characterization of immunity induced by m2e of influenza virus
TRANSCRIPT

A
Hbicc©
K
1
Hsaeatglvtcmtwccat
0d
Available online at www.sciencedirect.com
Vaccine 25 (2007) 8868–8873
Characterization of immunity induced by M2e of influenza virusFan Wu, Jing-He Huang, Xiao-Yi Yuan, Wei-Shan Huang, Ying-Hua Chen ∗Laboratory of Immunology, Department of Biology, Tsinghua University, Protein Science Laboratory of Ministry of Education,
Beijing 100084, PR China
Received 19 July 2007; received in revised form 25 September 2007; accepted 25 September 2007Available online 22 October 2007
bstract
The extracellular-domain of influenza Matrix 2 protein (M2e) is considered as a putative target for designing universal influenza vaccines.owever, the mechanism by which M2-based vaccine induces protection has not been clear. In this study, we analyzed the immunity inducedy free synthetic M2e peptide and found the peptide was highly immunogenic. Without carrier proteins, the synthetic M2e peptide could
nduce M2e-specific IgG antibodies in both incomplete Freund’s and aluminum adjuvant. The peptide could also provoke M2e-specific Tell response, which could not be mounted by influenza virus. Moreover, immunization with M2e peptide could protect mice from a lethalhallenge with influenza virus. These results provide useful information for the development of M2e-based influenza vaccine.2007 Elsevier Ltd. All rights reserved.
eywords: Influenza A virus; M2e; Immunity; Protection; Vaccine
pdtprccMvappdptMWt
. Introduction
The threat of highly virulent avian influenza, such as5N1 virus, brings out an urgent need to develop a univer-
al influenza vaccine, which could provide cross-protectiongainst different influenza virus strains [1]. The conservedxtra-domain of influenza M2 protein (M2e) is considered aspromising candidate target. Previous studies have revealed
hat M2e-specific monoclonal antibody 14C2 restricted viralrowth in vitro [2] and protected mice against virus chal-enge in passive transfer experiments [3]. Several M2-basedaccines have been proved to provide successful protec-ion against homologous and heterologous influenza virushallenge, including H5N1 subtype [4–7]. However, theechanism by which M2-based vaccine-induced protec-
ion was not clear. In some reports, M2e-specific antibodiesere suggested to play an important role. The M2-hepatitis
ore (M2-HBc) particles protected mice against influenza
hallenge by inducing M2e-specific antibodies, and passivedministration of antibodies could provide sufficient protec-ion [5]. A study by Jegerlehner et al. indicated that influenza∗ Corresponding author. Tel.: +86 10 6277 2267; fax: +86 10 6277 1613.E-mail address: [email protected] (Y.-H. Chen).
tabTar
264-410X/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2007.09.056
rotection induced by M2e-HBc was mediated via antibody-ependent cell cytotoxicity [8]. Fan et al. have also reportedhat passive administration of monkey anti-M2e sera couldrotect mice against influenza challenge [9]. In our previouseports, rabbit antibodies induced by M2e–BSA conjugatesould also inhibit influenza virus replication in vitro [10]. Inontrast, vaccination of a DNA plasmid containing influenza
genes protected mice against influenza infection by acti-ation of CD4+ and CD8+ T cells instead of protectiventibodies [11]. Vaccination with baculovirus-expressed M2rotein could also confer protection despite of absence ofrotective antibodies [4]. Protective immunity provoked byifferent M2e-based vaccines could be influenced by carrierroteins, adjuvant or routes of administration [12]. To inves-igate the influence of carrier proteins, we tested whether
2e peptide could induce protective immunity without them.e found that the M2e peptide was highly immunogenic. In
he absence of carrier protein, immunization of M2e pep-ide with both incomplete Freund’s adjuvant and aluminumdjuvant could induce high titres of M2e-specific IgG anti-
odies in mice. The peptide could also provoke M2e-specificcell response. Moreover, vaccination of the peptide couldlso protect mice against lethal influenza virus challenge. Ouresults suggested that carrier protein was not necessary for

ne 25 (
Mpmti
2
2
Hehbpa3f9bdNa[cca
2
wuo1nSia
2
lmpasitd(ra
mwMttapO
2
Pioncaa
2
capaiTBtbbRwta4atfwb
2
alow
F. Wu et al. / Vacci
2e-induced protection and there was T cell epitopes on M2eeptide. Our results also suggested that M2e-specific T cellight be involved in influenza protection induced by the pep-
ide, which would provide useful information for M2e-basednfluenza vaccine development.
