cell cycle dependent expression of a glucose regulated cell surface glycoprotein
TRANSCRIPT

0073-5655/81/0769-0776 $01.50/0 IN VITRO Vol. 17, No. 9, September 1981 �9 1981 Tissue Culture Association, Inc.
C E L L C Y C L E D E P E N D E N T E X P R E S S I O N O F A G L U C O S E R E G U L A T E D C E L L S U R F A C E G L Y C O P R O T E I N
BARBARA J. KEYS, MARIAN HOYLE, AND ALBERT J. T. MILLIS'
Department of Biological Sciences, State University of New York at Albany, 1400 Washington Avenue, Albany, New York 12222
(Received August 27, 1980; accepted January 13, 1981 )
SUMMARY
A cell surface associated "glucose regulated protein" has been described on nontrans- formed human fibroblasts. To examine the distribution of that protein on human fibro- blasts specific antisera were used. The antisera was used in conjunction with indirect immunofluorescence and revealed that the glucose regulated protein was present as fibers on spread cells. Further, the antisera was used in complement mediated cytotoxicity assays to examine cells during specific stages of the mitotic cell cycle. Fibroblasts were synchron- ized by serum starvation, hydroxyurea inhibition, or colcemid inhibition followed by mito- tic selection. The results demonstrated that the glucose regulated protein was maximally displayed during the G, phase of the cell cycle and minimally displayed during the S and M phases.
Key words: glucose regulated protein; glycoprotein; fibroblasts; cell cycle; fibronectin.
INTRODUCTION
Recently we identified a glycoprotein of 100,000 daltons as one of the two major iodin- atible cell surface proteins released by human fibroblasts into conditioned media (1). The pro- tein is distinctive from many other cell surface as- sociated glycoproteins in that it is exquisitely sensitive to glucose availability; a trait not shared with fibronectin (1,2). Cells cultured in glucose depleted media synthesize a 97,000 dalton protein in place of the 100,000 dalton component. Because of the glucose sensitivity, molecular weight, and similarities between it and cell sur- face associated proteins from 3T3 cells (2) and chick embryo fibroblasts (3) we refer to the pro- tein as the "lOOK-glucose regulated protein" (100K-GRP).
Because the 100K-GRP has unique properties we examined its display at the cell surface during the different phases of the mitotic cell cycle. The technique of indirect immunofluorescence re- vealed that the antigen was organized into small spots on rounded ceils and into a fibrous pattern on spread cells. Although in some cases the fibers appeared to form intercellular connections there
'To whom all correspondence should be addressed.
was no evidence of antigen deposited directly on the glass substrate. Further, we used a comple- ment mediated cytotoxicity assay (4) and estab- lished that the 100K-GRP was maximally dis- played during the G, phase of the cell cycle and minimally displayed during the S phase.
MATERIALS AND METHODS
Cells and cell culture. The cell strain used in this work was a culture of human diploid fibro- blasts (HDF) that was initiated in this laboratory from a foreskin biopsy. Culture methods have been published previously (5). All experiments were performed on rapidly growing cells that did not exhibit the senescent or postreplicative growth state during this study. Cultures were maintained in Medium 199 containing 20% fetal bovine serum (Flow Laboratories, Inc., Rockville, MD), penicillin, streptomycin, 20 m M HEPES, sodium bicarbonate, and sodium hydroxide in a 3% CO2:97% air environment at 37 ~ C. Cultures of both rapidly growing and confluent cells used in the cytotoxicity assays were obtained by plating cells at 6.6 x l0 s cell/cm 2 and collecting them either after 48 h or 168 h. Cultures used for syn- chrony and cell cycle experiments were subcon- fluent at the time of each experiment.
