carbon and nitrogen assimilation in deep subsea … · carbon and nitrogen assimilation in deep...
TRANSCRIPT

Carbon and nitrogen assimilation in deep subseafloormicrobial cellsYuki Moronoa,1, Takeshi Teradab, Manabu Nishizawac, Motoo Itoa, François Hilliond, Naoto Takahatae, Yuji Sanoe,and Fumio Inagakia,1
aKochi Institute for Core Sample Research, Japan Agency for Earth-Marine Science and Technology (JAMSTEC), Nankoku, Kochi 783-8502, Japan; bDepartmentof Ocean Drilling Science Technical Support, Marine Works Japan LTD, Yokohama 236-0042, Japan; cPrecambrian Ecosystem Laboratory, Japan Agency forEarth-Marine Science and Technology (JAMSTEC), Yokosuka 237-0061, Japan; dCAMECA Simplified Anonymous Society, 92622 Cedex, Gennevilliers, France;and eAtmosphere and Ocean Research Institute, University of Tokyo, Kashiwa 277-8564, Japan
Edited by David M. Karl, University of Hawaii, Honolulu, HI, and approved September 9, 2011 (received for review May 16, 2011)
Remarkable numbers of microbial cells have been observed inglobal shallow to deep subseafloor sediments. Accumulating evi-dence indicates that deep and ancient sediments harbor livingmicrobial life, where the flux of nutrients and energy are extremelylow. However, their physiology and energy requirements remainlargely unknown. We used stable isotope tracer incubation andnanometer-scale secondary ion MS to investigate the dynamics ofcarbon and nitrogen assimilation activities in individual microbialcells from 219-m-deep lower Pleistocene (460,000 y old) sedimentsfrom the northwestern Pacific off the Shimokita Peninsula of Japan.Sediment samples were incubated in vitro with 13C- and/or 15N-labeled glucose, pyruvate, acetate, bicarbonate, methane, ammo-nium, and amino acids. Significant incorporation of 13C and/or 15Nand growth occurred in response to glucose, pyruvate, and aminoacids (∼76% of total cells), whereas acetate and bicarbonate wereincorporated without fostering growth. Among those substrates, amaximum substrate assimilation rate was observed at 67 × 10−18
mol/cell per d with bicarbonate. Neither carbon assimilation norgrowth was evident in response to methane. The atomic ratiosbetween nitrogen incorporated from ammonium and the total cel-lular nitrogen consistently exceeded the ratios of carbon, suggest-ing that subseafloor microbes preferentially require nitrogenassimilation for the recovery in vitro. Our results showed that themost deeply buried subseafloor sedimentary microbes maintainpotentials for metabolic activities and that growth is generally lim-ited by energy but not by the availability of C and N compounds.
subseafloor life | metabolic activity | carbon and nitrogen fixation |marine sedimentary habitat
Numerous studies of microbial ecology have shown that thedynamics of microbial activities in any ecosystem on this
planet largely depend on the availability of energy and nutrientsubstrates in the environment (reviewed by Morita in ref. 1). Mi-crobial life has been identified in various natural environments,including deep and ancient subseafloor sediments down to 1,627 mbelow the seafloor (mbsf) (2). Estimates of naturally occurringmicrobial populations have suggested that a significant fraction ofthe total living biomass on Earth is present in the subseafloorsedimentary biosphere (3–5). Chemical profiles in pore water fromsediment drill cores have suggested that the metabolic activities ofsubseafloor life are generally extremely low (6–8) because of se-vere limitations in the availability of electron donors and acceptorsfor cell respiration and growth (9). Radioactive tracer incubationstudies have consistently shown that the theoretical mean gener-ation time of subseafloor microbial cells is extremely long, rangingfrom a few to thousands of years (9, 10). The net activity of thesubseafloor sedimentary microbial ecosystem plays an ecologicallyimportant role in biogeochemical cycles, such as the degradationof buried organic matter and methane production.Other studies of subseafloor life in sediment on the continental
margins have shown that the microbial biomass positively corre-lated with the organic concentration in sediment (5, 8). This
finding indicates that sedimentary microbial ecosystems on mostcontinental margins consist mainly of organic-fueled heterotrophswith relatively minor autotrophic components (e.g., methanogens)(11). The composition of carbon isotopes (δ13C) in both extractedintact polar lipids and cellular bodies assessed by secondary ionMS (SIMS) from core sediments off the coast of Peru revealedconstant values similar to the δ13C of buried organic matter, evenat the sulfate–methane transition zones where 13C-depleted bio-genic methane is expected to mediate microbial carbon metabo-lism through anaerobic oxidation of methane (12). Molecularecological studies based on PCR-amplified 16S rRNA (and itsDNA) have revealed the presence of diverse archaea and bacteriafrom organic-rich subseafloor sediments, of which most sequencesare phylogenetically distinct from physiologically known isolates(13–16), and their community compositions are stratified withdepth and redox interfaces such as sulfate–methane transitionzones (17, 18). Metagenomic analysis of the Peru margin sedimentrevealed that over 80% of the genetic assemblages were func-tionally unknown (19), whereas some key functional genes andactivities related to biogeochemical carbon cycles from variouslocations on continental margins have been reported (14, 20–22).Despite the widespread occurrence of significant populations of
microbial communities in the subseafloor sedimentary environ-ment, the physiological nature and nutrient energy requirementsfor the long-term maintenance of essential life functions haveseldom been tested (23, 24). For example, whether the low avail-ability of nutrient and energy substrates constrains the physiolog-ical status and growth potential of deeply buried microbial cells isessentially unknown. More generally, differentiating whetherthese subseafloor microbes are alive, growing, simply surviving,dormant, or dead fossils is important. To address these funda-mental questions regarding the ecological physiology of subsea-floor sedimentary microbial life, we studied the carbon andnitrogen assimilation in a deeply buried microbial community atthe single-cell level using stable isotope (13C and/or 15N) tracers,image-based cell enumeration, catalyzed reporter deposition(CARD) -FISH, and nanometer-scale SIMS (NanoSIMS).
