carbon sequestration in forest soils: effects of soil type, atmospheric co2 enrichment, and n...
TRANSCRIPT

Carbon sequestration in forest soils: effects of soil type,atmospheric CO2 enrichment, and N deposition
F . H A G E D O R Na , S . M A U R E R
b , P . E G L Ia , P . B L A S E R
a , J . B . B U C H E Ra & R . S I E G W O L F
b
aSwiss Federal Institute for Forest, Snow and Landscape Research (WSL), ZuÈrcherstrasse 111, 8903 Birmensdorf, and bPaul Scherrer
Institut, 5232 Villigen-PSI, Switzerland
Summary
Soil contains the major part of carbon in terrestrial ecosystems, but the response of this carbon to
enriching the atmosphere in CO2 and to increased N deposition is not completely understood. We studied
the effects of CO2 concentrations at 370 and 570�mol CO2 mol±1 air and increased N deposition (7
against 0.7 g N m±2 year±1) on the dynamics of soil organic C in two types of forest soil in model
ecosystems with spruce and beech established in large open-top chambers containing an acidic loam and
a calcareous sand. The added CO2 was depleted in 13C and thus the net input of new C into soil organic
carbon and the mineralization of native C could be quanti®ed.
Soil type was the greatest determining factor in carbon dynamics. After 4 years, the net input of
new C in the acidic loam (670 6 30 g C m±2) exceeded that in the calcareous sand (340 6 40 g C m±2)
although the soil produced less biomass. The mineralization of native organic C accounted for
700 6 90 g C m±2 in the acidic loam and for 2800 6 170 g C m±2 in the calcareous sand. Unfavourable
conditions for mineralization and a greater physico-chemical protection of C by clay and oxides in
the acidic loam are probably the main reasons for these differences. The organic C content of the
acidic loam was 230 g C m±2 more under the large than under the small N treatment. As suggested
by a negligible impact of N inputs on the fraction of new C in the acidic loam, this increase
resulted mainly from a suppressed mineralization of native C. In the calcareous sand, N deposition
did not in¯uence C concentrations. The impacts of CO2 enrichment on C concentrations were small.
In the uppermost 10 cm of the acidic loam, larger CO2 concentrations increased C contents by 50±
170 g C m±2. Below 10 cm depth in the acidic loam and at all soil depths in the calcareous sand,
CO2 concentrations had no signi®cant impact on soil C concentrations. Up to 40% of the `new'
carbon of the acidic loam was found in the coarse sand fraction, which accounted for only 7% of
the total soil volume. This suggests that a large part of the CO2-derived `new' C was incorporated
into the labile and easily mineralizable pool in the soil.
Introduction
The increase in atmospheric CO2 is indisputable, but sinks for
this carbon are not well understood. Combustion of fossil fuel
and deforestation lead to an emission of 6.6 3 1015 g C year±1,
whereas the atmospheric increase and the oceanic uptake
account for only about 5.2 3 1015 g C year±1. This leaves an
unknown sink of about 1.4 3 1015 g C year±1 (Schimel, 1995).
One hypothesis is that this `missing' C is being stored in
terrestrial ecosystems of the northern hemisphere, probably as
a result of increased plant growth induced by larger
concentrations of CO2 and a greater deposition of N (Ciais
et al., 1995; Townsend et al., 1996; White et al., 2000).
Increases in plant growth in response to CO2 enrichment are
experimentally well documented. Forest inventories show an
accelerated forest growth in the northern hemisphere (Speicker
et al., 1996). Carbon ®xed within plant biomass ultimately
enters the soil, where it may reside for hundreds of years. The
ability of soils to store this additional C, however, is highly
controversial, because there are two contrasting ways in which
the increased input of C may be processed in the soil. First, the
extra-®xed C may become soil organic C. Second, this readily
available source of C may stimulate soil microbial processes
by providing substrates that enhance decomposition of the
organic matter through the so-called `priming-effect' (Paterson
et al., 1997). Strong evidence for a long-term sink for
increased atmospheric CO2 in soils is still lacking
(Schlesinger, 1990; Schimel, 1995; Canadell et al., 1996).
Paper given at Eurosoil 2000.
Correspondence: F. Hagedorn. E-mail: [email protected]
Received 12 October 2000; revised version accepted 3 April 2001
European Journal of Soil Science, December 2001, 52, 619±628
# 2001 Blackwell Science Ltd 619

The impacts of CO2 enrichment on soil C dynamics have
been studied mainly in grasslands and agricultural systems
(e.g. Leavitt et al., 1994; Torbert et al., 1997; van Kessel
et al., 2000). Inputs of `new' C from CO2 into soils of
these ecosystems were estimated by using CO2 that was
depleted in 13C compared with ambient atmospheric CO2.
Results indicated that substantial amounts of atmospheric
CO2 are ®xed in soil: up to 25% of the total organic C in
the soil (700 g C m±2) can be derived from the added
atmospheric CO2 after 4 years (van Kessel et al., 2000).
Forests play a major role in the global carbon cycle.
Between 62 and 78% of the global terrestrial C is
sequestered in forests, and about 70% of this C is stored
in the soil (Dixon et al., 1994; Schimel, 1995). Forest soil
tends to accumulate more C than does soil under
agriculture, because the carbon turns over more slowly
(Guggenberger et al., 1994). Thus, forest soils may store
more C than agricultural soils, and their responses to
increasing atmospheric CO2 concentrations will be signi®-
cant for the future global carbon cycle.
Canadell et al. (1996) have suggested that soil quality may
in¯uence sequestration of C in response to increased atmo-
spheric CO2. Soil fertility may control the C inputs into the
soil, since CO2 enrichment can stimulate plant growth only in
soils with adequate nutrients (Egli et al., 1998). Moreover, soil
properties in¯uence the decomposition of organic C in the soil,
because they determine the living conditions for microbes and
protect C in the soil (Martin & Haider, 1986; Christensen,
1992). Most studies on CO2, however, have been made on only
one soil each, and if fertility was taken into account, fertilizers
were applied in different amounts.
The quantitative contribution of increased deposition of
nitrogen to C sequestration by forest ecosystems of the
northern hemisphere is much debated. Estimates of the
effects of N deposition on C sequestration in forests range
from less than 0.1 up to 1.3 3 1015 g C year±1 (Townsend
et al., 1996; Nadelhoffer et al., 1999; White et al., 2000),
which corresponds to 5±90% of the putative terrestrial C
sink. As increased deposition of N affects both the amount
and the quality of litter, it is not clear how the carbon
dynamics of the soil respond to increased N inputs to
forests.
