- dult.
Selection of parents
From a large PMxE hexaploid population, seven plants were selected on the basis of leafiness, num- ber of tillers, high biomass, and seed production. This hexaploid population originated from a series of crosses involving the cytoplasmic male-sterile in- bred 'Tiff 23DA' (dwarf) pearl millet, 'Mott ' dwarf elephantgrass, and the tall PMxE hexaploid hybrids MN3 and MN33 (2n = 6x = 42) obtained from Dr. W.W. Hanna, USDA-ARS, Tifton, Georgia. MN3 and MN33 were derived from the crosses 23DA x N13 and 23DB x N9, respectively. Both hybrids were treated with colchicine to double the chromo- some number. The population was generated as fol- lows: In 1986, Tilt 23DA was crossed to Mott and triploid progeny were obtained. Selected plants were then grown in tissue culture (Rajasekaran et al., 1986), and through this process, two hexaploid plants were obtained. One of these plants (P3) was then crossed to MN3. The following year, hybrids of P3 × MN3 were crossed to MN33 which contains re- storer genes and hence, male fertility was improved. Mass selection for biomass, persistence, and fertil-

ity during 2 years resulted in the population from which the seven plants were selected.
The seven selected plants were selfed as previ- ously described (Diz & Schank, 1990). Seed derived from each selected plant was classified as a S 1 family. The seed of each S 1 family was divided into two seed sizes, large and small, using a series of sieves. The large seed included half of the total seed which was retained on the upper sieves (larger holes), while the small seed comprised the half retained on the lower sieves. A factorial experiment with a split- plot design was planted at the DRU on May 3,1990. Main plots consisted of the two seed sizes, while the subplots consisted of the seven S 1 families (selfed seed from the previously selected parents, 45B, 109A, 127A, 128B, 131B, 140B, and 144B). Seeds were planted in plastic cavity trays containing Met- romix 200 TM, approximately one month before transplanting the seedlings into the field. Each plot consisted of six plants per row, spaced 0.9 m within the row and 2.7 m between rows (plots). Plots were irrigated four times during May and June, to help establish the plants due to lower than normal rain- fall. Plots received 190 kg N ha -~ in a 4-1-2 ratio with P205 and K20. This rate was split in two equal appli- cations on May 24 and July 7, 1990.
145
wering, number of panicles per plant, panicle length, seed set, seed weight per panicle, number of seeds per panicle, total seed yield per plant, 100- seed weight, and seed germination.
Days to flowering was calculated as the total number of days from transplanting into the field un- til three panicles were fully exerted from their flag leaves. Number of panicles per plant was calculated by multiplying the number of tillers by the average number of panicles per tiller (panicles that had un- dergone anthesis). Panicle length, seed set, and seed weight were measured on one primary panicle per plant. Seed set was estimated by taking a sample of 50 spikelets from the middle of the panicle and threshing them by hand. The number of seeds ob- tained was then divided by 50 (spikelets) and mul- tiplied by 100(%). Seed weight per panicle was ob- tained by partially threshing the panicle with a Forsberg TM portable scarifier, followed by hand- threshing on a rubber board to finish removing the seed hulls. Seeds per panicle was calculated by di- viding the seed weight per panicle by the 100-seed weight, and multiplying by 100. The total seed yield for primary panicles was calculated by multiplying the seed weight per panicle by the number of pri- mary panicles per plant.
Morphological measurements
Morphological characterization included plant height, number of tillers, leaf length, and leaf width. These were measured on every plant on July 27-29 [86 days after planting (DAP)], and on October 24- 28 (approximately 176 DAP). Plant height was mea- sured without extending the top leaves on the first date, and to the top of the inflorescences on the sec- ond date. Four mature leaf blades per plant were sampled from nodes at mid-height of the plant, for both leaf length and width measurements. A mean of these four measurements was calculated for sta- tistical analysis.
Seed-related measurements
Statistical analysis
For both morphological and seed-related charac- teristics, analyses of variance were performed using the general linear model procedure of the Statisti- cal Analysis System (SAS Institute Inc., 1985). This procedure identified differences among families and seed sizes for the characteristics under study, and significance for the family x seed-size interac- tion. Duncan's Multiple Range test was used when the F-test for S 1 families showed significance at the 5 % level of probability. The morphological charac- teristics measured at two growth stages were ana- lyzed separately. Correlation coefficients were cal- culated between certain characteristics to deter- mine their relationship.