Results and DiscussionSampling Location and Geochemical Settings. The sample studiedherein represents typical organic-rich deep (sediment depth of
Author contributions: Y.M. and F.I. designed research; Y.M., T.T., and F.I. performedresearch; M.N., M.I., F.H., N.T., and Y.S. contributed new reagents/analytic tools; Y.M.,T.T., and M.N. analyzed data; and Y.M. and F.I. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.
See Commentary on page 18193.1To whom correspondence may be addressed. E-mail: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1107763108/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1107763108 PNAS | November 8, 2011 | vol. 108 | no. 45 | 18295–18300
ENVIRONMEN
TAL
SCIENCE
SSE
ECO
MMEN
TARY
219 m) and ancient (ca. 460,000 y old) sedimentary habitats inthe northwestern Pacific Ocean (25). The drilling vessel (DV)Chikyu launched the shakedown cruise CK06-06 at Site C9001 inthe northwestern Pacific Ocean (41°10′38.28″ N, 142°12′04.89″E) off the Shimokita Peninsula of Japan in 2006 (Fig. S1). Thecored sediment was diatom-rich hemipelagic clay containinga large amount of methane and organic carbon (1.2% weight oftotal organic carbon). The pore water from the examined sedi-ment core at 219 mbsf was pH 7.8, and it contained dissolvedinorganic carbon (DIC), acetate, and ammonium concentrationsof ∼104, 0.23, and 15 mM, respectively. Neither sulfate nor ni-trate was detected in the pore water. Detailed geochemicalprofiles of the squeezed pore waters have been described else-where (26). Sediment slurry was aseptically prepared in the mi-crobiology laboratory on DV Chikyu and portioned underanaerobic conditions. Subsamples were collected immediatelyafter core recovery. During subsample processing on board, thecore was placed in a laminar flow clean cabinet, and its centerwas sampled (50 cm3) using a 50-mL tip-cut sterilized syringe.Two volumes of sulfate-free, anaerobic artificial seawater (pH7.8) were added to prepare a slurry sample under an atmosphereof 0.22-μm filtered nitrogen.
13C and 15N Incorporation Analyzed Using NanoSIMS. Aliquots wereseparately supplemented with either one 13C-labeled carbonsubstrate (1 mM glucose, 1 mM acetate, 1 mM pyruvate, and1 mM bicarbonate or 3 atm of methane in headspace) and a15N-labeled nitrogen source (0.1 mM ammonium) or a 13C- and15N-labeled mixture of 20 aa (1 mM in total). After a 65-d sta-tionary incubation at 10 °C in the dark, the total cell abundanceand number of cells incorporating carbon (13C) and nitrogen(15N) were estimated by a fluorescent microscopic image-basedcell enumeration technique (27) and NanoSIMS.
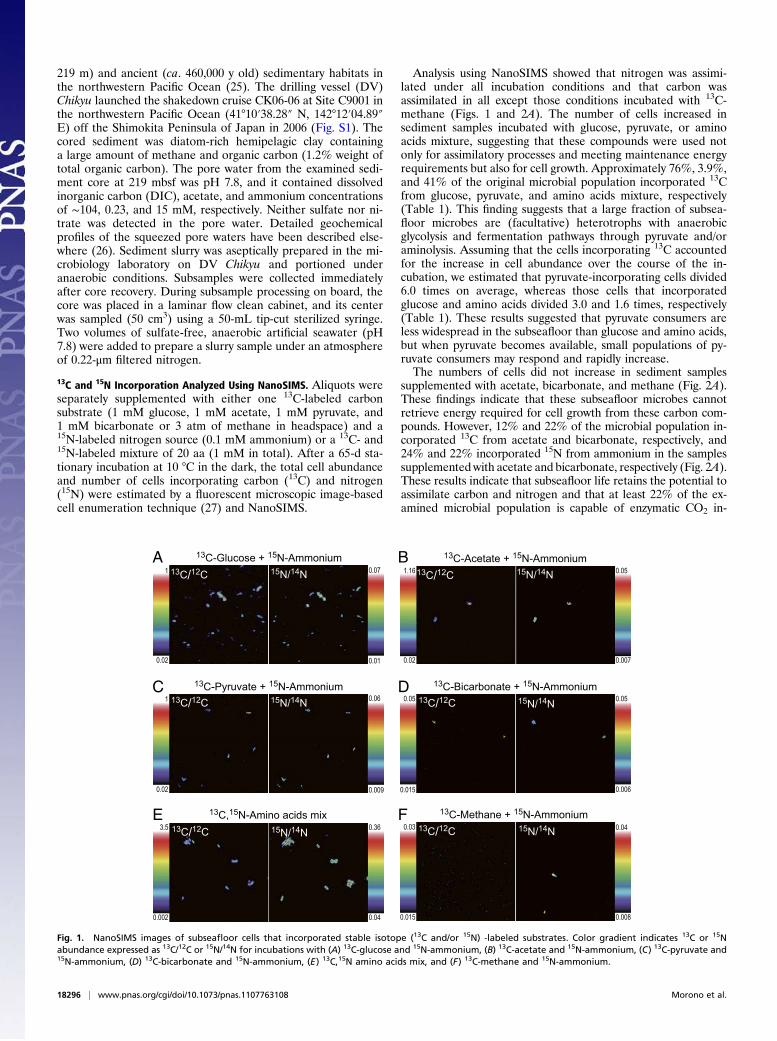
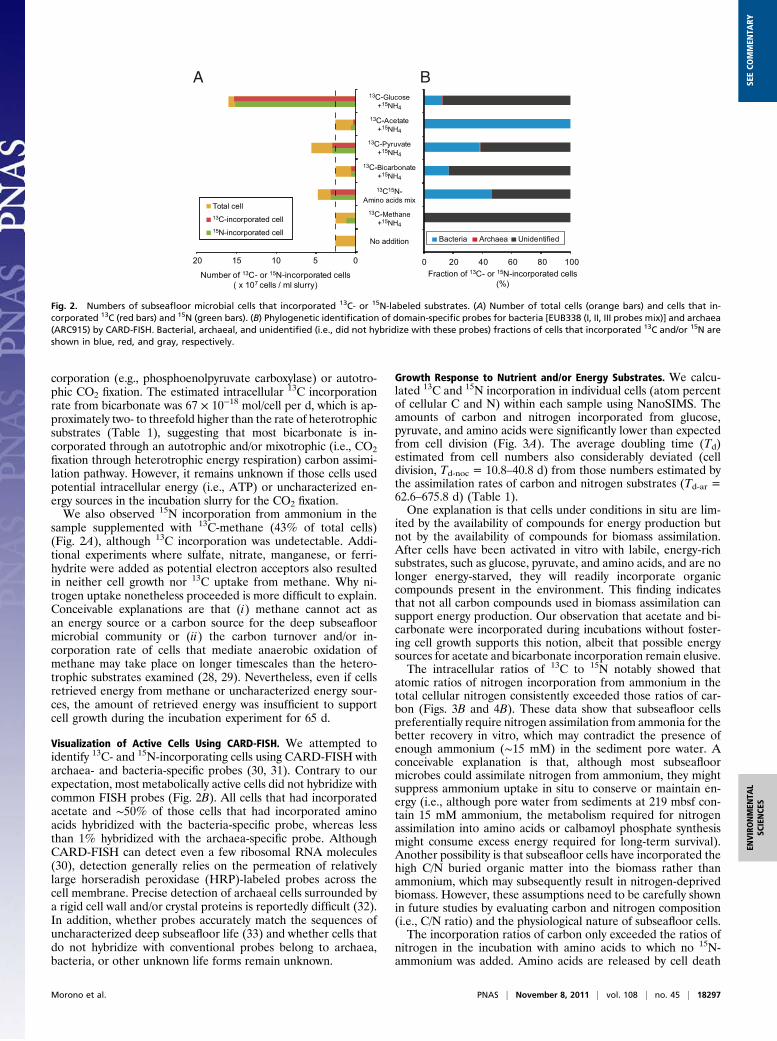
Analysis using NanoSIMS showed that nitrogen was assimi-lated under all incubation conditions and that carbon wasassimilated in all except those conditions incubated with 13C-methane (Figs. 1 and 2A). The number of cells increased insediment samples incubated with glucose, pyruvate, or aminoacids mixture, suggesting that these compounds were used notonly for assimilatory processes and meeting maintenance energyrequirements but also for cell growth. Approximately 76%, 3.9%,and 41% of the original microbial population incorporated 13Cfrom glucose, pyruvate, and amino acids mixture, respectively(Table 1). This finding suggests that a large fraction of subsea-floor microbes are (facultative) heterotrophs with anaerobicglycolysis and fermentation pathways through pyruvate and/oraminolysis. Assuming that the cells incorporating 13C accountedfor the increase in cell abundance over the course of the in-cubation, we estimated that pyruvate-incorporating cells divided6.0 times on average, whereas those cells that incorporatedglucose and amino acids divided 3.0 and 1.6 times, respectively(Table 1). These results suggested that pyruvate consumers areless widespread in the subseafloor than glucose and amino acids,but when pyruvate becomes available, small populations of py-ruvate consumers may respond and rapidly increase.The numbers of cells did not increase in sediment samples
