characteristics of thyroid lysosomal cathepsin
TRANSCRIPT

BIOCHIMICA ET BIOPHYSICA ACTA
BBA 65344
CHARACTERISTICS OF THYROID LYSOSOMAL CATHEPSIN
KANDIAH BALASUBRAMANIAM AND WILLIAM P. DEISS, JR.
Departments of Biochemistry and Medicine, Indiana University School of Medicine, Indianapolis,Ind. (U.S.A.)
(Received April znd, 1965)
(Revised manuscript received August 23rd, 1965)
SUMMARY
Some biochemical characteristics of thyroid lysosomes were studied. About40% of the cathepsin of thyroid homogenates was present in a sedimentable form inthe IS 000 X g pellet. The stability of the lysosomes was affected by pH and osmolarity. The lysosomal cathepsin is latent between pH 5.1 and 7.3 in isotonic sucrose at37° for 30 min.
In thyroid extracts and homogenates, only one cathepsin with an acid pHoptimum was observed. No neutral or alkaline protease could be demonstrated. Thecharacteristics of this acid lysosomal cathepsin resemble those reported for cathepsinD (EC 3+4.23).
The formation of polypeptides of high molecular weights as products of autolysis of thyroid iodoproteins was demonstrated by gel filtration. In addition to undegraded thyroglobulin and iodoamino acids, four intermediate polypeptides wereeluted with characteristics suggesting molecular weights of approx. 70000, 50000,30000, and 18000, respectively.
INTRODUCTION
Proteolysis in the thyroid gland has been of particular interest because of itsrelevance to the release of the thyroid hormones, thyroxine (T4) and 3,S,3'-triiodothyronine (Ta) from their peptide linkage in thyroglobulin to the circulation. In vitro,the proteolysis of thyroglobulin by the thyroid protease occurs optimally betweenpH 3 and 5 (refs. 1-3). Of the several proteolytic enzymes that have been reported tohave their pH optimum in the acid, neutral or alkaline ranges4 - 10, only the proteasewith the acid pH optimum has been well characterized and consistently shown by allworkers.
Study of proteolysis in animal tissue has been stimulated by the observations ofDE DUVEll on lysosomes, the subcellular organelles rich in acid hydrolases including
Abbreviation: TSH, thyrotropin.
Biochim, Biopbys. Acta, lIO (1965) 564-575

THYROID LYSOSOMAL CATHEPSIN 565
cathepsin with an optimum activity at pH 3.6. WEISS12 has shown catheptic activityin the 20 000 X g particulate fraction of the thyroid homogenate and DE DUVEll hasshown the presence of lysosomes in the thyroid gland by electron microscopy. Oneof the earliest effects of TSH is to stimulate the migration of the luminal colloid intothe intracellular particulate droplets. This has been shown by histochemistry and bythe combined techniques of electron microscopy and autoradiography-e.>. Similarconclusions have recently been drawn from differential centrifugation of homogenatesof dog thyroid tissue". WOLLMAN, SPICER AND BURSTONE16 have shown histochemically the fusion of lysosomes with these colloid droplets in about IS min after theadministration ofTSH. ROUS17 has demonstrated that the acidity in the phagocytosedvacuoles can be as low as pH 3.0 in monocytes. The above studies suggest the possibility of the intracellular release of thyroxine at acid pH in the lysosomes as postulatedby NOVIKOFF18. An understanding of thyroid proteolysis must therefore include adetailed biochemical analysis of the lysosomal cathepsin. The use of gel filtration inthe study of the proteolysis of thyroglobulin by the thyroid acid protease and studiesdefining the characteristics of the thyroid lysosomes and their cathepsin are reportedin this paper.
METHODS AND MATERIALS
Preparation of lysosomal cathepsinThe fresh, cleaned beef thyroid glands were homogenized in 0.25 1'1 sucrose for
15-20 sec in a Potter-Elvehjem teflon homogenizer with working capacity of 10 mland chamber clearance of 0.004--0.005 in. in a cold room. The volume of the homogenate was adjusted to 10 ml per g wet weight ofthyroid with 0.25 M sucrose and wasfurther homogenized for about 10 sec in grinding vessels with a working capacity of30 ml and chamber clearance of 0.005-0.007 in. The homogenate was centrifuged ina refrigerated Serval centrifuge at 800 X g for 10 min and the supernatant was centrifuged at IS 000 X g for 20 min. The IS 000 X g pellet contained the lysosomes alongwith the mitochondriav.w. The supernatant was designated as the IS 000 X g supernatant. The pellets were washed twice and resuspended in 4 ml of sucrose per g wetweight of original thyroid tissue.