769

770 KEYS, HOYLE, AND MILLIS
Cell synchrony. To develop synchronous populations, actively growing cells were growth arrested either by serum starvation, serum starva- tion followed by 10 -s M hydroxyurea treatment, or colcemid treatment. Those procedures pro- duced cell populations that were arrested in early G,, G,-S phase boundary or during mitosis. Growth was then initiated by the addition of 20% serum. At the indicated times the degree of syn- chrony was estimated by pulse labeling with 1/~Ci/m of [3H]thymidine for 60 min. At the time the cells were harvested, counted, and treated with 15~ cold trichloroacetic acid. The acid in- soluble material was counted in a Beckman liquid scintillation spectrophotometer. Although the incorporation of thymidine is a good indicator of the temporal position of the S phase it does not reveal the actual percentage of S phase cells. To obtain that information the cells were pulsed with 1 ~Ci/m [3H]thymidine for 30 min, fixed with 2% glutaraldehyde (Sigma Chemical Co., St. Louis, MO) for 2 h on ice, then dehydrated with 25, 50, 70, and 100% ethanol for 15 min each. The cultures were then overlayed with NB2 photo- graphic emulsion (Kodak). After 3 wk incubation in the dark at 4 ~ C, the plates were developed, stained with Giemsa (GIBCO, Grand Island, NY) and counted to determine the percent labeled nuclei (see Fig. 1 and Table 1).
To select mitotic populations, subconfluent cul- tures of fibroblasts were treated with 0.04 ~g/ml colcemid (GIBCO) for 4 h. At that time the vessels were shaken and the detached cells de- canted. After collection by centrifugation at 1500 xg for 5 min, a sample was removed to determine the mitotic index t6). Cultures that were >85% mitotic were used in the cytotoxicity assays.
Antisera production. The antigens used in this work were prepared either from human fibroblast conditioned media or from human plasma. To produce antisera to the 100K-GRP tanti-100K- GRP) conditioned media t5) were brought to 35% saturation with iNH4hSO4 and the insoluble frac- tion dialyzed against phosphate buffered saline (PBS) and passed over a Sephadex G-100 column (RP-35L Approximately 200 ~g protein, contain- ing 30 ~g 100K-GRP was electrophoresced into sodium dodeeyl sulfate (SDSj-polyacrylamide gel. After staining with Coomassie Brilliant Blue the 100K-GRP band was sliced from the gel, tri- turated, and suspended in PBS (7). New Zealnd white male rabbits were each immunized with 200 ~g of that material in Freunds adjuvant
15- /
% o . f
o
%_
l l4
4 I I I I I0 20 30
Time, hrs.
FIG. 1. Cell synchrony induced by serum starvation. Subconfluent cultures were exposed to 0.2% fetal bovine sera for 48 h. At that time the serum concentration was increased and at subsequent times both ( �9 ) [3H]thymi- dine incorporation and (O) cell density were monitored as described in Methods.
(Calbiochem, San Diego, CA) via subcutaneous injections into the neck and shoulder regions. A second injection followed after 10 d and a third injection followed the first by 30 d. The rabbits were bled from an ear vein 5 to 7 d after the third injection.
Antisera to fibronectin (antifibronectin) was produced in rabbits using either gelatin- Sepharose (8), purified conditioned media fibro- nectin (1), or human plasma fibronectin ("cold in-
TABLE 1
PERCENT LABELED NUCLEI IN SYNCHRONIZED C ULTURES OF H UMAN F IBROBLASTS
Time After Labeled Synchronizing Treatment Release Nuclei
h % Serum starvation
Hydroxyurea inhibition
0 11 29 70 30 71 31 68
0 14 1 64 2 69 3 67
Replicate cultures were released from serum starva- tion in early G, and from hydroxyurea inhibition in late G,, early S were pulse labeled with 1 ~Ci/ml[3H]thymi - dine for 30 min, fixed with glutaraldehyde, and examined by autoradiography. The percentage of labeled nuclei was determined by counting 150 to 200 cells in three trials for each time point.
EXPRESSION OF A GLUCOSE REGULATED PROTEIN 771
soluble globulin") purified in a similar manner. Anti-100K-GRP and antifibronectin do not cross react in immune double diffusion gels (1 }. In addi- tion, using conditioned media as the source of antigens, anti-100K-GRP will only precipitate a 100K protein whereas antifibronectin precipitates a 220K protein, but not a 100K component (9).