We have investigated the effect of soil type, N deposition,
and increased atmospheric CO2 on soil organic carbon and
attempted to determine the input of added CO2 into two
different forest soils experimentally. We established spruce±
beech model ecosystems in large open-top chambers on an
acidic loam and a calcareous sand. The ecosystems were
exposed to an atmosphere enriched in CO2 and increased N
inputs for 4 years. The CO2 used for the enrichment was
depleted in 13C relative to the ambient atmospheric CO2,
creating an opportunity to trace C inputs in the plant±soil
system.
Materials and methods
Experimental design
Model ecosystems with spruce (Picea abies Karst) and beech
(Fagus sylvatica L.) were established in 16 open-top chambers
in 1994. Each chamber was 3 m high, had a diameter of 3 m
and contained two soil compartments of 3 m2 surface area.
Each compartment was 1.5 m deep, served as a non-weighable
lysimeter, and had a 0.5-m thick layer of quartz sand with a
drainage outlet at the bottom (for details see Sonnleitner et al.,
2001). The treatments were as follows: ambient CO2
(370�mol mol±1) + low N deposition (0.7 g NH4NO3-N m±2
year±1); enriched CO2 (570�mol mol±1) + low N; ambient
CO2 + high N (7 g NH4NO3-N m±2 year±1); enriched CO2 +
high N. The treatments were arranged as a Latin square with
four replicates for each CO2 3 N treatment. Atmospheric CO2
enrichment started in January 1995. The incoming ®ltered air
was blown through textile tubes at a rate of 3000 m3 per hour
(Landolt et al., 1997). The chambers were irrigated with
electro-osmotically puri®ed tap water with ions added in
concentrations usually found in rain water. Nitrogen deposi-
tion was manipulated by adding NH4NO3 to the irrigation
water. Concentrations of nitrogen in the irrigation water were
0.88 mg N l±1 under low N deposition and 8.8 mg l±1 under high
N deposition. The irrigation water was applied once or twice a
week through nozzles which were kept just above the plant
canopy. To avoid external water and nutrient inputs by rain, a
transparent roof automatically closed the chambers at the onset
of rain.
The experiment was done on two soils transferred from
natural spruce±beech forest sites in Switzerland. One of the
soils was an acidic sandy loam, a Haplic Alisol, referred to
as `acidic loam'. The other soil was a calcareous loamy
sand, a Calcaric Fluvisol, quoted henceforth as `calcareous
sand'. None of the soils showed signs of hydromorphy.
Each soil was ®lled into one of the two separate
gravitational lysimeters (surface area 3 m2) of each open-
top chamber in spring 1994. The acidic loam was
transferred in two layers, a 0.4-m topsoil layer and a 0.6-
m subsoil layer, into the lysimeters. The calcareous sand,
which had no distinct soil horizons, was transferred in one
layer. Soil properties are given in Table 1. To minimize the
effects of soil disturbance, oats and barley were cropped in
the summer of 1994. In October 1994, model ecosystems
were established in each of the 32 lysimeters (16 per soil
type). Each model ecosystem was composed of eight beech
and eight spruce trees with ®ve understorey species (Carex
sylvatica, Geum urbanum, Ranunculus ®caria, Viola
sylvatica, and Hedera helix). Trees were grown from seeds
collected from selected provenances (beech) and from
clonal cuttings (spruce). The beech trees were 2±3 years
old at the time of planting. Details are given in Egli et al.
(1998).
620 F. Hagedorn et al.
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628
Sampling and analysis
After four growing seasons, in September 1998, the trees
were harvested. Leaves, stems, and roots were sampled and
dried separately. For isotopic analysis, one leaf was taken
from the top, middle, and lower crown section of each of
the eight beech trees. The 24 leaves taken per lysimeter
were pooled for analysis. In each lysimeter, three needles
per age class from the top and lower crown position were
collected from each of the eight spruce trees and pooled.
Fine roots were sampled at three locations per lysimeter
from 0 to 20 cm soil depth with a soil corer (3.5 cm in
diameter), separated into beech and spruce roots and
washed thoroughly. All plant samples were dried at 65°C
for 72 h and ground with a mixer mill (Retsch MM2000,
Haan, Germany). Aliquots of 4.0±4.5 mg were placed in tin
capsules for mass spectrometry analysis.
The soil was sampled prior to tree harvest from 0 to 10 cm
depth with a sharpened metal tube (n = 6 per lysimeter). The
soil from 10±25, 25±50, 50±60, and 90±100 cm depth was
taken with a soil corer (n = 2 per lysimeter). All soil samples
were dried at 60°C for 48 h immediately after sampling and
then stored in a climate chamber. For analysis, visible plant
particles were removed from the soil by hand. After sieving the
soil through a 2-mm sieve, we ground the samples of the bulk
soil ®nely in a ball-mill. The organic C associated with the
sand was separated by dispersing soil aliquots of 30 g
ultrasonically with 3 kJ energy and passing them through a
250-�m sieve as described by Amelung & Zech (1999). The
remaining sand fraction was dried at 60°C for 48 h and ground
with a ball-mill. To remove carbonates from the soil prior to C
analysis, aliquots of 15 mg soil were placed into silver
elemental analysis capsules, moistened, and exposed to vapour
from concentrated HCl in a desiccator for 12 h (David Harris,
personal communication, 1999). This procedure yielded the
same �13C values as the removal of carbonates by titration.
The C content and the �13C values of plant and soil samples
were determined with an automated elemental analyser±
continuous ¯ow isotope ratio mass spectrophotometer (EA-
1110, Carlo Erba, Italy, interfaced with a CONFLO II, Delta-S
Finnigan MAT, Bremen). Results of the C isotope analysis are
expressed in � units (½):
d13C � 1000
Rsample
Rstandard
ÿ 1
� �; �1�
where R = 13C/12C.
The �13C values were referenced to the Pee Dee Belemnite
(PDB) standard.
