caracterización de hongos entomopatógenos … hongos entomopatgeno… · caracterización de...
TRANSCRIPT

Caracterización de aislamientos de hongos entomopatógenos de
los géneros Beauveria y Metarhizium asociados a insectos plaga
de palma de aceite (Elaeis guineensis Jaqc.)
Carolina Valencia Cortés
Universidad Nacional de Colombia
Facultad de Agronomía
Bogotá, Colombia
2015

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
Caracterización de aislamientos de hongos entomopatógenos de
los géneros Beauveria y Metarhizium asociados a insectos plaga
de palma de aceite (Elaeis guineensis Jaqc.)
Carolina Valencia Cortés
Tesis o trabajo de investigación presentado como requisito parcial para optar al título de:
Magister en Ciencias Agrárias con énfasis en Entomología
Director: Ph.D., Profesora Liliana María Hoyos Carvajal.
Universidad Nacional de Colombia
Facultad de Agronomía
Bogotá, Colombia
2015

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
Con todo mi amor para mi familia que en este mundo
me acompaña y para mis ángeles que desde el cielo me cuidan,
Mateo Andrés y Mauricio, para quien sino para ustedes.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
Agradecimientos
A la Universidad Nacional de Colombia por permitirme culminar este trabajo.
A el centro de investigación en Palma de Aceite Cenipalma por facilitarme el material
biológico para la realización de este trabajo.
A la Dr. Lilliana María Hoyos Carvajal, docente de la Universidad Nacional de Colombia,
por su orientación, gran paciencia y por alentarme a culminar este proceso.
A mi Madre, a mi Padre y mis Hermanos por su tiempo y colaboración.
A mis compañeros, Hanna Lorena Alvarado, Angie Barragán, Sandra Castillo, Estefanía
Macías, Camilo Gómez Correa.
A Jesús Alberto León y Wadith de León por toda la colaboración en la etapa de
laboratorio.
A mi gran amigo Iván Ayala Díaz.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
Tabla de contenido
Resumen ............................................................................................................................. 9
LISTA DE TABLAS ........................................................................................................... 11
LISTA DE FIGURAS ......................................................................................................... 13
LISTA DE SIMBOLOS Y ABREVIATURAS ..................................................................... 14
INTRODUCCIÓN GENERAL ............................................................................................... 15
El control biológico con hongos entomopatógenos en el contexto de producción de
palma de aceite ................................................................................................................. 15
Generalidades de los hongos entomopatógenos. ........................................................... 17
Clasificación de los hongos entomopatógenos ................................................................ 20
Beauveria Vuillemin (1912) ........................................................................................... 22
Metarhizium Metschnikoff (1883) ................................................................................. 23
Proceso infectivo de los hongos entomopatógenos en los insectos. .............................. 23
Bibliografía ........................................................................................................................ 27
CAPITULO 1: CRECIMIENTO DE Beauveria sp. y Metarhizium sp. EN DIFERENTES
MEDIOS Y TEMPERATURAS ............................................................................................. 38
1.1 Introducción ................................................................................................................ 38
1.2 Objetivo Específico ..................................................................................................... 40
1.3 Metodología ................................................................................................................ 40
1.3.1 Aislamientos utilizados en el estudio ................................................................... 40
1.3.2 Pruebas de crecimiento radial ............................................................................. 41
1.3.3 Análisis estadístico............................................................................................... 41
1.4. Resultados ................................................................................................................. 42
1.4.1. Crecimiento radial de los aislamientos evaluados en medio de cultivo Agar
Papa Dextrosa y Sabureaud Dextrosa Agar. ............................................................... 42

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
1.4.2 Crecimiento radial de los aislamientos evaluados a 10°C en dos medios de
cultivo ............................................................................................................................ 43
1.4.3 Crecimiento radial de los aislamientos evaluados a 26.5°C en dos medios de
cultivo ............................................................................................................................ 45
1.4.4 Crecimiento radial de los aislamientos evaluados a 30°C en dos medios de
cultivo ............................................................................................................................ 46
1.5 Discusión..................................................................................................................... 48
1.6 Bibliografía .................................................................................................................. 53
CAPITULO 2: TOLERANCIA DE LOS HONGOS ENTOMOPATÓGENOS A LA
RADIACIÓN ULTRAVIOLETA ............................................................................................. 57
2.1 Introducción ................................................................................................................ 57
2.1.1 Efecto de la RUV-B sobre los hongos ................................................................. 58
2.2 Objetivo Específico ..................................................................................................... 59
2.3 Metodología ................................................................................................................ 60
2.3.1 Aislamientos utilizados en el estudio ................................................................... 60
2.3.2 Aplicación de tratamiento con UV-B. ................................................................... 60
2.3.4 Cálculo de porcentaje de germinación. ............................................................... 61
2.3.5 Análisis estadístico............................................................................................... 61
2.4 Resultados .................................................................................................................. 61
2.4.1 Efecto de la UV-B sobre aislamientos de Beauveria .......................................... 61
2.5 Discusión..................................................................................................................... 67
2.6 Bibliografía .................................................................................................................. 71
CAPITULO 3: DETERMINACIÓN DE LA HIDROFOBICIDAD DE CONIDIAS DE HONGOS
ENTOMOPATÓGENOS. ...................................................................................................... 76
3.1 Introducción ................................................................................................................ 76
3.2 Hipótesis y Objetivos Específicos .............................................................................. 77
3.3 Metodología. ............................................................................................................... 78
3.3.1 Aislamientos utilizados en el estudio ................................................................... 78

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
3.3.2 Pruebas de hidrofobicidad ................................................................................... 78
3.3.4 Análisis estadístico............................................................................................... 78
3.4 Resultados .................................................................................................................. 79
3.5 Discusión..................................................................................................................... 81
3.6 Bibliografía .................................................................................................................. 83
CONCLUSIONES ................................................................................................................. 86
RECOMENDACIONES ........................................................................................................ 87
ANEXOS ........................................................................................................................... 88
Anexo 1. Aislamientos de Beauveria y Metarhizium colectados en diferentes zonas
palmeras procedentes de insectos plaga del cultivo de palma de aceite. ...................... 88
Anexo 2. Registro de cada aislamiento ............................................................................ 90

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
9
Resumen
Se realizó la caracterización de 36 aislamientos de Beauveria y Metarhizium en ellos se
evaluó la tasa de crecimiento en dos medios de cultivo Papa Dextrosa Agar (PDA) y
Sabouraud Dextrosa Agar (SDA), la tolerancia a radiación ultravioleta (UV-B) y la
hidrofobicidad. En los análisis se encontraron diferencias significativas P=0.05 en todos
los parámetros evaluados. Con respecto a la tasa de crecimiento en diferentes medios y
temperaturas se observó un mayor efecto de la temperatura que del medio de cultivo en el
crecimiento de los hongos. Ninguno de los aislamientos evaluados de Metarhizium tiene
la capacidad de crecer a 10°C independiente del medio de cultivo en el que sea
sembrado. Beauveria en general tiene la capacidad de crecer en las tres temperaturas
evaluadas existiendo variaciones en la tasa de crecimiento dependiendo de la
temperatura de incubación. Se observaron diferentes grados de tolerancia a la radiación
UV-B en los aislamientos de Beauveria y Metarhizium. En general en la medida que
aumentó el tiempo de exposición a UV-B también se incrementó la pérdida de
germinación de los aislamientos con respecto al testigo. Se encontraron aislamientos
como B028 con alta sensibilidad y B001 con alta tolerancia a la exposición UV-B. Con
respecto a la hidrofobicidad los aislamientos de Metarhizium y Beauveria evaluados se
comportaron como aislamientos hidrofóbicos y no presentaron diferencias significativas
estadísticas P= 0.05. Los aislamientos de Beauveria mostraron diferencias significativas
P=0.05 mostrando algunos de ellos porcentaje de permanencia de las conidias por
encima del 80% después de la aplicación de los tratamientos, lo que indicaría la presencia
de una cantidad mayor de hidrofobinas con respecto a los aislamientos que mostraron
porcentajes de retención de conidias inferiores al 22%.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
10
Summary
A characterization of 36 isolates of Beauveria and Metarhizium was done to evaluate the
grown rate in two culture media such as Potato Dextrose Agar (PDA) and Dextrose Agar
Sabouraud (SDA), the tolerance to UV radiation (UV -B) and the hydrophobicity. All
parameters evaluated showed significant differences (P=0.05). The growth rate had a
higher effect due temperature rather than media culture. None of Metarhizium isolates
tested has the ability to grow at 10 ° C independently of the culture medium evaluated.
Whereas, Beauveria isolates have the ability to grow at all three temperatures tested
showing variations in growth rate depending on the incubation temperature. Different
degrees of tolerance to UV -B radiation in Metarhizium and Beauveria were observed. In
general, when the exposure time to UV-B is increased, the germination of the isolates was
decreased compared with the control. We found isolates with high sensitivity to UV-B such
the isolate B028, and isolates with high tolerance as to UV-B such the B001. With respect
to the hydrophobicity of Metarhizium and Beauveria all isolates tested behaved as
hydrophobics (P=0.05). Beauveria isolates showed significant differences in
hydrophobicity (P=0.05) with percentages of permanence of conidia over 80 % in some
isolates after application of the treatments to evaluate the hydrophobicity, probability
related to the presence of a greater amount of hydrophobins compared to the isolates that
showed percentages retention of less than 22 % conidia.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
11
LISTA DE TABLAS
Tabla 1. Microorganismos entomopatógenos empleados para control de insectos plaga en
palma de aceite en Colombia bajo condiciones de laboratorio y campo.
Tabla 1.4. Ejemplos de algunos estudios recientes de investigaciones realizadas con
hongos entomopatógenos de los géneros Beauveria y Metarhizium para el manejo de
insectos plaga en diversos cultivos alrededor del mundo.
Capítulo 1
Tabla 1.1. Análisis de varianza para el área bajo la curva acumulada hasta el día 18 de
evaluación de crecimiento en aislamientos de Beauveria y Metarhizium procedentes de
palma de aceite.
Tabla 1.2. Grupos de crecimiento radial de aislamientos de Beauveria sembrados en
PDA y sometidos a diferentes temperaturas de crecimiento.
Tabla 1.3. Grupos de Crecimiento radial de aislamientos de Metarhizium en medios de
cultivo PDA y SDA incubados a 26.5°C y 30°C.
Capítulo 2
Tabla 2.1. Porcentaje de germinación de Beauveria de cada aislamiento, en función del
tiempo de exposición a la radiación UV-B1.
Tabla 2.2. Disminución de germinación de Beauveria en función del tiempo de
exposición a la radiación UV-B.
Tabla 2.3. Porcentaje de germinación de Metarhizium de cada aislamiento, en función
del tiempo de exposición a la radiación UV-B.
Tabla 2.4. Disminución de germinación de Metarhizium en función del tiempo de
exposición a la radiación UV-B.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
12
Capítulo 3
Tabla 3.1. Grupos de medias de los aislamientos de Beauveria en función del número de
conidias suspendidos en fase acuosa.
Tabla 3.2. Grupos de medias de los aislamientos de Metarhizium en función del número
de conidias suspendidos en fase acuosa.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
13
LISTA DE FIGURAS
Figura 1.1. Ubicación Taxonómica de los géneros de hongos entomopatógenos
Beauveria y Metarhizium.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
14
LISTA DE SIMBOLOS Y ABREVIATURAS
ADN: Acido Desoxirribonucleico
g: gramo
°C: grados centígrados
ha: hectárea
L: Litro
µl: microlitro
ml: mililitro
mm/día: milímetros / día
nm: nanómetro

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
15
INTRODUCCIÓN GENERAL
El control biológico con hongos entomopatógenos en el contexto de producción de
palma de aceite
La palma de aceite Elaeis guineensis Jacq. es una planta tropical que se desarrolla hasta
los 500 metros sobre el nivel del mar, entre las semillas oleaginosas es el cultivo de palma
de aceite el que produce mayor cantidad de aceite por hectárea. Su centro de origen es el
Golfo de Guinea (África occidental). Los principales países productores son Indonesia,
Malasia, Tailandia, Nigeria y Colombia (Fedepalma, 2013). En el año 2013, Colombia tuvo
un área sembrada de 452.435 de las cuales 299.953 ha se encontraban en producción y
152.482 ha en desarrollo (Fedepalma, 2013). En el mismo año Colombia contribuyó con el
1,81% de la producción mundial, ubicandose en el país con el primer lugar de producción
en el continente americano (Torres, 2013).
En cifras económicas internas, el sector palmero representa el 3.3% del PIB Agrícola. En
el año 2013, las siembras de palma de aceite se encontraban distribuidas en 116
municipios en 20 departamentos (Fedepalma, 2013) siendo en muchas poblaciones la
fuente primaria de empleo.
En el contexto productivo en Colombia, el cultivo de palma de aceite tiene problemas
fitosanitarios, entre los que se se cuentan un amplio número de especies de insectos y
ácaros fitófagos que causan la reducción del área foliar por consumo directo, como
Stenoma cecropia Meyrick y Loxotoma elegans Zeller (Lepidoptera: Elachistidae), otros
que atacan la raíz como Sagalassa valida Walter (Lepidoptera: Glyphipterygidae);
barrenadores del tallo como Rhynchophorus palmarum L., y Metamasius hemipterus (L.)
(Coleoptera: Curculionidae) o raspadores de fruto como Demotispa neivai (Bondar)
(Coleoptera: Chrysomelidae) (Genty et al. 1978; Calvache, 2003). Existe un grupo de
insectos como Rhynchophorus palmarum (Coleoptera: Dryophthoridae) o Leptopharsa
gibbicarina Froeschner (Hemiptera: Tingidae) considerados inductores o vectores de
algunos problemas de carácter patológico en el cultivo (Aldana et al. 2004).
Durante años se han venido desarrollando diferentes estrategias para el manejo de estos
insectos fitófagos, entre ellas la aplicación de insecticidas químicos, el control etológico
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
16
que incluye la captura y recolección de insectos en diferentes estados de desarrollo
mediante el uso de trampas con atrayentes, y el control biológico como el uso de insectos
depredadores (Aldana et al. 2004b; Bustillo et al. 2013) y parasitoides (Aldana et al.
2004a, Castillo et al. 2000). Con respecto a hongos entomopatógenos, los trabajos más
recientes asociados a insectos plaga del cultivo han permitido conocer la actividad
patogénica de algunos aislamientos sobre insectos de importancia económica en el cultivo
de la palma de aceite como L. femoratus, S. cecropia, D. neivai, L. elegans y S. aloeus
entre otros (Tabla 1).
Tabla 1 Hongos entomopatógenos empleados para control de insectos plaga en palma de aceite
en Colombia bajo condiciones de laboratorio y campo.
Microorganismo Clase Insecto plaga Estadío Referencia
Beauveria sp. Hongo
Ascomycete
Demotispa neivai
(Coleoptera:
Chrysomelidae)
Adultos Valencia et al. 2007.
Beauveria sp. Hongo
Ascomycete
Stenoma cecropia
(Lepidoptera: Stenomidae) Larvas
Valencia y Torres,
2007.
Beauveria sp. Hongo
Ascomycete
Leucothyreus femoratus
(Coleoptera:
Scarabaeidae)
Larvas Rodríguez et al.
2007.
Beauveria bassiana Hongo
Ascomycete
Loxotoma elegans
(Lepidoptera: Elachistidae) Larvas Monroy et al. 2011.
Beauveria bassiana Hongo
Ascomycete
Rhynchophorus palmarum
(Coleoptera:
Dryophthoridae)
Adultos y
larvas Alvarado et al. 2013.
Metarhizium sp. Hongo
Ascomycete
Leucothyreus femoratus
(Coleoptera: Scarabaeidae) Larvas
Rodríguez et al.
2007.
Metarhizium sp. Hongo
Ascomycete
Strategus aloeus
(Coleoptera: Scarabaeidae) Larvas Valencia et al. 2011.
Metarhizium
anisopliae
Hongo
Ascomycete
Rhynchophorus palmarum
(Coleoptera:
Dryophthoridae)
Adultos y
larvas Alvarado et al. 2013.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
17
La eficiencia del uso de estos agentes de control depende entre otros del conocimiento
sobre la biología del insecto y los mecanismos de interacción de estos con el hospedero,
además del ambiente en el cual se aplican. Para lograr programas eficientes de control
utilizando los hongos entomopatógenos es necesario desarrollar estudios de virulencia
biología y genética de estos enemigos naturales que permitan su selección y posterior
incorporación en programas de manejo integrado de plagas por regiones palmeras.
Generalidades de los hongos entomopatógenos.
En los últimos 50 años, la investigación en la protección de los cultivos contra los insectos
plaga ha estado enfocada al uso de agentes de biocontrol (BCA) (Whipps y Lumsden,
2001). La creciente inquietud por los efectos nocivos sobre la salud causados por las
prácticas de la agricultura tradicional intensiva que involucra el uso de insecticidas
sintéticos; la aparición de resistencia a los insecticidas químicos en algunos insectos de
importancia económica hacen que haya un interés cada vez mayor en el uso de
insecticidas de origen biológico (Glare et al. 2012). En consecuencia, desde mediados del
siglo XX ha habido gran expansión del mercado en base a hongos patógenos con
actividad selectiva contra las plagas de insectos (López-Pérez et al. 2014). El uso de
estos métodos de control se deriva de una disciplina denominada control biológico, que
se basa en el principio natural en que muchas especies de organismos se alimentan,
viven y se reproducen sobre otras, cuyas poblaciones son reguladas por las primeras en
los diferentes ecosistemas, indirectamente esto logra que las cantidades alcanzadas por
los grupos poblacionales no sean nocivas al sistema (Madrigal, 2001). Ernest y Brown
(2001) definen la homeostasis como “las interacciones compensatorias entre especies,
que producen estabilidad en un ecosistema”, en consecuencia los hongos
entomopatógenos y las enfermedades que de forma natural causan en los insectos son
un ejemplo claro de esta relación y el control biológico es el aprovechamiento de estas
interacciones regulatorias para disminuir las poblaciones de una plaga.
Entre los microorganismos entomopatógenos, los hongos son considerados controladores
promisorios, ya que pueden infectar a los insectos directamente a través de la penetración
de la cutícula y poseen múltiples mecanismos de acción que les confieren una alta
capacidad para evitar que el hospedero despliegue mecanismos de resistencia (Hayek y
Leger, 1994). Sin embargo, su crecimiento y desarrollo está limitado por las condiciones
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
18
ambientales externas, en particular por la humedad, la radiación solar y la temperatura,
que determinan la adecuada esporulación y germinación de las conidias (Tanada y Kaya,
1993).
Las características de los hongos entomopatógenos y su potencial en el control de
insectos plaga han dado lugar a una disciplina que ha desarrollado un gran número de
estudios encaminados a seleccionar los aislamientos más patogénicos para el control de
insectos plaga de diferentes órdenes en diversos cultivos alrededor del mundo (Tabla 2).
Tabla 2. Ejemplos de algunos estudios recientes de investigaciones realizadas con hongos
entomopatógenos de los géneros Beauveria y Metarhizium para el manejo de insectos plaga en
diversos cultivos alrededor del mundo.
Hongo
Entomopatógeno Insecto blanco
Área de
Economía
Cultivo o
Problema
asociado
Referencia
Beauveria bassiana Anopheles stephensi
(Diptera: Culicidae) Salud Publica
Transmisor de
Malaria Vogels et al. 2014.
Beauveria bassiana Arge Rosae
(Hymenoptera: Argidae)
Agricultura
Ornamentales Flores Khosravi et al. 2014.
Metarhizium
bruneum
Agriotes lineatus
Agriotes obscurus
Agriotes sputator
(Coleoptera: Elateridae)
Agricultura Papa Eckard et al. 2014.
Beauveria spp.
Metarhizium spp.
Cosmopoplites sordidus
(Coleoptera: Curculionidae) Agricultura Banano Lopes et al. 2013.
Debido al potencial de estos hongos para el manejo de plagas en diversos
agroecosistemas, muchos han sido modificados genéticamente con diferentes propósitos
entre los que se encuentran generar resistencia al fungicida benomyl (Goettel et al. 1990),
a los herbicidas bialaphos y al glufosinato de amonio (Inglis et al. 2000) para que sean
compatibles con planes de manejo integrado en los cuales se usan estos agentes
xenobióticos, también se han realizado modificaciones genéticas en puntos enzimáticos
importantes para el aumento de patogenicidad, la disminución de los tiempos letales
(Leger et al. 1996), sobreproducción de quitinasas en B. bassiana y la producción de
cutinasas en M. anisopliae (Fang et al. 2007) otro tipo de transformaciones incluyen el
aumento o resistencia al estrés particularmente al causado por la radiación ultravioleta
(Tseng et al. 2011).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
19
En la actualidad, se han utilizado entre 12 a 15 especies de hongos entomopatógenos
para el desarrollo de aproximadamente 170 productos (Faria y Wraight, 2007) sin
embargo, el número de especies de hongos que afectan insectos es sin duda superior
parasitando individuos en todos los órdenes de insectos, en su mayoría pertenecientes a
los órdenes Hemiptera, Diptera, Coleoptera, Lepidoptera, Hymenoptera y Orthoptera
(Ferron, 1978).
Según Faria y Wraight (2007), entre insecticidas, fungicidas y acaricidas, más 170
productos fueron desarrollados en las últimas tres décadas, 75% de los cuales
actualmente se hallan en proceso de registro. Los microorganismos como agentes de
control de plagas y enfermedades, son considerados un mercado en aumento que
incrementó sus ventas de cerca de 150 millones USD en 2000 a más de 500 millones
USD en 2008 (Ravensberg, 2011). El desarrollo de biopesticidas exitosos en el campo
depende entre otras, de la selección de aislamientos altamente patogénicos partiendo de
un número importante de candidatos que además deben ser resistentes a la desecación,
tolerantes a la radiación ultravioleta, de fácil producción masiva, amigables con el
ambiente e inocuos para seres vivos no blanco (Glare et al. 2012).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
20
Clasificación de los hongos entomopatógenos
Desde los primeros estudios realizados con hongos la clasificación de estos ha tenido
cambios importantes y se halla en constante revisión. El Phylum Ascomycota, que agrupa
una considerable cantidad de hongos asociados a insectos, se ha dividido en tres
subphylum Taphirinomycotina (140 especies), Saccharomycotina (915 especies) y
Pezizomycotina (63.011 especies) muchos miembros de Pezizomycotina incluyen
parásitos de abejas y algunos parásitos biótrofos de insectos (Hibbet et al. 2007;
Blackwell, 2011). Los hongos pertenecientes al subphylum Taphirinomycotina son
saprofitos y parásitos de plantas y vertebrados (Schoch et al. 2009), respecto al
subphylum Saccharomycotina se han reportado asociaciones mutualistas con insectos en
donde el insecto sirve como medio de dispersión del hongo y este a su vez provee
enzimas y otras sustancias útiles para el insecto (Vega y Dowd, 2005). El subphylum
Pezizomycotina se considera el más numeroso, además de ser ecológica y
morfológicamente el más complejo (Schoch et al. 2009), su ciclo de vida tiene dos
estados anamorfo y teleomorfo, ambos de ocurrencia en infecciones en insectos. Según
Vega et al. (2012) dentro del orden Hypocreales se evidencian varios ejemplos de este
fenómeno, pues incluye los hongos entomopatógenos más estudiados y empleados
comercialmente. El orden Hypocreales a su vez está dividido en siete familias, dentro de
estas, la familia polifilética Clavicipitaceae que incluye tres familias monofiléticas
Clavicipetaceae propiamente dicha, Cordycipetaceae y Ophiocordycipetaceae.
Comprende géneros como Aschersonia, Hypocrella, Regiocrella y Metarhizium, siendo
este último asociado al teleomorfo Metacordyceps (Vega et al. 2012; Humber, 2009;
Sung et al. 2007), también Beauveria, Isaria, Lecanicillium entre otros géneros (Vega et
al. 2012) (Figura 1.1).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
21
Figura 1.1. Ubicación Taxonómica de los géneros de hongos entomopatógenos Beauveria y
Metarhizium
Los anamorfos del orden Hypocreales entre los que se encuentran Beauveria y
Metarhizium son considerados generalistas, oportunistas, hemibiótrofos ya que
inicialmente atacan tejidos vivos del insecto en una fase biotrófica y luego mediante la
producción de toxinas pasan a la fase necrótrofa y luego saprofita (Hensket et al. 2010).
Los parásitos biótrofos de insectos son considerados extraños y únicamente se reportan
algunos casos exitosos en agricultura en el orden Laboulbeniales (Vega et al. 2008).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
22
Beauveria Vuillemin (1912)
Es un género cosmopolita, necrótrofo que se encuentra en el suelo, o como endófito en
plantas, saprofito facultativo de importancia para el control de insectos plaga alrededor del
mundo (Vega et al. 2008; Roberts y Hajek 1992; Goettel et al. 2005). También se
considera promisorio en la industria farmacéutica, en la agricultura por el gran número de
metabolitos y enzimas que produce (Vey et al. 2001). Dentro de las especies de este
género Beauveria bassiana (Bals.-Criv.) Vuill. (Ascomycota: Hypocreales) ha sido
reportado en varios tipos de interacciones con plantas entre ellos como supresor de
enfermedades y plagas (Vega, 2008; Vega et al. 2008) (Goettel et al. 2008; Ownley et al.
2008).
El género Beauveria Vuillemin (1912) está en constante revisión; Petch en 1926 reconoció
dos especies, B. bassiana (Bals.) Vuill. y B.brongniartii (Sacc.) Petch diferenciados por
sus conidias, posteriormente De Hoog en 1972 describió tres especies B. bassiana, B.
brongniartii y B. alba (Limber) Sacc, que posteriormente este mismo autor, renombra
como Engyodontium album (Limber) (De Hoog, 1978). Más tarde se describen cuatro
nuevas especies B. vermiconia De Hoog y Rao (1975), B. amorpha Samson y Evans
(1982), B. caledonica Bisset Widden (1986) y B. malawiensis Rehner et al. (2006).
Estudios de filogenética molecular ubican a Beauveria dentro de la familia
Cordycipitaceae (Hypocreales) (Sung et al. 2007). En la actualidad B. bassiana y B.
brongniartii son consideradas especies crípticas, por ejemplo B. bassiana incluye un
número aún no determinado de linajes muchos de ellos con distribuciones
intercontinentales (Rehner et al. 2006; Ghikas et al. 2010), y que se encuentran como
complejos de especies múltiples tanto en ambientes agrícolas como en los no
intervenidos (Rehner et al. 2006, Meyling et al. 2009).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
23
Metarhizium Metschnikoff (1883)
Desde su descripción en 1883 las especies del genero Metarhizium han sido ampliamente
estudiadas (Rocha et al. 2013), las investigaciones más recientes del complejo
Metarhizium anisopliae s.l. (Metschnikoff) reconocen diez especies: M. anisopliae s.s. y M.
pingshaense y las nuevas especies M. frigidum, M. globosum y M. robertsii, recupera M.
brunneum, eleva al nivel de especies tres variedades M. acridum, M. lepidiotae y M. majus
y finalmente reconoce a M. taii como una sinonimia de M. guizhouense. (Bischoff et al.
2006; Bischoff et al. 2009; Rocha et al. 2013).
Proceso infectivo de los hongos entomopatógenos en los insectos.
La mayoría de estos hongos son patógenos obligados o facultativos que causan en el
insecto enfermedades denominadas micosis (Tanada y Kaya, 1993). El proceso infectivo
comienza con la Adhesión de la conidia a la cutícula del insecto este proceso está
directamente relacionado con el tipo de conidias y la hidrofobobicidad de la cutícula del
insecto, cuando las conidias crecen aéreamente se unen bien a superficies hidrofobicas
pero la adhesión a superficies hidrofílicas es débil; las conidias que crecen sumergidas se
unen bien a ambos tipos de superficies y las blastosporas (In vitro) se unen de una
manera fuerte a superficies hidrofílicas y débilmente a superficies hidrofóbicas (Holder y
Keyhani, 2005). La adhesión de las conidias también puede estar influenciada por la
topografía cuticular, la presencia de setas o espinas puede facilitar este proceso (Sosa-
Gomez et al. 1997). Una vez la conidia se ha adherido comienza el proceso de
germinación que es el retorno de la actividad o metabolismo vegetativo, morfológicamente
es la emergencia de la célula vegetativa en forma de un tubo germinativo que crece sobre
la superficie cuticular formándose un apresorio que penetra la cutícula (Vargas, 2003),
factores como la humedad ambiental, la temperatura y estado de desarrollo, parte del
cuerpo y balance nutricional del insecto pueden influenciar la germinación de la conidia de
un hongo entomopatógeno (Volcy y Pardo, 1994; Tanada y Kaya, 1993). La rápida
germinación de las conidias puede ser una ventaja ya que evita la exposición a ambientes
que pueden ser adversos para el hongo y reduce la posibilidad de que el insecto tenga
una respuesta al ataque del mismo (Vega et al. 2012). Una vez la conidia ha germinado
comienza la penetración en la cutícula del hospedero que ocurre como resultado de una