Immunofluorescence. For immunofluores- cence, cells were plated on glass cover slips and incubated for 4 to 48 h. At the desired times the cells were fixed for 30 min in 3% paraformalde- hyde, washed, and incubated with a 1:40 dilution of primary antibody. After 30 min of incubation the cell layer was washed in PBS- and subse- quently incubated in a 1:40 dilution of fluorescein isothiocyanate-conjugated sheep antirabbit anti- body (Miles, Elkhart, Indiana) for an additional 30 min. After extensive washing in PBS- the cells were examined using a Zeiss Photoscope required for epifluorescence and photographed with Kodak Tri-X film. Antibody blocking experiments were performed to assay for cross-reacting components in the antibodies to fibronectin or 100K-GRP. In those experiments cells were pretreated with a 10- fold excess (1:4 dilution) of sheep antifibronectin before addition of rabbit anti-100K-GRP. Con- versely we attempted to block antifibronectin ac- tivity with anti-100K-GRP. Neither assay re- vealed cross-reacting material in the antisera. Other evidence that the two antibodies do not cross react is provided by experiments ~not shown) in which anti-100K-GRP stains mouse endodermal cells that are not labeled with anti- fibronectin ~McCormick, et al., in preparation).
Complement mediated cytotoxicity test. The complement mediated cytotoxicity assay was a modification of Terasaki's microdroplet lympho- cyte cytotoxicity test 14). Briefly, target fibro- blasts were harvested with EDTA and PBS- un- less otherwise indicated. Exactly 2 x 103 cells in 10/~1 were incubated in each well of a Cooke coni- cal well microtiter plate at room temperature with the appropriate serum and 5 ~1 mineral oil. After 30 min, 5~1 rabbit complement ~GIBCO) or PBS- was added to each well and incubated for an additional 60 min. At that time 3 t~l freshly made 0.75% trypan blue dye was added to each well and incubated for 2 min. The cells were then fixed with 8 #1 3% formaldehyde and the percentage of cell lysis was determined using an inverted phase contrast microscope tWild}. For each determina- tion approximately 200 to 300 cells were counted. The cytotoxicity index was calculated as follows ~10}:
% dead - % dead in complement control { 1 )
100% - % dead in complement control The percentage of dead cells in the absence of
complement ranged from 20 to 22% for each point.
Protein determination. Protein concentrations of antisera samples were determined by measur- ing the optical density of the sample at 278 nm with a Gilford spectrophotometer. Protein con- centrations were calculated using known extinc- tion coefficients for bovine serum albumin (0.66 A27s U at 1 mg/ml) rabbit IgG (1.49 A27s U at 1 mg/ml}, and fibronectin I1.28 A2so U at 1 mg/ml} (11}.
RESULTS
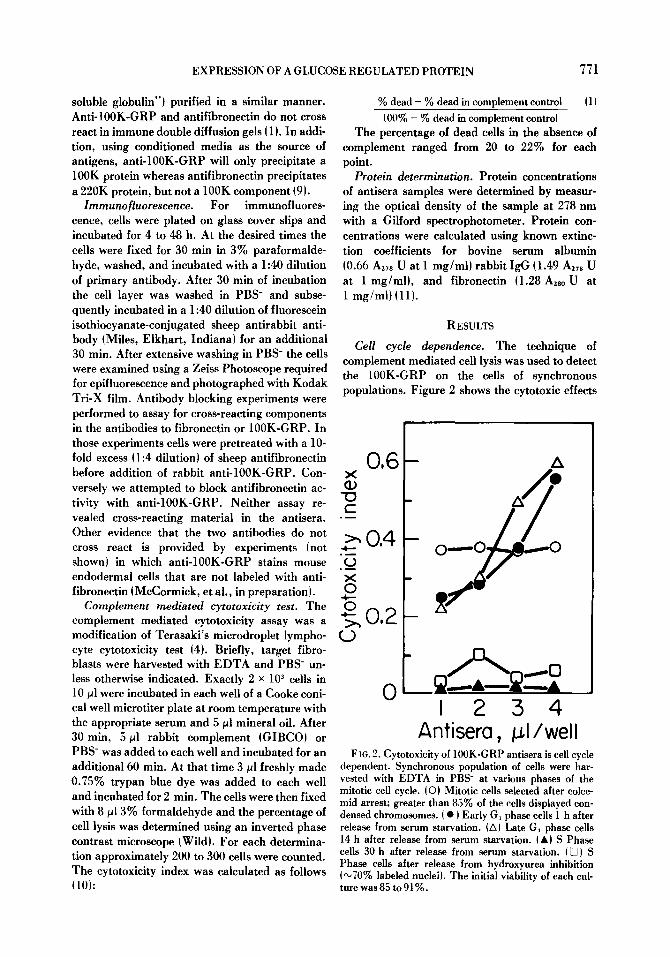
Cell cycle dependence. The technique of complement mediated cell lysis was used to detect the 100K-GRP on the cells of synchronous populations. Figure 2 shows the cytotoxic effects
0,6 X CD
" 0 t -
> , 0,4 ~
0 ~
X 0
o 0 , 2
A
m
o ~ O ,
I 2 3 4 Antisera, I/well
F 1G. 2. Cytotoxicity of 100K-GRP antisera is cell cycle dependent. Synchronous population of cells were har- vested with EDTA in PBS- at various phases of the mitotic cell cycle, iO) Mitotic ceils selected after colce- mid arrest; greater than 85% of the cells displayed con- densed chromosomes, i �9 ) Early G~ phase cells 1 h after release from serum starvation. (A) Late G~ phase cells 14 h after release from serum starvation. (A} S Phase cells 30 h after release from serum starvation. I D) S Phase cells after release from hydroxyurea inhibition (",70% labeled nuclei). The initial viability of each cul- ture was 85 to 91%.
772 KEYS, HOYLE, AND MILLIS
0,8
x o 0,6 c -
�9 - ~ 0.4 x 0
.4---
0
c~ 0.2
0
O
i:
M G~ S Cell cycle phose
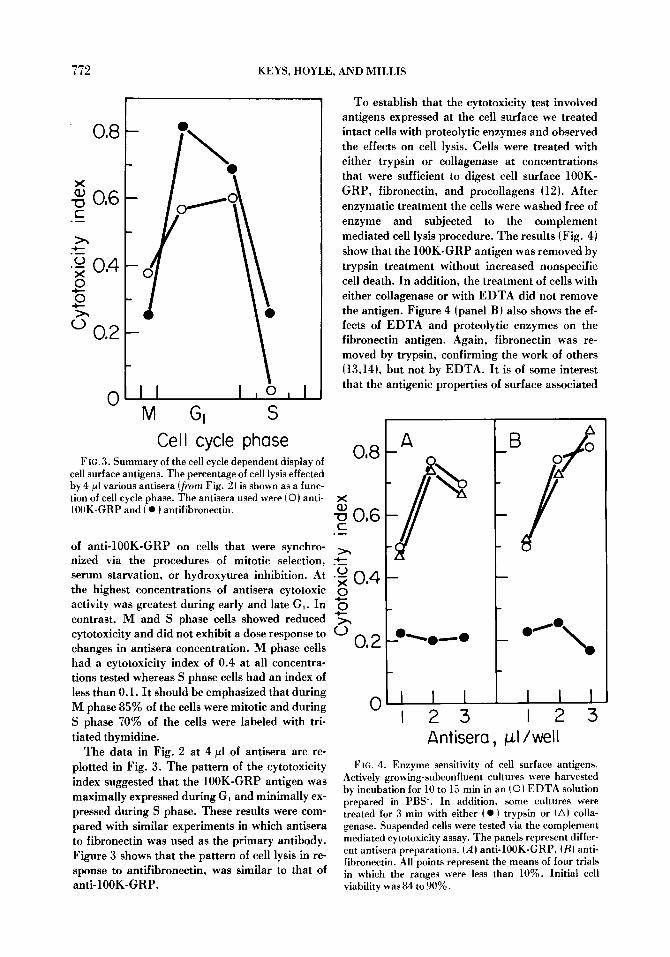
F IG. 3. Summary of the cell cycle dependent display of cell surface antigens. The percentage of cell lysis effected by 4/~1 various antisera (from Fig. 2) is shown as a func- tion of cell cycle phase. The antisera used were (0) anti- 100K-GRP and ( �9 ) antifibronectin.