Enzyme assayThe total activity of the lysosomal suspension was determined by assaying the
enzymes that were released into the IS 000 X g supernatant after disrupting the lysosomes in a high-speed Virtis homogenizer for 5 min at 5°. In experiments measuringrelease of free cathepsin, aliquots of the lysosomal suspension were recentrifuged atIS 000 X g for 20 min at zero time and after incubation. The supernatants wereassayed for cathepsin by a modification of the method of ANSON2o against 2% Hb in0.1 1'1 citrate (pH 3.6) at 37°. At 2 and 20 min, the reaction was stopped with 0.3 Mtrichloroacetic acid. The trichloroacetic acid soluble prodncts were measured eitherby their absorption at 280 mfl or with the phenol reagent.
Protein-bound iodineProtein-bound iodine was determined after precipitation with 5% trichloro
acetic acid. 40 mg bovine serum albumin were added to coprecipitate the thy-
Biochim, Biophys. A cia, lIO (1965) 564-575

566 K. BALASUBRAMANIAM, W. P. DEISS JR.
roid proteins. The precipitate was digested with chloric acid as described by ZAKet al.21 and analyzed for total iodine using the eerie ammonium sulfate method asmodified by GROSSMANN AND GROSSMANN22•
Beej-thyroid extractSome studies of proteolysis were performed with an extract prepared by the
method of TODD AND TRIKOJUSB which consists essentially of homogenizing fresh beefthyroid tissue and extracting it overnight with isotonic saline (I :3, w/v) . This extractwas separated by centrifuging at 3000 X g for 2 h.
Partial purification of cathepsin from thyroid homogenate on eM-celtulose columnThe CM-cellulose (purchased from Carl Schleicher and Schuell , Co.) was washed
with 0.2 N HCI, water and 0.01 N NaOH and equilibrated with 0.005 :M citrate-o.ocrM phosphate (pH 4.7). The sample, IO ml (1:4, w]», in sucrose) of beef thyroid homogenate , disrupted in a Virtis homogenizer was applied to the column and eluted witha gradient phosphate buffer (pH 6.8) of increasing concentration from 0.001 M to 1.0M. The reservoir contained r.o M phosphate buffer, which flowed into a 500-rnl mixercontaining the 0 .001 M buffer, which in tum was connected to the column. The sizeof the column was I em X 30 em and 5-rnl fractions were collected in an automaticfraction collector. The flow rate was IS ml jh.
Gelfiltration of the products oj proteolysis oj thyroid proteinsA dog was injected with lStI, and after 72 h, the thyroid was removed, homo
genized in 0.25 M sucrose, and the IS 000 X g supernatant was incubated at 370 in0.03 M citrate (pH 3.6). z-ml aliquots were removed at zero time, 0.5 and 8 h. Reactionwas stopped by increasing the pH to 8.0. The z-ml samples were applied to a 2.5 emX ISO em column packed under gravity with Sephadex G-IOO (purchased fromPharmacia, Uppsala, Sweden) previously equilibrated with 0.02 Mphosphate (pH 6.6)containing I mg merthiolate per 40 ml and eluted with the same buffer. A constantflow rate of IS ml jh was maintained with an accu-flo pump (Beckman Instruments,Inc., Calif.). s-ml fractions were collected in an automatic fraction collector. In another experiment 10 mg of ammonium sulfate purified beef thyroglobulin, 10 mg ofbovine serum albumin, 6 mg of myoglobin (gift from Dr. F. R. N. Gurd) and 128 000
counts/min of 1311 in 2.0 ml were applied and eluted under same conditions.