. Materials and methods
.1. Virus and peptide
Mouse-adapted influenza virus, A/Purto Rico/8/34 (PR8;1N1) was propagated in allantoic fluids of 10-day-old
mbryonated eggs at 37 ◦C for 3 days. Allantoic fluids werearvested and stored at −80 ◦C. Virus titres were measuredy plaque assay on MDCK cell monolayer as describedreviously [13]. To inactivate allantoic fluids, formalin wasdded at a final concentration of 0.1% before incubation at7 ◦C for 24 h. The inactivated influenza virus was enrichedrom allantoic fluids by centrifugation at 75 600 × g for0 min. Influenza virus pellet was dissolved in phosphate-uffered saline (PBS) and virus protein concentration wasetermined by Cormass Blue dying assay. An M2e peptide,-KSLLTEVETPIRNEWGCRCNDSSD, which containeduniversal sequence of extracellular part of M2 protein
5], was synthesized at Genemed Synthesis Inc. (San Fran-isco, USA). Stock solution of peptide was prepared atoncentration of 10 mg/mL in distilled water and storedt −20 ◦C.
.2. Immunization
Female Balb/C mice were intraperitoneally immunizedith M2e peptide (10 �g per mouse) in incomplete Fre-nd’s adjuvant, aluminum adjuvant or PBS at final volumef 200 �L. Another group of mice were immunized with0 �g inactivated PR8 virus as controls. A booster immu-ization was given with the same immunogen 2 weeks later.era were collected 7 days after final immunization. Pre-
mmunized sera (MNS) were prepared before immunizations negative controls.
.3. Antibody detection
The M2e-specific antibodies were detected by enzyme-inked immunosorbent assay (ELISA). Ninety-six-well
icrotitre plates (Costar) were coated with 50 �L M2eeptide (5 �g/mL) in PBS for 2 h at room temperaturend blocked with PBS containing 0.25% gelatin. Serumamples were serially diluted and added, followed by 1 hncubation at room temperature. After extensive washes,he bound antibodies were detected by addition of 1:2000
iluted horseradish peroxidase-linked anti-mouse antibodiesDAKO) and substrate o-phenylenediamine dihydrochlo-ide peroxide solution (OPD, Sigma) sequentially. Thebsorbance at 450 nm (A450) was recorded.wwPt
2007) 8868–8873 8869
To determine isotypes of M2e-specific antibodies,icrotitre plates were coated with M2e peptide and blockedith blocking buffer. Fifty microliters of serial diluted2e-positive sera were added. After incubation at room
emperature and extensive washes, the bound antibody iso-ypes were detected by sequentially addition of 50 �L goatnti-mouse heavy chain antibodies (sigma), horseradisheroxidase-linked rabbit anti-goat antibodies and substratePD. The A450 was measured.
.4. Flow cytometry
Monolayer of MDCK Cells was infected overnight withR8 influenza virus at 0.5 multiplicity of infection. 1 × 106
nfected cells were incubated with 1:100 diluted sera for 1 hn ice. The uninfected MDCK cells were stained and used asegative controls. After washes with PBS, 1:40 diluted FITC-onjugated rabbit anti-mouse immunoglobulin were addednd incubated for 1 h on ice. The cells were washed twicend analyzed in a FACS-Calibur (Becton-Dickinson, USA).