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
24
combinación entre la degradación enzimática de la cutícula y la presión mecánica ejercida
por el extremo de una hifa invasiva que forma un botón de penetración donde la capa
cuticular es deformada por presión (Tanada y Kaya, 1993; Humber, 2009). El modo de
penetración principalmente depende de las propiedades de la cutícula del insecto, grosor,
esclerotización y la presencia de sustancias antifúngicas y nutricionales en ella (Charleyn,
1989). El mecanismo químico de penetración de la cutícula involucra una serie de
enzimas como proteasas, aminopeptidasas, esterasas, lipasas y quitinasas, las cuales
causan degradación del tejido en la zona de penetración (Vega et al. 2012). Durante este
proceso de penetración en algunos casos se produce una respuesta no celular, llamada
melanización cuticular que puede retardar o detener el proceso de germinación mediante
el endurecimiento de la cutícula que forma una barrera contra la penetración del hongo
(Humber, 2009; Schrank y Vainstein, 2010). Una vez que el hongo atraviesa la cutícula
debe vencer el sistema inmunológico del hospedero antes de entrar a la hemolinfa y
desarrollarse dentro del insecto (Madrigal, 2001). Después de llegar al hemocele, la
mayoría de los hongos pasan a una fase levaduriforme o de crecimiento por gemación.
Las infecciones causadas por hongos en su gran mayoría tardan más de dos días en
causar la muerte del hospedero. Se producen toxinas y enzimas, aunque algunos hongos
aparentemente no poseen toxinas, matan el insecto al consumir todos los nutrientes o por
destrucción física de sus tejidos (Bustillo, 2001). En la medida que avanza el proceso de
infección el hongo crece y comienza a absorber una proporción cada vez mayor de
nutrientes del hospedero, se da una creciente competencia por el oxígeno disuelto
disponible y por el crecimiento de las estructuras del hongo se produce una invasión del
espacio que impide la circulación de la sangre a través del hemocele (Humber, 2009). La
colonización del hemocele por el hongo produce una alteración del estado fisiológico del
insecto y de sus funciones de excreción y respiración provocando la muerte del insecto
(Humber, 2009). Los insectos tienen una serie de mecanismos de respuesta al ataque de
los hongos como la melanización, la fagocitosis o la encapsulación, también pueden
modificar su comportamiento esponiendose al sol para incrementar su temperatura
corporal (Rolff y Reynolds, 2009; Elliot et al. 2002; Roy et al. 2006). Por otro lado, los
hongos pueden evitar la defensa inmune de los insectos mediante diferentes mecanismos
como desarrollo de protoplastos que no son reconocidos por los hemocitos del insecto, la
formación de cuerpos hifales multiplicándose y dispersándose rápidamente (Samson et al.
1988) y la producción de micotoxinas (Tanada y Kaya, 1993). El papel de las toxinas en el
proceso de patogénesis es entre otros actuar como inhibidores de las reacciones de

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
25
defensa del hospedante por alteraciones de los hemocitos y retardo en la agregación de
las células de la hemolinfa (López, 1994). Posterior al crecimiento del hongo en el
hemocele, la micosis induce a síntomas fisiológicos anormales en el insecto y cambios en
la coloración en insectos atacados. Después de muerto el insecto, si existe una alta
disponibilidad de agua los hongos emergen al exterior a través de la cutícula y esporulan
sobre el cadáver produciendo inóculo para infectar a otros insectos. Si las condiciones no
son favorables, estos quedan dentro del cadáver, donde pueden sobrevivir por algunos
meses y eventualmente producir conidias cuando lleguen las condiciones favorables.
Las enzimas juegan un papel importante en el proceso patogénico de los hongos
entomopatógenos se han descubierto en el tubo germinativo proteasas, aminopeptidasas,
lipasas, esterasas, y N-acetil-glucosamidasa (quitinasas). Estudios in vitro indican que en
la digestión del integumento sigue una secuencia de lipasa-proteasa-quitinasa (Tanada y
Kaya, 1993). Los hongos B. bassiana, M. anisopliae, Paecilomyces spp. y L. lecanii,
producen grandes cantidades de proteasas y quitinasas en medios de cultivo líquido
(Guillespie, 1988). En varios aislamientos de B. bassiana y M. anisopliae la enzima
principal es una endoproteasa que disuelve la proteína matriz que cubre la quitina
cuticular. Por lo tanto, la producción de quitinasa ocurre después del proceso de infección
y una vez que el hongo atraviesa la cutícula debe vencer el sistema inmunológico del
hospedero antes de entrar a la hemolinfa y desarrollarse dentro del insecto (Guillespie et
al. 1997).
Beauveria spp. y Metarhizium producen ciclodepsipéptidos que alteran la permeabilidad
de las membranas celulares (Ngoka et al. 1999), estos tienen actividad antibiótica y
producen efectos letales en insectos aún en bajas concentraciones para el caso de
Beauveria el ciclodepsipéptido más conocido y estudiado es la beauvericina, también se
han descubierto otros como beauverolides (Ellsworth y Grove, 1977), bassianolides
(Suzuki et al. 1977), beauveriolides (Mochizuki et al. 1993) y bassiatina (Kagamizono et al.
1995). La oosporina es una dibenzonquinona tóxica producida especialmente por hongos
del género Beauveria, presenta propiedades repelentes y antialimentarias en insectos
(Rosas - Acevedo et al. 2003). Quesada-Moraga y Vey (2004) reportaron la existencia de
otra toxina a la que llamaron bassiacridina demostrando su toxicidad al ser inyectada
dentro del hemocele de langostas. En Metarhizium anisopliae el primer ciclodepsipéptido
aislado fue llamado Dextrusina, esta toxina aparentemente abre los canales de calcio en

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
26
las membranas de los músculos de los insectos (Schrank y Vainstein, 2010). Metarhizium
spp. produce entre otras toxinas citocalacinas, miroridinas (Kondo et al. 1980), viridoxina
(Gupta et al. 1993), macrolidos (Kozone et al. 2009) y tirosina betaina (Carollo et al.
2010).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
27
Bibliografía
Aldana de la Torre, J., Calvache G., H., Daza, C., y Aldana de la Torre, J. 2004.
Alternativas para siembra de plantas nectaríferas. Revista Palmas, 25 (especial, 2): 194-
204.
Aldana de la Torre, R., Calvache G., H., Higuera, O., Vanegas, M., y Ayala, L. 2004a.
Control de Cyparissius daedalus Cramer (Lepidóptera: Castniidae) con el nematodo
Steinernema carpocapsae. Revista Palmas 25 (especial, 2): 259-267.
Aldana de la Torre, J., Calvache G., H., Castaño, J., Valencia C., C., y Hernández S., J.
2004b. Aspectos biológicos y alternativas de control de Imatidium neivai Bondar
(Coleoptera: Chrysomelidae) raspador del fruto de la palma de aceite. Revista Palmas, 25
(especial, 2): 240-248.
Alvarado M., H., Montes, L., Gomes de Oliveira, H., Bustillo P., A., y Mesa F., E. 2013.
Patogenicidad de cepas de Metarhizium anisopliae (L.) y Beauveria bassiana sobre
Rhynchophorus palmarum. Revista Palmas, 34(2): 11-20.
Bischoff, J.F., Rehner, A.S., y Humber, R.A., 2006. Metarhizium frigidum sp. nov.: a
cryptic species of M. anisopliae and a member of the M. flavoviride. Mycologia 98: 737–
745.
Bischoff, J.F., Rehner, A.S., y Humber, R.A., 2009. A multilocus phylogeny of the
Metarhizium anisopliae lineage. Mycologia 101: 508–528.
Blackwell, M. 2011. The fungi: 1, 2, 3, 5.1 million species? Am. J. Bot., 98: 426-438.
Bustillo, A., Aldana de la T., R. & Serna, F. (2013, Diciembre 30). Corrección en la
identificación de la especie del barrenador gigante de la palma de aceite (Lepidoptera:
Castniidae). Palmas, 34(4), 21-22.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
28
Bustillo, A., 2001. Hongos en insectos y posibilidades de uso en el control biológico de
plagas en Colombia. In: seminario Uso de entomopatógenos en Colombia. Sociedad
Colombiana de Entomología. Bogotá, pp. 30–53.
Calvache, H. 2003. Manejo integrado de plagas en palma de aceite. Centro de
Investigación en palma de aceite. Nueva Gente Ltda, Bogotá.
Carollo, C. A., Calil, A. L. A., Schiave, L. A., Guaratini, T., Roberts, D. W., Lopes, N. P., y
Braga, G. U. L. 2010. Fungal tyrosine betaine, a novel secondary metabolite from conidia
of entomopathogenic Metarhizium spp. fungi. Fungal Biol., 114: 473-480.
Castillo M., S., Aldana de la Torre, J., Calvache G., H., y Grijalva, O. 2000. Evaluación de
técnicas de liberación de Trichogramma pretiosum Riley (Hymenoptera:
Trichogrammatidae) para el manejo de Stenoma cecropia Meyrick (Lepidoptera:
Stenomidae) en el cultivo de palma de aceite (Elaeis guineensis Jacq.). Revista Palmas,
21(especial, 1): 203-211.
Charleyn, A. K. 1989. Mycoinsecticides: present use and future prospects. BCPC mono.
Progress and prospects. USA. Insect control (43): 165-181.
Eckard, S., Ansari, MA, Bacher, S., Butt, TM, Enkerli, J., y Grabenweger, G. 2014.
Virulence of in vivo and in vitro produced conidia of Metarhizium brunneum strains for
control of wireworms. Crop Protection, 64: 137-142.
Elliot, S. L., Blanford, S., y Thomas, M. B. 2002. Host pathogen interactions in a varying
environment: temperature, behavioural fever and fitness. Proc. R. Soc. Lond. B, 269:
1599-1607.
Ellsworth, J. F., y Grove, J. F. 1977. Cyclodepsipeptides from Beauveria bassiana Bals.
Part 1. Beauverolides H and I. J. Chem. Soc. Perkin, 1(1): 270-273.
Ernest, S. K., y Brown, J. H. 2001. Homeostasis and compensation: the role of species
and resources in ecosystem stability. Ecology, 82(8): 2118-2132.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
29
Fan, Y., Fang, W., Guo, S., Pei, X., Zhang, Y., Xiao, Y., Li, D., Jin, K., Bidochka, M. J., y
Pei, Y. 2007. Increased insect virulence in Beauveria bassiana strains overexpressing an
engineered chitinase. Appl. Environ. Microbiol. 73: 295-302.
Faría M.R., y Wraight S.P. 2007. Mycoinsecticides and Mycoacaricides: A comprehensive
list with worldwide coverage and international classification of formulation types. Biological
Control 43: 237-256.
Federación Nacional de Cultivadores de Palma de Aceite FEDEPALMA. 2013. Anuario
estadístico 2013.
Ferron, P. 1978. Biological control of insect pest by entomopathogenous fungi. In: Annual
review of Entomology of United States (23). Annual Reviews 409-442.
Genty, P., Desmier De Chenon, R., Morin, J. 1978. Les ravages du palmier a huile en
Ámerique Latine. Oleagineux 33(7): 326-420.
Ghikas, D.V., Kouvelis, V.N., y Typas, M.A. 2010. Phylogenetic and biogeographic
implications inferred by mitochondrial intergenic region analyses and ITS1-5.8S-ITS2 of
the entomopathogenic fungi Beauveria bassiana and B. brongniartii. BMC Microbiol
10:174.
Glare, T., Caradus, J., Gelernter, W., Jackson, T., Keyhani, N., Köhl, J., y Stewart, A.
2012. Have biopesticides come of age?. Trends in biotechnology, 30 (5): 250-258.
Goettel, M.S., Eilenberg, J., y Glare, T., 2005. Entomopathogenic fungi and their role in
regulation of insect populations. En: Gilbert, L.B., Latrou, K. (eds.), Comprehensive
molecular insect science. Elservier Pergamon, Oxford, UK.
Goettel, M. S., Koike, M., Kim, J. J., Aiuchi, D., Shinya, R., y Brodeur, J. 2008. Potential of
Lecanicillium spp. for management of insects, nematodes and plant diseases. J. Invertebr.
Pathol., 98, 256:261.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
30
Goettel, M. S., St. Leger, R. J., Bhairi, S., Jung, M. K., Oakley, B. R., Roberts, D. W., y
Staples, R. C. 1990. Pathogenicity and growth of Metarhizium anisopliae stably
transformed to benomyl resistance. Curr. Genet., 17: 129-132.
Guillespie, A. 1988. Use of fungi to control pest of agricultural importance. In: Burge, M.
(Ed) Fungi in biological control systems. Manchester University Press, Manchester,
England. 269 p.
Guillespie, J. P., Kanost, M. R., y Trenczek, T. 1997. Biological mediators of insect
immunity. Annu. Rev. Entomol., 42: 611-643.
Gupta, S., Krasnoff, S. B., Renwick, J. A. A., Roberts, D. W., Steiner, J. R. y Clardy, J.
1993. Viridoxins A and B: novel toxins from the fungus Metarhizium flavoviride. J. Org.
Chem., 58: 1062-1067.
Hayek, A. y Leger, R. 1994. Interactions between fungal pathogens and insect host. In:
Annual Review of Entomology United States (39). USA: Annual Reviews. 293 – 322 p.
Hesketh H., E. Roy H., Eilenberg J., K. Pell J., y Hails R. S. 2010. Challenges in modelling
complexity of fungal entomopathogens in semi-natural populations of insects. BioControl
55: 55–73.
Holder, D. J., y Keyhani, N. O. 2005. Adhesion of the entomopathogenic fungus
Beauveria (Cordyceps) bassiana to substrates. Appl. Environ. Microbiol. 71: 5260-5266.
Humber, R. A. 2009. Entomogenous fungi. In M. Schaechter (Ed.) (3rd Ed.). San Diego:
Academic Press. Encyclopedia of Microbiology 3:443-456.
Inglis, P. W., Araga˜o, F. J. L., Frazao, H., Magalhaes, B. P., y Valadares- Inglis, M. C.
2000. Biolistic co-transformation of Metarhizium anisopliae var. acridum strain GC423 with
green fluorescent protein and resistance to glufosinate ammonium. FEMS Microbiol. Lett.,
191: 249-254.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
31
Kagamizono, T., Nishino, E., Matsumoto, K., Kawashima, A., Kishimoto, M., Sakai, N.,
He, B.-M., Chen, Z.-X., Adachi, T., Morimoto, S., y Hanada, K. 1995. Bassiatin, a new
platelet aggregation inhibitor produced by Beauveria bassiana K-717. J. Antibiot., 48,
1407:1412.
Khosravi, R., Sendi, J. J., Zibaee, A., y Shokrgozar, M. A. 2014. Virulence of four
Beauveria bassiana (Balsamo) (Asc. Hypocreales) isolates on rose sawfly, Arge rosae
under laboratory condition. Journal of King Saud University-Science.
Kondo, S., Meguriya, N., Mogi, H., Aota, T., Miura, K., Fujii, T., Hayashi, I., Makino, K.,
Yamamoto, M., y Nakajima, N. 1980. K- 582, a new peptide antibiotic. I. J. Antibiot., 33:
533-542.
Kozone, I., Ueda, J.-Y., Watanabe, M., Nogami, S., Nagai, A., Inaba, S., Ohya, Y., Tagaki,
M., y Shin-ya, K. 2009. Novel 24-membered macrolides, JBIR-19 and -20 isolated from
Metarhizium sp. fE61. J. Antibiot., 62: 159-162.
Leger, R. J., Joshi, L., Bidochka, M. J., y Roberts, D. W. 1996. Construction of an
improved mycoinsecticide overexpressing a toxic protease. Proc. Natl. Acad. Sci. USA,
93: 6349-6354.
López, F., Martínez, M.D. y Barandica, J.M. 1994. Four new species of the genus
Leptanilla from Spain - Relationships to other species and ecological issues. Sociobiology,
24 (2): 179-212.
López-Pérez, M., Rodríguez-Gómez, D., y Lopera, O. 2014. Production of conidia of
Beauveria bassiana in solid-state culture: current status and future perspectives. Critical
reviews in biotechnology, (0): 1-8.
Lopes, R. B., Mesquita, A. L. M., Tigano, M. S., Souza, D. A., Martins, I., y Faria, M. 2013.
Diversity of indigenous Beauveria and Metarhizium spp. in a commercial banana field and
their virulence toward Cosmopolites sordidus (Coleoptera: Curculionidae). Fungal
Ecology, 6(5): 356-364.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
32
Madrigal, A. 2001. Fundamentos de Control Biológico. Universidad nacional de Medellín,
Facultad de Ciencias. Medellín: Universidad Nacional de Colombia. 453 p.
Meyling, N.V., Lubeck, M., Buckley, E.P., Eilenberg, J., y Rehner, S.A., 2009. Community
composition, host range and genetic structure of the fungal entomopathogen Beauveria in
adjoining agricultural and seminatural habitats. Molecular Ecology 18: 1282–1293.
Mochizuki, K., Ohmori, K., Tamura, H., Shizuri, Y., Nishiyama, S., Miyoshi, W., y
Yamamura, S. 1993. The structures of bioactive cyclodepsipeptides, beauveriolides I and
II, metabolites of entomopathogenic fungi Beauveria sp. Bull Chem. Soc. Jpn, 66: 3041-
3046.
Monroy S., M., Aldana de la Torre, J., Mesa, E., Aldana de la Torre, R., y Oliveira, H.
2011. Patogenicidad de diferentes aislamientos del género Beauveria sobre larvas de
Loxotoma elegans (Lepidóptera: Elachistidae), defoliador de la palma de aceite en los
Llanos Orientales. Revista Palmas, 32 (1): 47-55.
Ngoka, L. C. M., Gross, M. L., y Toogood, P. L. 1999. Sodium-directed selective cleavage
of lactones: a method for structure determination of cyclodepsipeptides. Int. J. Mass
Spectrom, 182/183. 289 -298.
Ownley, B.H., Griffin, M.R., Klingeman, W.E., Gwinn, K.D., Moulton, K.J., y Pereira, R.M.
2008 Beauveria bassiana: Endophytic colonization and plant disease control. Journal of
invertebrate pathology, 98(3): 267-70.
Quesada-Moraga, E., y Vey, A. 2004. Bassiacridin, a protein toxic for locusts secreted by
the entomopathogenic fungus Beavueria bassiana. Mycol. Res., 108: 441-452.
Ravensberg, W. J. 2011. A Roadmap to the Successful Development and
Commercialization of Microbial Pest Control Products for Control of Arthropods. In:
Progress in Biological Control, Vol. 10.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
33
Rehner, S.A., Posada, F., Buckley, E.P., Infante, F., Castillo, A., y Vega, F.E. 2006.
Phylogenetic origins of African and Neotropical Beauveria bassiana s.l. pathogens of the
coffee berry borer, Hypothenemus hampei. J Invert Pathol 93:11–23.
Roberts, D.W., y A.E. Hajek. 1992. Entomopathogenic fungi as bioinsecticides, pp. 144-
159. In Frontiers in Industrial Mycology. G.F. Leatham, ed. Chapman & Hill, NY.
Rocha, L.F., Inglis, P.W., Humber, R.A., Kipnis, A., y Luz C. 2013. Occurrence of
Metarhizium spp. in Central Brazilian soils. Journal Basic Microbiol. 53(3): 251-259.
Rodriguez, F., Aldana, R. y Cayón, G. 2007. Evaluación de hongos entomopatógenos
para el control de larvas de Leucothyreus femoratus Burmeister (Coleoptera:
Scarabaeidae) defoliador de palma de aceite. Ceniavances. 146: 4.
Rolff, J. y Reynolds, S. E. (Eds.). 2009. Insect Infection and Immunity: Evolution, Ecology
and Mechanisms. Oxford: Oxford University Press.
Rosas-Acevedo, J. L., Boucias, D. G., Lezama R., K. Sims., y Pescador., A. 2003.
Exudate from sporulating cultures of Hirsutella thompsonii, inhibit oviposition by the
twospotted spider mites Tetranychus urticae. Applied and Experimental Acarology 39:
213-225.
Roy, H. E., Steinkraus, D., Eilenberg, J., Hajek, A. E., y Pell, J. K. 2006. Bizarre
interactions and endgames: entomopathogenic fungi and their arthropod hosts. Annu.
Rev. Entomol., 51: 331-357.
Samson, R.A., Evans, H.C., y Latge, J.P. 1988. Atlas of Entomopathogenic Fungi.
Springer-Verlag, New York, pp. 187.
Schoch, C. L., Sung, G.-H., López-Giráldez, F., Towsend, J. P., Miadlikowska, J.,
Hofstetter, V., Robbertse, B., Matheny, P. B., Kauff, F., Wang, Z., Gueidan, C., Andrie, R.
M., Trippe, K., Ciufetti, L. M., Wynns, A., Fraker, E., Hodkinson, B. P., Bonito, G.,
Groenewald, J. Z., Arzanlou, M., de Hoog, G. S., Crous, P. W., Hewitt, D., Pfister, D. H.,
Peterson, K., Gryzenhout, M., Wingfield, M. J., Aptroot, A., Suh, S.-O., Blackwell, M.,