Seed-related characteristics included days to rio-
146
Results and discussion
Morphological characteristics
Table 1 s u m m a r i z e s the July and O c t o b e r f ie ld m e a -
s u r e m e n t s o b t a i n e d for the m o r p h o l o g i c a l charac -
ter is t ics s tud ied . B o t h fami ly a n d seed-s ize fac tors
a re i n c o r p o r a t e d in to this t ab le for each charac te r -
istic s tudied . Fami ly x seed-s ize in te rac t ions were
no t s ignif icant for any of the m o r p h o l o g i c a l charac -
ter is t ics s tud ied .
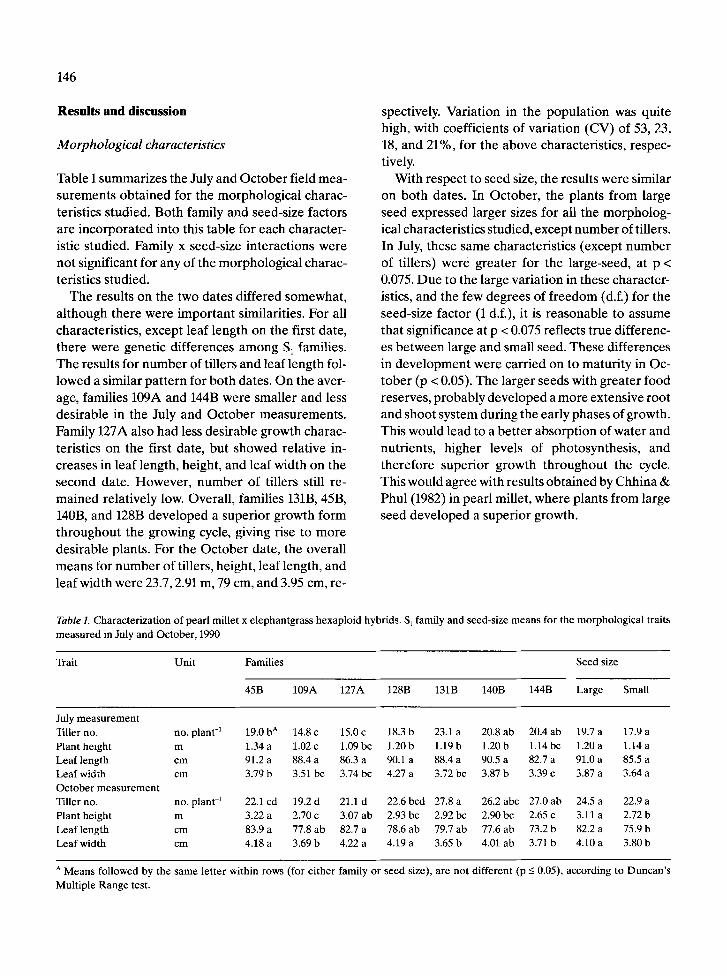
T h e resul ts on the two da tes d i f fe red s o m e w h a t ,
a l t hough the re were i m p o r t a n t s imilar i t ies . F o r all
charac ter i s t ics , excep t leaf l ength on the first date ,
t h e r e were gene t i c d i f f e rences a m o n g S I famil ies .
T h e resul ts for n u m b e r of t i l lers and lea f l eng th fol-
l owed a s imi lar p a t t e r n for bo th dates . O n the aver-
age, fami l ies 109A and 144B were sma l l e r and less
des i r ab l e in the July and O c t o b e r m e a s u r e m e n t s .
Fami ly 127A also had less des i r ab l e g rowth charac -
ter is t ics on the first date , bu t s h o w e d re la t ive in-
c reases in leaf length, height , and leaf wid th on the
s econd date . H o w e v e r , n u m b e r of t i l lers still re-
m a i n e d re l a t ive ly low. Overa l l , fami l ies 131B, 45B,
140B, and 128B d e v e l o p e d a s u p e r i o r g rowth fo rm
t h r o u g h o u t the g rowing cycle, giving rise to m o r e
des i r ab l e p lants . Fo r the O c t o b e r date , the overa l l
m e a n s for n u m b e r o f t i l lers , height , leaf length , and
leaf wid th were 23.7, 2.91 m, 79 cm, and 3.95 cm, re-
spectively. Var ia t ion in the p o p u l a t i o n was qui te
high, wi th coeff ic ients of va r i a t ion (CV) of 53, 23,
18, and 21%, for the a b o v e charac ter i s t ics , r e spec-
tively.