.5. Enzyme-linked immunospot (ELISPOT) assay
The number of M2e-specific IL-4 and IFN-�-secretingells in mouse was counted using commercial ELISPOTssay kits (U-Cytech, Netherlands). The procedure waserformed as described in the instruction manual. Briefly,nti-mouse IL-4 or anti-mouse IFN-� monoclonal antibod-es were coated on 96-well ELISPOT plates at 4 ◦C for 8 h.he plates were blocked with 200 �L PBS containing 1%SA at 37 ◦C for 4 h. Mice were sacrificed at day 7 post
he second immunization. Spleen lymphocytes were isolatedy density separation on mouse lymphocyte cell separationuffer (LTS1092, TBD, China) and resuspended in completePMI-1640 medium (Hyclone) with 10% FCS. 1 × 105 cellsere transferred to ELISPOT plates and 1 �g/mL M2e pep-
ide was added as stimulator. Medium without peptide wasdded as a negative control. After incubation at 37 ◦C for0 h, 5% CO2, biotinylated detected antibodies and �-labelednti-biotin antibodies (GABA) solution were added, sequen-ially. After incubation with freshly prepared activator I/IIor 15 min, the visible spots of IL-4 or IFN-�-secreting cellsere photographed with microscope system and enumeratedy TotalLab software.
.6. Virus challenge
Mice were immunized with M2e peptide as describedbove. Seven days post final immunization, mice were chal-enged intranasally with one MLD90 (dose lethal to 90%f mice) of live influenza virus PR8. Un-immunized miceere challenged as negative controls. The loss of body
eight and death were monitored every day. Survival curvesere calculated by Kaplan–Meier methods with Graphpadrism software and survival data were compared by Logrankest.
8 ne 25 (
3
3o
b2c
FataPiibmidm
pba[ia
870 F. Wu et al. / Vacci
. Results
.1. Free synthetic M2e peptide could induce high titresf M2e-specific IgG antibodies
To test whether M2e peptide could induce specific anti-
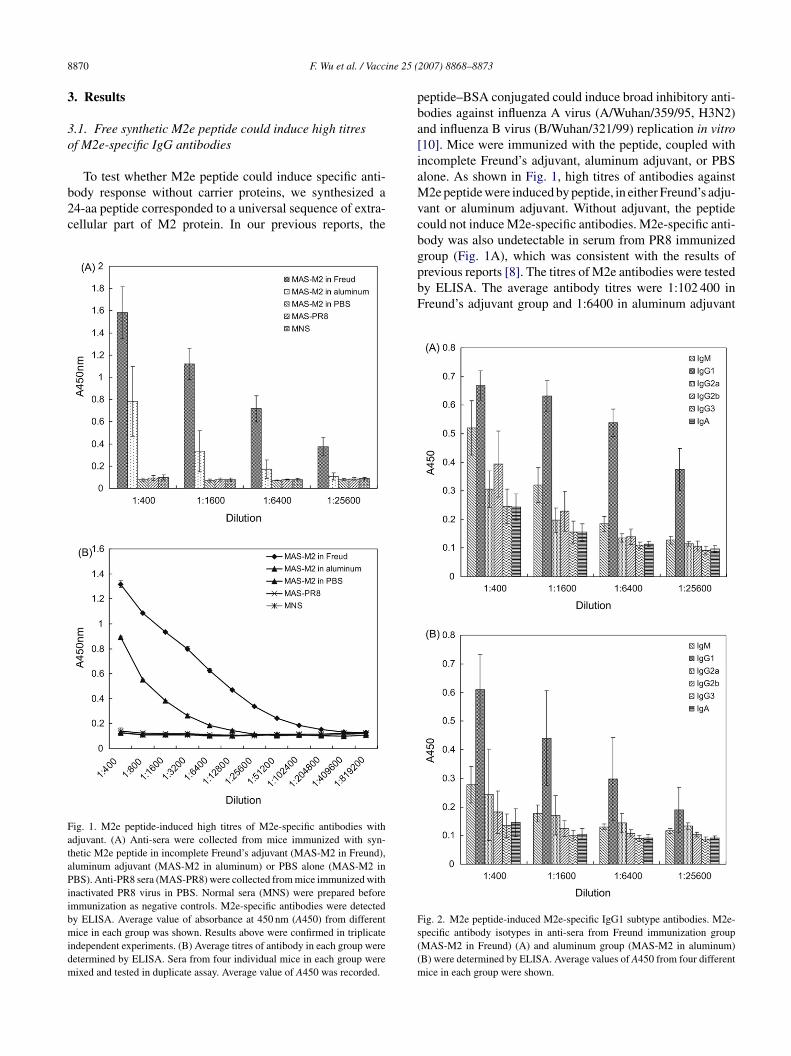
ody response without carrier proteins, we synthesized a4-aa peptide corresponded to a universal sequence of extra-ellular part of M2 protein. In our previous reports, theig. 1. M2e peptide-induced high titres of M2e-specific antibodies withdjuvant. (A) Anti-sera were collected from mice immunized with syn-hetic M2e peptide in incomplete Freund’s adjuvant (MAS-M2 in Freund),luminum adjuvant (MAS-M2 in aluminum) or PBS alone (MAS-M2 inBS). Anti-PR8 sera (MAS-PR8) were collected from mice immunized with