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
34
Hillis, D. M., Griffith, G. W., Castlebury, L. A., Rossman, A. Y., Lumbsch, H. T., Lu¨ cking,
R., Bu¨ del, B., Rauhut, A., Diederich, P., Ertz, D., Geiser, D. M., Hosaka, K., Inderbitzin,
P., Kohlmeyer, J., Volkmann-Kohlmeyer, B., Mostert, L., O’Donnell, K., Sipman, H.,
Rogers, J. D., Shoemaker, R. A., Sugiyama, J., Summerbell, R. C., Untereiner, W.,
Johnston, P. R., Stenroos, S., Zuccaro, A., Dyer, P. S., Crittenden, P. D., Cole, M. S.,
Hansen, K., Trappe, J. M., Yahr, R., Lutzoni, F., y Spatafora, J. W. 2009. The Ascomycota
tree of life: a phylum-wide phylogeny clarifies the origin and evolution of fundamental
reproductive and ecological traits. Syst. Biol., 58: 224-239.
Schrank, A., y Vainstein, M. H. 2010. Metarhizium anisopliae enzymes and toxins.
Toxicon, 56: 1267-1274.
Sosa-Gomez, D. R., Boucias, D. G., Nation, J. L. 1997. Attachment of Metarhizium
anisopliae to the southern green stink bug Nezara viridula cuticle and fungistatic effect of
cuticular lipids and aldehydes. J. Invertebr. Pathol. 69: 31-39.
Sung, G. H., Hywel-Jones, N. L., Sung, J. M., Luangsa-ard, J. J., Shrestha, B., y
Spatafora, J. W. 2007. Phylogenetic classification of Cordyceps and the clavicipitaceous
fungi. Studies in mycology, 57: 5-59.
Suzuki, A., Kanaoka, M., Isogai, A., Murakoshi, S., Ichinoe, M., y Tamura, S. 1977.
Bassionalide, a new insecticidal cyclodepsipeptide from Beauveria bassiana and
Verticillium lecanii. Tetrahedron Lett., 25: 2167-2170.
Tseng, M. N., Chung, P. C., y Tzean, S. S. 2011. Enhancing the stress tolerance and
virulence of an entomopathogen by metabolic engineering of dihydroxynaphthalene
melanin biosynthesis genes. Applied and environmental microbiology, 77(13), 4508-4519.
Tanada, Y., y Kaya, H. 1993. Insect Pathology. Academy Press. New York: Mc-Graw Hill.
Torres Carrasco, R. 2013. Evolución y perspectivas de la palma de aceite en
América. Revista Palmas, 34: 236-244.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
35
Valencia C., C., Ayala D., I., Benítez, E., Torres, N., Barrera, E., y Herrera H., A. 2007.
Evaluación de estrategias de control y cuantificación de las pérdidas causadas por
Demotispa neivai Bondar, raspador del fruto de la palma de aceite. Revista Palmas,
28(1): 41-51.
Valencia C., C., Pérez, S., Aldana de la Torre, R., Mesa, E., y Gomes de Olivera, H. 2011.
Patogenicidad de hongos entomopatógenos del género Metarhizium sobre larvas de
Strategus aloeus L. (Coleoptera: Scarabaeidae), en condiciones de laboratorio. Revista
Palmas, 32 (4): 30-40.
Valencia, C., C., y Torres, N. 2007. Patogenicidad de hongos entomopatógenos del
género Beauveria sp., sobre larvas de Stenoma cecropia (Lepidoptera: Elaschistidae), en
condiciones de laboratorio. Colombia, Cenipalma, Ceniavances 147: 4.
Vargas, Flores., M.E. 2003. Caracterización de tres cepas de Beauveria brongniartii
(Saccardo) Petch y su virulencia en Phthorimaea operculella (Zeller) y Zymmetrischema
tangolias (Gyen). Oficina general del sistema de bibliotecas y biblioteca general Central
UNMSM. Perú. 14p.
Vega, Fernando E., Posada, Francisco., Aime, M. Catherine., Pava-Ripoll, Monica.,
Infante, Francisco y Rehner, Stephen A., 2008. Entomopathogenic fungal endophytes.
Publications from USDA-ARS / UNL Faculty.Paper 385. Biological Control 46: 72–82.
Vega, F.E. 2008. Insect pathology and fungal endophytes. Journal of invertebrate
pathology, 98(3): 277-9.
Vega, F. E., Meyling N.V., Luangsa-ard J.J y Blackwell M. 2012. Fungal
Entomopathogens. En: insect pathology 2da edición.
Vega, F. E., y Dowd, P. F. 2005. The role of yeasts as insect endosymbionts. In F. E.
Vega y M. Blackwell (Eds.), Insect Fungal Associations: Ecology and Evolution. New
York: Oxford University Press. p. 211:243.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
36
Vey, A., Hoagland, R.E., y Butt, T.M. 2001. Toxic Metabolites of Fungal Biocontrol Agents.
Fungi as Biocontrol Agents. Progress, Problems and Potential (Butt TM, Jackson C y
Magan N, Eds), pp. 311-346. CABI Publishing, Oxford, UK.
Vogels, C. B., Bukhari, T., y Koenraadt, C. J. 2014. Fitness consequences of larval
exposure to Beauveria bassiana on adults of the malaria vector Anopheles
stephensi. Journal of invertebrate pathology, 119: 19-24.
Volcy C., y V. Pardo. 1994. Principios de Micología. Décimo novena edición. Centro de
publicaciones, Universidad Nacional de Colombia. 141 p.
Whipps, J. M., y Lumsden, R. D. 2001. Commercial use of fungi as plant disease
biological control agents: status and prospects. Fungal biocontrol agents: progress,
problems and potential, 9-22.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
37
OBJETIVO GENERAL
Objetivo general
Caracterizar algunos parametros fisiológicos de hongos entomopatógenos de los
géneros Beauveria y Metarhizium aislados de insectos plagas de la palma de aceite, que
puedan ser útiles en su control.
Objetivos específicos:
Determinar la respuesta en el crecimiento de aislamientos de Beauveria y
de Metarhizium en medios de cultivo incubados a diferentes temperaturas.
Establecer el efecto de la luz ultravioleta, fracción UV-B de la luz solar, sobre la
germinación de aislamientos de Beauveria y Metarhizium.
Establecer la capacidad de adhesión de conidias de Beauveria y Metarhizium a través de
su hidrofobicidad.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
38
CAPITULO 1: CRECIMIENTO DE Beauveria sp. y Metarhizium sp.
EN DIFERENTES MEDIOS Y TEMPERATURAS
1.1 Introducción
El crecimiento de los microorganismos es un delta positivo entre la acumulación de
biomasa en relación con el tiempo, mediado por división celular y la formación de la
pared en hongos; está influenciado entre otros, por la luz, la temperatura, iones en el
medio, siendo estos factores limitantes si no se encuentran en las condiciones
específicas para cada tipo de organismo. El conocimientos de las condiciones óptimas
de cultivo de un organismo de importancia en control biológico permite ajustar los
parámetros para su multiplicación masiva con fines comerciales, además conocer las
condiciones favorables para su desarrollo, sugiere los rangos de zonas de vida y
sustratos que puede colonizar, además aporta elementos para el uso diferencial de
cepas de hongos en determinados ambientes. La velocidad a la cual ocurre el
crecimiento de un hongo a diferentes temperaturas y sustratos o medios de cultivo, está
determinada en gran parte por la capacidad del organismo de aprovechar las fuentes de
nutrientes disponibles (carbohidratos y proteínas, entre otros), de degradar las
sustancias presentes en el sustrato de crecimiento y por la habilidad del metabolismo de
ese organismo de funcionar dentro del rango de temperatura ofrecido por el medio
(Iskandarov et al. 2006; Fargues et al. 1992). La tasa de crecimiento de un
microorganismo es un factor importante a la hora de seleccionar un individuo para el
desarrollo de producciones masivas puesto que los hongos con una mayor celeridad de
crecimiento pueden ser buenos candidatos para formulaciones de bioinsecticidas
(Varela y Morales, 1996), un organismo de rápido crecimiento tiene cierta ventaja
competitiva una vez es aplicado en el campo.
Iskandarov et al. (2006) sostienen que la actividad de los hongos entomopatógenos
depende en gran medida de la composición nutricional del medio de cultivo en
laboratorio, ya que la relación C/N necesaria, la presencia de proteínas, y la demanda
de cada nutriente es diferente no sólo entre especies sino entre cepas de la misma
especie, por esto al composición de cada medio de cultivo usado para el crecimiento de
los hongos entomopatógenos tiene una directa relación con la producción y germinación
de las conidias y con la producción de biomasa. Vega et al. (2012) reportan los

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
39
rendimientos de esporas de tres hongos entomopatógenos en seis diferentes medios
líquidos contrastantes en la relación C/N encontrando que los rendimientos más altos de
esporas se obtuvieron en un medio que contiene una relación C/N de 10:1. En el caso
de M. anisopliae la nutrición influye en variables como el crecimiento, la esporulación y
la virulencia del hongo, encuentran que una relación C/N de 5,2:1 produjo las conidias
más virulentas entre los aislamientos utilizados en su estudio.
Iskandarov et al. (2006) mostraron que la composición del medio de cultivo influye en la
germinación de las conidias en M. anisopliae y B. bassianna comprobando que cada
uno requiere de diferentes condiciones para iniciar la germinación de las mismas, por
ejemplo M. anisopliae únicamente requiere sucrosa. Rangel et al. (2008) y Rangel et al.
(2011) plantean diferentes componentes o condiciones nutricionales en los medios de
crecimiento para inducir la producción de conidias más tolerantes a estrés en M.
robertsii, las conidias producidas bajo estrés nutritivo o estrés osmótico fueron
aproximadamente dos veces más tolerantes al calor y la radiación UV-B y germinaban
en un menor tiempo que las conidias producidas en un medio rico nutricionalmente. En
B. bassiana se han desarrollado estudios en donde se evidenció que la acumulación de
trehalosa y otros azucares está involucrada en el desarrollo de mecanismos de defensa
que protegen a las proteínas y las membranas celulares de la inactivación y
desnaturalización causadas por factores como calor, desecación o congelamiento y
estrés oxidativo causado por radicación solar (Liu et al. 2009; Ferreira et al. 2007).
En general, la esporulación y otros procesos que hace parte del ciclo infeccioso de los
hongos entomopatógenos es altamente dependiente no sólo de nutrientes, sino también
de condiciones ambientales adecuadas principalmente la temperatura y la humedad
relativa (Arthurs y Thomas, 2001). La mayoría de los hongos viven a condiciones
cálidas, con azúcares, medios ácidofílicos con pH entre 4 y 6 y aerobicos, y por
generalidad, la mayoría de los hongos filamentosos se desarrollan a 25°C, sin embargo,
tienen la capacidad de habitar en medios con temperaturas bajas psicrófilos, medias
mesófilos y altas termófilos, presentándose entre los dos primeros grupos los
termolerantes, con temperaturas óptimas de crecimiento por debajo de los rangos a los
cuales pueden resistir sin sufrir daños celulares (Walker y White, 2005). La mayoría de
los hongos entomopatógenos son mesófilos, con un crecimiento entre 10°C y 40 °C con
una temperatura óptima entre 25 y 35°C (Fernandes et al. 2010). De acuerdo a

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
40
McCammon y Rath (1994), las cepas de M. anisopliae pueden ser separadas por las
tasas de germinación a diferentes temperaturas, pues esta variable tiene un efecto
importante sobre la germinación y el crecimiento micelial de aislamientos por lo que esta
característica es utilizada como criterio de selección de aislamientos en programas de
manejo de plagas. Si bien algunos aislamientos o variedades pueden tolerar estar a
temperaturas altas, pueden sufrir un proceso denominado demora de crecimiento pos
estrés (post-stress growth delay PSGD), este término es acuñado por Keyser et al.
(2014) quienes evaluaron el efecto de la exposición de Metarhizium a 40°C y 5°C por
diferentes periodos de tiempo encontrando que los aislamientos sometidos a altas
temperaturas sobreviven, sin embargo, la exposición a 40°C afecta de una forma
importante el crecimiento, retardando el inicio del proceso de germinación cuando son
llevados a una temperatura de incubación de 28°C. Soares et al. (1983) y Ferron (1978)
afirman que el crecimiento de un aislamiento a una temperatura inferior de la óptima
retarda el desarrollo de la micosis pero no necesariamente afectan la mortalidad total
causada por cada hongo.
1.2 Objetivo Específico
Objetivo específico 1: Determinar la respuesta en el crecimiento de aislamientos
de Beauveria y de Metarhizium en medios de cultivo incubados a diferentes
temperaturas.
1.3 Metodología
1.3.1 Aislamientos utilizados en el estudio
Se evaluaron 31 aislamientos de hongos entomopatógenos, 24 de Beauveria sp. y siete
de Metarhizium , procedentes de diferentes zonas palmeras del país (Anexo 1) que en la
actualidad pertenecen a la colección de hongos entomopatógenos de la Corporación
Centro de Investigación en Palma de Aceite (Cenipalma). Los aislamientos fueron
encontrados afectando insectos asociados al cultivo de la palma. Se realizó su
aislamiento y conservación en Agar Sabureaud Dextrosa (SDA) a 4°C. Para cada vuna