supplemented with acetate, bicarbonate, and methane (Fig. 2A).These findings indicate that these subseafloor microbes cannotretrieve energy required for cell growth from these carbon com-pounds. However, 12% and 22% of the microbial population in-corporated 13C from acetate and bicarbonate, respectively, and24% and 22% incorporated 15N from ammonium in the samplessupplemented with acetate and bicarbonate, respectively (Fig. 2A).These results indicate that subseafloor life retains the potential toassimilate carbon and nitrogen and that at least 22% of the ex-amined microbial population is capable of enzymatic CO2 in-
13C-Glucose + 15N-Ammonium 13C-Acetate + 15N-Ammonium
0.02
1.16
0.007
0.0513C/12C 15N/14N13C/12C 15N/14N
0.01
0.07
0.02
1
13C-Pyruvate + 15N-Ammonium 13C-Bicarbonate + 15N-Ammonium
0.006
0.05
0.015
0.05 13C/12C 15N/14N
0.009
0.0613C/12C 15N/14N
0.02
1
13C,15N-Amino acids mix 13C-Methane + 15N-Ammonium
0.04
0.36
0.002
3.5 13C/12C 15N/14N
0.008
0.04
0.015
0.03 13C/12C 15N/14N
BA
C D
FE
Fig. 1. NanoSIMS images of subseafloor cells that incorporated stable isotope (13C and/or 15N) -labeled substrates. Color gradient indicates 13C or 15Nabundance expressed as 13C/12C or 15N/14N for incubations with (A) 13C-glucose and 15N-ammonium, (B) 13C-acetate and 15N-ammonium, (C) 13C-pyruvate and15N-ammonium, (D) 13C-bicarbonate and 15N-ammonium, (E) 13C,15N amino acids mix, and (F) 13C-methane and 15N-ammonium.
18296 | www.pnas.org/cgi/doi/10.1073/pnas.1107763108 Morono et al.
corporation (e.g., phosphoenolpyruvate carboxylase) or autotro-phic CO2 fixation. The estimated intracellular 13C incorporationrate from bicarbonate was 67 × 10−18 mol/cell per d, which is ap-proximately two- to threefold higher than the rate of heterotrophicsubstrates (Table 1), suggesting that most bicarbonate is in-corporated through an autotrophic and/or mixotrophic (i.e., CO2fixation through heterotrophic energy respiration) carbon assimi-lation pathway. However, it remains unknown if those cells usedpotential intracellular energy (i.e., ATP) or uncharacterized en-ergy sources in the incubation slurry for the CO2 fixation.We also observed 15N incorporation from ammonium in the
sample supplemented with 13C-methane (43% of total cells)(Fig. 2A), although 13C incorporation was undetectable. Addi-tional experiments where sulfate, nitrate, manganese, or ferri-hydrite were added as potential electron acceptors also resultedin neither cell growth nor 13C uptake from methane. Why ni-trogen uptake nonetheless proceeded is more difficult to explain.Conceivable explanations are that (i) methane cannot act asan energy source or a carbon source for the deep subseafloormicrobial community or (ii) the carbon turnover and/or in-corporation rate of cells that mediate anaerobic oxidation ofmethane may take place on longer timescales than the hetero-trophic substrates examined (28, 29). Nevertheless, even if cellsretrieved energy from methane or uncharacterized energy sour-ces, the amount of retrieved energy was insufficient to supportcell growth during the incubation experiment for 65 d.