nactivated PR8 virus in PBS. Normal sera (MNS) were prepared beforemmunization as negative controls. M2e-specific antibodies were detectedy ELISA. Average value of absorbance at 450 nm (A450) from differentice in each group was shown. Results above were confirmed in triplicate
ndependent experiments. (B) Average titres of antibody in each group wereetermined by ELISA. Sera from four individual mice in each group wereixed and tested in duplicate assay. Average value of A450 was recorded.
MvcbgpbF
Fs((m
2007) 8868–8873
eptide–BSA conjugated could induce broad inhibitory anti-odies against influenza A virus (A/Wuhan/359/95, H3N2)nd influenza B virus (B/Wuhan/321/99) replication in vitro10]. Mice were immunized with the peptide, coupled withncomplete Freund’s adjuvant, aluminum adjuvant, or PBSlone. As shown in Fig. 1, high titres of antibodies against2e peptide were induced by peptide, in either Freund’s adju-
ant or aluminum adjuvant. Without adjuvant, the peptideould not induce M2e-specific antibodies. M2e-specific anti-ody was also undetectable in serum from PR8 immunizedroup (Fig. 1A), which was consistent with the results of
revious reports [8]. The titres of M2e antibodies were testedy ELISA. The average antibody titres were 1:102 400 inreund’s adjuvant group and 1:6400 in aluminum adjuvantig. 2. M2e peptide-induced M2e-specific IgG1 subtype antibodies. M2e-pecific antibody isotypes in anti-sera from Freund immunization groupMAS-M2 in Freund) (A) and aluminum group (MAS-M2 in aluminum)B) were determined by ELISA. Average values of A450 from four differentice in each group were shown.
ne 25 (
giw
mMba
tssFMtsP
acr
3l
icwsf
FPMw
F. Wu et al. / Vacci
roup, respectively. And the M2e-specific antibodies titresn sera from PR8 immunized group and adjuvant-free groupere lesser than 1:400 (Fig. 1B).M2e-specific antibody isotypes in anti-sera were deter-
ined by ELISA with isotype-specific antibodies. Synthetic2e peptide induced specific IgG1 subtype-dominant anti-
odies both in Freund’s adjuvant (Fig. 2A) and aluminumdjuvant groups (Fig. 2B).