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
41
de las pruebas cada uno de los aislamientos se sembraron en cajas de Petri con SDA
las cuales fueron incubadas a 26.5 °C durante 10 días, tiempo en el que se verificó la
esporulación.
1.3.2 Pruebas de crecimiento radial
Para evaluar el crecimiento radial de los aislamientos se usaron dos medios de cultivo,
Agar Sabouraud Dextrosa (SDA) Oxoid® (composición g.L-1: peptona micológica 10,
glucosa (dextrosa) 40, agar 15) y Agar Papa Dextrosa (PDA) Oxoid® (composición g.L-1:
extracto de papa 4, dextrosa 20, agar 15) a tres temperaturas 10°C; 26.5°C y 30°C. A
partir de cajas de Petri que contenían los aislamientos a evaluar, se preparó una
suspensión de cada uno y se ajustó la concentración a 1x106 conidias.ml-1. En el centro
de cada caja que contenía el respectivo medio (PDA ó SDA), se ubicó un disco de papel
filtro de 0.64 mm de diámetro, sobre el cual se depositaron 5 µl de suspensión de
conidias de cada aislamiento (aprox. 5000 conidias). Cada combinación aislamiento x
medio fue incubado a tres temperaturas diferentes 10°C, 26.5°C y 30°C. El tiempo de
incubación y evaluación comprendió un total de 30 días, durante los cuales se midió el
diámetro de las colonias cada tercer día con un calibrador Vernier. Se realizaron cuatro
repeticiones por tratamiento por cada uno de los medios de cultivo y temperaturas
evaluadas.
1.3.3 Análisis estadístico
Se realizó un análisis de varianza, a partir del cual se detectó que la interacción:
aislamiento por temperatura por medio de cultivo, posteriormente mediante la función
AUDPC, del paquete Agricolae, desarrollado por De Mendiburu (2009), se obtuvo el
área bajo la curva (ABC) para cada una de las combinaciones aislamiento X medio de
cultivo X temperatura hasta la evaluación registrada el día 18 después de siembra,
obteniéndose una base de datos con estructura factorial: 31 aislamientos x 3
temperaturas x 2 medios y, una sola variable respuesta, el ABC acumulada.
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
42
1.4. Resultados
1.4.1. Crecimiento radial de los aislamientos evaluados en medio de cultivo Agar
Papa Dextrosa y Sabureaud Dextrosa Agar.
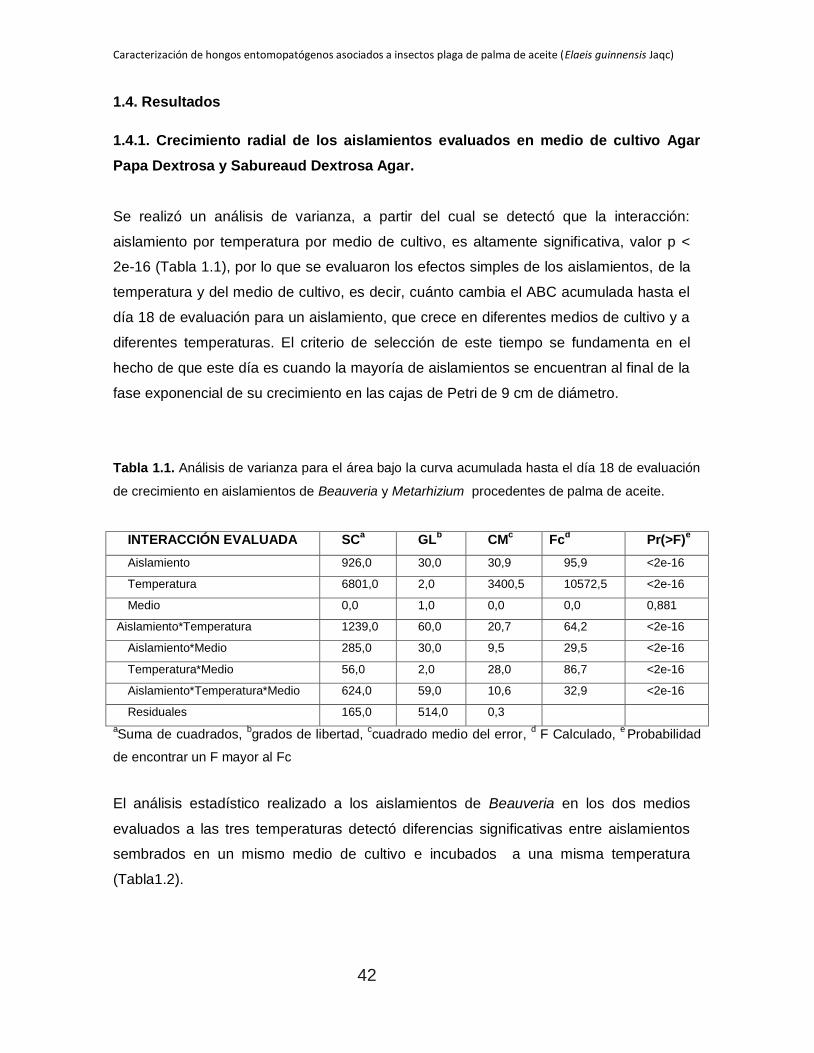
Se realizó un análisis de varianza, a partir del cual se detectó que la interacción:
aislamiento por temperatura por medio de cultivo, es altamente significativa, valor p <
2e-16 (Tabla 1.1), por lo que se evaluaron los efectos simples de los aislamientos, de la
temperatura y del medio de cultivo, es decir, cuánto cambia el ABC acumulada hasta el
día 18 de evaluación para un aislamiento, que crece en diferentes medios de cultivo y a
diferentes temperaturas. El criterio de selección de este tiempo se fundamenta en el
hecho de que este día es cuando la mayoría de aislamientos se encuentran al final de la
fase exponencial de su crecimiento en las cajas de Petri de 9 cm de diámetro.
Tabla 1.1. Análisis de varianza para el área bajo la curva acumulada hasta el día 18 de evaluación
de crecimiento en aislamientos de Beauveria y Metarhizium procedentes de palma de aceite.
INTERACCIÓN EVALUADA SCa
GLb
CMc
Fcd
Pr(>F)e
Aislamiento 926,0 30,0 30,9 95,9 <2e-16
Temperatura 6801,0 2,0 3400,5 10572,5 <2e-16
Medio 0,0 1,0 0,0 0,0 0,881
Aislamiento*Temperatura 1239,0 60,0 20,7 64,2 <2e-16
Aislamiento*Medio 285,0 30,0 9,5 29,5 <2e-16
Temperatura*Medio 56,0 2,0 28,0 86,7 <2e-16
Aislamiento*Temperatura*Medio 624,0 59,0 10,6 32,9 <2e-16
Residuales 165,0 514,0 0,3
aSuma de cuadrados,
bgrados de libertad,
ccuadrado medio del error,
d F Calculado,
e Probabilidad
de encontrar un F mayor al Fc
El análisis estadístico realizado a los aislamientos de Beauveria en los dos medios
evaluados a las tres temperaturas detectó diferencias significativas entre aislamientos
sembrados en un mismo medio de cultivo e incubados a una misma temperatura
(Tabla1.2).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
43
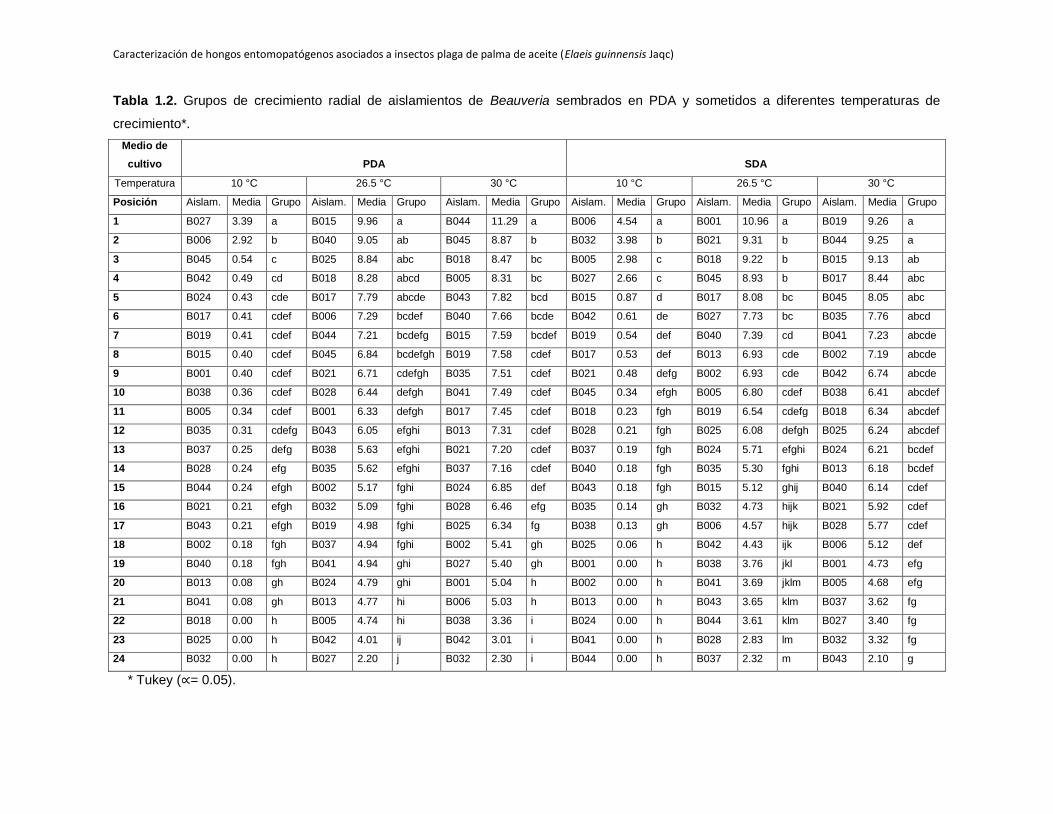
1.4.2 Crecimiento radial de los aislamientos evaluados a 10°C en dos medios de
cultivo
En general, se observó que los aislamientos de Beauveria estudiados tienen la
capacidad de crecer a esta temperatura de incubación en los dos medios de cultivo
evaluados. En PDA, 21 de los 24 aislamientos mostraron algún tipo de crecimiento, en
SDA solo 18 de la totalidad de aislamientos evaluados crecieron a esta temperatura de
incubación después de 18 días de evaluación.
La máxima ABC registrada corresponde a B006 en PDA (4.54), este mismo aislamiento
en SDA se ubicó entre los que tuvieron las mayores ABC hasta el día 18 de evaluación,
B027 presentó un comportamiento similar en los dos medios de cultivo evaluados, en
PDA obtiene el mayor ABC 3.39 y en SDA también se ubica entre los aislamientos que
presentan los mayores valores de ABC 2.66, lo que indicaría que estos dos tienen un
comportamiento psicrófilo. Los aislamientos B005 y B032 que en SDA se ubican entre
los que muestran las mayores ABC 2.98 y 3.98 respectivamente, en PDA tienen valores
mínimos de ABC de 0.34 y 0 respectivamente.
Las ABC registradas en PDA son inferiores a las observadas en SDA en todos los
aislamientos a excepción de B027 que muestra una mayor ABC en PDA (3.39) que en
SDA (2.66). Algunos aislamientos como B001, B002, B013, B024, B041 y B044 no tienen
la capacidad de crecer en SDA mientras que cuando son cultivados en PDA a la misma
temperatura de incubación muestran crecimiento después de 18 días de incubación
(Tabla 1.2). Al realizar un análisis estadístico de los aislamientos sembrados en ambos
medios de cultivo en PDA y en SDA se forman varios grupos estadisticamente diferentes
entre los aislamientos con mayores ABC y con respecto a los demás (Tabla 1.2). De los
siete aislamientos de Metarhizium evaluados en los dos medios de cultivo SDA y PDA
ninguno muestra crecimiento a esta temperatura de incubación.
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
Tabla 1.2. Grupos de crecimiento radial de aislamientos de Beauveria sembrados en PDA y sometidos a diferentes temperaturas de
crecimiento*.
Medio de
cultivo PDA SDA
Temperatura 10 °C 26.5 °C 30 °C 10 °C 26.5 °C 30 °C
Posición Aislam. Media Grupo Aislam. Media Grupo Aislam. Media Grupo Aislam. Media Grupo Aislam. Media Grupo Aislam. Media Grupo
1 B027 3.39 a B015 9.96 a B044 11.29 a B006 4.54 a B001 10.96 a B019 9.26 a
2 B006 2.92 b B040 9.05 ab B045 8.87 b B032 3.98 b B021 9.31 b B044 9.25 a
3 B045 0.54 c B025 8.84 abc B018 8.47 bc B005 2.98 c B018 9.22 b B015 9.13 ab
4 B042 0.49 cd B018 8.28 abcd B005 8.31 bc B027 2.66 c B045 8.93 b B017 8.44 abc
5 B024 0.43 cde B017 7.79 abcde B043 7.82 bcd B015 0.87 d B017 8.08 bc B045 8.05 abc
6 B017 0.41 cdef B006 7.29 bcdef B040 7.66 bcde B042 0.61 de B027 7.73 bc B035 7.76 abcd
7 B019 0.41 cdef B044 7.21 bcdefg B015 7.59 bcdef B019 0.54 def B040 7.39 cd B041 7.23 abcde
8 B015 0.40 cdef B045 6.84 bcdefgh B019 7.58 cdef B017 0.53 def B013 6.93 cde B002 7.19 abcde
9 B001 0.40 cdef B021 6.71 cdefgh B035 7.51 cdef B021 0.48 defg B002 6.93 cde B042 6.74 abcde
10 B038 0.36 cdef B028 6.44 defgh B041 7.49 cdef B045 0.34 efgh B005 6.80 cdef B038 6.41 abcdef
11 B005 0.34 cdef B001 6.33 defgh B017 7.45 cdef B018 0.23 fgh B019 6.54 cdefg B018 6.34 abcdef
12 B035 0.31 cdefg B043 6.05 efghi B013 7.31 cdef B028 0.21 fgh B025 6.08 defgh B025 6.24 abcdef
13 B037 0.25 defg B038 5.63 efghi B021 7.20 cdef B037 0.19 fgh B024 5.71 efghi B024 6.21 bcdef
14 B028 0.24 efg B035 5.62 efghi B037 7.16 cdef B040 0.18 fgh B035 5.30 fghi B013 6.18 bcdef
15 B044 0.24 efgh B002 5.17 fghi B024 6.85 def B043 0.18 fgh B015 5.12 ghij B040 6.14 cdef
16 B021 0.21 efgh B032 5.09 fghi B028 6.46 efg B035 0.14 gh B032 4.73 hijk B021 5.92 cdef
17 B043 0.21 efgh B019 4.98 fghi B025 6.34 fg B038 0.13 gh B006 4.57 hijk B028 5.77 cdef
18 B002 0.18 fgh B037 4.94 fghi B002 5.41 gh B025 0.06 h B042 4.43 ijk B006 5.12 def
19 B040 0.18 fgh B041 4.94 ghi B027 5.40 gh B001 0.00 h B038 3.76 jkl B001 4.73 efg
20 B013 0.08 gh B024 4.79 ghi B001 5.04 h B002 0.00 h B041 3.69 jklm B005 4.68 efg
21 B041 0.08 gh B013 4.77 hi B006 5.03 h B013 0.00 h B043 3.65 klm B037 3.62 fg
22 B018 0.00 h B005 4.74 hi B038 3.36 i B024 0.00 h B044 3.61 klm B027 3.40 fg
23 B025 0.00 h B042 4.01 ij B042 3.01 i B041 0.00 h B028 2.83 lm B032 3.32 fg
24 B032 0.00 h B027 2.20 j B032 2.30 i B044 0.00 h B037 2.32 m B043 2.10 g
* Tukey (∝= 0.05).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
45
1.4.3 Crecimiento radial de los aislamientos evaluados a 26.5°C en dos medios de cultivo
Todos los aislamientos evaluados de Beauveria y Metarhizium mostraron crecimiento a 26.5°C
tanto en PDA como en SDA. Con respecto a Beauveria en SDA, el rango de ABC oscila entre
10.96 (B001) y 2.32 (B037) Aislamientos como B001, B021, B018, B045 y B017 muestran
ABC con valores superiores 10.96, 9.31, 9.22, 8.93 y 8.08 por lo que se podrían considerar
aislamientos con una tasa de crecimiento alta, dentro de este grupo es estadisticamente
diferente de los demás únicamente el aislamiento B001, en contraste este aislamiento no tuvo
la capacidad de crecer en el mismo medio de cultivo cuando fue incubado a 10°C. Los demás
aislamientos con ABC inferiores a 7.73 son estadisticamente diferentes entre ellos Tukey (∝=
0.05). El comportamiento de las ABC de los aislamientos sembrados en PDA es similar
respecto a los valores presentados en SDA, independientemente del hongo evaluado. Las
mayores ABC en PDA fueron las presentadas por B015, B040 y B025 con un valor medio de
9.96, 9.05 y 8.84 respectivamente, siendo entre estos B015 estadisticamente diferente de
B040 y B025 y de los demás aislamientos evaluados Tukey (∝= 0.05). Los aislamientos B017
y B018 se ubican entre los aislamientos con mayores ABC tanto en PDA como en SDA. El
aislamiento B027 en SDA a 26.5°C se ubicó en el tercio de los aislamientos con mayor media
de ABC (7.73) en contraste en PDA ocupó el último lugar en media de ABC (2.20) entre los
aislamientos evaluados lo que indica que es más beneficioso para su desarrollo el medio con
más contenido de proteínas y que a esta temperatura de incubación hay un mayor efecto del
medio de cultivo que de la temperatura sobre la tasa de crecimiento (Tabla 1.2). Aunque B025
no presentó crecimiento a 10°C en PDA, cuando se expone a 26.5°C se ve beneficiado y junto
con B015 y B040 presentan mayores valores de ABC; pero su desarrollo disminuye
nuevamente cuando se incuba a 30°C, este aislamiento entonces se puede considerar un
organismo con un rango de temperatura específico y estrecho para su crecimiento adecuado.
Con respecto a Metarhizium todos los aislamientos sembrados en PDA e incubados a esta
temperatura muestran crecimiento. Mt009 tuvo la mayor ABC (13.82) y Mt010 tuvo la menor
ABC en esta temperatura de incubación (5.16), aunque no son amplias las diferencias, si se
puede deducir un posible efecto del medio de cultivo ya que en PDA, en general las ABC son
levemente mayores que en SDA en donde estadísticamente se forman dos grupos,
Mt006(9.26), Mt002(8.68), Mt009(8.51), Mt005(8.34) y Mt010(7.98) se ubican dentro del grupo
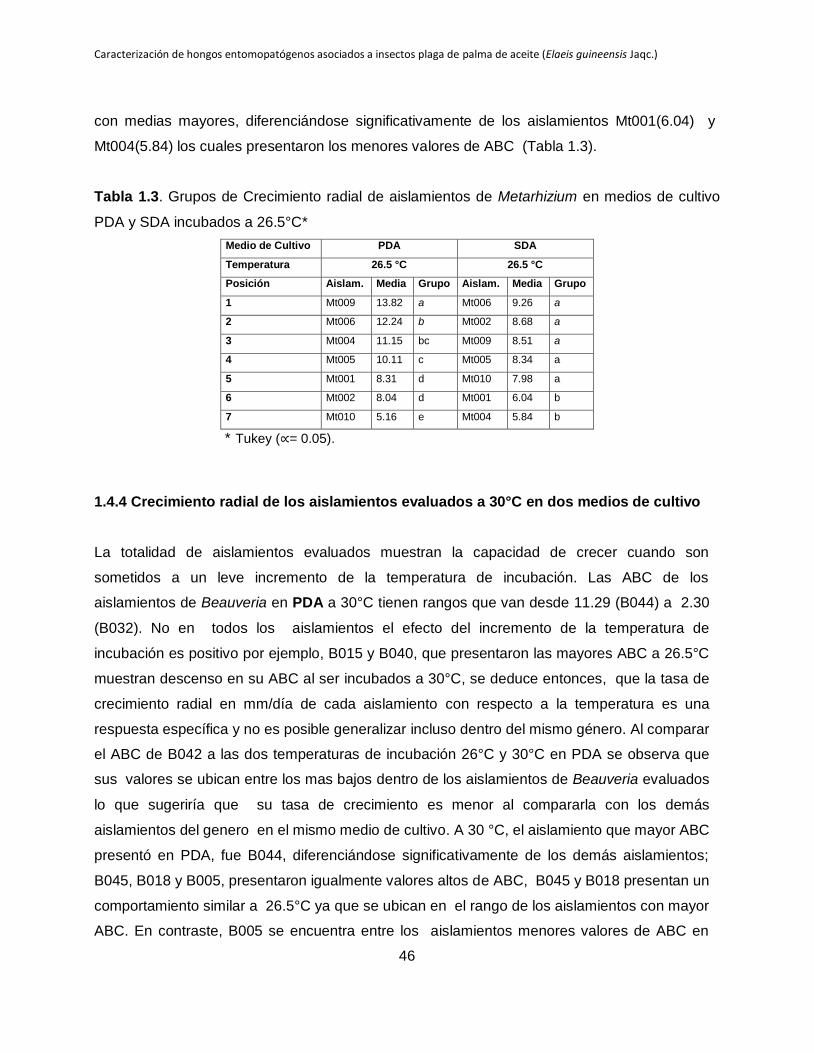
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
46
con medias mayores, diferenciándose significativamente de los aislamientos Mt001(6.04) y
Mt004(5.84) los cuales presentaron los menores valores de ABC (Tabla 1.3).
Tabla 1.3. Grupos de Crecimiento radial de aislamientos de Metarhizium en medios de cultivo
PDA y SDA incubados a 26.5°C*
Medio de Cultivo PDA SDA
Temperatura 26.5 °C 26.5 °C
Posición Aislam. Media Grupo Aislam. Media Grupo
1 Mt009 13.82 a Mt006 9.26 a
2 Mt006 12.24 b Mt002 8.68 a
3 Mt004 11.15 bc Mt009 8.51 a
4 Mt005 10.11 c Mt005 8.34 a
5 Mt001 8.31 d Mt010 7.98 a
6 Mt002 8.04 d Mt001 6.04 b
7 Mt010 5.16 e Mt004 5.84 b
* Tukey (∝= 0.05).
1.4.4 Crecimiento radial de los aislamientos evaluados a 30°C en dos medios de cultivo
La totalidad de aislamientos evaluados muestran la capacidad de crecer cuando son
sometidos a un leve incremento de la temperatura de incubación. Las ABC de los
aislamientos de Beauveria en PDA a 30°C tienen rangos que van desde 11.29 (B044) a 2.30
(B032). No en todos los aislamientos el efecto del incremento de la temperatura de
incubación es positivo por ejemplo, B015 y B040, que presentaron las mayores ABC a 26.5°C
muestran descenso en su ABC al ser incubados a 30°C, se deduce entonces, que la tasa de
crecimiento radial en mm/día de cada aislamiento con respecto a la temperatura es una
respuesta específica y no es posible generalizar incluso dentro del mismo género. Al comparar
el ABC de B042 a las dos temperaturas de incubación 26°C y 30°C en PDA se observa que
sus valores se ubican entre los mas bajos dentro de los aislamientos de Beauveria evaluados
lo que sugeriría que su tasa de crecimiento es menor al compararla con los demás
aislamientos del genero en el mismo medio de cultivo. A 30 °C, el aislamiento que mayor ABC
presentó en PDA, fue B044, diferenciándose significativamente de los demás aislamientos;
B045, B018 y B005, presentaron igualmente valores altos de ABC, B045 y B018 presentan un
comportamiento similar a 26.5°C ya que se ubican en el rango de los aislamientos con mayor
ABC. En contraste, B005 se encuentra entre los aislamientos menores valores de ABC en

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
47
PDA a 26.5 °C. Con respecto al ABC de los aislamientos de Beauveria sembrados en SDA e
incubados a 30°C los valores oscilan entre 9.26 (B019) y 2.10 (B043), se observó que las ABC
en general tienen un comportamiento similar al presentado cuando los aislamientos son
incubados a 26.5°C. A 30 °C los aislamientos que en SDA a 10°C mostraban el mayor ABC
como B005, B006, B027 y B032, pasan al tercio de aislamientos con menor ABC con valores
de 4.68, 5.12, 3.40 y 3.32 respectivamente, lo que sugiere un comportamiento psicrófilo,
diferenciándose significativamente de los aislamientos B019, B044, B015, B017, B045 los
cuales presentan la mayor ABC a 30 °C. En SDA a 30 °C, los aislamientos B019 y B044
presentaron la mayor ABC y presentaron diferencias significativas con el resto de
aislamientos, sin embargo, B044 se encontró entre los aislamientos con menor ABC, tanto a
10 °C como a 26.5 °C sugieriendo un efecto positivo del incremento de la temperatura de
incubación.
El ABC de los aislamientos de Metarhizium sembrados en PDA oscila entre 11.24 (Mt006) y
4.44 (Mt002). Al realizar una prueba de Tukey (∝= 0.05) se forman dos grupos de aislamientos
con ABC que muestran diferencias significativas (a,b) en el primero se incluyen los
aislamientos Mt006, Mt009, Mt004 y Mt005 con medias 11.24, 10.49, 9.91, 9.50
respectivamente. En el segundo grupo, se ubican tres aislamientos de Metarhizium Mt010,
Mt001 y Mt002 con medias de ABC 4.83, 4.63 y 4.44 respectivamente (Tabla 1.4). Los
aislamientos de Metarhizium sembrados en SDA incubados a 30°C muestran ABC con medias
que varían entre 12.85 (Mt005) y 9.26 (Mt010). En general, en SDA las medias de las ABC
para los aislamientos de Metarhizium son superiores a las presentadas en PDA, por ejemplo,
Mt002 que tuvo una media de ABC de 4.44 en PDA y de 12,11 en SDA (Tabla 1.4). Al realizar
una prueba de Tukey (∝= 0.05), se observan diferencias significativas en las medias del ABC
formando dos grupos en el primero se todos los aislamientos a excepción de Mt010 que se
ubica en el segundo grupo (Tabla 1.4). Particularmente se debe resaltar que Mt010, presentó
medias bajas de ABC tanto en PDA como en SDA a 30°C sin embargo, esta última a pesar de
ser la más baja dentro del tratamiento es la mejor ABC expresada por este hongo en todo el
ensayo, pudiendo ser que presente intrínsicamente tasas de crecimiento bajas o que el rango
de temperaturas evaluadas no incluyen el óptimo para su crecimiento.
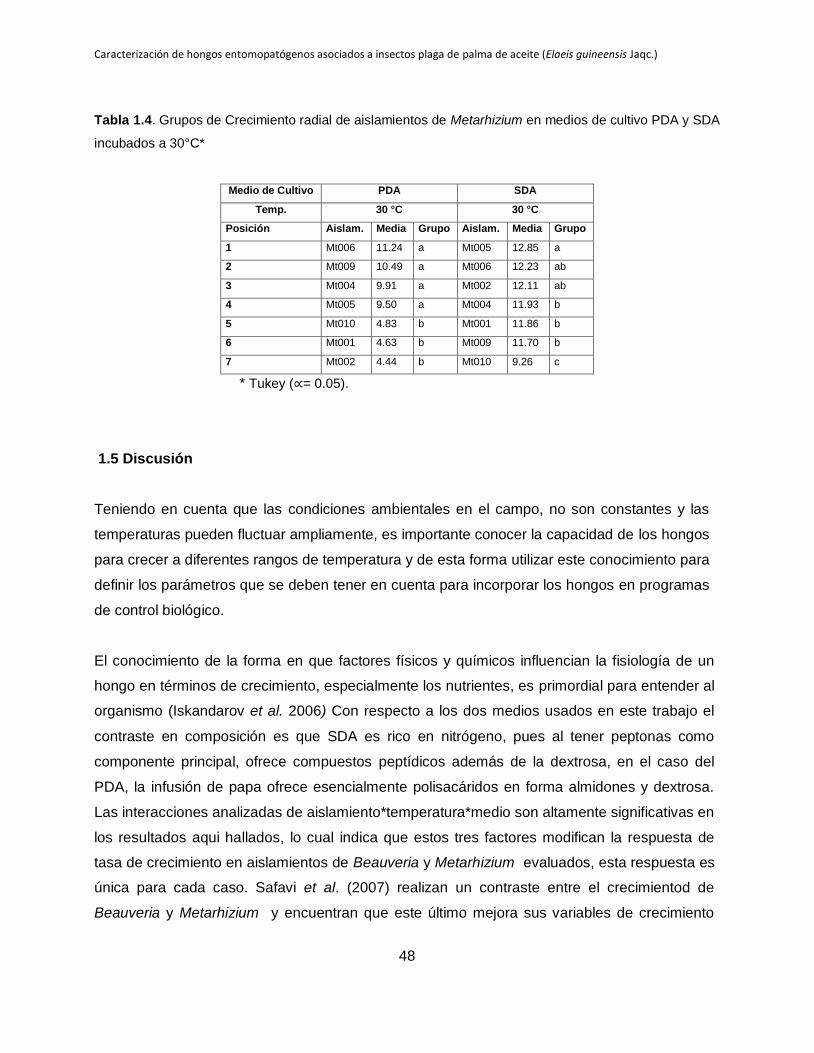
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
48
Tabla 1.4. Grupos de Crecimiento radial de aislamientos de Metarhizium en medios de cultivo PDA y SDA
incubados a 30°C*
Medio de Cultivo PDA SDA
Temp. 30 °C 30 °C
Posición Aislam. Media Grupo Aislam. Media Grupo
1 Mt006 11.24 a Mt005 12.85 a
2 Mt009 10.49 a Mt006 12.23 ab
3 Mt004 9.91 a Mt002 12.11 ab
4 Mt005 9.50 a Mt004 11.93 b
5 Mt010 4.83 b Mt001 11.86 b
6 Mt001 4.63 b Mt009 11.70 b
7 Mt002 4.44 b Mt010 9.26 c
* Tukey (∝= 0.05).
1.5 Discusión
Teniendo en cuenta que las condiciones ambientales en el campo, no son constantes y las
temperaturas pueden fluctuar ampliamente, es importante conocer la capacidad de los hongos
para crecer a diferentes rangos de temperatura y de esta forma utilizar este conocimiento para
definir los parámetros que se deben tener en cuenta para incorporar los hongos en programas
de control biológico.
El conocimiento de la forma en que factores físicos y químicos influencian la fisiología de un
hongo en términos de crecimiento, especialmente los nutrientes, es primordial para entender al
organismo (Iskandarov et al. 2006) Con respecto a los dos medios usados en este trabajo el
contraste en composición es que SDA es rico en nitrógeno, pues al tener peptonas como
componente principal, ofrece compuestos peptídicos además de la dextrosa, en el caso del
PDA, la infusión de papa ofrece esencialmente polisacáridos en forma almidones y dextrosa.
Las interacciones analizadas de aislamiento*temperatura*medio son altamente significativas en
los resultados aqui hallados, lo cual indica que estos tres factores modifican la respuesta de
tasa de crecimiento en aislamientos de Beauveria y Metarhizium evaluados, esta respuesta es
única para cada caso. Safavi et al. (2007) realizan un contraste entre el crecimientod de
Beauveria y Metarhizium y encuentran que este último mejora sus variables de crecimiento