RESULTS
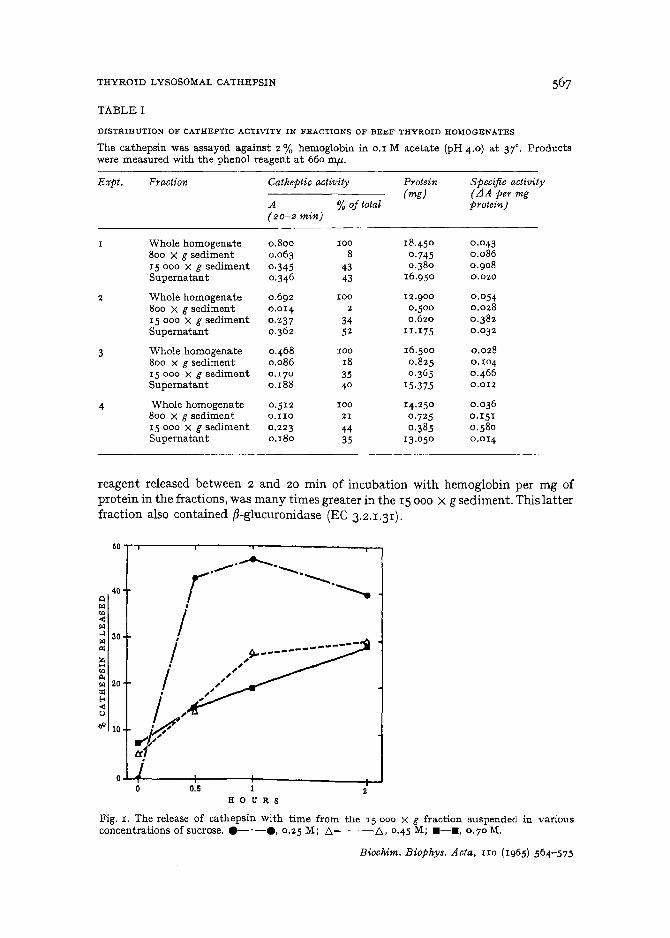
Distribution of cathepsin in various fractions oj thyroid homogenaiesHomogenates of lobes from four separate beef thyroids were prepared in 0.25 M
sucrose (IO ml per g wet weight) . 5-ml aliquots were removed, disrupted in a highspeed Virtis homogenizer, diluted r:r with sucrose and assayed for total cathepsin.The remainder of the homogenates were subjected to differential centrifugation. Theparticulate fractions were suspended in sucrose, disrupted in the homogenizer andassayed. The results are shown in Table 1. About 40% ofthe total cathepsin was sedimentable between 800 and IS 000 X g. The supernatant contained roughly similarquantities of enzyme. The specific activity of enzyme, expressed as change in absorbancy at 660 mil- of trichloroacetic acid filtrable products reacting with the phenol
Biochim, Biophys . A cta. IIO (1965) 564-575
THYROID LYSOSOMAL CATHEPSIN 567
TABLE I
DISTRIBUTION OF CATHEPTIC ACnVITY IN FRACTIONS OF BEBF THYROID HOMOGENATES
The cathepsin was assayed against 2 % hemoglobin in 0.1 M acetate (pH 4 .0) at 37°. Productswere measured with the phenol reagent at 660 mfl-.
E:o:pt. Fraction Catheptic activity Protein Specific activity(mg) (LlA per mg
A %of total protein)(20-2 min)
Whole homogenate 0 .800 100 18.450 0 .043
800 X g sediment 0.0 63 8 0 ·745 0 .086IS 000 X g sediment 0 ·345 4 3 0 .3 8 0 0 .90 8
Supern at ant 0.346 43 16.950 0.020
2 Whole homogenate 0 .692 10 0 12 .9 0 0 0 .0548 0 0 X g sediment 0.0 14 2 0 .5 0 0 0.028
15 000 X g sediment 0 .237 34 0.620 0 .3 82Supernatant 0.36 2 5 2 II .175 0 .03 2
3 Whole homogenate 0 .468 100 16 .5 0 0 0.028800 X g sediment 0.086 18 0 .8 25 0. 104
IS 000 X s sed iment 0.170 35 0 .3 65 0 .46 6
Supernatant 0 .188 4 0 15 ·375 0.012
4 Whole homogenate 0 .5 1 2 100 14.250 0 .036
800 X g sediment 0.1I0 21 0 .725 0.151
IS 000 X g sediment 0 .223 44 0 ·385 0 .58 0
Supernatant 0 .180 35 13 .050 0.0 14
reagent released between 2 and 20 min of incubation with hemoglobin per mg ofprotein in the fractions. was many times greater in the IS 000 X g sediment. This latterfraction also contained ~-glucuronidase (EC 3.2.1.31).