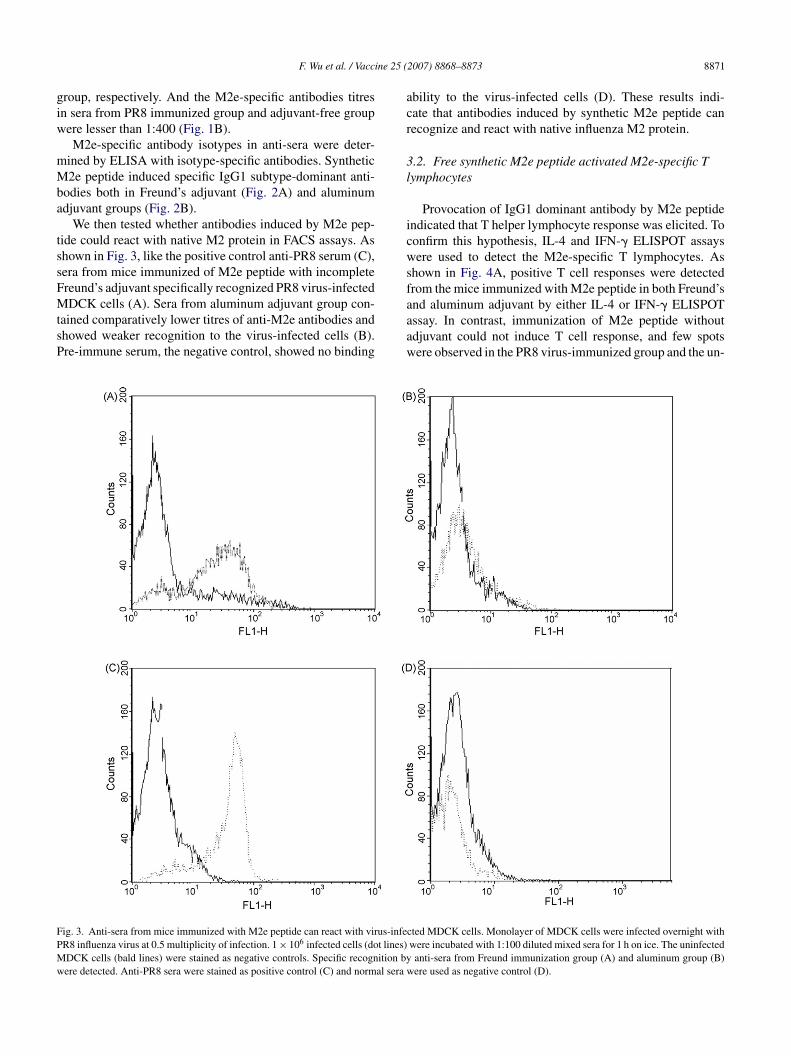
We then tested whether antibodies induced by M2e pep-ide could react with native M2 protein in FACS assays. Ashown in Fig. 3, like the positive control anti-PR8 serum (C),era from mice immunized of M2e peptide with incompletereund’s adjuvant specifically recognized PR8 virus-infected
DCK cells (A). Sera from aluminum adjuvant group con-ained comparatively lower titres of anti-M2e antibodies andhowed weaker recognition to the virus-infected cells (B).re-immune serum, the negative control, showed no binding
aaaw
ig. 3. Anti-sera from mice immunized with M2e peptide can react with virus-infeR8 influenza virus at 0.5 multiplicity of infection. 1 × 106 infected cells (dot lines)DCK cells (bald lines) were stained as negative controls. Specific recognition byere detected. Anti-PR8 sera were stained as positive control (C) and normal sera w
2007) 8868–8873 8871
bility to the virus-infected cells (D). These results indi-ate that antibodies induced by synthetic M2e peptide canecognize and react with native influenza M2 protein.