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
49
radial, producción de esporas y porcentaje de germinación en medios con altos contenidos de
C y bajos de peptonas. En contraste, tres aislamientos de Beauveria evaluados, crecen mejor
en medios con contenidos de N (SDA) en forma de extracto de levadura o peptonas, sin
embargo, es posible notar que esta respuesta es bastante específica. Keyser et al. (2014)
mencionan que Metarhizium sp. en PDA a 30°C crece menos que en SDA a 26.5°C estos
hallazgos resultan contrastantes con los hallados en este estudio ya que solo algunos de los
Metarhizium aquí evaluados tienen esta tendencia, lo que permite deducir que un adecuado
medio de cultivo contrarresta el posible efecto negativo que sobre el hongo pueden tener las
altas temperaturas, por ello esta interacción o requerimiento nutricional es exclusivo de cada
aislamiento y al parecer no está asociados a taxa particulares.
La temperatura óptima de crecimiento de un hongo entomopatógeno es determinante en la
eficiencia con la que actúe el microorganismo dentro de un programa de manejo de plagas ya
que no solo afecta procesos fisiológicos en general, sino también la virulencia y eficiencia de la
infección. Iskandarov et al. (2006) con respecto al crecimiento de aislamientos de B. bassiana y
M. anisopliae a diferentes temperaturas, definen que los hongos incubados entre 15°y 35°C
tienen una alta germinación (95%) después de 10 a 15 horas de incubación, en contraste, en
los mismos aislamientos incubados a 10°C durante 50 horas no superan el 10% de
germinación. A 40°C las conidias comienzan a germinar a las 48 horas de incubación
expresando una mejor tasa de germinación en M. anisopliae lo que indica una adaptación de
este hongo a las altas temperaturas. Autores como Keyser et al. 2014; Rangel, 2005;
Fernandes et al. 2010 afirman que Metarhizium puede desarrollarse bien a temperaturas
entre 37 a 40°C e incluso por encima de 45°C, los aislamientos de Metarhizium evaluados en
este trabajo muestran un comportamiento similar ya que se observa una tendencia a mayores
ABC acumuladas en la medida en que se incrementa la temperatura de incubación, sin
embargo, las temperaturas aquí evaluadas no son superiores a 30°C, por lo que se tendría que
evaluar el crecimiento de estos a temperaturas entre 35°C y 40°C. Según Fernandes et al.
(2010), ciertas cepas de M. anisopliae que crecen a 37°C no crecen a bajas temperaturas
(5°C), mientras que los aislados que crecen a 5°C no crecen a temperaturas más elevadas,
algunos aislamientos no crecen a 5 o 37°C, pero lo hacen a 25°C. Pero en general en M.
anisopliae se encuentran amplios rangos de termotolerancia (Rangel et al. 2005; Fernandes et
al. 2008). La capacidad de crecer y ser infectivo a bajas o altas temperaturas está relacionado
con el área en donde sea encontrado el hongo De cross y Bidochka (1999) aislaron M.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
50
anisopliae en Canada, muchos de los cuales pudieron germinar a 8°C. Fernandes et al. (2008)
y Bidochchka (2006) sugieren que la capacidad de crecimiento de un hongo a diferentes
temperaturas está limitada por la temperatura promedio del sitio en donde se realizó el
aislamiento, los aislamientos evaluados en este estudio proceden de áreas con temperatura
promedio superior a 25°C lo que podría estar relacionado con la incapacidad de estos para
crecer a 10°C. Keyser et al. (2014) también reportaron que aislamientos de Metarhizium
sometidos a 5°C no mostraron ningún crecimiento, coincidente con Fernandes et al. (2010)
reportan que 37 aislamientos de Metarhizium spp. y M.a. var acridum incubados a 5°C durante
15 días, presentan grandes pérdidas en la germinación, y sólo en dos de la totalidad de
aislamientos el efecto de la temperatura no causa grandes pérdidas, igualmente exponerlos a
10°C durante 15 días manifiesta el efecto negativo de la temperatura sobre la germinación, ya
que hace que ésta sea menor. Sin embargo, Keyser et al. (2014) observaron que si los
aislamientos después de ser incubados a 5°C eran expuestos a 28°C su desarrollo volvía a ser
normal, lo que sugeriría que la temperatura induce un periodo de latencia en las conidias que
puede romperse una vez ésta vuelve a ser favorable para el hongo, este comportamiento no
puedo ser corroborado en este trabajo debido que las cajas de petri que contenían el medio de
cultivo en donde se había depositado la suspensión de los hongos eran descartadas una vez
se realizaban las mediciones necesarias y por esto no fueron sometidas a nuevos periodos de
incubación a temperaturas mayores. La mayoría de hongos entomopatógenos crecen a
temperaturas entre los 10°C y 40°C y por ello son clasificados como mesofílicos (Roberts y
Campbell, 1977). Según Walstad et al. (1970) la temperatura máxima en la que muchas
especies de M. anisopliae germinan es 37°C, sin embargo, existen variaciones importantes y
los aislamientos que crecen a altas temperaturas no tienen la capacidad de crecer a
temperaturas inferiores, lo que ha sido utilizado como uno de los criterios de clasificación del
género (Bischoff et al. 2006). Estos hallazgos coinciden con los hechos en este trabajo ya que
se observa que los aislamientos de Metarhizium aquí evaluados tienen tasas de crecimiento
superiores a las de Beauveria a 26.5°C y 30°C, por lo que se podría considerar que leves
aumentos en la temperatura de incubación favorecen su desarrollo. En cuanto a las
preferencias en temperatura Fargues et al. (1992) determinaron que M. anisopliae requiere
para su crecimiento micelial temperaturas entre 25 y 28°C, en este trabajo los aislamientos de
Metarhizium evaluados en las dos temperaturas en donde se presentó crecimiento 26.5°C y
30°C no muestran diferencias altamente contrastantes en cuanto al ABC, independientemente
del medio de cultivo. Berlanga y Hernández (2002) determinaron que M.a. var acridum tiene un

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
51
crecimiento más lento al ser incubado a 20°C, siendo más tolerante a altas temperaturas en
comparación con algunos aislamientos de M. anisopliae y B. bassiana, en este trabajo se
observó que hay un incremento de las ABC acumuladas cuando los hongos son incubados a
una temperatura mas alta (30°C).
Beauveria bassiana crece en un rango de temperatura entre 8°C y 35°C (Fargues et al. 1992).
El punto de muerte térmico para conidias de B. bassiana reportados por Liu et al. (2009) está
entre 45°C y 50°C incluso algunos aislamientos es cercana a 55°C (Varela y Morales, 1996).
Devi et al. (2005) evaluaron el efecto de la exposición a temperaturas entre 32°C y 42°C en
ciclos de 8 horas/ 16 horas a 25 °C sobre el crecimiento de 29 aislamientos de B. bassiana en
donde observaron que a 32°C no hay un efecto importante sobre la germinación y no se
encuentran diferencias con el testigo, al incrementar la temperatura de incubación a 35°C y
38°C comienzan a verse diferencias entre los aislamientos y su tolerancia. Estos autores
sugieren que la exposición a altas temperaturas por periodos cortos de tiempo seguidas de la
incubación a temperaturas óptimas favorece el desarrollo de los hongos. El rango de
temperatura evaluado en este trabajo es más estrecho que el evaluado por los anteriores
autores, sin embargo se evidencia un comportamiento similar en cuanto al crecimiento ya que
la gran mayoría crece en los rangos aquí evaluados, nueve de los 24 aislamientos evaluados
no muestran ningún tipo de crecimiento en alguno de los dos medios evaluados cuando son
incubados a 10°C lo que sugiere un rango de tolerancia menor al reportado por los anteriores
autores.
Con respecto a la capacidad patogénica, esta se puede ver afectada por la temperatura y el
medio en el que se desarrolle el hongo sin embargo, algunos hongos pueden permanecer
infectivos a bajas temperaturas incluso a 2°C (Vega et al. 2012). Se ha encontrado que
algunas variedades responden de manera diferencial al aumento de la temperatura,
Fernandes et al. (2010) reportan que en la medida que se incrementa el tiempo de exposición
a altas temperaturas (45°C), los aislamientos de Metarhizium spp. disminuyen su germinación
mientras que los aislamientos de M.a. var acridum no tienen grandes pérdidas en ella, los
aislamientros que este trabajo presentaron los rangos de crecimiento mas amplios muy
seguramente tendrán un mejor comportamiento en el caso dew que sean utilizados en el
campo.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
52
El aumento en la temperatura corporal de algunos insectos como mecanismo de defensa al
ataque de los hongos entomopatógenos restringe severamente el desarrollo del patógeno, lo
que conduce a un retraso considerable en la mortalidad inducida por estos organismos (Inglis
et al. 1997; Blanford y Thomas, 1999; Arthurs y Thomas, 2000; Blanford y Thomas, 2000;
Ouedraogo et al. 2004). Por ejemplo, el límite superior para el crecimiento de M.anisopliae var.
acridum está en el rango de 35-40 °C tales temperaturas corporales afectan el proceso
infectivo de hongos no tolerantes a las mismas.En condiciones de temperatura ambiental
consideradas ideales 25°C - 32 °C, el tiempo de la muerte de langostas después de ser
tratadas con M. anisopliae var. acridum es de 7 días (Lomer et al. 2001). Welling et al. (1994)
mencionan que aislamientos de M.anisopliae y M.a. var. acridum pueden resistir temperaturas
de 40 ºC o mayores durante algunas horas, pero con una marcada reducción en su grado de
crecimiento, indicando que el aislamiento de M.a. var. acridum fue más resistente a
temperaturas entre 44°C y 25 ºC. Lanza et al. (2009) evaluaron el efecto de la temperatura y
la humedad en tres tipos de suelo sobre la supervivencia de M. anisopliae, encontrando que la
supervivencia de los aislamientos de M. anisopliae evaluados se ve afectada a 31.5°C. pero
no entre 21°C y 26°C valores de temperatura que se encuentran en los evaluados en este
trabajo en donde los resultados fueron similares.
Las temperaturas a las que fueron sometidas los aislamientos evaluados en este trabajo
permiten evidenciar que de acuerdo a lo reportado por otros autores hay un efecto sobre la
tasa de crecimiento, y que la temperatura optima de crecimiento de cada género es diferente,
sin embargo, es evidente la capacidad de los aislamientos a crecer en un amplio rango de
temperaturas, lo que coincide con lo encontrado en la naturaleza. Al igual que la temperatura el
medio en el que se desarrolle un hongo tiene un efecto importante sobre la tasa de crecimiento
y la patogenidad de los hongos. Por lo anterior, es necesario realizar estudios específicos entre
otros con respecto al medio de cultivo o propagación y la temperatura de incubación con los
aislamientos candidatos a ser incorporados como estrategia de manejo de insectos plaga.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
53
1.6 Bibliografía
Arthurs, S.P. y Thomas, M.B. 2000. Effects of a mycoinsecticide on feeding and fecundity of the
brown locust Locustana pardalina. Biocontrol Science and Technology, 10, 321 – 329.
Arthurs S. y M. B. Thomas. 2001. Effect of dose, pre-mortem host incubation temperature y
thermal behavior on host mortality, mycosis y sporulation of Metarhizium anisopliae var. acridum
in Schistocera gregaria. Biocontrol Science Technology. 11: 411-420.
Berlanga-Padilla, Angelica M., Hernández-Velázquez Víctor M. 2002. Efecto de la temperatura
sobre el crecimiento y la virulencia de Metarhizium anisopliae, M. a. var. acridum y Beauveria
bassiana en Schistocerca piceifrons piceifrons. Manejo Integrado de Plagas (Costa Rica) 63: 5 1
- 5 5.
Bischoff, J., Rehner, S. A., Humber, R.A. 2006. Metarhizium frigidum sp. nov.: a cryptic species
of M. anisopliae and a member of the M. flavoviride complex. Mycologia, 98 (5): 737–745.
Blanford, S. y Thomas, M.B. 1999. Role of thermal biology in disease dynamics. Aspects of
Applied Biology, 53:73 – 82.
Blanford, S. y Thomas, M.B. 2000 .Thermal behaviour of two acridid species: effects of habitat
and season on body temperature and the potential impact on biocontrol with pathogens.
Environmental Entomology, 29: 1060 – 1069.
Devi K. U, Sridevi V., Murali M. Ch, Padmavathi J. 2005. Efect of high temperature and water
stress on in vitro germination and growth in isolates of the entomopathogenic fungus Beauveria
bassiana (Bals.) Vuillemin. Journal of Invertebrate Pathology 88: 181–189.
De Mendiburu, F. 2009. Una herramienta de análisis estadístico para la investigación agrícola.
Tesis. Universidad Nacional de Ingeniería (UNI-PERU).
De Croos, J. N. A., y Bidochka, M. J. 1999. Effects of low temperature on growth parameters in
the entomopathogenic fungus Metarhizium anisopliae. Can. J. Microbiol., 45.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
54
Inglis, G.D., Johnson, D.L., Cheng, K.-L. y Goette , M.S . 1997. Use of pathogen combinations to
overcome the constraints of temperature on entomophagous hyphomycetes against
grasshoppers. Biological Control, 8: 143 – 152.
Iskandarov, U., Guzalova, A., y Davranov, K. 2006. Effects of Nutrient Medium Composition and
Temperature on the Germination of Conidia and the Entomopathogenic Activity of the Fungi
Beauveria bassiana and Metarhizium anisopliae. Applied Biochemistry and Microbiology. Rusia.
42(1): 72-76.
Fargues, J., N. K. Maniania, J. C. Delmas, y N. Smits. 1992. Influence de température sur la
croissance in vitro d'hyphomycètes entomopathogènes, Agronomie 12: 557–564.
Fernandes E., Keyser C. A., Rangel D., Foster R. N. y Roberts D. w. 2010. CTC medium: A
novel dodine-free selective medium for isolating entomopathogenic fungi, especially Metarhizium
acridum, from soil. Biological Control 54: 197–205.
Ferreira, A.S., Totola M.R. y Borges, A.C. 2007. Physiological implications of trehalose in the
ectomycorrhizal fungus Pisolithus sp. under thermal stress. Journal of Thermal Biology 32: 34-
41.
Ferron, P. 1978. Biological control of insect pests by entomogenous fungi. Annual Review of
Entomology 23:409-442.
Keyser C. A., Fernandes K.K., Rangel D. E.N., Roberts Donald W. 2014. Heat-induced post-
stress growth delay: A biological trait of many Metarhizium isolates reducing biocontrol
efficacy?. Journal of Invertebrate Pathology 120: 67–73.
Lanza L.M., Monteiro A.C., Malheiros E.B., .2009. Sensibilidade de Metarhizium anisopliae à
temperatura e umidade em três tipos de solos Sensibility of Metarhizium anisopliae to
temperature and moisture in three soil types. Ciência Rural, 39(1): 6-12.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
55
Lomer, C. J., Bateman, R. P., Johnson, D. L., Langewald, J. y Thomas, M. 2001. Biological
control of locust and grasshoppers. Annu. Rev. Entomol. (46): 667-702.
Liu, Q., Ying. S.H., Feng, M.G y Jiang XH. 2009. Physiological implication of intracellular
trehalose and mannitol changes in response of entomopathogenic fungus Beauveria bassiana to
thermal stress. Antonie van Leeuwenhoek 95: 65–75.
McCammon, S.A. y Rath, A.C. 1994: Separation of Metarhizium anisopliae strains by
temperature dependent germination rates. Mycol. Res. 98: 1253-1257.
Ouedraogo, R.M., Goettel, M.S. y Brodeur, J. 2004. Behavioral thermoregulation in the migratory
locust: a therapy to overcome fungal infection. Oecologia 138: 312 – 319.
Rangel D.E.N, Anderson A.J y Roberts D.W. 2008. Evaluating physical and nutritional stress
during mycelial growth as inducers of tolerance to heat and UV-B radiation in Metarhizium
anisopliae conidia. Mycol Res 112: 1362–1372.
Rangel, D.E.N., Fernandes, E.K.K., Braga, G.U.L., Roberts, D.W., 2011. Visible light during
mycelial growth and conidiation of Metarhizium robertsii produces conidia with increased stress
tolerance. FEMS Microbiol. Lett. 315: 81–86.
Roberts, D.W. y Campbell, A.S. 1977. Stability of entomopathogenic fungi. Misc Publ
Entomol Soc Am 10:19–76.
Soares, G. G., Marchal, M. y Ferron, P. 1983. Susceptibility of Otiorhynchus sulcatus
(Coleoptera: Curculionidae) larvae to Metarhizium anisopliae and Metarhizium flavoviridae
(Deuteromycotina: Hyphomycetes) at two different temperatures. Environmental Entomology
12:1887-1891.
Varela, A. y Morales, E. 1996. Characterization of some Beauveria bassiana isolates and their
virulence toward the coffee berry borer hypothenemus hampei Journal of invertebrate pathology
67 (2):147–152.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
56
Vega, F., Meyling, N., Luangsa-ard, J., Blackwell, M. 2012. Fungal Entomopathogens. En: Vega,
F. y Kaya, H. Insect Pathology (Second Edition). p. 172-206.
Walstad, J.D., Anderson, R.F. y Stambaugh, W.J. 1970. Effects of environmental conditions on
two species of muscardine fungi (Beauveria bassiana and Metarhizium anisopliae). Journal
Invertebrate Pathology 16: 221–226. 39
Welling M., Nachtigall G. y Zimmermann G. 1994. Metarhizium Spp, isolates from Madagascar:
Morphology and effect of high temperature on growth and infectivity to the migratory locust,
locusta migratoria. Entomophaga 39 (3/4): 351-361.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
57
CAPITULO 2: TOLERANCIA DE LOS HONGOS ENTOMOPATÓGENOS
A LA RADIACIÓN ULTRAVIOLETA
2.1 Introducción
La radiación consiste en la propagación de energía en forma de ondas electromagnéticas o
partículas subatómicas a través del vacío o de un medio material. La energía requerida para
los procesos físicos y biológicos terrestres depende en casi un 99% de la radiación proveniente
del sol, la cual es de tipo electromagnética, y va desde los 15 a 400 nm (radiación ultravioleta -
UV-). La radiación solar afecta a los hongos en condiciones de campo a través de dos
procesos diferentes: el efecto fotónico, causado por longitudes de onda corta (UV y fotones
visibles) y el efecto térmico causado por la radiación infrarroja cercana (700 – 3000 nm), lo
cual resulta en un incremento de la temperatura (Pavone, 2003).
La radiación responsable del efecto fotónico UV se divide en tres tipos: Radiación Ultravioleta
A (UV-A) entre 315- 400 nm; Radiación Ultravioleta B (UV-B) entre 280-315 nm y Radiación
Ultravioleta C (U\/- C) entre 200-280 nm (Devotto y Gerding, 2003; Paul y Gwynn-Jones,
2003). Aunque esta clasificación es arbitraria, se hace útil para considerar los efectos
biológicos y ecológicos de la UV. La energía de un fotón de radiación es inversamente
proporcional a su longitud de onda, y por lo tanto la radiación UV-C, es la más energética de la
tres bandas de frecuencia, por tener la menor longitud de onda (Paul y Gwynn-Jones, 2003).
La UV-C a pesar de ser la más nociva, es fuertemente absorbida por el oxígeno y el ozono en
la estratosfera. La UV-A y UV-B son capaces de atravesar la atmósfera terrestre, la UV-A no
es absorbida por el ozono y llega en su totalidad a la tierra, la radiación UV-B aunque es
absorbida en un 90% por la capa de ozono y solo el 10% logra ingresar a la litosfera es
nociva ya que afecta las moléculas de ADN induciendo reacciones fotoquímicas que llevan a
mutaciones.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
58
2.1.1 Efecto de la RUV-B sobre los hongos
Los hongos al igual que todos los demás organismos vivos son sensibles a la luz solar, por ello
son afectados luego de una exposición prolongada especialmente al componente UV-B de
ésta (Alves et al. 1998, Braga et al. 2001; 2002; Fernandes et al. 2007). La radiación
ultravioleta (UV) (entre 200 y 400 nm) es una causa importante de la disminución de la
viabilidad del inóculo de hongos aplicado en campo debido a los efectos fisiioloicos que tiene
sobre las conidias, este tipo de radiación varía en intensidad con el tiempo del día, el área
geográfica y la estación del año (Huang y Feng, 2009). Para los organismos expuestos la UV
impone la selección intensa, resultando en cambios en las especies y en la composición de
estas en una comunidad en escalas de tiempo incluso de horas (Jacobs y Sunding, 2001). Por
lo anterior, la tolerancia a la radiación UV es una característica importante en hongos y
bacterias, ya que durante una parte de su ciclo de vida están expuestos a esta condición,
siendo especialmente vulnerables durante procesos como la esporulación, dispersión y el
proceso de infección en sus hospederos.
Según Diffey, 1991 y Griffiths et al. 1998 el efecto que tiene la UV sobre los microorganismos,
se debe a la presencia de grupos cromóforos situados en diferentes estructuras celulares,
estos son capaces de absorber ciertas longitudes de onda, que a su vez pueden ocasionar
daños directos o indirectos sobre algunas moléculas (p. ej.; carotenoides, esteroides,
quinonas, proteínas y ácidos nucleicos entre otros). Paul y Gwynn-Jones (2003) mencionan
que el daño en el Acido Desoxirribucleico ADN es probablemente el principal efecto de la UV
induciendo cambios heredables traducidos en variación genética. En el ADN la radiación UV-C
y UV-B forma dímeros de bases pirimidinas sobre la misma cadena tipo timina- timina, timina -
citosina y citosina-citosina (Nicholson et al. 2005), además, se da la formación de hidratos de
pirimidina y entrecruzamientos entre ADN y proteínas (Devotto y Gerding, 2003). Los daños
indirectos de la radiación sobre la célula se deben principalmente a la formación de peróxido
de hidrógeno y radicales libres que oxidan la pentosa presente en el ADN rompiendo la hebra
de la molécula; en general los daños ocasionados se reflejan en mutaciones, retraso en el
crecimiento o muerte celular (Braga et al. 2001a). La UV también causa daños a proteínas y
peroxidación de lípidos que reaccionan con los ácidos grasos de los fosfolípidos, afectando la
fluidez de la membrana celular (Pavone, 2003; Gutteridge y Halliwell, 1990; Aikens y Dix,
1991). El ADN mitocondrial también puede sufrir alteraciones (Krutmann, 2006).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
59
En términos de efectos en poblaciones, se considera que el aumento de radiación UV-B puede
cambiar no sólo los individuos, sino también la estructura de las comunidades, considerado
este uno de los factores que reducen la eficiencia de los agentes de control biológico (Johnson,
2003). Durante la aplicación en el día de las conidias de hongos, la principal preocupación es
la 'inactivación' debida a la UV. Ignoffo et al. (1977) propusieron al peróxido producido por la
fotooxidación de los aminoácidos debido a UV-A y UV-B como el responsable de la falta de
efectividad de un hongo entomopatógeno sometido a UV. Por ello en la actualidad se emplean
protectores de luz UV y se seleccionan aislamientos basados en su tolerancia a la radiación,
todo esto con el fin de conservar su efecto en campo.
El presente estudio se realiza una prueba de la tolerancia a luz UV-B en aislamientos de
Beauveria y Metarhizium procedentes de insectos plaga asociados al cultivo de la palma de
aceite.
2.2 Objetivo Específico
Objetivo específico 2: Establecer el efecto de la luz ultravioleta, fracción UV-B de la luz solar,
sobre la germinación de aislamientos de Beauveria y Metarhizium .