of anti-100K-GRP on cells that were synchro- nized via the procedures of mitotic selection, serum starvation, or hydroxyurea inhibition. At the highest concentrations of antisera cytotoxic activity was greatest during early and late G,. In contrast, M and S phase cells showed reduced cytotoxicity and did not exhibit a dose response to changes in antisera concentration, n phase cells had a cytotoxicity index of 0.4 at all concentra- tions tested whereas S phase cells had an index of less than 0.1. It should be emphasized that during M phase 85~ of the cells were mitotic and during S phase 70% of the cells were labeled with tri- tiated thymidine.
The data in Fig. 2 at 4/~1 of antisera are re- plotted in Fig. 3. The pattern of the cytotoxicity index suggested that the 100K-GRP antigen was maximally expressed during G, and minimally ex- pressed during S phase. These results were com- pared with similar experiments in which antisera to fibronectin was used as the primary antibody. Figure 3 shows that the pattern of cell lysis in re- sponse to antifibronectin, was similar to that of anti-100K-GRP.
To establish that the cytotoxicity test involved antigens expressed at the cell surface we treated intact cells with proteolytic enzymes and observed the effects on cell lysis. Cells were treated with either trypsin or collagenase at concentrations that were sufficient to digest cell surface 100K- GRP, fibronectin, and procollagens (12). After enzymatic treatment the cells were washed free of enzyme and subjected to the complement mediated cell lysis procedure. The results (Fig. 4) show that the 100K-GRP antigen was removed by trypsin treatment without increased nonspecific cell death. In addition, the treatment of cells with either collagenase or with EDTA did not remove the antigen. Figure 4 (panel B) also shows the ef- fects of EDTA and proteolytic enzymes on the fibronectin antigen. Again, fibronectin was re- moved by trypsin, confirming the work of others (13,14), but not by EDTA. It is of some interest that the antigenic properties of surface associated
x
-o 0 , 6 r--
. D
(.) �9 ~ 0 . 4 0 - I - -
0
0.2
o, sL A
o ~ o . . . . - o
_B o
oIe%,~
0 I I I I I I I 2 3 I 2 3
Anfisera, p l /wel l FIG. 4. Enzyme sensitivity of cell surface antigens.
Actively growing-subconfluent cultures were harvested by incubation for 10 to 15 rain in an (�9 EDTA solution prepared in PBS-. In addition, some cultures were treated for 3 min with either ( �9 ) trypsin or (A) colla- genase. Suspended cells were tested via the complement mediated cytotoxicity assay. The panels represent differ- ent antisera preparations. (A) anti- 100K-GRP, (B) anti- fibronectin. All points represent the means of four trials in which the ranges were less than 10%. Initial cell viability was 84 to 90%.

EXPRESSION OF A GLUCOSE REGULATED PROTEIN 773
fibronectin are unaffected by collagenase inas- much as fibronectin is intimately associated with collagens (15) and procollagens (16) at the cell surface.