.2. Free synthetic M2e peptide activated M2e-specific Tymphocytes
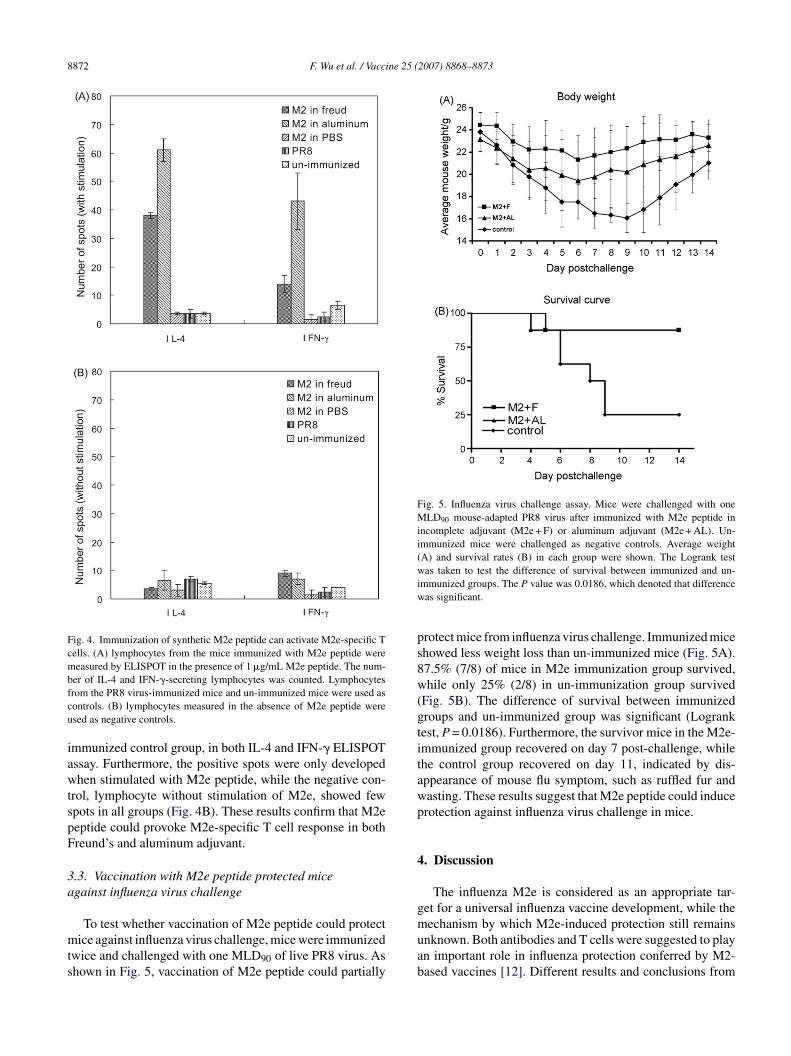
Provocation of IgG1 dominant antibody by M2e peptidendicated that T helper lymphocyte response was elicited. Toonfirm this hypothesis, IL-4 and IFN-� ELISPOT assaysere used to detect the M2e-specific T lymphocytes. As
hown in Fig. 4A, positive T cell responses were detectedrom the mice immunized with M2e peptide in both Freund’s
nd aluminum adjuvant by either IL-4 or IFN-� ELISPOTssay. In contrast, immunization of M2e peptide withoutdjuvant could not induce T cell response, and few spotsere observed in the PR8 virus-immunized group and the un-cted MDCK cells. Monolayer of MDCK cells were infected overnight withwere incubated with 1:100 diluted mixed sera for 1 h on ice. The uninfectedanti-sera from Freund immunization group (A) and aluminum group (B)ere used as negative control (D).
8872 F. Wu et al. / Vaccine 25 (2007) 8868–8873
Fig. 4. Immunization of synthetic M2e peptide can activate M2e-specific Tcells. (A) lymphocytes from the mice immunized with M2e peptide weremeasured by ELISPOT in the presence of 1 �g/mL M2e peptide. The num-bfcu
iawtspF
3a
mts
Fig. 5. Influenza virus challenge assay. Mice were challenged with oneMLD90 mouse-adapted PR8 virus after immunized with M2e peptide inincomplete adjuvant (M2e + F) or aluminum adjuvant (M2e + AL). Un-immunized mice were challenged as negative controls. Average weight(A) and survival rates (B) in each group were shown. The Logrank testwiw
ps8w(gtitawp
4
g
er of IL-4 and IFN-�-secreting lymphocytes was counted. Lymphocytesrom the PR8 virus-immunized mice and un-immunized mice were used asontrols. (B) lymphocytes measured in the absence of M2e peptide weresed as negative controls.
mmunized control group, in both IL-4 and IFN-� ELISPOTssay. Furthermore, the positive spots were only developedhen stimulated with M2e peptide, while the negative con-
rol, lymphocyte without stimulation of M2e, showed fewpots in all groups (Fig. 4B). These results confirm that M2eeptide could provoke M2e-specific T cell response in bothreund’s and aluminum adjuvant.