Visualization of Active Cells Using CARD-FISH. We attempted toidentify 13C- and 15N-incorporating cells using CARD-FISH witharchaea- and bacteria-specific probes (30, 31). Contrary to ourexpectation, most metabolically active cells did not hybridize withcommon FISH probes (Fig. 2B). All cells that had incorporatedacetate and ∼50% of those cells that had incorporated aminoacids hybridized with the bacteria-specific probe, whereas lessthan 1% hybridized with the archaea-specific probe. AlthoughCARD-FISH can detect even a few ribosomal RNA molecules(30), detection generally relies on the permeation of relativelylarge horseradish peroxidase (HRP)-labeled probes across thecell membrane. Precise detection of archaeal cells surrounded bya rigid cell wall and/or crystal proteins is reportedly difficult (32).In addition, whether probes accurately match the sequences ofuncharacterized deep subseafloor life (33) and whether cells thatdo not hybridize with conventional probes belong to archaea,bacteria, or other unknown life forms remain unknown.
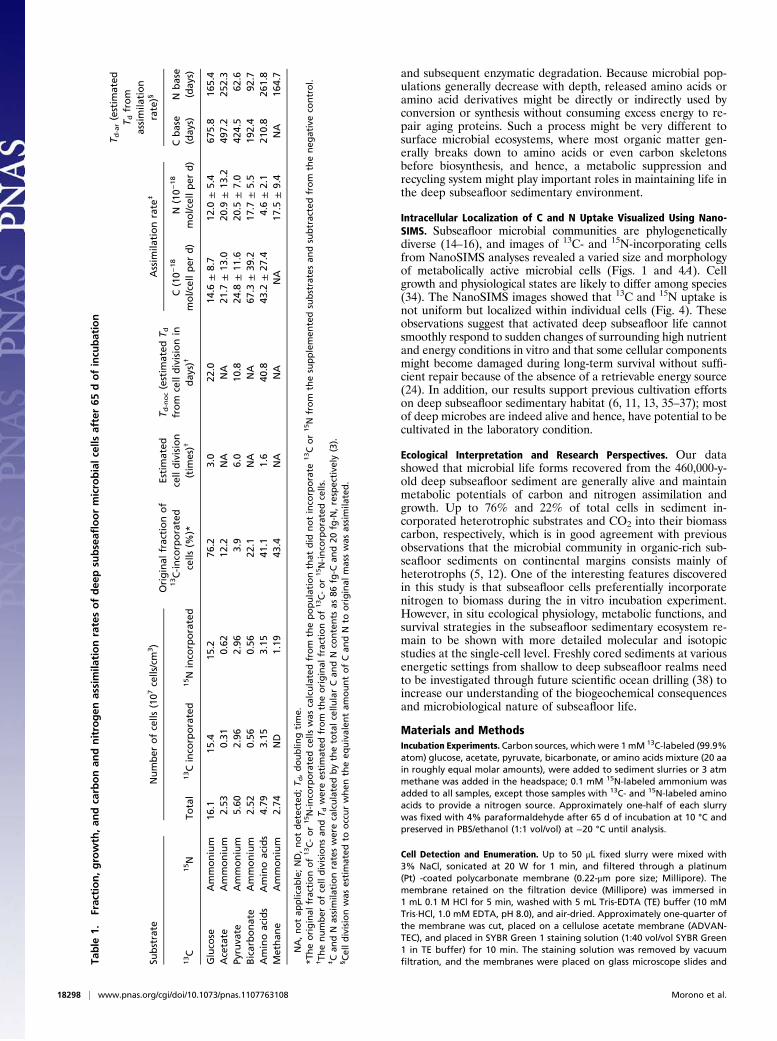
Growth Response to Nutrient and/or Energy Substrates. We calcu-lated 13C and 15N incorporation in individual cells (atom percentof cellular C and N) within each sample using NanoSIMS. Theamounts of carbon and nitrogen incorporated from glucose,pyruvate, and amino acids were significantly lower than expectedfrom cell division (Fig. 3A). The average doubling time (Td)estimated from cell numbers also considerably deviated (celldivision, Td-noc = 10.8–40.8 d) from those numbers estimated bythe assimilation rates of carbon and nitrogen substrates (Td-ar =62.6–675.8 d) (Table 1).One explanation is that cells under conditions in situ are lim-
ited by the availability of compounds for energy production butnot by the availability of compounds for biomass assimilation.After cells have been activated in vitro with labile, energy-richsubstrates, such as glucose, pyruvate, and amino acids, and are nolonger energy-starved, they will readily incorporate organiccompounds present in the environment. This finding indicatesthat not all carbon compounds used in biomass assimilation cansupport energy production. Our observation that acetate and bi-carbonate were incorporated during incubations without foster-ing cell growth supports this notion, albeit that possible energysources for acetate and bicarbonate incorporation remain elusive.The intracellular ratios of 13C to 15N notably showed that
atomic ratios of nitrogen incorporation from ammonium in thetotal cellular nitrogen consistently exceeded those ratios of car-bon (Figs. 3B and 4B). These data show that subseafloor cellspreferentially require nitrogen assimilation from ammonia for thebetter recovery in vitro, which may contradict the presence ofenough ammonium (∼15 mM) in the sediment pore water. Aconceivable explanation is that, although most subseafloormicrobes could assimilate nitrogen from ammonium, they mightsuppress ammonium uptake in situ to conserve or maintain en-ergy (i.e., although pore water from sediments at 219 mbsf con-tain 15 mM ammonium, the metabolism required for nitrogenassimilation into amino acids or calbamoyl phosphate synthesismight consume excess energy required for long-term survival).Another possibility is that subseafloor cells have incorporated thehigh C/N buried organic matter into the biomass rather thanammonium, which may subsequently result in nitrogen-deprivedbiomass. However, these assumptions need to be carefully shownin future studies by evaluating carbon and nitrogen composition(i.e., C/N ratio) and the physiological nature of subseafloor cells.The incorporation ratios of carbon only exceeded the ratios of
nitrogen in the incubation with amino acids to which no 15N-ammonium was added. Amino acids are released by cell death
0 20 40 60 80 100
UnidentifiedArchaeaBacteria
05101520
Number of 13C- or 15N-incorporated cells( x 107 cells / ml slurry)
Total cell13C-incorporated cell15N-incorporated cell
13C-Glucose+15NH4
13C-Acetate+15NH4
13C-Pyruvate+15NH4
13C-Bicarbonate+15NH4
13C15N-Amino acids mix
13C-Methane+15NH4
No addition
Fraction of 13C- or 15N-incorporated cells(%)
BA
Fig. 2. Numbers of subseafloor microbial cells that incorporated 13C- or 15N-labeled substrates. (A) Number of total cells (orange bars) and cells that in-corporated 13C (red bars) and 15N (green bars). (B) Phylogenetic identification of domain-specific probes for bacteria [EUB338 (I, II, III probes mix)] and archaea(ARC915) by CARD-FISH. Bacterial, archaeal, and unidentified (i.e., did not hybridize with these probes) fractions of cells that incorporated 13C and/or 15N areshown in blue, red, and gray, respectively.