Visualization of IOOK-GRP. The technique of indirect immunofluorescence was used to examine the display of the 100K-GRP antigen on the cell surface. Cells were grown on glass cover slips, fixed, and stained for immunofluorescence as shown in Fig. 5. Figure 5.4, B show a single cell that was treated first with anti-100K-GRP and subsequently stained with goat antirabbit Ig conjugated to fluorescein isothiocyanate. The 100K-GRP antigen was organized into fibers that were evident on the cell surface. Examination of the corresponding phase-contrast photograph suggested that the fibers followed the outline of the cell. That observation was enhanced by exam- ining several cells in a more confluent culture ~Fig. 5C, DL In this case the fibers were present on the cell surface and between cells. There was no evidence of antigen deposited directly on the glass in regions that were free of cells. The fibers were less evident on rounded cells tFig. 5E) and the fluorescence was detected in bright regions or spots. Comparison between 100K-GRP and fibronectin (another major surface associated pro- tein) revealed that both were organized into fibers {not shown} on spread cells and into spots on rounded cells tFig. 5EL
D ISCUSSION
The 100K-GRP is a membane associated glyco- protein whose glycosylation is particularly sensi- tive to the availability of glucose in the culture media (1 }. In this report we show, by immuno- fluorescence, that the antigen is organized into fibrous arrays at the cell surface. Those fibers may be a reflection either of a fibrous organiza- tion of 100K-GRP or that the antigen is bound to an unidentified fibrous component. The 100K- GRP fibers were similar, but not identical, to fibronectin fibers observed by us (not shown~ and by others (14,17L The fibers were not detected as deposits on the glass substrate or on regions free of cells. Glucose regulated proteins of similar molecular weights are known to exist on other vertebrate fibroblasts. Indirect immunofluores- cence, using an antisera to a mouse 97K-GRP, re- vealed that the membrane associated antigen of those 3T3 cells appeared as spots rather than fibers ~181. The differences between the two pat- terns may be due to the degree of cell spreading or
differences in the amount of extracellular matrix produced by different cell types. For example, on rounded human fibroblasts the 100K-GRP anti- gen was also present as "spots" (Fig. 5E). Under conditions that precede flattening and spreading, the usual staining pattern showed small loci of immunofluorescence distributed over the entire surface of the cell. The transition to the fibrous pattern required about 48 h (Chandrasekhar and Millis, in preparation). In data not shown, we examined 3T3 cells using anti-100K-GRP and ob- served a similar transition from foci to a fibrous pattern associated with increased cell spreading. However, the antigen on 3T3 cells never becomes as fibrous as that observed on human fibroblasts. 3T3 cells do not spread as well as human fibro- blasts and apparently synthesize a less complex extracellular matrix (19). Although we have not examined the mechanism of fiber formation we do not believe that it results from disulfide bonds. Unreduced 100K-GRP runs faster on SDS- polyacrylamide gels than does reduced 100K- GRP. This suggests the presence of intrachain but not interchain disulfide bonds. I t is possible that the fibers result from interchain bonds catalyzed by cellular transglutaminases. A recent report shows that transglutaminases are associated with cell surface proteins on nontransformed cells (20).
To examine the expression of cell surface 100K- GRP during the cell cycle we used a complement mediated cytotoxicity assay and synchronized populations of cells. The results demonstrated that fibroblasts were maximally sensitive to anti- 100K-GRP during the GI phase. Cells were mini- mally sensitive during the M and S phases. During those latter two phases we were unable to demonstrate a dose response to the antibody sug- gesting a residual level of nonspecific complement dependent cell lysis. In addition, we used anti- fibronectin in our assay and found that surface fibronectin was also highest in G, and minimally expressed during S and M phases. Previously, a cell cycle dependent distribution was reported for fibronectin by two groups using different tech- niques. Both found high levels in G, low levels in M (13,14) but disagreed on the S phase ex- pression. Our results with complement mediated cytotoxicity more closely support the results ob- tained by Hynes and Bye t13).
In experiments using cells that had been pre- treated with either trypsin or collagenase we es- tablished that both the anti-100K-GRP and anti- fibronectin targets were removed by trypsin, but not by collagenase. This is further evidence that

774 KEYS, HOYLE, AND MILLIS
neither glycoprotein is collagenous (121 and sug- gests that they are not attached to the cells via a collagenous link.
There are other reports of specific cell surface changes during the cell cycle (21,22) associated with max imum expression during the G, phase.
FIG. 5. Distribution of 100K-GRP on human fibroblasts. IA) 100K-GRP on single cell, {B) phase contrast of A, ~C) 100K-GRP on sheet of cells, ID) phase contrast of C, (E} 100K-GRP on rounded cell, iFi fibronectin on rounded cell, IG) preimmune sera control, and lHI phase contrast of G. Cells were stained with primary and secondary antibodies, visualized via epifluorescent illumination using a 40"Z objective with oil.