.3. Vaccination with M2e peptide protected micegainst influenza virus challenge
To test whether vaccination of M2e peptide could protectice against influenza virus challenge, mice were immunized
wice and challenged with one MLD90 of live PR8 virus. Ashown in Fig. 5, vaccination of M2e peptide could partially
muab
as taken to test the difference of survival between immunized and un-mmunized groups. The P value was 0.0186, which denoted that differenceas significant.
rotect mice from influenza virus challenge. Immunized micehowed less weight loss than un-immunized mice (Fig. 5A).7.5% (7/8) of mice in M2e immunization group survived,hile only 25% (2/8) in un-immunization group survived
Fig. 5B). The difference of survival between immunizedroups and un-immunized group was significant (Logrankest, P = 0.0186). Furthermore, the survivor mice in the M2e-mmunized group recovered on day 7 post-challenge, whilehe control group recovered on day 11, indicated by dis-ppearance of mouse flu symptom, such as ruffled fur andasting. These results suggest that M2e peptide could inducerotection against influenza virus challenge in mice.
. Discussion
The influenza M2e is considered as an appropriate tar-et for a universal influenza vaccine development, while the
echanism by which M2e-induced protection still remainsnknown. Both antibodies and T cells were suggested to playn important role in influenza protection conferred by M2-ased vaccines [12]. Different results and conclusions from

ne 25 (
ptstMtv(cpMrTiMtetgs(rcccmvTavdMMehi
pNiviwiipatTbs
A
Sm
R
[
[
[
[
[
F. Wu et al. / Vacci
revious studies may be due to the difference of carrier pro-eins, adjuvant or routes of administration. In the presenttudy, we investigated the immunity induced by a free syn-hetic M2e peptide without carrier protein. We found that
2e peptide was highly immunogenic. The peptide was ableo induce high titres of M2e-specific IgG antibodies whenaccinated with incomplete Freund’s and aluminum adjuvantantibody dilution: 1:102 400 and 1:6400, respectively). Vac-ination of M2e peptide with adjuvant could also significantlyrotect mice against lethal dose of influenza virus challenge.oreover, the peptide provoked M2e-specific T helper cell
esponse, indicated by both IL-4 and IFN-� ELISPOT assay.hese results suggested that T cells might participate in
nfluenza protection conferred by M2e peptide. However,2e-specific T helper cell could not provide direct protec-
ion without antibody. In a passively transferred experiment,nriched T cells from M2e-peptide-immunized mice wereransferred to naı̈ve mice. Compared with un-immunizedroup, the mice which received T cells suffered even moreevere symptom after sub-lethal influenza virus challengedata not shown). The T cell subset may have influence on theesult because transfer of influenza virus-specific Th2 T celllone could exacerbate pulmonary pathology and delay viruslearance in mice [14]. Besides, the M2e-specific T helperell may drive B cell to produce anti-M2e antibody and pro-ote immune memory [15]. Compared with M2e peptide,
accination with influenza virus did not activate M2e-specifichelper cell, in accordance with undetectable M2e-specific
ntibody response. These results were consistent with pre-ious reports, in which M2e-specific T cells were absenturing virus infection [16]. The immuno-recessiveness of2e-specific T cell may account for the absence of anti-2e antibodies during virus vaccination. However, further
xploration is needed to investigate the role of M2e-specific Telper cell during influenza protection and the T cell responsenduced by different M2e conjugated.