Morono et al. PNAS | November 8, 2011 | vol. 108 | no. 45 | 18297
ENVIRONMEN
TAL
SCIENCE
SSE
ECO
MMEN
TARY
and subsequent enzymatic degradation. Because microbial pop-ulations generally decrease with depth, released amino acids oramino acid derivatives might be directly or indirectly used byconversion or synthesis without consuming excess energy to re-pair aging proteins. Such a process might be very different tosurface microbial ecosystems, where most organic matter gen-erally breaks down to amino acids or even carbon skeletonsbefore biosynthesis, and hence, a metabolic suppression andrecycling system might play important roles in maintaining life inthe deep subseafloor sedimentary environment.
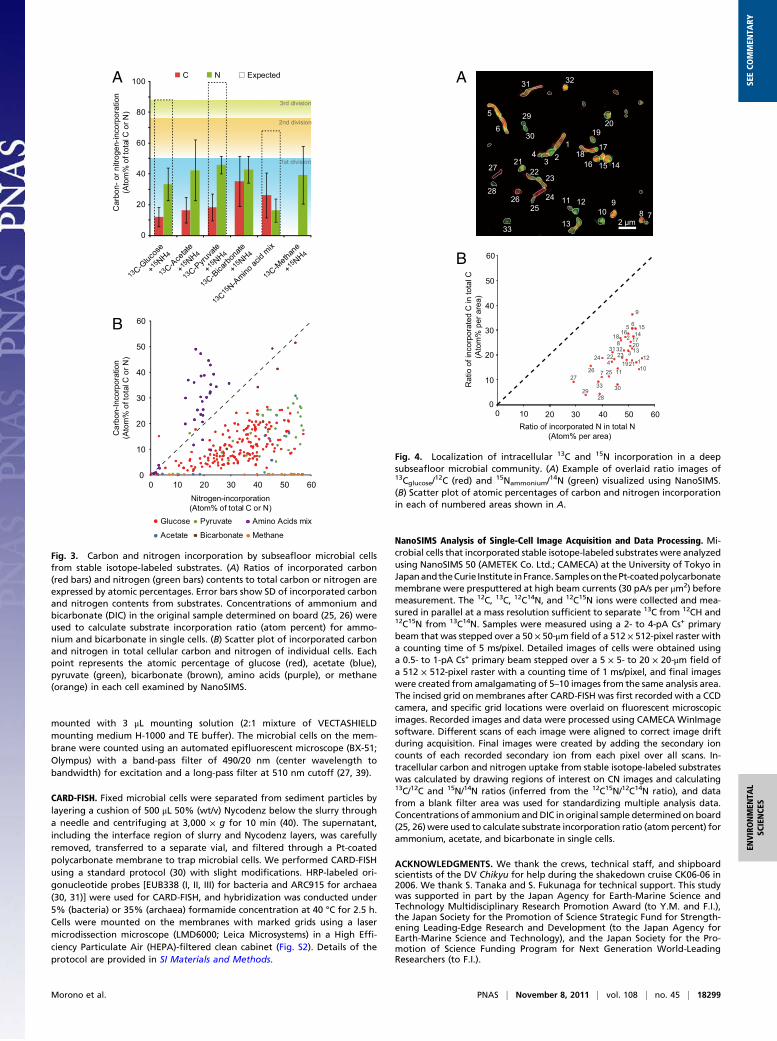
Intracellular Localization of C and N Uptake Visualized Using Nano-SIMS. Subseafloor microbial communities are phylogeneticallydiverse (14–16), and images of 13C- and 15N-incorporating cellsfrom NanoSIMS analyses revealed a varied size and morphologyof metabolically active microbial cells (Figs. 1 and 4A). Cellgrowth and physiological states are likely to differ among species(34). The NanoSIMS images showed that 13C and 15N uptake isnot uniform but localized within individual cells (Fig. 4). Theseobservations suggest that activated deep subseafloor life cannotsmoothly respond to sudden changes of surrounding high nutrientand energy conditions in vitro and that some cellular componentsmight become damaged during long-term survival without suffi-cient repair because of the absence of a retrievable energy source(24). In addition, our results support previous cultivation effortson deep subseafloor sedimentary habitat (6, 11, 13, 35–37); mostof deep microbes are indeed alive and hence, have potential to becultivated in the laboratory condition.
Ecological Interpretation and Research Perspectives. Our datashowed that microbial life forms recovered from the 460,000-y-old deep subseafloor sediment are generally alive and maintainmetabolic potentials of carbon and nitrogen assimilation andgrowth. Up to 76% and 22% of total cells in sediment in-corporated heterotrophic substrates and CO2 into their biomasscarbon, respectively, which is in good agreement with previousobservations that the microbial community in organic-rich sub-seafloor sediments on continental margins consists mainly ofheterotrophs (5, 12). One of the interesting features discoveredin this study is that subseafloor cells preferentially incorporatenitrogen to biomass during the in vitro incubation experiment.However, in situ ecological physiology, metabolic functions, andsurvival strategies in the subseafloor sedimentary ecosystem re-main to be shown with more detailed molecular and isotopicstudies at the single-cell level. Freshly cored sediments at variousenergetic settings from shallow to deep subseafloor realms needto be investigated through future scientific ocean drilling (38) toincrease our understanding of the biogeochemical consequencesand microbiological nature of subseafloor life.