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
60
2.3 Metodología
2.3.1 Aislamientos utilizados en el estudio
Se evaluaron 31 aislamientos de hongos entomopatógenos, 29 de Beauveria sp. y siete de
Metarhizium (Anexo 1), estos en la actualidad pertenecen a la colección de hongos
entomopatógenos de la Corporación Centro de Investigación en Palma de Aceite (Cenipalma).
Los aislamientos fueron mantenidos en Agar Sabouraud Dextrosa (SDA) Oxoid® (composición
g.L-1: peptona micológica 10, glucosa 40, agar 15) y multiplicados según necesidad.
2.3.2 Aplicación de tratamiento con UV-B.
Se evaluó el efecto de la radiación ultravioleta (UV-B) sobre el porcentaje de germinación al
exponer los hongos a esta radiación durante diferentes tiempos: 0, 15, 30, 45 y 60 minutos.
Para ello, se tomaron conidias de cada uno de los aislamientos a evaluar y se se preparó una
suspensión en 10 ml de agua destilada estéril + Tween 80 al 0,1%, se homogenizó la
suspensión y se ajustó la concentración a 1 x 10 7conidias.ml-1. Luego, en cajas de Petri que
contenían SDA se depositó 1 µl de suspensión de cada aislamiento sobre cinco puntos
diferentes del medio de cultivo, lo que constituyó una unidad experimental. Las unidades
experimentales se expusieron a radiación UV-B en una cámara de flujo laminar con una
lámpara General Electric® que produce una radiación de 300 nm ubicando las cajas de Petri a
una distancia de 30 cm de la lámpara. Luego de someter cada unidad experimental a los
tratamientos, éstas se llevaron a la incubadora por 24 horas a 26.5 °C en oscuridad total para
evitar efectos de fotoreparación. Los testigos (no expuestos a UVB) se dispusieron igualmente
en la incubadora en similares condiciones.
Para cada aislamiento se realizaron cuatro repeticiones por cada uno de los tiempos de
exposición. La variable evaluada fue el porcentaje de germinación como indicador del número
de conidias que sobrevivieron a la exposición a la radiación. Una vez se cumplió el tiempo de
incubación, se realizó el conteo de conidias germinadas y no germinadas utilizando el objetivo
40X en un microscopio marca Olympus® CX21.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
61
2.3.4 Cálculo de porcentaje de germinación.
Se consideró que una conidia estaba germinada cuando el tubo germinal observado es como
mínimo el doble del diámetro de la conidia. La determinación del porcentaje de germinación se
hizo mediante la utilización de la siguiente fórmula:
2.3.5 Análisis estadístico
Para el análisis estadístico mediante Tukey, se agruparon las medias de cada uno de los
aislamientos en función del tiempo de exposición a RUV-B con un nivel de significancia del
95%; en todos los casos, se cumplieron con los supuestos de normalidad y homocedasticidad
(valor p > 0.01).
2.4 Resultados
2.4.1 Efecto de la UV-B sobre aislamientos de Beauveria
En general, para todos los aislamientos de Beauveria evaluados se oberva que en la medida
que aumenta el tiempo de exposición a la UV-B, disminuye el porcentaje de germinación de
las conidias con respecto a su testigo, este es mayor y difiere significativamente en la medida
que aumenta el tiempo de exposición a 30, 45 y 60 minutos (Tabla 2.1). Con respecto al
menor tiempo de exposición evaluado 15 minutos, en la mayoría de aislamientos de Beauveria
aunque las medias de germinación varían no se observan diferencias significativas entre 0
minutos (testigo) y 15 minutos. Algunos aislamientos como B028 y B042 presentaron las
mayores diferencias entre el tiempo 0 y 15 minutos. B028 al tiempo 0 tuvo un porcentaje de
germinación del 95.06% y luego de 15 minutos de exposición el valor fue 72.71%, para B042
a los 0 minutos de exposición la media de germinación fue 92.35% y después de 15 minutos
de exposición fue 74.82%. En B007 aunque la media del porcentaje de germinación es mayor

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
62
a los 15 minutos de exposición a UV-B (90.25%), con respecto a 0 minutos de exposición
(testigo) (89.37%) estos valores no tienen diferencia estadística significativa. En los primeros
15 minutos de exposición a UV-B entre los aislamientos que no muestran diferencias
significativas al compararlos con el tiempo 0 se presentan disminuciones porcentuales de la
germinación en rangos que van desde -0.98 (B007) a 16.74 (B019) (Tabla 2.2).
Al comparar los porcentajes de germinación de los aislamientos cuando son expuestos por 15
y 30 minutos a UV-B a excepción de B006, B019, B035 y B036 que no muestran diferencias
significativas entre estos dos tiempos de exposición, los demás aislamientos si las presentan
lo que indica que es después de este tiempo de exposición cuando se empieza a evidenciar el
efecto que sobre la germinación de las conidias tiene la UV-B. La disminución en el porcentaje
de germinación después de 30 minutos de exposición con respecto al testigo oscila entre el
22.73% para B002 y 37.61% para B042 superiores a los porcentajes de disminución al ser
expuestos durante 15 minutos a UV-B (Tabla 2.2). Al comparar las medias de germinación de
los aislamientos a los 30 y 45 minutos de exposición únicamente B036 muestra diferencias
estadísticamente significativas entre estos dos tiempos de exposición sin embargo, esto no
sugiere que este aislamiento tenga una menor tolerancia a la UV-B, ya que al observar las
medias de disminución en el porcentaje de germinación de las conidias después de 45
minutos de exposición con respecto al tiempo 0 algunos aislamientos como B042, B041, B035
tienen medias de disminución del porcentaje de germinación de 46.92%, 45.17% y 45.20%
respectivamente, superiores a B036 cuya valor de disminución del porcentaje de germinación
de las conidias fue 40.17% (Tabla 2.2).
Después de 60 minutos de exposición a UV-B todos los aislamientos tienen pérdidas en la
germinación y muestran diferencias estadísticamente significativas con respecto a las conidias
no expuestas a radiación. Al realizar la comparación entre los valores medios de germinación
a 45 y 60 minutos de exposición únicamente los aislamientos B002, B013, B016, B025, B028,
B038 y B044 muestran diferencias significativas (Tabla 2.2). El mayor porcentaje de
disminución de la germinación al compararlo con la presentada al tiempo 0 después de 60
minutos de exposición a UV-B se presentó en B025 60.52%, los demás aislamientos tienen
porcentajes de disminución de la germinación en un rango entre 57.53% y 38.79% siendo
B001 el que menor porcentaje de pérdida presentó 38.79% (Tabla 2.2). B036, se caracterizó
por presentar un mayor porcentaje de germinación cuando se expuso a 60 minutos de UV-B,
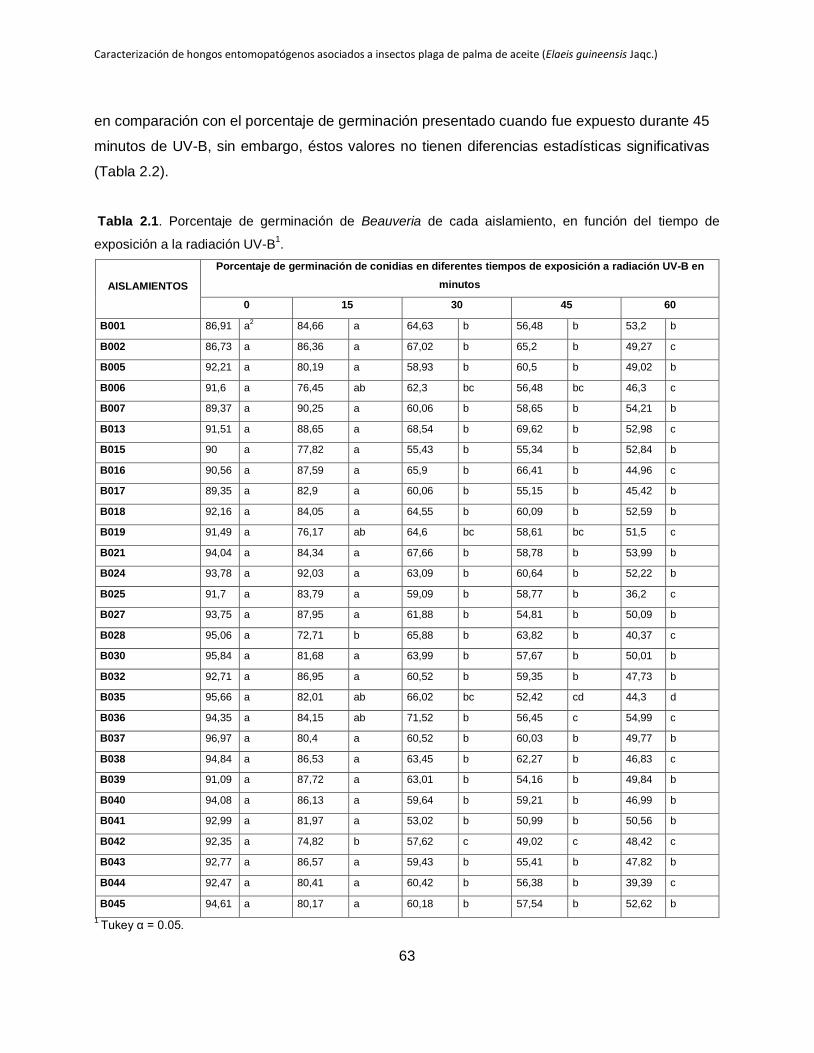
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
63
en comparación con el porcentaje de germinación presentado cuando fue expuesto durante 45
minutos de UV-B, sin embargo, éstos valores no tienen diferencias estadísticas significativas
(Tabla 2.2).
Tabla 2.1. Porcentaje de germinación de Beauveria de cada aislamiento, en función del tiempo de
exposición a la radiación UV-B1.
AISLAMIENTOS
Porcentaje de germinación de conidias en diferentes tiempos de exposición a radiación UV-B en
minutos
0 15 30 45 60
B001 86,91 a2
84,66 a 64,63 b 56,48 b 53,2 b
B002 86,73 a 86,36 a 67,02 b 65,2 b 49,27 c
B005 92,21 a 80,19 a 58,93 b 60,5 b 49,02 b
B006 91,6 a 76,45 ab 62,3 bc 56,48 bc 46,3 c
B007 89,37 a 90,25 a 60,06 b 58,65 b 54,21 b
B013 91,51 a 88,65 a 68,54 b 69,62 b 52,98 c
B015 90 a 77,82 a 55,43 b 55,34 b 52,84 b
B016 90,56 a 87,59 a 65,9 b 66,41 b 44,96 c
B017 89,35 a 82,9 a 60,06 b 55,15 b 45,42 b
B018 92,16 a 84,05 a 64,55 b 60,09 b 52,59 b
B019 91,49 a 76,17 ab 64,6 bc 58,61 bc 51,5 c
B021 94,04 a 84,34 a 67,66 b 58,78 b 53,99 b
B024 93,78 a 92,03 a 63,09 b 60,64 b 52,22 b
B025 91,7 a 83,79 a 59,09 b 58,77 b 36,2 c
B027 93,75 a 87,95 a 61,88 b 54,81 b 50,09 b
B028 95,06 a 72,71 b 65,88 b 63,82 b 40,37 c
B030 95,84 a 81,68 a 63,99 b 57,67 b 50,01 b
B032 92,71 a 86,95 a 60,52 b 59,35 b 47,73 b
B035 95,66 a 82,01 ab 66,02 bc 52,42 cd 44,3 d
B036 94,35 a 84,15 ab 71,52 b 56,45 c 54,99 c
B037 96,97 a 80,4 a 60,52 b 60,03 b 49,77 b
B038 94,84 a 86,53 a 63,45 b 62,27 b 46,83 c
B039 91,09 a 87,72 a 63,01 b 54,16 b 49,84 b
B040 94,08 a 86,13 a 59,64 b 59,21 b 46,99 b
B041 92,99 a 81,97 a 53,02 b 50,99 b 50,56 b
B042 92,35 a 74,82 b 57,62 c 49,02 c 48,42 c
B043 92,77 a 86,57 a 59,43 b 55,41 b 47,82 b
B044 92,47 a 80,41 a 60,42 b 56,38 b 39,39 c
B045 94,61 a 80,17 a 60,18 b 57,54 b 52,62 b
1 Tukey α = 0.05.
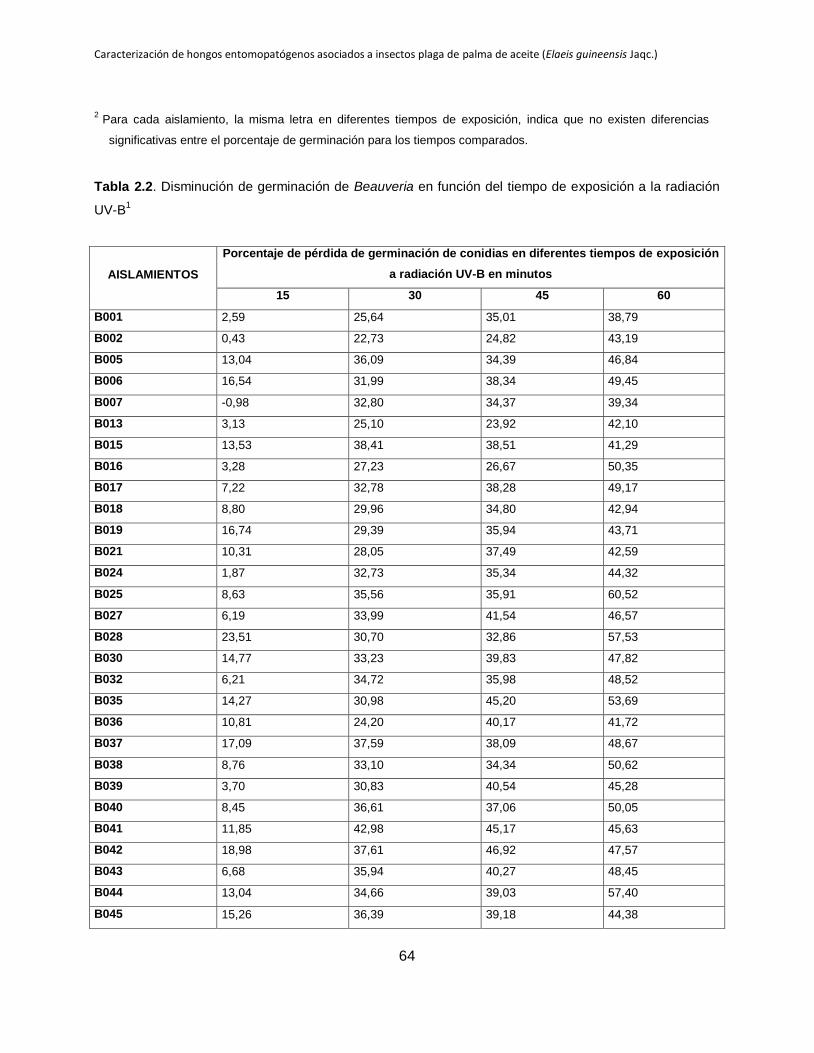
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
64
2 Para cada aislamiento, la misma letra en diferentes tiempos de exposición, indica que no existen diferencias
significativas entre el porcentaje de germinación para los tiempos comparados.
Tabla 2.2. Disminución de germinación de Beauveria en función del tiempo de exposición a la radiación
UV-B1
AISLAMIENTOS
Porcentaje de pérdida de germinación de conidias en diferentes tiempos de exposición
a radiación UV-B en minutos
15 30 45 60
B001 2,59 25,64 35,01 38,79
B002 0,43 22,73 24,82 43,19
B005 13,04 36,09 34,39 46,84
B006 16,54 31,99 38,34 49,45
B007 -0,98 32,80 34,37 39,34
B013 3,13 25,10 23,92 42,10
B015 13,53 38,41 38,51 41,29
B016 3,28 27,23 26,67 50,35
B017 7,22 32,78 38,28 49,17
B018 8,80 29,96 34,80 42,94
B019 16,74 29,39 35,94 43,71
B021 10,31 28,05 37,49 42,59
B024 1,87 32,73 35,34 44,32
B025 8,63 35,56 35,91 60,52
B027 6,19 33,99 41,54 46,57
B028 23,51 30,70 32,86 57,53
B030 14,77 33,23 39,83 47,82
B032 6,21 34,72 35,98 48,52
B035 14,27 30,98 45,20 53,69
B036 10,81 24,20 40,17 41,72
B037 17,09 37,59 38,09 48,67
B038 8,76 33,10 34,34 50,62
B039 3,70 30,83 40,54 45,28
B040 8,45 36,61 37,06 50,05
B041 11,85 42,98 45,17 45,63
B042 18,98 37,61 46,92 47,57
B043 6,68 35,94 40,27 48,45
B044 13,04 34,66 39,03 57,40
B045 15,26 36,39 39,18 44,38

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
65
2.4.2 Efecto de la UV-B sobre aislamientos de Metarhizium
Las conidias de Metarhizium presentan la misma tendencia de pérdida de la germinación que
las de los aislamientos de Beauveria cuando se incrementa el tiempo de exposición a UV. Las
medias del porcentaje de germinación de cada uno de los hongos después de ser expuestos a
la UV durante 0 y 15 minutos, aunque muestran una disminución en el porcentaje de
germinación no tienen diferencias significativas en estos dos tiempos, a excepción de Mt010
(Tabla 2.3). Se observa en los aislamientos expuestos a UV-B durante 15 minutos, que Mt010
muestra el mayor porcentaje de pérdida de germinación del 31.07% con respecto al testigo,
los demás aislamientos Mt004; Mt001; Mt006 y Mt009 muestran disminuciones en
germinación del 16.29%; 9.96%; 9.36% y 8.39% respectivamente. En Mt002 y Mt005 la
disminución en la germinación a los 15 minutos de exposición con respecto a la presentada en
el testigo no supera el 3% (Tabla 2.4). Cuando aumenta el tiempo de exposición a UV-B a 30
minutos todos los tratamientos, a excepción de Mt001 muestran diferencias significativas al
compararlas con el testigo, se observa una disminución generalizada en las medias de los
porcentajes de germinación (Tabla 2.3). El comportamiento de los aislamientos expuestos 45
minutos a la radiación UV-B es similar al presentado cuando fueron expuestos a la misma
condición 30 minutos, y en ninguno se presentan diferencias significativas con respecto al
tiempo de exposición inmediatamente anterior. Después de 60 minutos de exposición, los
aislamientos de Metarhizium evaluados tienen una disminución de mínimo el 39.55% (Mt001)
y máximo el 57.33% (Mt010) en la media del porcentaje de germinación respecto al alcanzado
cuando los aislamientos no son expuestos a UV-B. En este tiempo de evaluación no se
presentan diferencias estadísticas de los aislamientos con respecto a 45 minutos de
exposición. El aislamiento Mt005, se caracterizó porque el porcentaje de germinación fue
mayor cuando el aislamiento se expuso a 45 minutos de UV-B, en comparación con el
porcentaje de germinación obtenido cuando éste se expuso a 30 minutos de UV-B, sin
embargo estadísticamente no se presentan diferencias. En los aislamientos Mt002, Mt005 y
Mt009 no se encontraron diferencias estadísticas significativas después de 30 y hasta 60
minutos de exposición a UV-B.
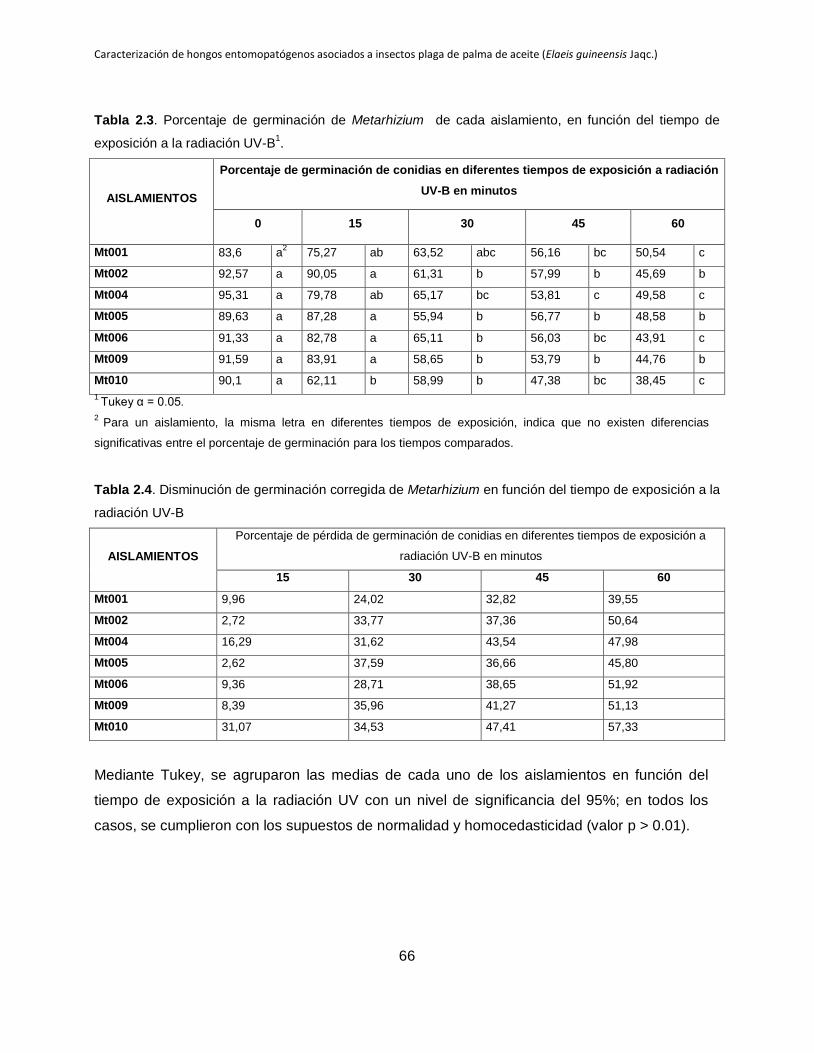
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
66
Tabla 2.3. Porcentaje de germinación de Metarhizium de cada aislamiento, en función del tiempo de
exposición a la radiación UV-B1.
AISLAMIENTOS
Porcentaje de germinación de conidias en diferentes tiempos de exposición a radiación
UV-B en minutos
0 15 30 45 60
Mt001 83,6 a2
75,27 ab 63,52 abc 56,16 bc 50,54 c
Mt002 92,57 a 90,05 a 61,31 b 57,99 b 45,69 b
Mt004 95,31 a 79,78 ab 65,17 bc 53,81 c 49,58 c
Mt005 89,63 a 87,28 a 55,94 b 56,77 b 48,58 b
Mt006 91,33 a 82,78 a 65,11 b 56,03 bc 43,91 c
Mt009 91,59 a 83,91 a 58,65 b 53,79 b 44,76 b
Mt010 90,1 a 62,11 b 58,99 b 47,38 bc 38,45 c
1 Tukey α = 0.05.
2 Para un aislamiento, la misma letra en diferentes tiempos de exposición, indica que no existen diferencias
significativas entre el porcentaje de germinación para los tiempos comparados.
Tabla 2.4. Disminución de germinación corregida de Metarhizium en función del tiempo de exposición a la
radiación UV-B
AISLAMIENTOS
Porcentaje de pérdida de germinación de conidias en diferentes tiempos de exposición a
radiación UV-B en minutos
15 30 45 60
Mt001 9,96 24,02 32,82 39,55
Mt002 2,72 33,77 37,36 50,64
Mt004 16,29 31,62 43,54 47,98
Mt005 2,62 37,59 36,66 45,80
Mt006 9,36 28,71 38,65 51,92
Mt009 8,39 35,96 41,27 51,13
Mt010 31,07 34,53 47,41 57,33
Mediante Tukey, se agruparon las medias de cada uno de los aislamientos en función del
tiempo de exposición a la radiación UV con un nivel de significancia del 95%; en todos los
casos, se cumplieron con los supuestos de normalidad y homocedasticidad (valor p > 0.01).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
67
2.5 Discusión
Los resultados del efecto de la UV-B sobre la germinación de hongos de los géneros
Beauveria y Metarhizium encontrados este trabajo coinciden con lo reportado por autores
como Long et al. 2013 y Fernandes et al. 2007 en donde al exponer conidias de diferentes
aislamientos de los géneros Beauveria y Metarhizium a UV-B registran una disminución en la
tasa de germinación, la cual se comporta inversamente proporcional al tiempo de exposición.
Fargues et al. (1997), determinaron la influencia de la luz solar artificial en la supervivencia de
esporas de una gran cantidad de aislamientos de 4 especies de hongos entomopatógenos (B.
bassiana, M. anisopliae, M. flavoviridae y Paecilomyces fumosoroseus) al irradiar estos
hongos con luz solar artificial durante 0, 1, 2, 4 y 8 horas, la supervivencia de todos los
aislamientos disminuyó con el aumento del tiempo de exposición, siendo la exposición de dos
horas la más perjudicial para todos los aislamientos. Generalmente, una hora de exposición a
rayos UV-B en irradiancias similares que se encuentran en la naturaleza no perjudica
notablemente conidias de hongos entomopatógenos, pero después de dos horas, una mayor
variabilidad en la tolerancia a la radiación UV-B se encuentra entre las especies y cepas
(Braga et al. 2001a; Fargues et al. 1996; Rangel et al. 2006). Con respecto a los resultados
anteriores en esta investigación, se observó que después de 30 minutos de exposición a UV-B
las conidias de Beauveria tienen una disminución de mínimo el 20% de germinación con
respecto al testigo en los aislamientos de Metarhizium evaluados, la situación es similar,
inclusive Mt010, pierde el 31.07% de germinación con respecto al testigo no irradiado después
de tan solo 15 minutos de exposición a UV-B, lo que sugiere que esta tolerancia es especifica
por aislamiento y que es después de aumentar el tiempo de exposición que realmente se
evidencia la susceptibilidad o tolerancia de cada aislamiento La vida media de un
entomopatógeno en condiciones de campo es estimada en menos de una hora para el más
susceptible y de cerca de 80 horas para el más resistente, por ejemplo en Noumarea rileyi, se
reporta hasta un máximo de 60 horas de supervivencia (Pavone, 2003). Velez y Montoya,
1995 sostienen que la vida media de los diferentes tipos de inóculo de hongos expuestos a la
luz solar es de un tiempo aproximado de 1 hora para el hongo entomopatógeno más
susceptible y de 96 horas para el más resistente de los evaluados por estos investigadores.
Los resultados hallados en este trabajo hacen pensar que las pérdidas en campo puedan ser
similares, más aun cuando allí se conjugan además de la radiación, la humedad relativa, la
desecación, la precipitación y la competencia con otros microorganismos presentes en el