50
40Qt>l
'"-<f>l..l 30f>l(Z;
~'"""f>l 20:I:f<-<uIi< 10
o..l.....1~---+----+--------r-J0.5o 1
H 0 U R S
Fig. I . The release of cathepsin with time from the J5000 X g fraction suspended in variousconcentrations of sucrose. e-·-e. 0.25 M; f::,-- -f::" 0 .4 5 M; . - • • 0.70 M.
Biochim. Biophys. Acta. lID (1965) 564-575

568 K. BALASUBRAMANIAM, W. P. DEISS JR.
Latency of cathepsin in beef-thyroid lysosomesThe latent catheptic activity in the IS 000 X g fraction could be released by
low osmolar solutions as shown in Fig. 1. Aliquots of the IS 000 X g suspension indifferent concentrations of sucrose on 0.1 M Tris (pH 7.3) were incubated at 37° forvarious times and then centrifuged at IS 000 X g for 20 min. Their supernatants wereassayed for released cathepsin. In 0.45 M and 0.7 M sucrose, the release of cathepsinwas almost linear with time up to 2 h, while in 0.25 M sucrose there was a rapid releaseof cathepsin during the first 30 min.
Fig. 2 shows the release of cathepsin at various pH values. At pH 2.8, cathepsinwas released almost instantaneously, while at pH 3.7 and 8.2, there was some delay.
100
9090
qBO
80 \ » p-._.o\ ,\ I ,\ , "'l 70 ,
70 I ,\ I .... ,
I __.cf~
I ,\ ~ 60
80~
I .»:....,~i=l
\ I 50 ./I ....r<l 50 \ ,OJ \ , 0 'r\ I 40-< 40 \ , en i/r<l
\ I\ I Ii'!H 30 \ ..d 30
\r<l \ ,,"
\ ,-6" 20 ~._~...~.'!ill: 200'''
10 10
0 02 4 5 3.6 4.6 5.6 6.6 7.6
pH pH
Fig. 2. Effect of pH on the release of cathepsin from beef-thyroid lysosomes. The 15 000 X gfraction suspension in 0.25 M sucrose-o.oj M buffers of citrate (pH 2.8 and 3.7). acetate (pH 5. I)phosphate (pH 6.1) and Tris (pH 7.3·and 8.2) was incubated at 37°.•-e. zero time; 0- - -0,30 min.
Fig. 3. Solubility of the 15 000 X g fraction soluble proteins at various pH values. Incubationswere in 0.25 M sucrose-o.og M buffers of citrate (pH 3.6), acetate (pH 4.1, 4.6. 5.1 and 5.6) andphosphate (pH 6.1, 6.6 and 7.1). 0-0, cathcptic activity; 0---0. PBU7I; "-·-A.280 mf1..
Between pH 5.1 and 7.3 the release was slow. This latency of lysosomal cathepsinmay be explained on the basis of increased stability of the lysosomal membrane ordecreased solubility of the lysosomal cathepsin near its isoelectric point, or both.
The solubility of the lysosomal cathepsin at various pH values was investigatedafter lysing the IS 000 X g fraction in a high-speed blendor and recentrifuging at15 000 X g. This supernatant was mixed with various buffers and incubated for 20
min. At the end of incubation any precipitate was separated by centrifuging atIS 000 X g and the supernatant assayed for cathepsin and protein-bound iodine.The total protein in the supernatant was measured by its absorption at 280 mfA,. The
Biochim. Biophys. Acta, no (1965) 564-575

THYROID LYSOSOMAl. CATHEPSIN 569
results in Fig. 3 show that the thyroglobulin and cathepsin were least soluble atpH 4.6 and 5.1, respectively. At and above pH 5.6, both were soluble so that the observed latency oflysosomal cathepsin between pH 5.6 and 7.3 demonstrated in Fig . 2
must therefore be due to the binding of the cathepsin in or on the lysosomes.
Effects of sulfhydryl reagents, pH and temperature on the activity of lysosomal cathepsinThe lysosomal cathepsin was prepared as for the determination of the total
catheptic activity in 15 000 X g fraction. This cathepsin was incubated with I • ro-3 Mand 5' 10-8 M p-chloromercuribenzoate and iodoacetate at 37° in 0.05 M citrate
0.25 .,...,.....--,--r----r--,-r--r-r---,
106pH
Fig. 4. The effect of pH on the activity of lysosomal cathepsin. The lysosomal enzyme in 0 .1 Mbuffers was incubated for 2 h at 37° and then assayed.
o
'"..Eo<
-<I><U:z;-<~
~ 0.10
'"OJ-e<I
~ 0.20
(pH 4.0) for 1 h and then assayed for activity. There was no inhibition indicating thatthe lysosomal cathepsin is not a sulfhydryl enzyme.