Materials and MethodsIncubation Experiments. Carbon sources, which were 1mM 13C-labeled (99.9%atom) glucose, acetate, pyruvate, bicarbonate, or amino acids mixture (20 aain roughly equal molar amounts), were added to sediment slurries or 3 atmmethane was added in the headspace; 0.1 mM 15N-labeled ammonium wasadded to all samples, except those samples with 13C- and 15N-labeled aminoacids to provide a nitrogen source. Approximately one-half of each slurrywas fixed with 4% paraformaldehyde after 65 d of incubation at 10 °C andpreserved in PBS/ethanol (1:1 vol/vol) at −20 °C until analysis.
Cell Detection and Enumeration. Up to 50 μL fixed slurry were mixed with3% NaCl, sonicated at 20 W for 1 min, and filtered through a platinum(Pt) -coated polycarbonate membrane (0.22-μm pore size; Millipore). Themembrane retained on the filtration device (Millipore) was immersed in1 mL 0.1 M HCl for 5 min, washed with 5 mL Tris-EDTA (TE) buffer (10 mMTris·HCl, 1.0 mM EDTA, pH 8.0), and air-dried. Approximately one-quarter ofthe membrane was cut, placed on a cellulose acetate membrane (ADVAN-TEC), and placed in SYBR Green 1 staining solution (1:40 vol/vol SYBR Green1 in TE buffer) for 10 min. The staining solution was removed by vacuumfiltration, and the membranes were placed on glass microscope slides andTa
ble
1.Fraction,growth,an
dcarbonan
dnitrogen
assimila
tionratesofdee
psu
bseafl
oormicrobialcells
after65
dofincu
bation
Substrate
Number
ofcells
(107
cells/cm
3)
Original
fractionof
13C-inco
rporated
cells
(%)*
Estimated
celldivision
(tim
es)†
T d-noc(estim
ated
T dfrom
celldivisionin
day
s)†
Assim
ilationrate
‡
T d-ar(estim
ated
T dfrom
assimila
tion
rate)§
13C
15N
Total
13Cinco
rporated
15N
inco
rporated
C(10−
18
mol/cellper
d)
N(10−
18
mol/cellper
d)
Cbase
(day
s)N
base
(day
s)
Gluco
seAmmonium
16.1
15.4
15.2
76.2
3.0
22.0
14.6
±8.7
12.0
±5.4
675.8
165.4
Acetate
Ammonium
2.53
0.31
0.62
12.2
NA
NA
21.7
±13
.020
.9±
13.2
497.2
252.3
Pyruva
teAmmonium
5.60
2.96
2.96
3.9
6.0
10.8
24.8
±11
.620
.5±
7.0
424.5
62.6
Bicarbonate
Ammonium
2.52
0.56
0.56
22.1
NA
NA
67.3
±39
.217
.7±
5.5
192.4
92.7
Aminoacids
Aminoacids
4.79
3.15
3.15
41.1
1.6
40.8
43.2
±27
.44.6±
2.1
210.8
261.8
Methan
eAmmonium
2.74
ND
1.19
43.4
NA
NA
NA
17.5
±9.4
NA
164.7
NA,notap
plicab
le;ND,notdetected;T d
,doublin
gtime.
*Theoriginal
fractionof13C-or15N-inco
rporatedcells
was
calculatedfrom
thepopulationthat
did
notinco
rporate
13Cor15N
from
thesupplemen
tedsubstratesan
dsubtractedfrom
theneg
ativeco
ntrol.
†Th
enumber
ofcelldivisionsan
dT d
wereestimated
from
theoriginal
fractionof13C-or15N-inco
rporatedcells.
‡Can
dN
assimila
tionrateswerecalculatedbythetotalcellu
larCan
dN
contents
as86
fg-C
and20
fg-N
,respective
ly(3).
§Celldivisionwas
estimated
tooccurwhen
theeq
uivalen
tam
ountofCan
dN
tooriginal
masswas
assimila
ted.
18298 | www.pnas.org/cgi/doi/10.1073/pnas.1107763108 Morono et al.
mounted with 3 μL mounting solution (2:1 mixture of VECTASHIELDmounting medium H-1000 and TE buffer). The microbial cells on the mem-brane were counted using an automated epifluorescent microscope (BX-51;Olympus) with a band-pass filter of 490/20 nm (center wavelength tobandwidth) for excitation and a long-pass filter at 510 nm cutoff (27, 39).
CARD-FISH. Fixed microbial cells were separated from sediment particles bylayering a cushion of 500 μL 50% (wt/v) Nycodenz below the slurry througha needle and centrifuging at 3,000 × g for 10 min (40). The supernatant,including the interface region of slurry and Nycodenz layers, was carefullyremoved, transferred to a separate vial, and filtered through a Pt-coatedpolycarbonate membrane to trap microbial cells. We performed CARD-FISHusing a standard protocol (30) with slight modifications. HRP-labeled ori-gonucleotide probes [EUB338 (I, II, III) for bacteria and ARC915 for archaea(30, 31)] were used for CARD-FISH, and hybridization was conducted under5% (bacteria) or 35% (archaea) formamide concentration at 40 °C for 2.5 h.Cells were mounted on the membranes with marked grids using a lasermicrodissection microscope (LMD6000; Leica Microsystems) in a High Effi-ciency Particulate Air (HEPA)-filtered clean cabinet (Fig. S2). Details of theprotocol are provided in SI Materials and Methods.