Wi th r e spec t to seed size, the resul ts were s imi lar
on bo th dates . In O c t o b e r , the p lan t s f rom large
seed exp re s sed la rger sizes for all the m o r p h o l o g -
ical charac te r i s t i cs s tud ied , excep t n u m b e r of t i l lers.
In July, these s ame charac te r i s t i cs ( excep t n u m b e r
of t i l lers) were g r e a t e r for the l a rge-seed , at p <
0.075. D u e to the large va r i a t ion in these charac te r -
istics, and the few de g re e s of f r e e d o m (d.f.) for the
seed-s ize fac tor (1 d.f.), it is r e a s o n a b l e to assume
tha t s ignif icance at p < 0.075 ref lects t rue d i f ferenc-
es b e t w e e n la rge and smal l seed . T h e s e d i f fe rences
in d e v e l o p m e n t were ca r r i ed on to m a t u r i t y in Oc-
t o b e r (p < 0.05). The l a rge r seeds wi th g rea t e r food
reserves , p r o b a b l y d e v e l o p e d a m o r e ex tens ive roo t
and shoo t sys tem dur ing the ea r ly phases of growth.
This wou ld l ead to a b e t t e r a b s o r p t i o n of w a t e r and
nut r ien ts , h igher levels of pho tosyn thes i s , and
t h e r e f o r e s u p e r i o r g rowth t h r o u g h o u t the cycle.
Th is would ag ree wi th resul ts o b t a i n e d by Chh ina &
Phul (1982) in pea r l mi l le t , w h e r e p l an t s f rom large
seed d e v e l o p e d a s u p e r i o r growth.
Table 1. Characterization of pearl millet x elephantgrass hexaploid hybrids. S t family and seed-size means for the morphological traits measured in July and October, 1990
Trait Unit Families Seed size
45B 109A 127A 128B 131B 140B 144B Large Small
July measurement Tiller no. no. plant -1 19.0 b A 14.8 c 15.0 c 18.3 b 23.1 a 20.8 ab 20.4 ab 19.7 a 17.9 a Plant height m 1.34 a 1.02 c 1.09 bc 1.20 b 1.19 b 1.20 b 1.14 bc 1.20 a 1.14 a Leaf length cm 91.2 a 88.4 a 86.3 a 90.1 a 88.4 a 90.5 a 82.7 a 91.0 a 85.5 a Leaf width cm 3.79 b 3.51 bc 3.74 bc 4.27 a 3.72 bc 3.87 b 3.39 e 3.87 a 3.64 a October measurement Tiller no. no. plant -~ 22.1 cd 19.2 d 21.1 d 22.6 bcd 27.8 a 26.2 abc 27.0 ab 24.5 a 22.9 a Plant height m 3.22 a 2.70 c 3.07 ab 2.93 bc 2.92 bc 2.90 bc 2.65 c 3.11 a 2.72 b Leaf length cm 83.9 a 77.8 ab 82.7 a 78.6 ab 79.7 ab 77.6 ab 73.2 b 82.2 a 75.9 b Leaf width cm 4.18 a 3.69 b 4.22 a 4.19 a 3.65 b 4.01 ab 3.71 b 4.10 a 3.80 b
A Means followed by the same letter within rows (for either family or seed size), are not different (p -< 0.05), according to Duncan's Multiple Range test.