Calculations and statistical analysis
The fraction of soil C derived from plant input (fnew), the net
new soil C, was calculated with a simple mixing model
(Balesdent et al., 1988):
fnew � dsoil ÿ dsoil; native
dplant ÿ dsoil; native
� �; �2�
where �soil is the �13C value of the organic C in the soil at
harvest, �soil,native that of the organic C before the start of the
experiment (`native' SOC), and �plant is the �13C value of the
trees at harvest. As the biomass of foliage and ®ne roots at
harvest were approximately the same, and the contribution of
both root and ®ne root litter to soil organic C was not known,
we took the mean of the �13C values of leaves and ®ne roots
for �plant. The remaining proportion of native C in the soil is
equal to 1 ± fnew. The quantity of C in native C and the net
input of new C were calculated by multiplying the proportions
with soil C concentrations and dry masses. The mineralization
of native C was assessed by subtracting the amount of native
organic C at harvest from the value prior to the experiment.
Treatment effects were estimated with analyses of variance
(ANOVA). Following the experimental design, the main effects
of CO2 and N and their interaction were tested against the
`chamber mean squares', while the main effects of soil type
and its interactions with CO2, N, and CO2 3 N were tested
against the `lysimeter mean squares'. Effects of CO2 and N
were additionally tested in separate analyses for the acidic and
the calcareous soil.
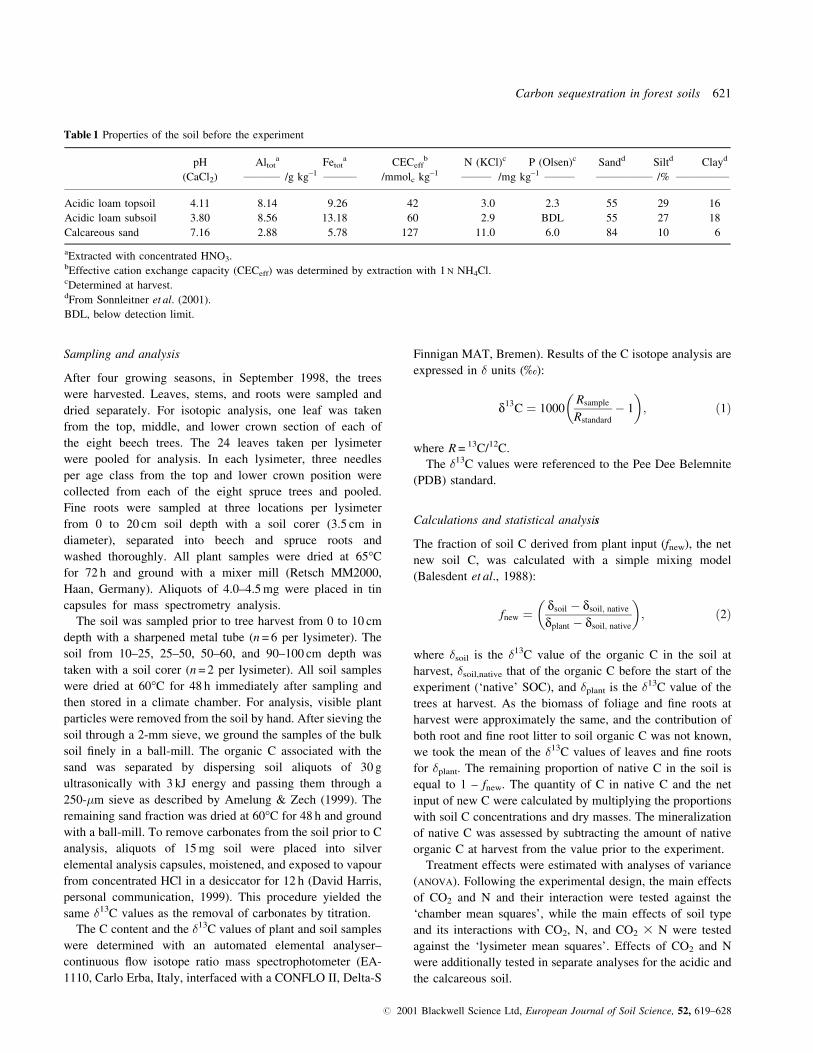
Table 1 Properties of the soil before the experiment
pH Altota Fetot
a CECeffb N (KCl)c P (Olsen)c Sandd Siltd Clayd
(CaCl2) ___________ /g kg±1 __________ /mmolc kg±1 _________ /mg kg±1 _________ _________________ /% ________________
Acidic loam topsoil 4.11 8.14 9.26 42 3.0 2.3 55 29 16
Acidic loam subsoil 3.80 8.56 13.18 60 2.9 BDL 55 27 18
Calcareous sand 7.16 2.88 5.78 127 11.0 6.0 84 10 6
aExtracted with concentrated HNO3.bEffective cation exchange capacity (CECeff) was determined by extraction with 1 N NH4Cl.cDetermined at harvest.dFrom Sonnleitner et al. (2001).
BDL, below detection limit.
Carbon sequestration in forest soils 621
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628

Results
Soil carbon
After the 4 years, the concentrations of organic C in the
calcareous sand decreased throughout all depths (see dotted
lines in Figure 1). This decrease reached 3 g C kg±1, which
corresponds to a loss of 23% of the initial carbon content. The
total loss of C in the whole soil compartment (100 cm depth) of
the calcareous sand ranged between 1300 and 2500 g C m±2. In
contrast, the changes of C concentrations in the acidic loam
were small (Figure 1). Contents of C in the acidic loam
decreased only below 25 cm depth. The total loss of C was
between 0 and 600 g C m±2.
Under large deposition of N, concentrations of C in the
uppermost 10 cm of the acidic loam were 9% greater than
when little N was added (P < 0.05, main effect pooled across
the CO2 treatments). At depths below 10 cm, N inputs had no
signi®cant effect on C concentrations (Figure 1). At the end of
the experiment, the C contents in the whole soil compartment
of the acidic loam were 230 g C m±2 greater in the model
ecosystem receiving 7 g N m±2 year±1 than in those receiving
0.7 g N m±2 year±1. In the calcareous sand, N deposition had no
effect on the C concentration of the topsoil, but concentrations
at 50 and 100 cm soil depth were about 14% (P < 0.05) greater
under the larger N inputs.
With increased atmospheric CO2, C concentrations in the
uppermost 10 cm of the acidic loam and of the calcareous sand
were 8% and 5% greater than at ambient CO2 (Figure 1), but
signi®cant only in the acidic soil (P < 0.05). Below 10 cm
depth, elevated CO2 had no signi®cant impact on the C
concentration of either of the soils. The increase in C content
of the uppermost 10 cm of the acidic loam induced by the CO2
accounted for 180 g C m±2 under low and for 70 g C m±2 under
high N deposition (Figure 4).