The effect of pH at 37° for 2 h shown in Fig. 4 demonstrates that the lysosomalcathepsin lost activity at and below pH 2.5. Between pH 3.6 and 8.4, the cathepsinwas stable under the conditions employed. The lysosomal cathepsin incubated at 60°in phosphate buffer (pH 7.3) lost 75% of its activity in the first 10 min and almost itswhole activity by 20 min.
pH optimum and partial purification of thyroid cathepsinIn Fig. 5 observation of the curve of absorbancy at 280 mp, of trichloroacetic
acid-soluble products formed by reacting thyroid extracts- with hemoglobin at different pH values suggests that there are two peaks of catheptic activity at pH 4.2 and7.5, respectively. However, when the trichloroacetic acid products were assayed withthe phenol reagent only the pH 4.2 peak was noted. Further evidence that the alkalinepeak is not due to catheptic activity is shown in Fig. 6. The absorption maximumof the pH 7.5 experiment is at 263 m,u, suggesting that these are not primarily products of proteolysis but possibly are nucleic acids, whereas the maximum of the pH-
Biochim, Biophys. Acta, IrO (1965) 564-575

570 K. BALASUBRAMANIAM, W. P. DEISS JR.
4.2 experiment is at 275 mJ.l which is compatible with the release of aromatic aminoacids by the cathepsin .
Additional proofthat there is only the acid cathepsin in the thyroid is presentedin Fig. 7 where it can be seen that autolysis of the IS 000 X g supernatant from dogthyroid occurs optimally at pH 3.6 and that a partially purified thyroid cathepsin(obtained from P eak II of F ig. 8) has maximal activity at the same pH. Fig. 8 also
s.t
/> -,
e.e f ~II,,,
>< a.5,I
t.l,,
z\
,< I
'" 0• •. ,
0: \\0
"' \;'" a.3<<l
0.2 III ,I II I, I,
~o. i II \, \
I,,
I ,I
' t>-" o._I
7 9 10 11
p I!
Fig. 5. Catheptic activity of thyroid extract at varying pH values against urea -denatured hemoglobin : .-e, a.bsorp tion of t richloroacetic acid-soluble produ cts a.t 280 mp: 0 - - - 0,tyrosine at 660 tsus: Bu ffer concentration was 0.04 M. Buffers were citrate, acetate, phosphate,Tris and glycine. Incubation : 1 ml thyroid homogenate with 4 ml hemoglobin at 37° for 2 and30 min .
demonstrates that 95% of the cathepsin is in Peak II, while most of the protein isin Peak 1.
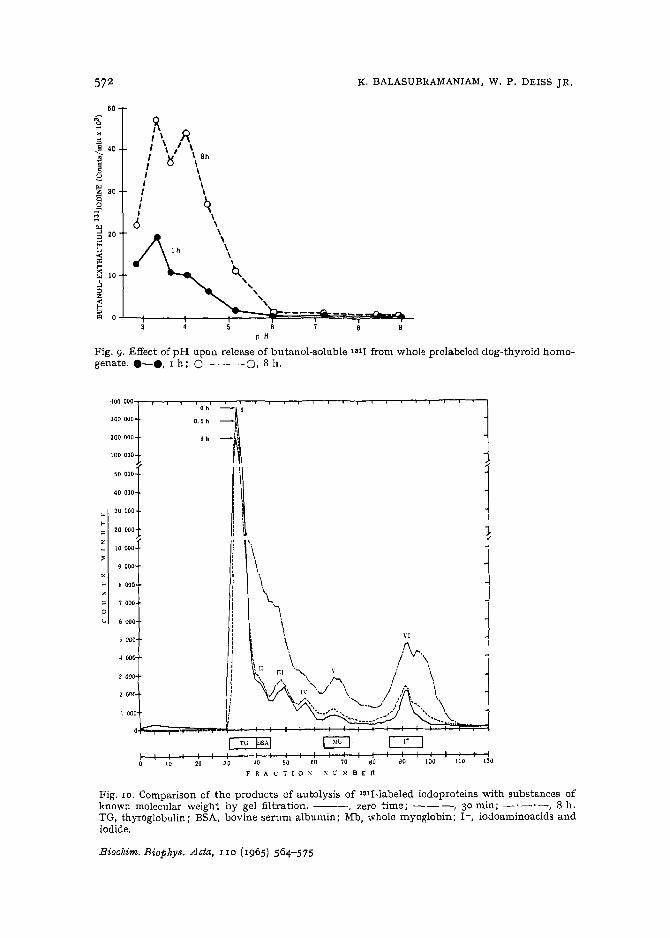