Seed-related characteristics
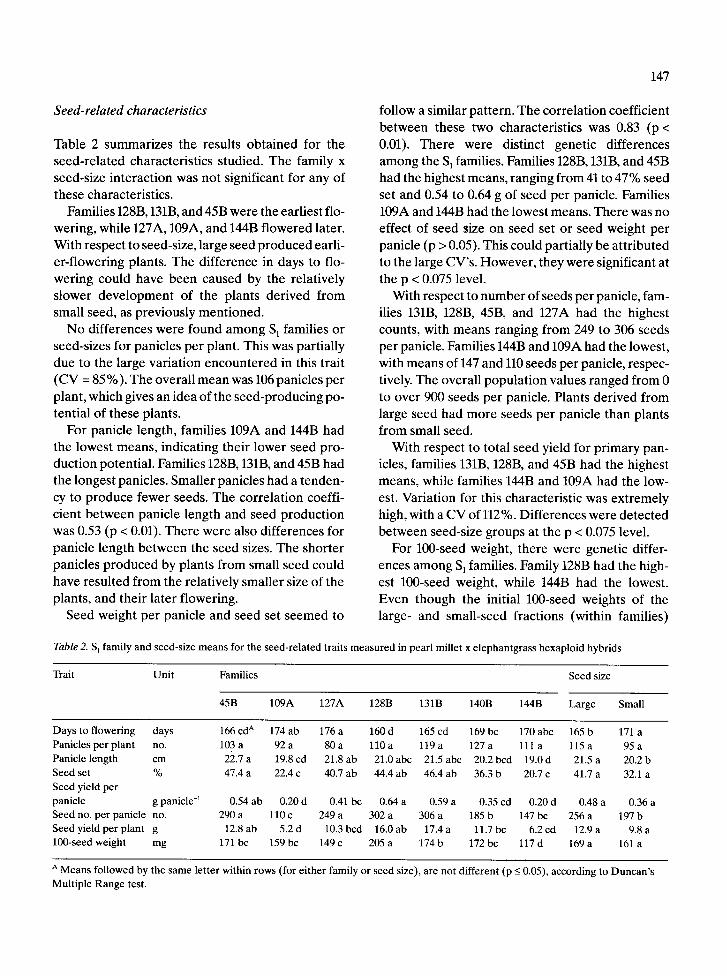
Table 2 summarizes the results obtained for the seed-related characteristics studied. The family x seed-size interaction was not significant for any of these characteristics.
Families 128B, 131B, and 45B were the earliest flo- wering, while 127A, 109A, and 144B flowered later. With respect to seed-size, large seed produced earli- er-flowering plants. The difference in days to flo- wering could have been caused by the relatively slower development of the plants derived from small seed, as previously mentioned.
No differences were found among S~ families or seed-sizes for panicles per plant. This was partially due to the large variation encountered in this trait (CV = 85 %). The overall mean was 106 panicles per plant, which gives an idea of the seed-producing po- tential of these plants.
For panicle length, families 109A and 144B had the lowest means, indicating their lower seed pro- duction potential. Families 128B, 131B, and 45B had the longest panicles. Smaller panicles had a tenden- cy to produce fewer seeds. The correlation coeffi- cient between panicle length and seed production was 0.53 (p < 0.01). There were also differences for panicle length between the seed sizes. The shorter panicles produced by plants from small seed could have resulted from the relatively smaller size of the plants, and their later flowering.
Seed weight per panicle and seed set seemed to
Table 2. S 1 family and seed-size means for the seed-related traits
147
follow a similar pattern. The correlation coefficient between these two characteristics was 0.83 (p < 0.01). There were distinct genetic differences among the S 1 families. Families 128B, 131B, and 45B had the highest means, ranging from 41 to 47% seed set and 0.54 to 0.64 g of seed per panicle. Families 109A and 144B had the lowest means. There was no effect of seed Size on seed set or seed weight per panicle (p > 0.05). This could partially be attributed to the large CV's. However, they were significant at the p < 0.075 level.
With respect to number of seeds per panicle, fam- ilies 131B, 128B, 45B, and 127A had the highest counts, with means ranging from 249 to 306 seeds per panicle. Families 144B and 109A had the lowest, with means of 147 and 110 seeds per panicle, respec- tively. The overall population values ranged from 0 to over 900 seeds per panicle. Plants derived from large seed had more seeds per panicle than plants from small seed.
With respect to total seed yield for primary pan- icles, families 131B, 128B, and 45B had the highest means, while families 144B and 109A had the low- est. Variation for this characteristic was extremely high, with a CV of 112 %. Differences were detected between seed-size groups at the p < 0.075 level.