New and old soil carbon
The 4-year exposure of the trees to increased atmospheric CO2
with a �13C value of about ±16½ decreased the �13C values of
®ne roots and leaves by 10½ as compared with the biomass
produced under ambient CO2 (Table 2). The ratios of C to N of
®ne roots were increased by CO2 enrichment, but were
decreased by the greater N deposition (Table 2; P < 0.01 for
beech and spruce across both soils). Hagedorn et al. (2000)
noticed similar effects for C/N ratios of foliage.
The decline in 13C through the fumigation with CO2 was
also re¯ected in soil organic C. The �13C values of the C were
signi®cantly less under enriched CO2 than under ambient
concentration (Figure 2), indicating that new C was incorpo-
rated into the soils. The shift in �13C was highly signi®cant
down to a depth of 50 cm. In the acidic loam, this decline in
�13C was more pronounced than in the calcareous sand
(P < 0.01). As the C concentrations of the two soils were
similar before the experiment, the greater decrease in 13C
indicates that more new C was incorporated in the course of
the experiment. In contrast to soil type, N inputs did not
signi®cantly in¯uence the shift in �13C (Figure 2).
Under CO2 enrichment, the calculated fraction of new C in
total organic C in the uppermost 10 cm was 22 6 2% (6 SE) in
the acidic loam and 17 6 2% in the calcareous sand (Figure 4).
In the acidic loam, the net input of new C under elevated CO2
was 670 6 30 g C m±2 in the whole soil compartment with no
Figure 1 Concentrations of soil organic carbon in the acidic loam and in the calcareous sand of the spruce±beech model ecosystem after
4 experimental years. The dotted lines represent the carbon concentrations before the experiment. Low N represents a deposition of 0.7 g N m±2
year±1 and high N a deposition of 7 g N m±2 year±1. Means and standard errors of four replicates per treatment.
622 F. Hagedorn et al.
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628
effects of N additions (Figure 3). In the calcareous sand, the
net input of new C was 450 6 20 g C m±2 and 320 6 50 g C m±2
at low and at high N inputs (P < 0.05). The mineralization of
native C was signi®cantly greater in the calcareous sand than
in the acidic loam (P < 0.001). Increasing the N deposition did
not lead to an increase in net input of new C in the acidic loam,
but decreased the mineralization of native C. In the whole soil
compartment of the acidic loam, the decrease in mineralization
of native C accounted for 240 g C m±2 (Figure 3), but it was
signi®cant only at 0±10 cm depth (P < 0.05; Figure 4). In the
calcareous sand, N deposition did not signi®cantly in¯uence
the mineralization of native C.
Carbon associated with sand
After the 4 years, the concentrations of C in the coarse sand
fraction, representing the particulate organic matter, were
substantially larger than those of the native acidic loam (Table
3). In contrast, C concentrations in the particulate fraction of
the calcareous sand remained constant at enriched CO2 and
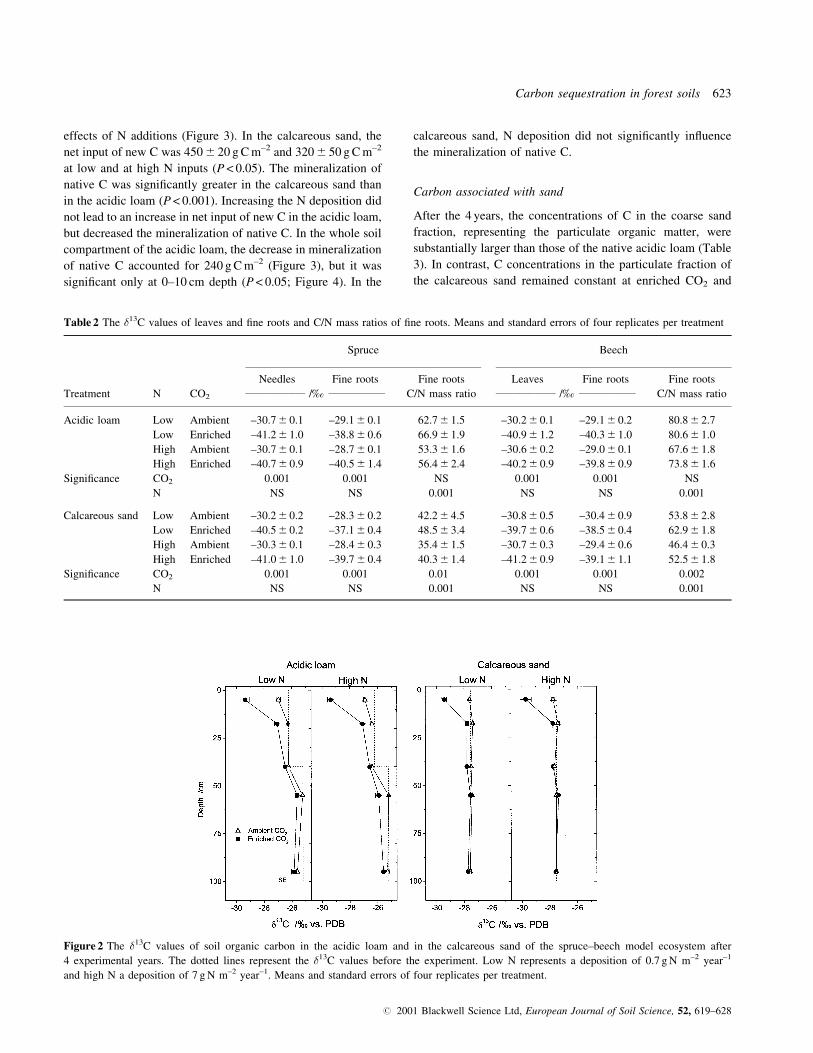
Table 2 The �13C values of leaves and ®ne roots and C/N mass ratios of ®ne roots. Means and standard errors of four replicates per treatment
Spruce Beech
Needles Fine roots Fine roots Leaves Fine roots Fine roots
Treatment N CO2__________________ /½ _________________ C/N mass ratio __________________ /½ _________________ C/N mass ratio
Acidic loam Low Ambient ±30.7 6 0.1 ±29.1 6 0.1 62.7 6 1.5 ±30.2 6 0.1 ±29.1 6 0.2 80.8 6 2.7
Low Enriched ±41.2 6 1.0 ±38.8 6 0.6 66.9 6 1.9 ±40.9 6 1.2 ±40.3 6 1.0 80.6 6 1.0
High Ambient ±30.7 6 0.1 ±28.7 6 0.1 53.3 6 1.6 ±30.6 6 0.2 ±29.0 6 0.1 67.6 6 1.8
High Enriched ±40.7 6 0.9 ±40.5 6 1.4 56.4 6 2.4 ±40.2 6 0.9 ±39.8 6 0.9 73.8 6 1.6
Signi®cance CO2 0.001 0.001 NS 0.001 0.001 NS
N NS NS 0.001 NS NS 0.001
Calcareous sand Low Ambient ±30.2 6 0.2 ±28.3 6 0.2 42.2 6 4.5 ±30.8 6 0.5 ±30.4 6 0.9 53.8 6 2.8
Low Enriched ±40.5 6 0.2 ±37.1 6 0.4 48.5 6 3.4 ±39.7 6 0.6 ±38.5 6 0.4 62.9 6 1.8
High Ambient ±30.3 6 0.1 ±28.4 6 0.3 35.4 6 1.5 ±30.7 6 0.3 ±29.4 6 0.6 46.4 6 0.3
High Enriched ±41.0 6 1.0 ±39.7 6 0.4 40.3 6 1.4 ±41.2 6 0.9 ±39.1 6 1.1 52.5 6 1.8
Signi®cance CO2 0.001 0.001 0.01 0.001 0.001 0.002
N NS NS 0.001 NS NS 0.001
Figure 2 The �13C values of soil organic carbon in the acidic loam and in the calcareous sand of the spruce±beech model ecosystem after
4 experimental years. The dotted lines represent the �13C values before the experiment. Low N represents a deposition of 0.7 g N m±2 year±1
and high N a deposition of 7 g N m±2 year±1. Means and standard errors of four replicates per treatment.
Carbon sequestration in forest soils 623
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628

decreased by about 25% at ambient CO2. The estimated
fraction of new C in the C associated with sand of the acidic