Another effort to demonstrate an alkaline protease in thyroid tissue that iscapable of attacking thyroglobulin was made by incubating fresh whole homogenatefrom a dog prelabeled 72 h previously with 1811. The homogenate was prepared in0 .25 M sucrose, strained through gauze, and brought to a volume of 44 ml withsucrose containing methiofate. After adding 3-ml aliquots to equal volumes of 0 .1 Mbuffer (glycine, citrate, acetate, phosphate, and Tris) and incubating for I and 8 h,the reactions were stopped with 0.2 ml of ro % H 2S0 4 and extracted with 4 ml ofbutanoltwice. Fig.9 shows that butanol-soluble lSII-containing products of proteolysiswere released maximally at pH 3.3 and 4 .0 and that essentially no evidence of proteolysis was evident above pH 6.0.
Biocbim. Biopbys . Acta, lID (1965) 564-575

THYROID LYSOSOMAL CATHEPSIN 57!
P.0.8 >
:l:~....zn.......
0.4 ~
o
0.2
• , 7
_"
?"~t1./ II
I ,I •I ,i ,
I I
9 \: ~I Ir •I 'I \I •
: \\I I! I
I I, ,I I
i ~ 0\& III\\,\ e__•b
,--,----r---,,....---,-----,---r,O.•
'000
~
~
z_ 4000
".>,
'"~z~
oU 2000
4
I 1'00
I I200 270 280 390
W A VEL ENe T H (m,u.)
250
o.e
01 I
J. ar-r---,.......---r----,----..-----,---,
I .•
..-< 0.8
..o "1.2Z
<..= 1. 0o
Fig. 6. Absorption spectra ofthe trichloroacetic acid-soluble products. .--e. pH 4.2; 0---0,pH 7+ Upper curves, 30 min; lower curves, 2 min.
Fig. 7. Comparison of pH optimum of partially purified cathepsin against (urea-denatured)hemoglobin with autolysis in the 15 000 x g supernatant containing 19BI-Iabeled iodoproteinsfrom dog thyroid labeled in vivo. Incubation: 37°, o. I M citrate (pH 3.5). 0- - -0. 280 tnu,trichloroacetic acid-soluble products between 2 and 30 min; e-e. counts/min, of trichloroacetic acid-soluble radioactivity between 0 and 8 h.
20 T"""--r--,---,---.,--.,--,.--,.--.,--..----.
18
0.5
\•
•
Cf-<rI I
: \: \I \I II II I
: 9I \I \I ,J ,I ,I II II I
1·....\ \., V·~.-.-.-~ \ \
1 ....,0-<),,'" ....,d ...~_~
'"oV1
'"<
~ Ho"" 12""r<< IIIr<z~ 11~
...l...c,w..o;.. 6:.JZ<'"
homogenate. eo effluent absorbancy;
Biochim, Biophys. Acta, IIO (1965) 564-575
572 K. BALASU BRAMANIAM, W. P. DEISS J R.
R 0I \ "I \ I \I .,/ \ BhI U \I \, \
I \
: qI \
6 \\,
\\
\c.,,,"
50
::J'" 20;::u
~ 10.!loz~ill O.L_ +-__-; -+====:=:~===;pJ-==~?=~-~-
6P H
Fig. 9. E ffect of pH upon release of butanol-soluble lOll from whole prc1a belecldog-thyroid homo.gen ate. e-e, 1 h; 0- - - 0, 8 h.
vt
'.\\\\
"\\\. ,'\r.
III \,. h. / \
,v<: \. / ,/. \."<; .....J. .. \
....., .., / \ .