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
68
ambiente que pueden afectan su desempeño ya que, aumentos en los tiempos de
germinación o la disminución de esta por efecto de la radiación en campo, aumenta la
posibilidad de que los insectos generen respuesta inmune de defensa al ataque de los
hongos.
Fernandes et al. (2007), evaluaron el efecto de la UV-B sobre aislamientos de B. bassiana,
Beauveria spp. y Engyodontium album (B. alba) encontrando una alta variabilidad a la
tolerancia a la exposición a la UV con valores entre 0% y 80% después de ser expuestas
durante dos horas a la radiación. El rango de disminución de la germinación de los
aislamientos evaluados en el presente trabajo con respecto a cada uno de los testigos
después de 60 minutos de exposición a UV-B, fue de 38.78% (B001) a 60.25% (B025) lo que
sugiere un rango más estrecho de tolerancia en estos aislamientos que los reportados por
estos autores. Loong et al. 2013 encontraron que algunos aislamientos de M. anisopliae y P.
fumosoroseus disminuyen su germinación hasta en un 50% después de una hora de
exposición a UV-B valor que va aumentando en la medida que se incrementa el tiempo de
exposición, estos resultados concuerdan con los encontrados para los aislamientos de
Metarhizium, que después de una hora de estar expuestos a UV-B disminuyen entre el
39.55% y el 57.33% su germinación. Posadas et al. 2012 describen una alta tolerancia natural
en los aislamientos de B. bassiana expuestos a UV-B incluso reportan que uno de los
aislamientos tiene una germinación del 78.5% después de exponer sus conidias a UV-B
durante cuatro horas, todos los aislamientos evaluados por estos autores después de 60
minutos de exposición a UV-B muestran porcentajes de germinación con respecto al testigo
superiores al 80 %, a pesar de que las condiciones de distancia en que la lámpara fue ubicada
con respecto a las cajas de Petri y las longitudes de onda de las lámparas utilizadas son
similares a las utilizadas en estos ensayos, sus resultados son contrastantes con los hallados
en este trabajo en donde ninguno de los aislamientos evaluados tiene porcentajes de
germinación superiores al 55% después de 60 minutos de exposición a UV-B, posiblemente el
medio de cultivo utilizado por Posadas et al. 2012, favorece el desarrollo de tolerancia a la UV-
B. A este respecto, Rangel et al. 2006 reportan que las conidias de M. anisopliae producidas
en medios que causen estrés al hongo, tienen al menos 2 veces más tolerancia a la radiación
UV-B que las producidas en medios enriquecidos, estos autores indican que posiblemente la
energía que se gasta en producir enzimas para fuentes no preferidas de carbono generan
conidias más tolerantes a la UV-B, en este trabajo se utilizaron SDA y PDA que proporcionan

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
69
las fuentes de carbono y nitrógeno para el crecimiento de estos hongos, por lo anterior es
pertinente evaluar el comportamiento de los aislamientos seleccionados por este y otros
parámetros en medios diferentes para medir si hay un incremento en la tolerancia a UV-B.
También la tolerancia de los hongos a la radiación UV-B y a las altas o bajas temperaturas
puede estar asociada con el lugar en donde se realiza el aislamiento del hongo ya que la
radiación aumenta con la disminución de la latitud y el incremento de la altitud (Fernandes et al.
2008; Rangel et al. 2005). En general, pocas horas de exposición directa a la radicación solar
en el trópico son suficientes para inactivar las conidias de la mayoría de hongos
entomopatógenos (Fernandes et al. 2007) a este respecto, Braga et al. 2001 afirman que los
aislamientos encontrados cerca del ecuador son más tolerantes a la radiación UV-B que los
encontrados en las latitudes superiores a 40° norte o sur de la línea ecuatorial. Fernandes et
al. 2007 observaron amplios rangos de tolerancia de entre 16% y 80% en aislamientos de
Beauveria sometidos a UV-B durante dos horas, siendo los aislamientos colectados en
diferentes locaciones de Brasil entre los 0° y 28° de latitud sur, los más tolerantes a la radiación,
lo que sugiere una adaptación latitudinal a la radiación, los aislamientos evaluados en este
trabajo han sido encontrados en zonas palmeras del territorio Colombiano ubicadas entre 12°
latitud norte y 4° latitud sur, por lo que se supondría que estos aislamientos tengan una
tolerancia relativamente alta a la UV, sin embargo, la disminución en la germinación de las
conidias para los aislamientos más tolerantes fue de 38.79% (Beauveria) y 39.55%
(Metarhizium ) después de tan solo 1 hora de exposición, lo que indica que estos tienen una
baja tolerancia a UV-B , los hallazgos realizados contrastan con los resultados de Mustafa y
Kaur, 2009; Fernandes et al. 2007 y Braga et al. 2001; quienes evaluaron diferentes
aislamientos de Beauveria y Metarhizium encontrados en latitudes ubicadas en la zona cálida
de la tierra entre los Trópicos de Cáncer y Trópico de Capricornio, encontrando que estos
aislamientos no tenían disminuciones importantes en el porcentaje de germinación de las
conidias después de su exposición a UV-B. Lo anterior sugiere que la latitud no es el único
factor que afecta la adaptación a la UV-B otros factores como la humedad atmosférica, la
inclinación del sol, el ecosistema e incluso el insecto hospedero influyen en esta característica
(Bidochka et al. 2001; Brooks et al. 2004; Polar et al. 2005).
Gahjar et al. (2006) determinaron que la exposición de conidias de Plectosporium alismatis a
UV-B por 15 o 30 minutos únicamente retrasa la germinación, por el contrario la exposición

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
70
por más de 60 minutos si causa muerte a las conidias, los resultados obtenidos en este
trabajo en los aislamientos de Beauveria y Metarhizium muestran pérdidas porcentuales en la
germinación de las conidias con valores que oscilan entre 33.71% y 55.50% al comparar la
media de germinación a 0 y a 60 minutos de exposición, probablemente la media de
germinación obtenida en los aislamientos después de 60 minutos de exposición a la UV-B
varía si el efecto del tratamiento fuera sobre la velocidad de germinación y no sobre la
supervivencia de las conidias.
Cepas tolerantes a la UV, emplean mecanismos de defensa tales como la secreción de
pigmentos de absorción (Hullo et al. 2001; Saxene et al. 2002). Braga et al. 2006 determinaron
que la pigmentación de las conidias de M. anisopliae confieren a este hongo una mayor
tolerancia a la radicación UV, sin embargo, al comparar la tolerancia de los aislamientos de
Beauveria y Metarhizium evaluados en este trabajo, el comportamiento es similar y no se
puede sugerir una mayor tolerancia de Metarhizium ya que entre los aislamientos de
Beauveria algunos muestran disminución en la germinación menor que los registrados en los
aislamientos de Metarhizium después de una hora de exposición.
Las diferencias encontradas entre la tolerancia de los aislamientos evaluados en este trabajo y
en los realizados por diferentes investigadores alrededor del mundo sugieren que además de
diferencias genéticas que confieren tolerancia a la radiación, las diferencias en la metodología
de realización de los trabajos influye directamente sobre los resultados obtenidos y hace que
las comparaciones sean siempre relativas, sin embargo, si se observa un efecto negativo de
la exposición a la UV-B, lo que coincide con los resultados obtenidos en los trabajos
presentados por diferentes autores.
El conocimiento de la tolerancia de los aislamientos a este y otros factores ambientales
permitirá orientar las investigaciones para el desarrollo de estrategias que minimizen el
impacto del ambiente cuando los hongos son aplicados en el campo, esto mediante el
desarrollo de formulaciones con filtros solares o sustancias que actúen como protectores
solares, o también mediante el uso de medios de cultivo que induzcan tolerancia a la UV
(Mulero, 2004).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
71
2.6 Bibliografía
Aikens, J., y Dix, T. A. 1991. Perhydroxyl radical (HOO.) initiated lipid peroxidation. The role of
fatty acid hydroperoxides. Journal of Biological Chemistry, 266(23): 15091-15098.
Alves, R.T., R.P. Bateman, C. Prior and S.R. Leather. 1998. Effects of simulated solar radiation
on conidial germination of Metarhizium anisopliae in different formulations. Crop Prot., 17: 675-
679.
Bidochka, M. J., Kamp, A. M., Lavender, T. M., Dekoning, J., y De Croos, J. A. 2001. Habitat
association in two genetic groups of the insect-pathogenic fungus Metarhizium anisopliae:
uncovering cryptic species?. Applied and Environmental Microbiology, 67(3):1335-1342.
Braga, G. U., Rangel, D. E., Flint, S. D., Anderson, A. J., y Roberts, D. W. 2006. Conidial
pigmentation is important to tolerance against solar simulated radiation in the Entomopathogenic
Fungus Metarhizium anisopliae.Photochemistry and photobiology, 82(2): 418-422.
Braga, G.U.L., D.E.N. Rangel, S.D. Flint, C.D. Miller, A.J., Anderson, y D.W. Roberts. 2002.
Damage and recovery from UV-B exposure in conidia of the entomopathogens Verticillium
lecanii and Aphanocladium album. Mycologia 94:912-920.
Braga, G.U.L., S.D. Flint, C.D. Miller, A.J. Anderson y D.W. Roberts. 2001. Variability in
response to UV-B among species and strains of Metarhizium isolated from sites at latitudes
from 61oN to 54oS. J. Invertebrate Pathol., 78: 98-108.
Braga, G.U.L., Flint, S.D., Messias, C.L., Anderson, A.J., Roberts, D.W. 2001a. Effect of UV-B
on conidia and germlings of the entomopathogenic hyphomycete Metarhizium anisopliae. Mycol.
Res. 105: 874–882.
Brooks, A. J., De Muro, M. A., Burree, E., Moore, D., Taylor, M. A., y Wall, R. 2004. Growth and
pathogenicity of isolates of the fungus Metarhizium anisopliae against the parasitic mite,
Psoroptes ovis: effects of temperature and formulation. Pest management science, 60(10):1043-
1049.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
72
Devotto, L.; Gerding, M. 2003. Respuesta de dos aislamientos chilenos de Metarhizium
anisopliae (Metschnikoff) sorokin a la adición de un protector solar. Agricultura Técnica 63: 339-
346.
Diffey, B. L. 1991. Solar ultraviolet radiation effects on biological systems.Physics in medicine
and biology, 36(3):299.
Fargues, J., Goettel, M.S., Smits, N., Ouedraogo, A., Vidal, C., Lacey, L.A., Lomer, C.J.,
Rougier, M., 1996. Variability in susceptibility to simulated sunlight of conidia among isolates of
entomopathogenic Hyphomycetes. Mycopathologia 135: 171–181.
Fargues, J., Rougier, M., Goujet, R., Smits, N., Coustere, C., Itier, B., 1997. Inactivation of
conidia of Paecilomyces fumosoroseus by near-ultraviolet (UVB and UVA) and visible radiation.
J. Invertebr. Pathol. 69: 70–78.
Fernandes, E.K., Rangel, D.E., Moraes, A.M., Bittencourt, V.R., Roberts, D.W. 2007. Variability
in tolerance to UV-B radiation among Beauveria spp. isolates. J. Invertebr. Pathol. 96: 237–243.
Fernandes, E.K.K., Rangel, D.E.N., Moraes, A.M.L., Bittencourt, V.R.E.P., Roberts, D.W., 2008.
Cold activity of Beauveria and Metarhizium , and thermotolerance of Beauveria. J. Invertebr.
Pathol. 98: 69–78.
Ghajar, F., Holford, P., Cother, E., y Beattie, A. 2006. Effects of ultraviolet radiation, simulated or
as natural sunlight, on conidium germination and appressorium formation by fungi with potential
as mycoherbistats. Biocontrol Science and Technology, 16(5):451-469.
Griffiths, H.R., Mistry, P., Herbert, K.E., Lunec, J. 1998. Molecular and cellular effects of
ultraviolet light-induced genotoxicity. Crit. Rev. Clin. Lab. Sci. 35: 189–237.
Gutteridge, J., y Halliwell, B. 1990. The measurement and mechanism of lipid peroxidation in
biological systems. Trends in biochemical sciences, 15(4): 129-135.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
73
Huang, BF, y feng, MG .2009. Tolerancias comparativos de diversos aislados de Beauveria
bassiana a los rayos UV-B de la irradiación con una descripción de un método de modelado
para evaluar dosis letal. Mycopathologia, 168 (3): 145-152.
Hullo, M., Moszer, I.; Danchin, A.; Martin, I. 2001. CotA of Bacillus subtilis is a copper dependent
laccase. Journal of Bacteriology 183: 5426-5430.
Ignoffo, C. M., Hostetter, D. L., Sikorowski, P. P., Sutter, G., y Brooks, W. M. 1977. Inactivation
of representative species of entomopathogenic viruses, a bacterium, fungus, and protozoan by
an ultraviolet light source. Environmental Entomology, 6(3):411-415.
Jacobs, J. L., y Sundin, G. W. 2001. Effect of solar UV-B radiation on a phyllosphere bacterial
community. Applied and environmental microbiology,67 (12): 5488-5496.
Johnson, D., 2003. Response of terrestrial microorganisms to ultraviolet-B radiation in
ecosystems. Res. Microbiol. 154: 315–320.
Krutmann, J. 2006. The interaction of UVA and UVB wavebands with particular emphasis on
signalling. Progress in biophysics and molecular biology, 92 (1):105-107.
Loong, C. Y., Sajap, A. S., Noor, H. M. Omar, D. Abood, F. 2013. Effects of UV-B and solar
Radiation on the efficacy of Isaria fumosorosea and Metharhizium anisopliae (Deuteromycetes:
Hyphomycetes) for controlling Bagworm, Pteroma pendula (Lepidoptera: Psychidae). Journal of
Entomology 10(2): 53-65.
Mulero, M. 2004. Efecto de la radiaci6n ultravioleta (RUV) sobre los procesos de estres
oxidativo e inmunosupresión cutánea. Efecto protector de los filtros solares. Tesis doctoral.
Facultad de Medicina. Universidad Rovira Virgili. 174 p.
Mustafa, U., y Kaur, G. 2009. UV-B radiation and temperature stress causes variable growth
response in Metarhizium anisopliae and Beauveria bassiana isolates. Internet Journal of
Microbiology, 7(1).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
74
Nicholson, W.; Schuerger A.; Setlow P. 2005. The solar UV environment and bacterial spore UV
resistance: considerations for earth - to Mars transport by natural processes and human
spaceflight. Mutation Research 571: 249-264.
Paul ND, Gwynn-Jones D. 2003. Ecological roles of solar UV radiation: towards an integrated
approach. Trends in Ecology and Evolution 18: 48–55.
Pavone D. 2003. Formulación de un Bioinsecticida con Base en el Hongo Nomuraea
rileyi (Farlow) Samson para el Control de Spodoptera frugiperda Smith (Lepidoptera:
Noctuidae). Tesis. Universidad Central de Venezuela. Caracas, Venezuela. 147 pp.
Polar, P., Kairo, M. T., Peterkin, D., Moore, D., Pegram, R., y John, S. A. 2005. Assessment of
fungal isolates for development of a mycoacaricide for cattle tick control. Vector-Borne &
Zoonotic Diseases, 5(3):276-284.
Posadas Julieta, B., Maricel, A. L., Mini Jorge, I., y Lecuona Roberto, E. 2012. Natural Tolerance
to UV-B and Assessment of Photoprotectants in Conidia of Six Native Isolates of Beauveria
bassiana (Bals-Criv) Vuillemin. World Applied Sciences Journal, 20(7):1024-1030.
Rangel, D.E.N., Braga, G.U.L., Anderson, A.J., Roberts, D.W., 2005. Influence of growth
environment on tolerance to UV-B radiation, germination speed, and morphology of Metarhizium
anisopliae var. acridum conidia. J. Invertebr. Pathol. 90: 55–58.
Rangel, D.E.N., Butler, M.J., Torabinejad, J., Anderson, A.J., Braga, G.U.L., Day, A.W., Roberts,
D.W., 2006. Mutants and isolates of Metarhizium anisopliae are diverse in their relationships
between conidial pigmentation and stress tolerance. J. Invertebr. Pathol. 93: 170–182.
Saxene, D., Bendov, E., Mansherob, R., Barak. Z., Boussiba, S., Zaritsky, A. 2002. A UV
tolerant mutant of Bacillus thuringiensis spp. Kurstaki producing melanin. Microbiology 44: 25-
32.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
75
Vélez, P. y Montoya, E. 1995. Efecto de la radiación solar en la supervivencia del hongo
Beauveria bassiana (Bals.) Vuill. en laboratorio y campo. Revista Colombiana de Entomología.
Vol. 21(2):91-98.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
76
CAPITULO 3: DETERMINACIÓN DE LA HIDROFOBICIDAD DE
CONIDIAS DE HONGOS ENTOMOPATÓGENOS.
3.1 Introducción
La hidrofobicidad es una característica de repelencia de una sustancia al agua. En los hongos
miceliales está determinada por la proteína hidrofobina secretada únicamente por ellos
(Wessels, 2000).
Las hidrofobinas son proteínas anfipáticas compuestas por 100 a 150 aminoácidos, con una
característica típica, la presencia de ocho residuos de cisteína en lugares conservados a lo
largo de la cadena de aminoácidos (Wessels, 2000). Las hidrofobinas han evolucionado para
intervenir en diferentes procesos celulares como la adhesión de los conidias a la cutícula de los
insectos (Li Jun et al. 2010), que es el primer paso en la patogénesis en hongos como
B. bassiana y Metarhizium ( Boucias y Pendland, 1991) también permiten un primer intercambio
de señales para que el hongo desarrolle hifas aéreas implicadas en la adherencia a las cutícula
de los insectos (Boucias et al. 1988), la formación de cuerpos fructíferos (Wessels, 1997), la
difusión de esporas por las corrientes de aire (Wessels, 1997; Linder et al., 2005) en muchos
hongos el papel exacto de la hidrofobinas particularmente en términos de virulencia y
propiedades de la capa de esporas aun no es claro (Zang et al. 2011) (Linder et al. 2005).
Las hidrofobinas son capaces de reconvertir la carga de una superficie, de hidrofílica a
hidrofóbica y viceversa (Boucias et al. 1988, Acevedo 2003). Se ha encontrado que ejercen una
acción de protección de las conidias contra el calor, la desecación y condiciones adversas del
ambiente y son un mecanismo de defensa de los hongos contra la respuesta inmune de los
insectos enmascarando componentes de la pared celular de hongos altamente inmunogénicos
(Heddergott et al. 2012. Paris et al, 2003). La hidrofobicidad de las estructuras de los hongos
conidias y micelio es determinada por las hidrofobinas, el recubrimiento que estas proteínas
producen es llamado capa rodlet (Bayry et al. 2012). La formación de estos recubrimientos
hidrófobos se asocia con la maduración de las esporas (Kim et al., 2010).