loam ranged from 50 to 75%. In the calcareous sand, these
proportions were 24 to 31%. Fertilization with N increased
concentrations of C in the acidic loam, but had no signi®cant
effects in the calcareous sand. Enrichment with CO2, in
contrast, did not signi®cantly in¯uence this C fraction in the
acidic loam, but increased it in the calcareous sand (Table 3).
Discussion
Effects of soil type
Our results show that the soil itself was the most important
factor in¯uencing the carbon dynamics of the soils and the
response of soil carbon to CO2 enrichment. In the 4 years of
the experiment, the contents of organic C decreased in the
calcareous sand, probably as a result of disturbance. This loss
in C indicates that changes in land use, such as deforestation or
clear-cutting, strongly affect storage of C in soils (Balesdent
et al., 1988; Davidson & Ackerman, 1993). Our results show
that loss of C due to disturbance depends upon soil type, since,
in contrast to the calcareous sand, the concentrations of C in
the acidic loam remained almost constant during the 4 years
(Figure 1). The net input of new C was about 75% greater in
the acidic loam than in the calcareous sand (Figure 3), despite
a 40% greater biomass of trees on the calcareous sand (Dieter
Spinnler, personal communication, 2000; for year 2, see Egli
et al., 1998). These ®ndings strongly suggest that C turned
over more rapidly in the calcareous sand than in the acidic
loam. This is supported by a greater mineralization of native C
in the calcareous sand (Figure 3) and by a 50% increase in
respiration measured by the nightly CO2 ef¯ux (Maurer et al.,
1999). Further, losses of nitrate and dissolved organic carbon
through drainage were more than 10 times greater in the
calcareous sand than in the acidic loam (Hagedorn et al.,
2000).
There are several reasons for the organic C to mineralize
more rapidly in the calcareous sand than in the acidic loam.
First, many microbes are more active at a neutral pH than in an
acidic milieu. For instance, there was a net nitri®cation in the
calcareous soil, but not in the acidic one (Hagedorn et al.,
2000). Second, the calcareous sand was less dense than the
acidic loam, indicating better aeration and more favourable
conditions for C mineralization. Third, the acidic loam
contained more clay than the calcareous sand (Table 1).
Clay interacts with organic compounds and protects organic C
from microbial decay. Fourth, the acidic loam contained more
Fe and Al than the calcareous sand (Table 1), suggesting that it
contained more oxides on which organic C could build stable
organo-mineral complexes (Martin & Haider, 1986).
Figure 3 The net input of new carbon and the mineralization of
native soil organic carbon of the whole soil compartment down to
100 cm depth in the acidic loam and in the calcareous sand of the
spruce±beech model ecosystem after 4 experimental years. Low N
represents a deposition of 0.7 g N m±2 year±1 and high N a
deposition of 7 g N m±2 year±1. Means and standard errors of four
replicates per treatment.
Figure 4 Total soil organic carbon as well as new and old carbon
under enriched atmospheric CO2 at 0±10 cm depth. The black lines
represent the organic carbon contents of the soils before the
experiment. New and old carbon could not be estimated for ambient
CO2, since the CO2 was not labelled with 13C. Means and standard
errors of four replicates per treatment.
624 F. Hagedorn et al.
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628

The differences between the two soil types were even more
pronounced in the C associated with the coarse sand. This C
consists mainly of particles and represents the most labile pool
of C in the soil (Balesdent et al., 1988; Christensen, 1992). The
C in the coarse sand fraction of the calcareous sand declined
by as much as 30% during the 4 years, which is considerably
more than the loss of C from the bulk soil. In contrast, in the
acidic loam, the amount of C associated with coarse sand
doubled during the experiment. Since the physico-chemical
stabilization of organic C in the coarse sand fraction is likely to
be equally small in both soil types, the contrasting response of
the soils strongly suggests that microorganisms found living
conditions unfavourable in the acidic loam, and so the organic
C was preserved more than in the calcareous sand.
Net input of new C into the forest soils
For the net input of new C, the soil type was more important
than the productivity of trees, as indicated by a larger net input
of new C in the less productive acidic loam than in the
calcareous sand (Figure 3). The net input of new C into the
acidic loam was 670 g C m±2, corresponding to 45% of the C
stored in the trees (above- + below-ground biomass). In the
calcareous sand, the net input of new C was only about
380 g C m±2, which was equal to 18% of the total C in the trees.