,,,,!iIII
Ii,IIII
)
I
s 000
c
~ 000
=- 'j 000
I- tI 000
9000
_ 10 000
lJ 6000
~oo OOOlaoo 000
200 000
100 000
:::::f30000
..~ 20 000
[ Tv [DSAI G C2:::Jf- I I I I I I I I I I I I I I I I I0 '0 20 30 ~. so 6O 70 .0 go '00 110 120
r R A C T I 0 x x V ~f a E n
Fig . 10. Comparison of t he products of au tolysis of l8lI-labeled iodoproteins wi th substances ofkn own molecular weight by gel filt ration. - --, zero time ; - - -, 30 min ; - '- '-, 8 h.TG , thyroglobulin; BSA, bovine serum albumin; Mb , whole myoglobin; 1- , iodoaminoacids andiodide .
B iochim , Biophys. Acta, JI O (1965) 564-5 75

THYROID LYSOSOMAL CATHEPSIN 573
Products of proteolysis of iodoproieins by thyroid cathepsinHydrolysis of thyroglobulin by thyroid cathepsin as measured by the trichloro
acetic acid-soluble products shows that thyroglobulin is not readily attacked by thecathepsin. It seemed possible that the proteolytic activity against this protein wouldbe more adequately analyzed by looking for intermediate products by gel filtration.The results in Fig. 10 indicate that at pH 3.6 thyroid cathepsin attacks the iodoprotein selectively releasing iodopeptides of high molecular weights and smallpeptides and amino acids. The iodopeptides in Peaks II and V have filtration behaviorsimilar to bovine serum albumin and whale myoglobin, whose molecular weights are70000 and IS 000, respectively. Peak I contained the unhydrolyzed thyroglobulinand Peak VI the iodoamino acids and iodide. The iodotyrosines and iodothyroninesin Peak VI were identified by column chromatography-e. The calculated molecularweights of the large peptides in Peaks III and IV are approx. 50 000 and 30 000.
respectively.
DISCUSSION
The presence of lysosomes in the thyroid gland was demonstrated by the biochemical criteria set forward by DE DUVEll. The IS 000 X g fraction contains themajority of the sedimentable catheptic activity shown in Table I, but this is onlyabout 40% of the total present. It is probable that under the conditions used tohomogenize this fibrous tissue, damage to the lysosomes resulted in a lower recoveryas has been shown by others in other tissuesw.
Release of cathepsin from thyroid lysosomes was shown to vary with pHand osmolarity of the bathing fluid. Release of enzyme was inhibited by increasingconcentrations of sucrose and was enhanced at pH values below 5.1 and above7.3. Similar results have been obtained by others using liver lysosomal preparations24 . Since both thyroglobulin and cathepsin exist within intracellular particles, it seems highly probable that the controlled release of thyroxine from itspeptide linkage occurs in some modified intracellular particle-the "derived lysosomes" of DE DUVE. It follows then that this reaction must occur at an ambient pHabove 5.0 as the lysosomes are unstable below this pH, even though the optimumpH of the cathepsin in vitro is at 3.6. We have previously shown that thyrotropinenhances the migration of thyroglobulin from the lumen into intracellular droplets-"and studies in progress suggest that the lysis of thyroglobulin in these particles isoptimal at pH 5.0.
Evidence was presented that thyroid lysosomal cathepsin is not a sulfhydrylenzyme, that it attacks hemoglobin optimally at pH 3.6, and that it is inactivated atpH below 2.5 and 37° and at 60° at pH 7.3. These characteristics simulate those ofcathepsin D (EC 344.23) (ref. 25). The presence of more than one protease could notbe demonstrated from our studies of pH optima of catheptic activity against hemoglobin or of autolysis of labeled thyroid iodoprotein nor was more than one peak ofcathepsin found on CM-cellulose chromatography. While these studies effectivelyexclude significant proteolytic activity at alkaline pH in thyroid extracts and wholehomogenates, they cannot completely exclude the presence of several cathepsins withclosely similar characteristics.