EXPRESSION OF A GLUCOSE REGULATED PROTEIN 775
Those results together with the results reported here suggest that surface expression during G, may be the result of active protein synthesis and membrane assembly following cell division (23) and may reflect an increased requirement for transport proteins during the GI period of the cell cycle (24).
In this report we established the cell cycle de- pendent display of the 100K-GRP using the tech- nique of complement mediated cytotoxicity. Al- though the technique is quite sensitive it is always possible that the results may be dependent upon differential membrane fragility (25L or steric ar- rangement of the antigens (26). We cannot ex- clude these possibilities and, in fact, they may ex- plain the high level of cytotoxicity that we consis- tently observe with mitotic cells.
A relationship between the display of the 100K- GRP and cellular proliferation remains to be es- tablished. However, there is circumstantial evi- dence to suggest that the relationship exists. First, several recent reports associate glycoproteins of about 100,000 daltons with oncogenesis and transformation (27,28) and to be "glucose regu- lated" on transformed murine (2), avian (3), and human (1) fibroblasts. Second, there is the sugges- tion that glucose regulated proteins account for the increased rate of sugar uptake by glucose starved cells (29). Finally, this report demon- strates that the 100K-GRP is differentially ex- pressed at the cell surface during the cell cycle.
REFERENCES
1. McCormick, P. J.; Keys, B. J.; Pucci, C.; Millis, A. J. T. Human fibroblast-conditioned medium contains a 100K dalton glucose-regulated cell sur- face protein. Cell 18: 173-182; 1979.
2. Pouyssegur, J.; Yamada, K.M. Isolation and immunological characterization of a glucose- regulated fibroblast cell surface glycoprotein and its nonglycosylated precursor. Cell 13: 139-150; 1978.
3. Shin, R. P. C.; Pouyssegur, J.; Pastan, I. Glucose depletion accounts for the induction of two trans- formation-sensitive proteins in Rous sarcoma virus-transformed chick embryo fibroblasts. Proc. Natl. Acad. Sci. USA 74: 3840-3844; 1977.
4. Terasaki, P. I.; Park, M. S. Microdroplet lympho- cyte cycotoxicity test. Ray, J. G.; Hare, D. B.; Pederson, P. D.; Mullally, D. I. eds. NIAID manual of tissue typing techniques. Washington, DC: U.S. Government Printing Office; 1976: 69-8O.
5. Millis, A. J. T.; Hoyle, M.; Field, B. Human fibro- blast condkioned media contain growth promot- ing activities for low density cells. J. Cell Physiol. 93: 17-24; 1977.
6. Millis, A. J. T.; Forrest, G.; Pious, D. A. Cyclic AMP dependent regulation of mitosis in human lymphoid cells. Exp. Cell Res. 83: 335-343; 1974.
7. Goldman, J.E.; Schaumberg, H.H.; Norton, W. T. Isolation and characterization of glial fila- ments from human brain. J. Cell Biol. 78: 426-440; 1978.
8. Engvall, E.; Ruoslahti, E. Binding of soluble form of fibroblast surface protein, fihronectin, to colla- gen. Intl. J. Cancer 20: 1-5; 1977.
9. Garvey, J.S.; Cremer, N.E.; Sussdorf, D.H. Methods in immunology. Reading, MA: Addison-Wesley Publ. Co., Inc.; 1977.
10. Cheng, C. C.; Bennett, D, Nature of the antigenic determinants of T-locus. Cell 19: 537-543; 1980.
1l. Mosesson, M. W.; Umfleet, R. A. The cold in- soluble globulin of human plasma. J. Biol. Chem. 245: 5728-5736; 1970.
12. McCormick, P.J. ; Chandrasekhar, S.; Millis, A. J. T. Direct visualization of collagens and procollagens in polyacrylamide gels. Anal. Bioch. 97: 359-366; 1979.
13. Hynes, R. O.; Bye, J. M. Density and cell cycle dependence of cell surtace proteins in hamster fibroblasts. Cell 3:113-120; 1974.