The net input of new C per unit of plant biomass in the acidic
loam was greater than that under agriculture. In cropping
systems with sorghum and soybean, the net input of new C
corresponded to 15 and 22% of the biomass produced
(calculated from Torbert et al., 1997). In a pasture study, this
proportion was about 15% (calculated from Hebeisen et al.,
1997 and van Kessel et al., 2000). One explanation for the
larger C input per unit biomass in our acidic model forest is a
slower decomposition of the new C due either to the quality of
the litter or to unfavourable conditions for decomposers as
discussed above. Moreover, the litter input was probably larger
in our forest than in agriculture, where a part of the biomass
produced is harvested and removed from the land each year.
Another reason could be a greater investment of trees into
below-ground biomass. In our acidic forest, root biomass was
about ®ve times greater than recorded by Hebeisen et al.
(1997) in the pasture experiment. These ®ndings suggest that
acidic forest soils can sequester more C than soils under
agriculture.
A large proportion of the net input of new C was found in
the organic matter associated with the coarse sand fraction
(Table 3). Although the coarse sand fraction accounted for
only 7% of the total soil by volume in the acidic loam, up to
40% of the total new C input occurred in it. Because the
turnover of the C associated with the coarse sand is usually
rather rapid, with half-lifes less than 10 years (Martin et al.,
1990), the large proportion of new C in this fraction strongly
suggests that only a small portion of it becomes stable organic
matter. However, only long-term experiments can clarify how
much of new C inputs enters passive pools of organic C in the
soil.
Effects of CO2 enrichment
The impact of an atmosphere enriched in CO2 on the dynamics
of C in the soil was small in comparison with the large
differences between the two soil types and the large losses of C
Table 3 Coarse sand fraction: content in the total soil volume, soil organic carbon concentrations, �13C values, and the fraction of new C in
soil organic carbon (SOC). Means and standard errors of four replicates per treatment
Coarse sand SOC �13C New Ca
Treatment N CO2____________________ /% ____________________ /½ /%
Acidic loam Before experiment 6.9 1.53 ±27.7
Low Ambient 7.0 6 0.5 3.03 6 0.32 ±29.0 6 0.2 ND
Low Enriched 7.0 6 0.2 3.06 6 0.19 ±35.2 6 0.6 59 6 4
High Ambient 6.9 6 0.1 3.49 6 0.49 ±28.9 6 0.1 ND
(After 4 years) High Enriched 7.0 6 0.1 3.57 6 0.07 ±35.8 6 0.5 64 6 4
Signi®cance CO2 NS NS 0.001 ±
N NS 0.03 NS NS
Calcareous sand Before experiment 23.3 1.07 ±27.3
Low Ambient 22.7 6 0.3 0.76 6 0.08 ±27.1 6 0.2 ND
Low Enriched 23.2 6 0.9 1.03 6 0.11 ±30.1 6 0.5 24 6 4
High Ambient 22.5 6 0.8 0.84 6 0.07 ±27.6 6 0.1 ND
(After 4 years) High Enriched 21.9 6 0.4 1.05 6 0.09 ±31.5 6 0.9 31 6 6
Signi®cance CO2 NS 0.03 NS ±
N NS NS NS NS
aCalculated with the mean �13C value of ®ne roots and leaves.
ND, not determined.
NS, not signi®cant.
Carbon sequestration in forest soils 625
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628

from the calcareous sand during the experiment (Figure 4).
The small increase in C concentrations of the bulk soil in
response to CO2 enrichment accorded with the marginal and
insigni®cant increases of C contents under enriched CO2 as
reported by Leavitt et al. (1994), Hungate et al. (1996) and van
Kessel et al. (2000). The increase in organic C induced by CO2
in the uppermost 10 cm of the soil ranged between 50 g C m±2
in the calcareous sand and 180 g C m±2 in the acidic loam
(Figure 4). This is similar to the observed increases of up to
400 g C m±2 after 4 years in the pasture study of van Kessel
et al. (2000) and in a tallgrass prairie exposed to increased CO2
for 8 years as found by Williams et al. (2000). In contrast to
most other studies, the CO2-induced increase in soil C was
signi®cant in the acidic loam of our study. This was probably
because there was little variation in the organic carbon content
of the soil in this study (CV < 15%, Figure 4) as a result of the
homogenization prior to the experiment and the large number
of replicates ± eight per CO2 level and soil type.
The small increase in total organic C through CO2
enrichment can be attributed either to an increased input of
new C or to a decreased mineralization of native C under
enriched CO2. Because CO2 of ambient air was not labelled
with 13C, we could not distinguish between these two
mechanisms. However, ®ne roots were stimulated through
CO2 enrichment by about 20% (Dieter Spinnler, personal
communication, 2000), suggesting that a part of the CO2-
induced increase in soil C was caused by a greater net input of
new C into the soil. This is consistent with experiments in
grasslands that were previously cropped with C4 plants where
increased atmospheric CO2 led to an increase in new C by 37
and 50%, respectively, in the soil (Nitschelm et al., 1997;
Loiseau & Soussana, 1999).
Assuming that soils of temperate forests respond similarly to
increased CO2 as the two soils in this experiment, and given
that the world's temperate forests cover about 6 3 1012 m2
(Ajtay et al., 1979), we can estimate that the gain in soil
organic C by the experimental increase of 200�mol CO2
mol±1 amounts to 0.1±0.27 3 1015 g C year±1. This
corresponds to 7±20% of the yearly missing C sink (1.4 3
1015 g C year±1; Schimel, 1995). However, as the annual rise of
CO2 concentrations of the atmosphere is about 1.4�mol CO2
mol±1 year±1 (Houghton et al., 1990), the actual contribution of
a CO2-induced increase in organic C in the soil is probably at
least one order of magnitude less.
Effects of N deposition
The response of C dynamics to inputs of N depended upon the
type of soil. In the calcareous sand with much available N
(Hagedorn et al., 2000), increased deposition of N did not
affect the concentrations of organic C, but it caused a decrease
of new C (Figures 3 and 4). This is consistent with a smaller
amount of root biomass at high than at low N deposition under
CO2 enrichment in the calcareous sand. In the acidic loam with
little available N, concentrations of organic C in the uppermost
10 cm were increased by increased deposition of N (Figure 4).