To be mentioned, adjuvant also plays an important role inrovocation of M2e-specific humoral and T cell response.either M2e-specific antibodies nor T cell response was
nduced by vaccination with M2e peptide in absence of adju-ant. The adjuvant might adsorb the peptide and activatennate immune system. However, the exact mechanism byhich Freund’s adjuvant and aluminum adjuvant enhance
mmune response still remains unknown. Compared with thatn incomplete Freund’s adjuvant, the peptide in aluminumreferred eliciting T cell response to inducing high titres ofntibodies, which suggested that different antigen presen-
ation pathways were used by different kinds of adjuvant.hese results suggested that the effect of adjuvant shoulde appropriately tested before the M2e peptide was used asupplement for traditional influenza vaccine.[
[
2007) 8868–8873 8873
cknowledgments
This work is funded by the 973-2005CB522503 andRFDP-20050003064. We thank Ms. Yu Chen foranuscript revising.
eferences
[1] Gerhard W, Mozdzanowska K, Zharikova D. Prospects for universalinfluenza virus vaccine. Emerg Infect Dis 2006;12(4):569–74.
[2] Zebedee SL, Lamb RA. Influenza A virus M2 protein: monoclonalantibody restriction of virus growth and detection of M2 in virions. JVirol 1988;62(8):2762–72.
[3] Treanor JJ, Tierney EL, Zebedee SL, Lamb RA, Murphy BR. Passivelytransferred monoclonal antibody to the M2 protein inhibits influenzaA virus replication in mice. J Virol 1990;64(3):1375–7.
[4] Slepushkin VA, Katz JM, Black RA, Gamble WC, Rota PA,Cox NJ. Protection of mice against influenza A virus challengeby vaccination with baculovirus-expressed M2 protein. Vaccine1995;13(15):1399–402.
[5] Neirynck S, Deroo T, Saelens X, Vanlandschoot P, Jou WM, Fiers W.A universal influenza A vaccine based on the extracellular domain ofthe M2 protein. Nat Med 1999;5(10):1157–63.
[6] Tompkins SM, Zhao ZS, Lo CY, Misplon JA, Liu T, Ye Z, et al. Matrixprotein 2 vaccination and protection against influenza viruses, includingsubtype H5N1. Emerg Infect Dis 2007;13(3):426–35.
[7] Ernst WA, Kim HJ, Tumpey TM, Jansen AD, Tai W, Cramer DV,et al. Protection against H1, H5, H6 and H9 influenza A infectionwith liposomal matrix 2 epitope vaccines. Vaccine 2006;24(24):5158–68.
[8] Jegerlehner A, Schmitz N, Storni T, Bachmann MF. Influenza Avaccine based on the extracellular domain of M2: weak protec-tion mediated via antibody-dependent NK cell activity. J Immunol2004;172(9):5598–605.
[9] Fan J, Liang X, Horton MS, Perry HC, Citron MP, HeideckerGJ, et al. Preclinical study of influenza virus A M2 peptide con-jugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine2004;22(23/24):2993–3003.
10] Liu W, Li H, Chen YH. N-terminus of M2 protein could induce antibod-ies with inhibitory activity against influenza virus replication. FEMSImmunol Med Microbiol 2003;35(2):141–6.
11] Okuda K, Ihata A, Watabe S, Okada E, Yamakawa T, Hamajima K, etal. Protective immunity against influenza A virus induced by immu-nization with DNA plasmid containing influenza M gene. Vaccine2001;19(27):3681–91.
12] De Filette M, Min Jou W, Birkett A, Lyons K, Schultz B, Tonkyro A, etal. Universal influenza A vaccine: optimization of M2-based constructs.Virology 2005;337(1):149–61.
13] Li H, Ding J, Chen YH. Recombinant protein comprising multi-neutralizing epitopes induced high titer of antibodies against influenzaA virus. Immunobiology 2003;207(5):305–13.
14] Graham MB, Braciale VL, Braciale TJ. Influenza virus-specific CD4+T helper type 2 T lymphocytes do not promote recovery from experi-mental virus infection. J Exp Med 1994;180(4):1273–82.
15] Brown DM, Roman E, Swain SL. CD4 T cell responses to influenzainfection. Semin Immunol 2004;16(3):171–7.
16] Crowe SR, Miller SC, Brown DM, Adams PS, Dutton RW, HarmsenAG, et al. Uneven distribution of MHC class II epitopes within theinfluenza virus. Vaccine 2006;24(4):457–67.