NanoSIMS Analysis of Single-Cell Image Acquisition and Data Processing. Mi-crobial cells that incorporated stable isotope-labeled substrates were analyzedusing NanoSIMS 50 (AMETEK Co. Ltd.; CAMECA) at the University of Tokyo inJapanandtheCurie Institute inFrance. SamplesonthePt-coatedpolycarbonatemembrane were presputtered at high beam currents (30 pA/s per μm2) beforemeasurement. The 12C, 13C, 12C14N, and 12C15N ions were collected and mea-sured in parallel at a mass resolution sufficient to separate 13C from 12CH and12C15N from 13C14N. Samples were measured using a 2- to 4-pA Cs+ primarybeam that was stepped over a 50 × 50-μm field of a 512 × 512-pixel raster witha counting time of 5 ms/pixel. Detailed images of cells were obtained usinga 0.5- to 1-pA Cs+ primary beam stepped over a 5 × 5- to 20 × 20-μm field ofa 512 × 512-pixel raster with a counting time of 1 ms/pixel, and final imageswere created from amalgamating of 5–10 images from the same analysis area.The incised grid onmembranes after CARD-FISH was first recorded with a CCDcamera, and specific grid locations were overlaid on fluorescent microscopicimages. Recorded images and data were processed using CAMECA WinImagesoftware. Different scans of each image were aligned to correct image driftduring acquisition. Final images were created by adding the secondary ioncounts of each recorded secondary ion from each pixel over all scans. In-tracellular carbon and nitrogen uptake from stable isotope-labeled substrateswas calculated by drawing regions of interest on CN images and calculating13C/12C and 15N/14N ratios (inferred from the 12C15N/12C14N ratio), and datafrom a blank filter area was used for standardizing multiple analysis data.Concentrations of ammoniumandDIC in original sample determinedon board(25, 26)were used to calculate substrate incorporation ratio (atompercent) forammonium, acetate, and bicarbonate in single cells.
ACKNOWLEDGMENTS. We thank the crews, technical staff, and shipboardscientists of the DV Chikyu for help during the shakedown cruise CK06-06 in2006. We thank S. Tanaka and S. Fukunaga for technical support. This studywas supported in part by the Japan Agency for Earth-Marine Science andTechnology Multidisciplinary Research Promotion Award (to Y.M. and F.I.),the Japan Society for the Promotion of Science Strategic Fund for Strength-ening Leading-Edge Research and Development (to the Japan Agency forEarth-Marine Science and Technology), and the Japan Society for the Pro-motion of Science Funding Program for Next Generation World-LeadingResearchers (to F.I.).
Glucose
Acetate
Pyruvate
Bicarbonate
Amino Acids mix
Methane
10
20
30
40
50
60
10 20 30 40 50 600
0
0
20
40
60
80
100
13 C-Gluc
ose
+15 NH4
13 C-Ace
tate
+15 NH4
13 C-Pyru
vate
+15 NH4
13 C-Bica
rbona
te
+15 NH4
13 C15 N-Amino
acid
mix
13 C-Metha
ne
+15 NH4
C N Expected
Car
bon-
or n
itrog
en-in
corp
orat
ion
(A
tom
% o
f tot
al C
or N
)
1st division
2nd division
3rd division
A
B
Car
bon-
Inco
rpor
atio
n(A
tom
% o
f tot
al C
or N
)
Nitrogen-incorporation(Atom% of total C or N)
Fig. 3. Carbon and nitrogen incorporation by subseafloor microbial cellsfrom stable isotope-labeled substrates. (A) Ratios of incorporated carbon(red bars) and nitrogen (green bars) contents to total carbon or nitrogen areexpressed by atomic percentages. Error bars show SD of incorporated carbonand nitrogen contents from substrates. Concentrations of ammonium andbicarbonate (DIC) in the original sample determined on board (25, 26) wereused to calculate substrate incorporation ratio (atom percent) for ammo-nium and bicarbonate in single cells. (B) Scatter plot of incorporated carbonand nitrogen in total cellular carbon and nitrogen of individual cells. Eachpoint represents the atomic percentage of glucose (red), acetate (blue),pyruvate (green), bicarbonate (brown), amino acids (purple), or methane(orange) in each cell examined by NanoSIMS.
123
4
5
6
789
1011 12
13
141516
1718
1920
2122
23
2425
26
27
28
29
30
31 32
33
0
10
20
30
40
Rat
io o
f inc
orpo
rate
d C
in to
tal C
(Ato
m%
per
are
a)
Ratio of incorporated N in total N (Atom% per area)
20 30 40 50 60
1127
2928
33
26 7 25
30
2442231
231921 1
10
1232
3 13201714
155 6
9
216
188
50
60
100
2 µm
A
B
Fig. 4. Localization of intracellular 13C and 15N incorporation in a deepsubseafloor microbial community. (A) Example of overlaid ratio images of13Cglucose/
12C (red) and 15Nammonium/14N (green) visualized using NanoSIMS.
(B) Scatter plot of atomic percentages of carbon and nitrogen incorporationin each of numbered areas shown in A.
Morono et al. PNAS | November 8, 2011 | vol. 108 | no. 45 | 18299
ENVIRONMEN
TAL
SCIENCE
SSE
ECO
MMEN
TARY

1. Morita RY (1997) Bacteria in Oligotrophic Environments: Starvation-Survival Lifestyle(Chapman & Hall, New York), pp 1–529.
2. Roussel EG, et al. (2008) Extending the sub-sea-floor biosphere. Science 320:1046.3. Whitman WB, Coleman DC, Wiebe WJ (1998) Prokaryotes: The unseen majority. Proc
Natl Acad Sci USA 95:6578–6583.4. Parkes RJ, Barry AC, Peter W (2000) Recent studies on bacterial populations and
processes in subseafloor sediments: A review. Hydrogeol J 8:11–28.5. Lipp JS, Morono Y, Inagaki F, Hinrichs K-U (2008) Significant contribution of Archaea
to extant biomass in marine subsurface sediments. Nature 454:991–994.6. D’Hondt S, et al. (2004) Distributions of microbial activities in deep subseafloor
sediments. Science 306:2216–2221.7. D’Hondt S, Rutherford S, Spivack AJ (2002) Metabolic activity of subsurface life in
deep-sea sediments. Science 295:2067–2070.8. D’Hondt S, et al. (2009) Subseafloor sedimentary life in the South Pacific Gyre. Proc
Natl Acad Sci USA 106:11651–11656.9. Jørgensen BB, Boetius A (2007) Feast and famine—microbial life in the deep-sea bed.
Nat Rev Microbiol 5:770–781.10. Schippers A, et al. (2005) Prokaryotic cells of the deep sub-seafloor biosphere iden-
tified as living bacteria. Nature 433:861–864.11. Colwell FS, et al. (2008) Estimates of biogenic methane production rates in deep
marine sediments at Hydrate Ridge, Cascadia margin. Appl Environ Microbiol 74:3444–3452.