Since the increase of N inputs also stimulated tree growth on
this soil (root biomass: +25%; Dieter Spinnler, personal
communication, 2000), one would expect that the increase in
C resulted from enhanced C input into the soil. However, the
net input of new C was only slightly stimulated by increased N
deposition in the acidic loam (Figures 3 and 4). The N-induced
increase in C was caused mainly by a suppressed mineraliza-
tion of native C. Our results accord with the results from other
experiments; they show that additions of mineral N stimulate
the decomposition of plant residues (i.e. `new' C), but reduce
decomposition of recalcitrant organic matter (Fog, 1988;
Green et al., 1995). Magill & Aber (1998) measured an
accumulation of lignin under large deposition of N during a
long-term experiment, and they explained their ®nding as the
suppression of the production of ligninolytic enzyme by soil
microbes, particularly fungi. Global carbon models including
increased N deposition neglect potentially retarded decay of
soil C (Townsend et al., 1996; White et al., 2000). Our results
give this as about 240 g C m±2 during 4 years in the acidic
loam, which is consistent with the values estimated by Magill
& Aber (1998). This corresponds to 50% of the N fertilization
effect found for tree growth, and it is in the same order of
magnitude as the annual leaf fall in mature forests (Vogt et al.,
1986). Since soil organic C represents the pool of carbon with
the longest turnover in ecosystems, this effect of added N
could be quantitatively even more important with respect to
the global carbon cycle.
Conclusions
Our results show that soils play a key role in the sequestration
of carbon. The net input of new C was larger in the acidic loam
than in the calcareous sand, despite a considerably larger
biomass production in the fertile calcareous sand. In
comparison with agricultural systems, the net input of new C
per unit of biomass produced was large in the acidic loam,
suggesting that acidic forest soils have a large potential to store
C. The pronounced differences between the two soil types in C
sequestration indicate that properties such as soil pH, texture,
and mineralogy control the stabilization of C in the soil. The
mechanisms by which organic matter is stabilized in the soil
need further investigation, because they determine whether the
carbon will be respired to the atmosphere in a fairly short time
or remain in the soil and become `long-term' organic C. Our
®ndings that a large proportion of new C was in the coarse
sand fraction of the soil, the most labile pool of organic C,
suggest that only a small portion of the new C input becomes
stable. The different dynamics of C in the two soil types have
implications for estimates and models of global carbon
budgets, which currently neglect soil properties and concen-
trate mainly on the effects of temperature and litter input.
626 F. Hagedorn et al.
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628

The large losses of C from the calcareous sand during the
4 years indicate that changes of land use can severely decrease
the carbon storage in the soil. In comparison with the large
differences between the two soil types, the effects of
increasing the CO2 and the increased N deposition on the soil
carbon were small. Enrichment in CO2 increased the contents
of C signi®cantly only in the acidic loam at 0±10 cm depth.
The small increase in C through increased CO2 compared with
the missing carbon sink suggests that an accelerated storage of
C in soils makes a minor contribution to C sequestration.
Increased deposition of N increased concentrations of organic
C in the uppermost acidic loam signi®cantly. The major reason
was a suppressed mineralization of native C as indicated by a
negligible in¯uence of N inputs on the fraction of new C.
According to our results, the signi®cant reduction in miner-
alization of native C due to increased deposition of N was
quantitatively important and may lead to underestimates of the
effect of nitrogen fertilization on C sequestration.
Acknowledgements
This experiment was designed by C. KoÈrner, J.B. Bucher, and
W. Landolt as part of the Swiss contribution to COST 614
under the coordination of C. Brunold. The CO2 enrichment
was maintained by U. Bleuler and W. Landolt. We are grateful
to M. Bundt for helpful discussions and carefully reading the
script. Funding was provided by the Board of the Swiss
Federal Institute of Technology and by the Swiss National
Science Foundation.
References
Ajtay, G.L., Ketner, P. & Duvigneaud, P. 1979. Terrestrial primary
production and phytomass. In: The Global Carbon Cycle (Scope
Report 13) (eds B. Bolin, E.T. Degens, S. Kempe & P. Ketner), pp.
129±181. John Wiley & Sons, Chichester.
Amelung, W. & Zech, W. 1999. Minimisation of organic matter
disruption during particle-size fractionation of grasslands epipe-
dons. Geoderma, 92, 73±85.
Balesdent, J., Wagner, G.H. & Mariotti, A. 1988. Soil organic matter
turnover in long-term ®eld experiments as revealed by carbon-13
natural abundance. Soil Science Society of America Journal, 52,
118±124.
Canadell, J.G., Pitelka, L.F. & Ingram, J.S.I. 1996. The effects of
elevated [CO2] on plant±soil carbon below-ground: a summary and
synthesis. Plant and Soil, 187, 391±400.
Christensen, B.T. 1992. Physical fractionation of soil and organic
matter in primary particle size and density separates. Advances in
Soil Science, 20, 1±90.
Ciais, P., Tans, P.P., Trolier, M., White, J.W.C. & Francey, R.J. 1995.
A large northern hemisphere CO2 sink indicated by the 13C/12C
ratio of atmospheric CO2. Science, 269, 1098±1102.
Davidson, E.A. & Ackerman, I.L. 1993. Changes in soil carbon
inventories following cultivation of previously untilled soils.
Biogeochemistry, 20, 161±193.
Dixon, R.K., Brown, S., Houghton, R.A., Solomon, A.M., Trexler,
M.C. & Wisniewski, J. 1994. Carbon pools and ¯ux of global forest
ecosystems. Science, 263, 185±190.
Egli, P., Maurer, S., GuÈnthardt-Goerg, M. & KoÈrner, C. 1998. Effects
of elevated CO2 and soil quality on leaf gas exchange and above-
ground growth in beech±spruce model ecosystems. New
Phytologist, 140, 185±196.
Fog, K. 1988. The effect of added nitrogen on the rate of
decomposition of organic matter. Biological Reviews, 63, 433±462.
Green, C.J., Blackmer, A.M. & Horton, R. 1995. Nitrogen effects on
conservation of carbon during corn residue decomposition in soil.
Soil Science Society of America Journal, 59, 453±459.