For 100-seed weight, there were genetic differ- ences among S 1 families. Family 128B had the high- est 100-seed weight, while 144B had the lowest. Even though the initial 100-seed weights of the large- and small-seed fractions (within families)
measured in pearl millet x elephantgrass hexaploid hybrids
Trait Unit Families Seed size
45B 109A 127A 128B 131B 140B 144B Large Small
Days to flowering days 166 cd A 174 ab 176 a 160 d 165 cd 169 bc 170 abc 165 b 171 a
Panicles per plant no. 103 a 92 a 80 a 110 a 119 a 127 a 111 a 115 a 95 a
Panicle length cm 22.7 a 19.8 cd 21.8 ab 21.0 abc 21.5 abc 20.2 bcd 19.0d 21.5 a 20.2b
Seed set % 47.4 a 22.4 c 40.7 ab 44.4 ab 46.4 ab 36.3 b 20.7 c 41.7 a 32.1 a
Seed yield per
panicle g panicle -1 0.54 ab 0.20 d 0.41 bc 0.64 a 0.59 a 0.35 cd 0.20 d 0.48 a 0.36 a
Seed no. per panicle no. 290 a 110 c 249 a 302 a 306 a 185 b 147 bc 256 a 197 b
Seed yield per plant g 12.8 ab 5.2 d 10.3 bcd 16.0 ab 17.4 a 11.7 bc 6.2 cd 12.9 a 9.8 a
100-seed weight mg 171 bc 159 bc 149 c 205 a 174 b 172 bc 117 d 169 a 161 a
A Means followed by the same let ter within rows (for ei ther family or seed size), are not different (p <_ 0.05), according to Duncan 's
Multiple Range test.

148
were different, there was no difference in this char- acteristic for the plants derived from these large or small seeds. This would indicate that those initial differences in seed weight within families were due to the environment (probably differences in endo- sperm filling), and not to genetic differences. This environmental - not genetic - effect on seed size was first demonstrated by W. Johannsen in his clas- sical 1903 paper titled 'Heredity in populations and pure-lines' (Peters, 1959).
Results for the seed-size factor are summarized in Table 3, for both the morphological and seed-re- lated characteristics. Larger seeds gave rise to bet- ter plant growth than smaller seeds. This is in con- trast to results obtained by Maranville & Clegg (1977) and Singh & Chopde (1987) in sorghum, but agrees with results obtained by Chhina & Phul (1982) in pearl millet. Larger seeds have greater food reserves for embryo growth, which is advanta- geous during seedling establishment. The more vig- orous seedlings may initially produce a more exten- sive root and shoot system and therefore develop a superior growth form throughout their growth cy- cle. Selecting for seed size within families will not be effective, since the existing variation is primarily en-
vironmental. However, the differences in seed size among families is evidence of genetic variation. Family selection should be successful for increasing seed size.
In conclusion, genetic differences were evident among S 1 families in the seed-related characteristics studied. The different seed-yield components (till- ers per plant, panicles per tiller, number of seeds per panicle, and 100-seed weight) will be important in identifying families which are genetically superi- or for seed production. Families 128B, 131B, and 45B were the most promising for overall fertility characteristics. These were also the earliest flower- ing families, which would be advantageous in areas where early frosts are a concern for seed produc- tion. Families 109A and 144B were less desirable due to their low seed-yield components, late flower- ing, and less desirable morphological characteris- tics. Expression of deleterious genes were observed in several plants, which could be due to inbreeding depression suffered by these selfed progeny. This somewhat lowered the averages obtained for the traits studied. Further breeding with clear objec- tives will be needed in the future to develop pearl millet x elephantgrass hexaploid hybrid varieties.
Table 3. Resul ts for the seed-size factor on the morphological and seed-related characteristics studied in pearl millet x elephantgrass hexaploid hybrids. Co lumns show the mean values for the large and small seed, their percent difference, significance, and the coefficient
of variation
Characteristic Seed size Differ (%) Signif. C.V.
Morphological: L S (L-S)/S No. tillers 24.5 22.9 6.7 N.S. A 53.4 Height (m) 3.1 2.7 14.5 **B 22.9 Leaf length (cm) 82.2 75.9 8.3 ,c 18.4
Leaf width (cm) 4.1 3.8 8.1 * 20.6
Seed-related: D. to flowering 165 171 - 3.2 * 8.0 Panicles per plant 115 95 20.6 N.S. 84.7 Panicle length (cm) 21.5 20.2 6.4 * 20.4 Seed set (%) 41.7 32.1 29.9 N.S. 64.6 Seed yield per panicle (g) 0.48 0.36 35.2 N.S. 86.4 Seed no. per panicle 256 197 29.7 * 67.9 Seed yield per plant (g) 12.9 9.8 31.6 N.S. 112.0 100-seed weight (mg) 169 161 4.7 N.S. 30.1
A N.S.: Difference not significant (p _< 0.05). B **: Significant at p _< 0.01. c ,: Significant at p _< 0.05.