12. Biddle JF, et al. (2006) Heterotrophic Archaea dominate sedimentary subsurfaceecosystems off Peru. Proc Natl Acad Sci USA 103:3846–3851.
13. Inagaki F, et al. (2003) Microbial communities associated with geological horizons incoastal subseafloor sediments from the Sea of Okhotsk. Appl Environ Microbiol 69:7224–7235.
14. Inagaki F, et al. (2006) Biogeographical distribution and diversity of microbes inmethane hydrate-bearing deep marine sediments on the Pacific Ocean Margin. ProcNatl Acad Sci USA 103:2815–2820.
15. Teske AP (2006) Microbial communities of deep marine subsurface sediments: Mo-lecular and cultivation surveys. Geomicrobiol J 23:357–368.
16. Webster G, et al. (2006) Prokaryotic community composition and biogeochemicalprocesses in deep subseafloor sediments from the Peru Margin. FEMS Microbiol Ecol58:65–85.
17. Parkes RJ, et al. (2005) Deep sub-seafloor prokaryotes stimulated at interfaces overgeological time. Nature 436:390–394.
18. Sørensen KB, Teske A (2006) Stratified communities of active Archaea in deep marinesubsurface sediments. Appl Environ Microbiol 72:4596–4603.
19. Biddle JF, Fitz-Gibbon S, Schuster SC, Brenchley JE, House CH (2008) Metagenomicsignatures of the Peru Margin subseafloor biosphere show a genetically distinct en-vironment. Proc Natl Acad Sci USA 105:10583–10588.
20. Webster G, et al. (2009) Subsurface microbiology and biogeochemistry of a deep,cold-water carbonate mound from the Porcupine Seabight (IODP Expedition 307).Environ Microbiol 11:239–257.
21. Lever MA, et al. (2010) Acetogenesis in deep subseafloor sediments of the Juan deFuca Ridge flank: A synthesis of geochemical, thermodynamic, and gene-based evi-dence. Geomicrobiol J 27:183–211.
22. Futagami T, Morono Y, Terada T, Kaksonen AH, Inagaki F (2009) Dehalogenationactivities and distribution of reductive dehalogenase homologous genes in marinesubsurface sediments. Appl Environ Microbiol 75:6905–6909.
23. Hoehler TM (2004) Biological energy requirements as quantitative boundary con-ditions for life in the subsurface. Geobiology 2:205–215.
24. Price PB, Sowers T (2004) Temperature dependence of metabolic rates for microbialgrowth, maintenance, and survival. Proc Natl Acad Sci USA 101:4631–4636.
25. Aoike K (2007) CK06-06 D/V Chikyu Shakedown Cruise Offshore Shimokita, Labora-tory Operation Report (CDEX-JAMSTEC, Yokohama, Japan), pp 40–43.
26. Tomaru H, et al. (2009) Dating of dissolved iodine in pore waters from the gas hydrateoccurrence offshore Shimokita Peninsula, Japan: 129I results from the D/V Chikyushakedown cruise. Resour Geol 59:359–373.
27. Morono Y, Terada T, Masui N, Inagaki F (2009) Discriminative detection and enu-meration of microbial life in marine subsurface sediments. ISME J 3:503–511.
28. Treude T, et al. (2007) Consumption of methane and CO2 by methanotrophic mi-crobial mats from gas seeps of the anoxic Black Sea. Appl Environ Microbiol 73:2271–2283.
29. Wegener G, Niemann H, Elvert M, Hinrichs K-U, Boetius A (2008) Assimilation ofmethane and inorganic carbon by microbial communities mediating the anaerobicoxidation of methane. Environ Microbiol 10:2287–2298.
30. Pernthaler A, Pernthaler J, Amann R (2002) Fluorescence in situ hybridization andcatalyzed reporter deposition for the identification of marine bacteria. Appl EnvironMicrobiol 68:3094–3101.
31. Pernthaler A, Amann R (2004) Simultaneous fluorescence in situ hybridization ofmRNA and rRNA in environmental bacteria. Appl Environ Microbiol 70:5426–5433.
32. Kubota K, et al. (2008) Evaluation of enzymatic cell treatments for application ofCARD-FISH to methanogens. J Microbiol Methods 72:54–59.
33. Teske A, Sørensen KB (2008) Uncultured archaea in deep marine subsurface sedi-ments: Have we caught them all? ISME J 2:3–18.
34. Musat N, et al. (2008) A single-cell view on the ecophysiology of anaerobic photo-trophic bacteria. Proc Natl Acad Sci USA 105:17861–17866.
35. Toffin L, Webster G, Weightman AJ, Fry JC, Prieur D (2004) Molecular monitoring ofculturable bacteria from deep-sea sediment of the Nankai Trough, Leg 190 OceanDrilling Program. FEMS Microbiol Ecol 48:357–367.
36. Parkes RJ, et al. (2009) Culturable prokaryotic diversity of deep, gas hydrate sedi-ments: First use of a continuous high-pressure, anaerobic, enrichment and isolationsystem for subseafloor sediments (DeepIsoBUG). Environ Microbiol 11:3140–3153.
37. Imachi H, et al. (2011) Cultivation of methanogenic community from subseafloorsediments using a continuous-flow bioreactor. ISME J, 10.1038/ismej.2011.64.
38. Inagaki F, Hinrichs K-U, Kubo Y, Expedition 337 Project Team (2010) Deep coalbedbiosphere off Shimokita: Microbial processes and hydrocarbon system associated withdeeply buried coalbed in the ocean. The Integrated Ocean Drilling Program (IODP)Scientific Prospectus, 337, 10.2204/iodp.sp.337.2010.
39. Morono Y, Inagaki F (2010) Automatic slide-loarder fluorescence microscope fordiscriminative enumeration of subseafloorlife. Scientific Drilling 9:32–36.
40. Kallmeyer J, Smith DC, Spivack AJ, D’Hondt S (2008) New cell extraction procedureapplied to deep subsurface sediments. Limnol Oceanogr: Methods 6:236–245.
18300 | www.pnas.org/cgi/doi/10.1073/pnas.1107763108 Morono et al.