Guggenberger, G., Christensen, B.T. & Zech, W. 1994. Land-use
effects on the composition of organic matter in particle-size
separates of soils: I. Lignin and carbohydrate signature. European
Journal of Soil Science, 45, 449±458.
Hagedorn, F., Bucher, J.B., Tarjan, D., Rusert, P. & Bucher-Wallin, I.
2000. Responses of N ¯uxes and pools to elevated atmospheric CO2
in model forest ecosystems with acidic and calcareous soils. Plant
and Soil, 224, 273±286.
Hebeisen, T., LuÈscher, A. & Zanetti, S. 1997. Growth responses of
Trifolium repens L. and Lolium perenne L. as monoculture and bi-
species mixture to free air CO2 enrichment and management.
Global Change Biology, 3, 149±160.
Houghton, J.T., Jenkins, G.J. & Ephraums, J.J. 1990. Climate Change:
The IPCC Scienti®c Assessment. Cambridge University Press,
Cambridge.
Hungate, B.A., Jackson, R.B., Field, C.B. & Chapin, F.S. 1996.
Detecting changes in soil carbon in CO2 enrichment experiments.
Plant and Soil, 187, 135±145.
van Kessel, C., Nitschelm, J., Horwath, W.R., Harris, D., Walley, F.,
LuÈscher, A. & Hartwig, U. 2000. Carbon-13 input and turn-over in
a pasture soil exposed to long-term elevated atmospheric CO2.
Global Change Biology, 6, 123±135.
Landolt, W., Bucher, J.B., Schulin, R., KoÈrner, C. & Brunold, C.
1997. Effects of elevated CO2 concentration and N deposition on
spruce±beech model ecosystem. In: Impacts of Global Change on
Tree Physiology and Forest Ecosystems (eds G.M.J. Mohren, K.
Kramer & S. SabateÂ), pp. 317±324. Kluwer Academic Publishers,
Dordrecht.
Leavitt, S.W., Paul, E.A., Kimball, B.A., Hendrey, G.R., Mauney,
J.R., Rauschkolb, R. et al. 1994. Carbon isotope dynamics of free-
air CO2 enriched cotton and soils. Agricultural and Forest
Meteorology, 70, 87±101.
Loiseau, P. & Soussana, J.F. 1999. Elevated [CO2], temperature
increase and N supply effects on the turnover of below-ground
carbon in a temperate grassland ecosystem. Plant and Soil, 210,
233±247.
Magill, A.H. & Aber, J.D. 1998. Long-term effects of experimental
nitrogen additions on foliar litter decay and humus formation in
forest ecosystems. Plant and Soil, 203, 301±311.
Martin, A., Mariotti, A., Balesdent, J., Lavelle, P. & Vuattoux, R.
1990. Estimate of organic matter turnover rate in a savanna soil by13C natural abundance measurements. Soil Biology and
Biochemistry, 22, 517±523.
Martin, J.P. & Haider, K. 1986. In¯uence of mineral colloids on
turnover rates of soil organic carbon. In: Interactions of Soil
Minerals with Natural Organics and Microbes (eds P.M. Huang &
Carbon sequestration in forest soils 627
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628

M. Schnitzer), pp. 283±304. Soil Science Society of America,
Madison, WI.
Maurer, S., Egli, P., Spinnler, D. & KoÈrner, C. 1999. Carbon and
water ¯uxes in beech±spruce model ecosystems in response to
long-term exposure to atmospheric CO2 enrichment and increased
nitrogen deposition. Functional Ecology, 13, 748±755.
Nadelhoffer, K.J., Emmett, B.A., Gundersen, P., Kjùnnas, O.J.,
Koopmans, C.J., Schleppi, P. et al. 1999. Nitrogen deposition
makes a minor contribution to carbon sequestration in temperate
forests. Nature, 398, 145±148.
Nitschelm, J.J., LuÈscher, A., Hartwig, U.A. & van Kessel, C. 1997.
Using stable isotopes to determine soil carbon input differences
under ambient and elevated atmospheric CO2 conditions. Global
Change Biology, 3, 411±416.
Paterson, E., Hall, J.M., Rattray, E.A.S., Grif®ths, B.S., Ritz, K. &
Killham, K. 1997. Effect of elevated CO2 on rhizosphere carbon
¯ow and soil microbial processes. Global Change Biology, 3, 363±
377.
Schimel, D.S. 1995. Terrestrial ecosystems and the carbon cycle.
Global Change Biology, 1, 77±91.
Schlesinger, W.H. 1990. Evidence from chronosequence studies for a
low carbon-storage potential of soils. Nature, 348, 232±234.
Sonnleitner, M.A., GuÈnthardt-Goerg, M.S., Bucher-Wallin, I.,
Attinger, W., Reis, S. & Schulin, R. 2001. In¯uence of soil type
on the effects of elevated atmospheric CO2 and N deposition on
water balances and growth of a young spruce and beech forest.
Water, Air, and Soil Pollution, 126, 271±290.
Speicker, H., MielikaÈinen, K., KoÈhl, M. & Skovsgaard, J.P. 1996.
Growth Trends in European Forests. Springer, Berlin.
Torbert, H.A., Rogers, H.H., Prior, S.A., Schlesinger, W.H. & Runion,
G.B. 1997. Effects of elevated atmospheric CO2 in agro-ecosystems
on soil carbon storage. Global Change Biology, 3, 513±521.
Townsend, A.R., Braswell, B.H., Holland, E.A. & Penner, J.E. 1996.
Spatial and temporal patterns in potential terrestrial carbon storage
due to deposition of fossil fuel nitrogen. Ecological Applications, 6,
806±814.
Vogt, K.A., Grier, C.C. & Vogt, D.J. 1986. Production, turnover, and
nutrient dynamics of above- and belowground detritus of world
forest. In: Advances in Ecological Research 15 (eds A. Macfadyen
& E.D. Ford), pp. 303±377. Academic Press, London.
White, A., Cannell, M.G.R. & Friend, A.D. 2000. The high-latitude
terrestrial carbon sink: a model analysis. Global Change Biology, 6,
227±245.
Williams, M.A., Rice, C.W. & Owensby, C.E. 2000. Carbon dynamics
and microbial activity in tallgrass prairie exposed to elevated CO2
for 8 years. Plant and Soil, 227, 127±137.
628 F. Hagedorn et al.
# 2001 Blackwell Science Ltd, European Journal of Soil Science, 52, 619±628