14. Stenman, S.; Wartiovaara, J.; Vaheri, A. Changes in distribution of a major fibroblast protein, fibronectin, during mitosis and interphase. J. Cell Biol. 74: 453-467; 1977.
15. Yamada, K. M.; Olden, K. Fibronectin-adhesive glycoproteins of cell surface and blood. Nature 275: 179-184; 1978.
16. Duksin, D.; Maoz, A.; Fuchs, S. Differential cyto- toxic activity of anticollagen serum on rat osteo- blasts and fibroblasts in tissue culture. Cell 5: 83-86; 1975.
17. Chen, L. B.; Burridge, K.; Murray, A.; Walsh, M. L.; Copple, C. D.; Bushnell, A.; McDougal, J. K.; Gallimore, P. H. Modulation of cell sur- face glycocalyx: studies on large, external, trans- formation sensitive protein. Ann. NY Acad. Sci. 312: 366-381; 1978.
18. Pouyssegur, J.; Yamada, K.M. Isolation and immunological characterization of a glucose regu- lated fibroblast cell surface glycoprotein and its non-glycosylated precursor. Cell 13: 139-150; 1978.
19. Culp, L. A.; Bensusan, H. Search for collagen in substrate adhesion of two murine cell lines. Nature 273: 680-682; 1978.
20. Birckbichler, P.J. ; Patterson, M.K. Cellular transglutaminase, growth, and transformation. Ann. NY Acad. Sci. 312: 354-365; 1978.
21. Pellegrino, M.A.; Ferrone, S.; Natali, P.G.; Pelligrino, A.; Reisfeld, R. A. Variation in sus- ceptibility o[ a human lymphoid cell llne to im- mune lysis during the cell cycle. J. Immunol. 108: 573-576; 1972.
22. Cikes, M.; Friberg, S. Expression of cell surface antigens on cultured cells. Poste, G.; Nicolson, G. eds. Dynamic aspects of cell surface organiza- tion. New York: Elsevier-North Holland, Inc.; 1977: 473-511.

776 KEYS, HOYLE, AND MILLIS
23. Baserga, R. Multiplication and division in mam- malian cells. Marcel Dekker, Inc.; 1976:17-52.
24. Sander, G.; Pardee, A. B. Transport changes in synchronously growing CHO and L cells. J. Cell. Physiol. 80: 267-272; 1972.
25. Pellegrino, M.A.; Ferrone, S.; Cooper, N.R.; Dierich, M. P.; Reisfeld, R. A. Variation in sus- ceptibility of a human lymphoid cell line to im- mune lysis during the cell cycle. J. Exp. Med. 140: 578-590; 1974.
26. Moiler, E.; Moiler, G. Quantitative studies of the sensitivity of normal and neoplastic mouse cells to the cytotoxic action of isoantibodies. J. Exp. Med. 115: 537-553; 1962.
27. Bramwell, M. E.; Harris, H. An abnormal mem- brane glycoprotein associated with malignancy in a wide range of different tumors. Proc. R. Soc. Lond. {Biol. ) 201 : 87-106; 1978.
28. Kamm, A. R.; Grimes, W. J. Isolation and char- acterization of a tumor cell surface antigen from spontaneously transformed BALB/c mouse fibroblasts. Proc. Natl. Acad. Sci. USA 75: 5912-5916; 1978.
29. Banjo, B.; Perdue, J. F. Increased synthesis of selected membrane polypeptides correlated with increased sugar transport sites in glucose starved CEF (abstr.). J. Cell Biol. 70: 270a; 1976.
The authors thank Dr. George Feigen (Stanford U.) for advice and help with antisera production; Dr. L. B. Chen (Sidney Farber Cancer Institute) for contributing antibody to human plasma fibronectin; Dr. P. J. McCormick (Sloan Kettering Cancer Research Institute) for discussion during the course of the work; and Dr. J. Mazurkiewicz (Albany Medical College) for advice on immunofluorescence. Research was supported by contract AG00697 from the National Institutes of Health.