Acknowledgment
The research work reported in this paper was sup- ported by a cooperative agreement between the In- stitute of Food and Agricultural Sciences, Universi- ty of Florida, and the Gas Research Institute, Chica- go, Illinois. Florida Agricultural Experiment Sta- tion Journal Series Number R-02813.
References
Chhina, B.S. & RS. Phul, 1982. Association of seed size and seed- ling vigour with various morphological traits in pearl millet. Seed. Sci. Technol. 10: 541-545.
Diz, D.A. & S.C. Schank, 1990. Seed and seedling characteriza- tion of pearl millet x napiergrass hexaploid hybrids. Proc. Soil Crop Sci. Soc. Florida 50: 69-75.
Gonzalez, B. & W.W. Hanna, 1984. Morphological and fertility responses in isogenic triploid and hexaploid pearl millet x na- piergrass hybrids. J. Hered. 75: 317-318.
Hanna, W.W., 1981. Method of reproduction in napiergrass and in the 3X and 6X alloploid hybrids with pearl millet. Crop. Sci. 21: 123-126.
Jauhar, EE, 1981. Cytogenetics and breeding of pearl millet and related species. Alan R. Liss, Inc. New York.
Khan, M.D. & H. Rahman, 1963. Genome relationship and chro- mosome behaviour in the allotriploid hybrid of Pennisetum ty-
phoides and P. purpureum. W. Pak. J. Agric. Res. 1: 61-65. Lawan, M., EL. Barnett, B. Khaleeq & R.L. Vanderlip, 1985.
Seed density and seed size of pearl millet as related to field emergence and several seed and seedling traits. Agron. J. 77: 567-571.
Maranville, J.W. & M.D. Clegg, 1977. Influence of seed size and density on germination, seedling emergence, and yield of grain sorghum. Agron. J. 69: 329-330.
149
Mislevy, R, R.S. Kalmbacher, A.J. Overman & EG. Martin, 1986. Effect of fertilizer and nematicide treatments on crops grown for biomass. Biomass 11: 243-253.
Muldoon, D.K. & C.J. Pearson, 1979. The hybrid between Penni-
setum americanum and Pennisetum purpureum. Herbage Abstr. 49: 189-199.
Peters, J.A., 1959. Classic papers in genetics. Prentice-Hall Inc., Englewood Cliffs, N.J.
Powell, J.B. & G.W. Burton, 1966. A suggested commercial method of producing an interspecific hybrid forage in Pennise-
rum. Crop. Sci. 6: 378-379. Rajasekaran, K., S.C. Schank & I.K. Vasil, 1986. Characteriza-
tion of biomass production, cytology, and phenotypes of plants regenerated from embryogenic callus cultures of Pennisetum
americanum x P. purpureum (hybrid triploid napiergrass). Theor. Appl. Genet. 73: 4-10.
SAS Institute Inc., 1985. SAS user's guide: statistics. SAS Insti- tute, Inc., Cary, N.C.
Schank, S.C. & D.A. Diz, 1991. A seeded type of hybrid hexa- ploid elephantgrass with a potential for livestock production in the tropics and subtropics. In: Proc. Intern. Conf. Livestock in the Tropics. pp. A7-A13. Inst. Food Agric. Sci., Univ. of Florida, Gainesville.
Schank, S.C., D.E Chynoweth, C.E. Turick & P.E. Mendoza, 1993. Napiergrass genotypes and plant parts for biomass ener- gy. Biomass and Bioenergy 4: 1-7.
Singh, A.R, 1987. Effects of seed size on seed viability and seed- ling vigour in sorghum. J Maharashtra Agric. Univ. 12:141-142.
Singh, A.R. & P.R. Chopde, 1987. Effect of seed size on perform- ance of the resulting crop of Sorghum bicolor L. Moench. Seed. Res. 14: 244-247.
Sollenberger, L.E. & C.S. Jones, Jr., 1989. Beef production from nitrogen-fertilized Mott dwarf elephantgrass and Pensacola bahiagrass pastures. Trop. Grass. 23: 129-134.
Sollenberger, L.E., C.S. Jones, Jr., K.A. Albrecht & G.H. Rui- tenberg, 1990. Vegetative establishment of dwarf elephant- grass: effect of defoliation prior to planting stems. Agron. J. 82: 274-278.