Biochim. Biophys. Acta, IIO (Ig65) 564-575

574 K. BALASUBRAMANIAM, W. P. DEISS JR.
The gel filtration experiments show accumulation of large polypeptides whichwould not be soluble in trichloroacetic acid. The accumulation of these large polypeptides may reflect the specificity of the thyroid cathepsin. This would account forthe slow rate of further destruction of these peptides. Another possibility is that thetertiary structure of the thyroglobulin and the polypeptide products limits the rate ofproteolysis. These two possible explanations are not, of course, mutually exclusive.ALPERS, PETERMANN AND RALL26 have reported the formation of large fragments ofthyroglobulin in various states of aggregation along with small fragments during theautolysis of partially purified labeled thyroglobulin. LnoNJuA2? has shown therelease of a polypeptide of molecular weight 29000 in z h and smaller fragments ofmolecular weight 1500 in 6 h from the proteolysis of thyroglobulin by thyroid protease.Our experiments with gel filtration show the release of four polypeptides of molecularweights 70 000, 50 000, 30 000, and 18000 and a peak containing iodoamino acids,iodide and probably also small peptides. Peaks II, III and IV can be the tetramer,trimer and dimer, respectively, of the polypeptide in Peak V, and this possibilitycannot be excluded. Other similar experiments were performed, however, in whichthe reservoir and the eluting buffer contained 1.0 M NaCl, Exactly similar patternsof peak elution were observed, suggesting that such aggregation is not the likelyexplanation.
ACKNOWLEDGEMENTS
This investigation was supported by Research Grant AM-6IZ8 and TrainingGrant AM-SI73 from the National Institute of Arthritis and Metabolic Diseases,United States Public Health Service.
REFERENCES
I J. B. ALPERS, J. ROBBINS AND J. E. RALL, Endocrinology, 56 (1955) rIO.2 A. D. LITONJUA, Endocrinology, 67 (1960) 829.3 M. T. MCQUILLAN, J. D. MATHEWS AND V. M. TRIKOJUS, Nature, 192 (196 1) 333·4 E. DE ROBERTIS, Anat. Record. 80 (1941) 219.5 M. T. MCQUILLAN AND V. M. TRIKOJUS, Australian]. Bioi. s«., 6 (1953) 61 7.6 M. CLEMENTS AND A. GORBMAN, Bioi. tsuu., 108 (1955) 258.7 H. M. HADDAD AND J. E. RALL, Endocrinology, 67 (1960) 4- 13.8 P. E. E. TODD AND V. M. TRIKOJUS, Biochim, Biopbys, Acta, 45 (19 61) 234·9 L. F. KRESS, H. M. KLITGAARD AND R. J. PEANASKY, Federation Proc., 23 (1964) 434·
10 C. G. SHAPLAND, ]. Med. Lab. Technol., 21 (1964) I.II C. DE DUVE, in T. HAYASHI, Subcellular particles, Ronald Press, New York, 1959, p. 128.12 B. WEISS, ]. Bioi. Chem., 205 (1953) 193.13 N. J. NADLER, S. K. SARKAR AND C. P. LEBLOND, Endoorinology, 71 (1962) 120.14 O. STEIN AND J. GROSS, Endocrinology, 75 (19 64) 787.IS K. BALASUBRAMANIAM, W. P. DEISS, \V. C. TAN AND R. C. POWELL, Endocrinology, 77 (19 65)
54·16 S. H. WOLLMAN, S. S. SPICER AND M. S. BURSTONE, ]. Cell tuot., 21 (1964-) 191.17 P. Rous, j. Exptl. Med., 41 (1925) 399.18 A. B. NOVIKOFF. in A. V. S. DE REUCK AND M. P. CAMERON, Lysosomes, Little Brown,
Boston, Mass., 1963, p. 36.19 G. WEISSMANN AND L. THOMAS, j. E:r:ptl. Med., II6 (196:Z) 433·20 M. L. ANSON, j. Gen. Fhysiol., 22 (193 11 ) 79.21 B. ZAK, H. H. WILLARD, G. B. MYERS AND A. J. BOYLE, Analyt. Chem., 24 (1952) 1345·22 A. GROSSMANN AND G. F. GROSSMANN, Clin. En.docrinol., X5 (1955) 354·
Biochim, Biopbys, Acta. IIO (1965) 564-575

THYROID LYSOSOMAL CATHE PSIN 575
23 A. L. GREENBAUM. T. F. SLATER AND D. Y. WAN G, N ature, 188 (1960) 318.24 F . ApPLllMANS AND C. D E DVVE , Biochem, ]., 59 (1955) 426.25 E. M. PRESS. R. R. PORTER AN D J. CEBRA, B iochem , j., 74 (1960) 501.26 J. B. ALPERS, M. L. PETERMANN AN D J. E. RALL, A rch. Biochem. B iophys.• 65 (1956) 513.27 A. D. L1TONJUA, Nature, 191 (1961) 356.
Biochim, Biophys, A cta, lIO (1965) 564-575