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
77
Wessels (1994) describe dos clases de hidrofobinas y las denomina Clase I y Clase II. Los
hongos pueden producir conidias hidrofóbicas (baja afinididad con el agua) o hidrofílicas (alta
afinidad con el agua). La cubierta externa de los conidias hidrofóbicas como las de Beauveria
bassiana o Metarhizium anisopliae es la estructura inicial donde se lleva a cabo la interfase de
unión con la cutícula del insecto hospedero (Boucias et al. 1988). Algunos hongos
entomopatógenos como Hirsutella thompsonii, producen conidias hidrofílicas que se
caracterizan por que tienen una capa externa mucilaginosa producida durante su maduración
que sirve como antisecante y adhesivo que protege a las conidias de polifenoles tóxicos
presentes en la cutícula del hospedero (Boucias et al. 1988; Acevedo, 2003). Las especies de
hongos con conidias hidrofóbicas son generalmente más tolerantes a las altas temperaturas
que las especies con conidias hidrofílicas (Souza et al. 2014; Bayry et al. 2012).
Las propiedades de fijación de los tipos de células B. bassiana reveló que conidias aéreas se
adhieren mal a superficies débilmente polares, y rápidamente a ambas superficies hidrófobas e
hidrófilas. (Boucias et al. 1988; Holder y Keyhani, 2005). En blastosporas in vitro, sin embargo,
se observa que estas tienen una débil unión a superficies hidrófobas, superficies
moderadamente a débilmente polares, y rápidamente a las superficies hidrófilas. Conidias
sumergidas muestran un espectro más amplio de unión, y pueden adherirse a superficies
hidrófobas, débilmente polares e hidrófilas.
3.2 Hipótesis y Objetivos Específicos
Objetivo específico 3: Establecer la capacidad de adhesión de conidias
de Beauveria y Metarhizium a través de su hidrofobicidad.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
78
3.3 Metodología.
3.3.1 Aislamientos utilizados en el estudio
Se evaluaron 31 aislamientos de hongos entomopatógenos, 29 de Beauveria sp. y siete de
Metarhizium (Anexo 1), que en la actualidad pertenecen a la colección de hongos
entomopatógenos de la Corporación Centro de Investigación en Palma de Aceite (Cenipalma).
Los aislamientos fueron mantenidos en SDA Agar Sabouraud Dextrosa (SDA) Oxoid®
(composición g.L-1: peptona micológica 10, glucosa 40, agar 15).
3.3.2 Pruebas de hidrofobicidad
Se preparó una suspensión de conidias en 10 ml de agua destilada estéril + Tween 80 al 1%.
Se homogenizó la suspensión utilizando un agitador magnético. Mediante diluciones en serie,
se ajustó y determinó la concentración de conidias de la suspensión preparada a 1 x 10
6conidias.ml-1. Luego se tomó un ml de la suspensión de conidias y se depositó en un tubo de
ensayo, al que se le agregó un ml de tolueno. El tubo de ensayo fue centrifugado a 500 rpm
durante 40 minutos. Después de centrifugados los tubos de ensayo se ubicaron verticalmente y
de la fase acuosa se extrajeron diez µl que se depositaron en la cámara de Neubauer. Se
realizó el conteo de las conidias en la fase acuosa. El cálculo del porcentaje de conidias
presentes en la fase acuosa se realizó teniendo en cuenta el conteo inicial de conidias en la
suspensión y el conteo realizado en la muestra de la fase acuosa después de la centrifugación.
Para cada uno de los aislamientos se realizaron cinco repeticiones.
3.3.4 Análisis estadístico
Se realizó un análisis de varianza con un nivel de significancia del 95%. Para cada uno de los
generos Beauveria, y Metarhizium. En ambos casos, se verificó que se cumplieran los
supuestos de normalidad y homocedasticidad con un nivel de significancia del 99%.
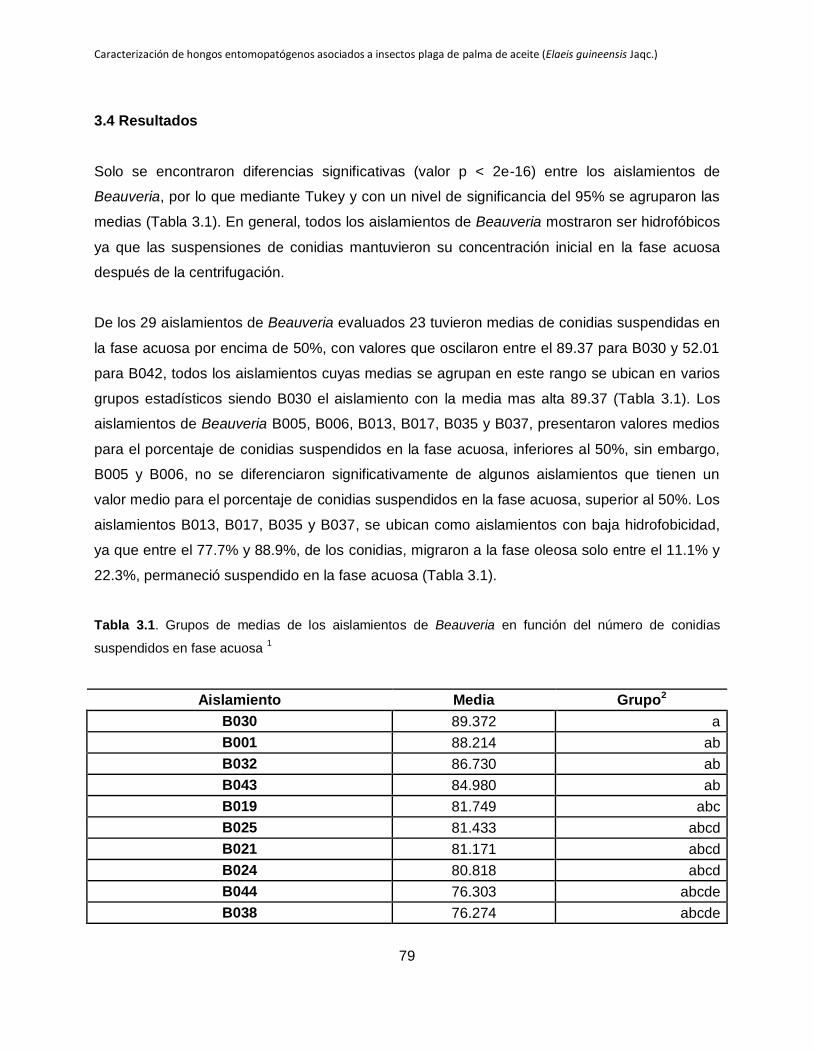
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
79
3.4 Resultados
Solo se encontraron diferencias significativas (valor p < 2e-16) entre los aislamientos de
Beauveria, por lo que mediante Tukey y con un nivel de significancia del 95% se agruparon las
medias (Tabla 3.1). En general, todos los aislamientos de Beauveria mostraron ser hidrofóbicos
ya que las suspensiones de conidias mantuvieron su concentración inicial en la fase acuosa
después de la centrifugación.
De los 29 aislamientos de Beauveria evaluados 23 tuvieron medias de conidias suspendidas en
la fase acuosa por encima de 50%, con valores que oscilaron entre el 89.37 para B030 y 52.01
para B042, todos los aislamientos cuyas medias se agrupan en este rango se ubican en varios
grupos estadísticos siendo B030 el aislamiento con la media mas alta 89.37 (Tabla 3.1). Los
aislamientos de Beauveria B005, B006, B013, B017, B035 y B037, presentaron valores medios
para el porcentaje de conidias suspendidos en la fase acuosa, inferiores al 50%, sin embargo,
B005 y B006, no se diferenciaron significativamente de algunos aislamientos que tienen un
valor medio para el porcentaje de conidias suspendidos en la fase acuosa, superior al 50%. Los
aislamientos B013, B017, B035 y B037, se ubican como aislamientos con baja hidrofobicidad,
ya que entre el 77.7% y 88.9%, de los conidias, migraron a la fase oleosa solo entre el 11.1% y
22.3%, permaneció suspendido en la fase acuosa (Tabla 3.1).
Tabla 3.1. Grupos de medias de los aislamientos de Beauveria en función del número de conidias
suspendidos en fase acuosa 1
Aislamiento Media Grupo2
B030 89.372 a
B001 88.214 ab
B032 86.730 ab
B043 84.980 ab
B019 81.749 abc
B025 81.433 abcd
B021 81.171 abcd
B024 80.818 abcd
B044 76.303 abcde
B038 76.274 abcde
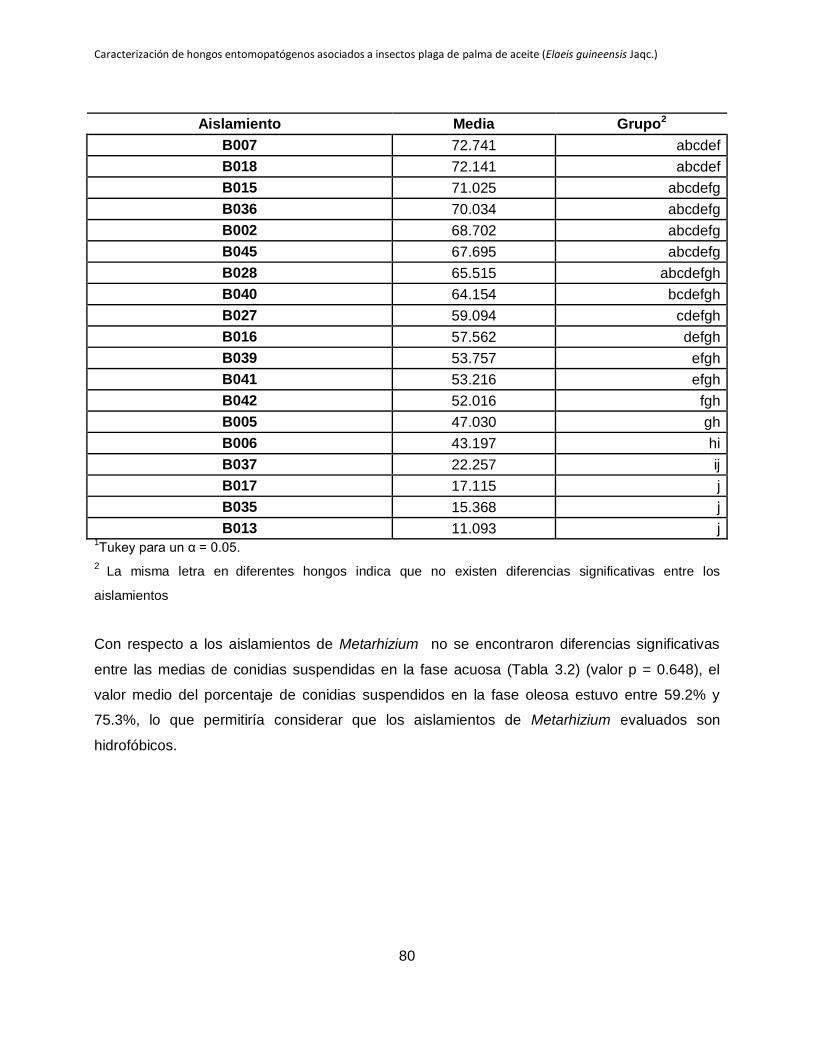
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
80
Aislamiento Media Grupo2
B007 72.741 abcdef
B018 72.141 abcdef
B015 71.025 abcdefg
B036 70.034 abcdefg
B002 68.702 abcdefg
B045 67.695 abcdefg
B028 65.515 abcdefgh
B040 64.154 bcdefgh
B027 59.094 cdefgh
B016 57.562 defgh
B039 53.757 efgh
B041 53.216 efgh
B042 52.016 fgh
B005 47.030 gh
B006 43.197 hi
B037 22.257 ij
B017 17.115 j
B035 15.368 j
B013 11.093 j 1Tukey para un α = 0.05.
2 La misma letra en
diferentes hongos indica que no existen diferencias significativas entre los
aislamientos
Con respecto a los aislamientos de Metarhizium no se encontraron diferencias significativas
entre las medias de conidias suspendidas en la fase acuosa (Tabla 3.2) (valor p = 0.648), el
valor medio del porcentaje de conidias suspendidos en la fase oleosa estuvo entre 59.2% y
75.3%, lo que permitiría considerar que los aislamientos de Metarhizium evaluados son
hidrofóbicos.
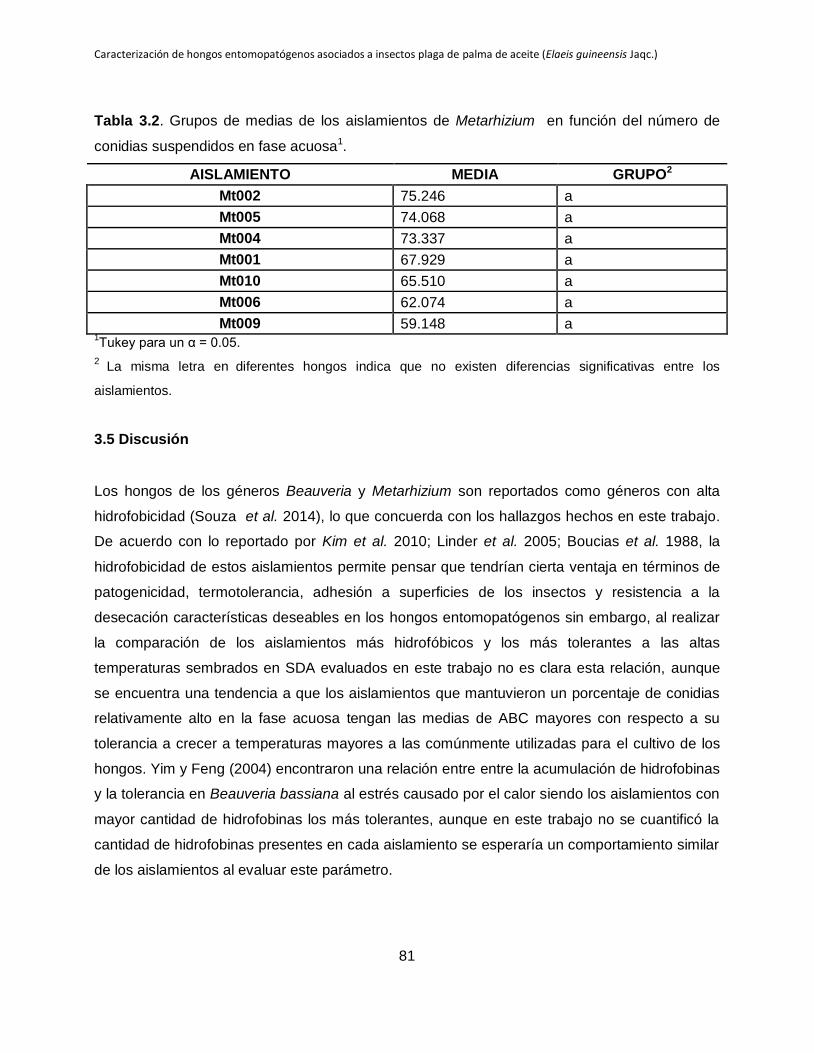
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
81
Tabla 3.2. Grupos de medias de los aislamientos de Metarhizium en función del número de
conidias suspendidos en fase acuosa1.
AISLAMIENTO MEDIA GRUPO2
Mt002 75.246 a
Mt005 74.068 a
Mt004 73.337 a
Mt001 67.929 a
Mt010 65.510 a
Mt006 62.074 a
Mt009 59.148 a 1Tukey para un α = 0.05.
2 La misma letra en
diferentes hongos indica que no existen diferencias significativas entre los
aislamientos.
3.5 Discusión
Los hongos de los géneros Beauveria y Metarhizium son reportados como géneros con alta
hidrofobicidad (Souza et al. 2014), lo que concuerda con los hallazgos hechos en este trabajo.
De acuerdo con lo reportado por Kim et al. 2010; Linder et al. 2005; Boucias et al. 1988, la
hidrofobicidad de estos aislamientos permite pensar que tendrían cierta ventaja en términos de
patogenicidad, termotolerancia, adhesión a superficies de los insectos y resistencia a la
desecación características deseables en los hongos entomopatógenos sin embargo, al realizar
la comparación de los aislamientos más hidrofóbicos y los más tolerantes a las altas
temperaturas sembrados en SDA evaluados en este trabajo no es clara esta relación, aunque
se encuentra una tendencia a que los aislamientos que mantuvieron un porcentaje de conidias
relativamente alto en la fase acuosa tengan las medias de ABC mayores con respecto a su
tolerancia a crecer a temperaturas mayores a las comúnmente utilizadas para el cultivo de los
hongos. Yim y Feng (2004) encontraron una relación entre entre la acumulación de hidrofobinas
y la tolerancia en Beauveria bassiana al estrés causado por el calor siendo los aislamientos con
mayor cantidad de hidrofobinas los más tolerantes, aunque en este trabajo no se cuantificó la
cantidad de hidrofobinas presentes en cada aislamiento se esperaría un comportamiento similar
de los aislamientos al evaluar este parámetro.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
82
Rangel et al. 2006; Kim et al. 2011 reportan que la producción de hidrofobinas y la inducción de
una mayor hidrofobicidad, está relacionada con el medio en el que se desarrollan los hongos
conidias de B. bassiana y M. anisopliae producidas en agar de millo son más hidrofobicas y
tienen una mayor tolerancia a altas temperaturas que las producidas en SDA o Agar extracto de
levadura los medios con lactosa también inducen una alta hidrofobicidad en las conidias,
aunque en un alto porcentaje los aislamientos evaluados en este trabajo mantuvieron el número
de conidias por encima del 70% en la fase acuosa este valor podría aumentar si se utiliza un
medio de cultivo diferente a SDA que induzca la producción de hidrofobinas.
Holder y Keyhani, 2005 detectaron que las conidias aéreas generan una capa de bastoncillos
que no permiten que los conidios se suspendan fácilmente en el agua, este comportamiento
causado por una capa hidrofobinas no se evidencia en blastosporas o conidias que crecen
sumergidas, las conidias utilizadas en este ensayo fueron conidias aéreas que no se suspenden
fácilmente en el agua lo que corrobora esta observación.
Kim et al. 2010 encontraron que la edad de las conidias y la termotolerancia de los aislamientos
esta relacionada con su capacidad hidrofóbica, las conidas más viejas son mas termotolerantes
que las más jóvenes por esta razon en futuros trabajos la combinación entre edad de las
conidias y temperaturas de incubación en relación con su capacidad hidrifóbica deberian ser
evaluados en condiciones de laboratorio antes de realizar pruebas en campo.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
83
3.6 Bibliografía
Acevedo, J. 2003. Actividad biológica de los exudados y filtrado crudo de Hirsutella thompsonii
FISHER (Cepa HtM120I) sobre Tetranychus urticae KOCH y otros artrópodos. Trabajo de grado
para optar el titulo de Doctor en ciencias, área biotecnología. Universidad de Colima. Tecoman
México. 193 p.
Bayry, J., Aimanianda, V., Guijarro, J. I., Sunde, M., y Latge, J. P. 2012. Hydrophobins – Unique
fungal proteins. Plos Pathogens, 8.
Boucias, D. G., Pendland, J. C., y Latge, J. P. 1988. Nonspecific factors involved in attachment
of entomopathogenic deuteromycetes to host insect cuticle. Applied and environmental
microbiology, 54(7), 1795-1805.
Boucias, D., y J. Pendland. 1991. Attachment of mycopathogens to cuticle, p. 101–127. In G. T.
Cole and H. C. Hoch (ed.), The fungal spore and disease initiation in plants and animals.
Plenum Press, New York, N.Y.
Heddergott, C., Bruns, S., Nietzsche, S., Leonhardt, I., Kurzai, O., Kniemeyer, O., y Brakhage,
A. A. 2012. The Arthroderma benhamiae hydrophobin HypA mediates hydrophobicity and
influences recognition by human immune effector cells. Eukaryotic Cell, 11, 673–682.
Holder, D. J., y Keyhani, N. O. 2005. Adhesion of the entomopathogenic fungus Beauveria
(Cordyceps) bassiana to substrata. Applied and environmental microbiology, 71(9), 5260-5266.
Kim, J. S., Kassa, A., Skinner, M., Hata, T., y Parker, B. L. 2011. Production of thermotolerant
entomopathogenic fungal conidia on millet grain. Journal of Industrial Microbiology &
Biotechnology, 38, 697–704.
Kim, J. S., Skinner, M., Hata, T., y Parker, B. L. 2010. Effects of culture media on hydrophobicity
and thermotolerance of Bb and Ma conidia, with description of a novel surfactant based
hydrophobicity assay. Journal of Invertebrate Pathology, 105, 322–328.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
84
Li, J., Ying, S. H., Shan, L. T., y Feng, M. G. 2010. A new non-hydrophobic cell wall protein
(CWP10) of Metarhizium anisopliae enhances conidial hydrophobicity when expressed in
Beauveria bassiana. Applied microbiology and biotechnology, 85(4), 975-984.
Linder, M. B., Szilvay, G. R., Nakari‐Setälä, T., y Penttilä, M. E. 2005. Hydrophobins: the protein‐
amphiphiles of filamentous fungi. FEMS microbiology reviews, 29(5), 877-896.
Paris, S., Debeaupuis, J.P., Crameri, R., Carey, M., Charles, F., Prevost, M.C., Schmitt, C.,
Philippe, B., y Latgé, J.P. 2003. Conidial hydrophobins of Aspergillus fumigatus. Appl Environ
Microbiol 69: 1581– 1588.
Rangel, D. E. N., Anderson, A. J., y Roberts, D. W. 2006. Growth of Metarhizium anisopliae on
non-preferred carbon sources yields conidia with increased UV-B tolerance. Journal of
Invertebrate Pathology. 93, 127–134.
Souza, R. K., Azevedo, R. F., Lobo, A. O., y Rangel, D. E. 2014. Conidial water affinity is an
important characteristic for thermotolerance in entomopathogenic fungi. Biocontrol Science and
Technology, 24(4).
Wessels, J. G. H. 2000. Hydrophobins, unique fungal proteins. Mycologist,14 (4), 153-159.
Wessels, J. G. H. 1997. Hydrophobins: proteins that change the nature of the fungal surface.
Advances in Microbial Physiology 38: 1-45.
Wessels, J. G. H. 1994. Developmental regulation of fungal cell wall formation. Annual
Review of Phytopathology 32: 413-437.
Ying, S. H., y Feng, M. G. 2004. Relationship between thermotolerance and hydrophobin‐like
proteins in aerial conidia of Beauveria bassiana and Paecilomyces fumosoroseus as fungal
biocontrol agents. Journal of applied microbiology, 97(2), 323-331.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
85
Zhang, S., Xia, Y. X., Kim, B., y Keyhani, N. O. 2011. Two hydrophobins are involved in fungal
spore coat rodlet layer assembly and each play distinct roles in surface interactions,
development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Molecular
microbiology, 80(3), 811-826.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
86
CONCLUSIONES
Los resultados obtenidos son contrastantes incluso en aislamientos colectados en la
misma zona geográfica, sugiriendo que la tasa de crecimiento en diferentes medios y
temperaturas, la tolerancia a luz ultra violeta y la hidrofobicidad son caracteres
particulares de cada aislamiento.
Se ratifica la importancia de la temperatura de incubación y su efecto positivo o
negativo en el crecimiento de los hongos, encontrándose en algunos casos que leves
incrementos en la temperatura favorecen el crecimiento de los hongos, lo que podría
indicar tolerancia o adaptación a las temperaturas de las zonas en donde fueron
encontrados los aislamientos objeto de estudio. La interacción medio de cultivo-
temperatura es determinante en el desarrollo de los aislamientos
La exposición de los hongos evaluados a radiación UV-B, afecta de forma gradual la
germinación de las conidias. Se observó que independientemente del género al que
pertenezca el aislamiento la tolerancia a la radiación UV-B es específica.
La hidrofobicidad de los aislamientos de Beauveria en general se considera alta,
posiblemente por un alto contenido de hidrofobinas característica deseable para los
hongos entomopatógenos esto les conferiría cierta ventaja en términos de
patogenicidad, termotolerancia, adhesión a superficies de los insectos y resistencia a
la desecación.

Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
87
RECOMENDACIONES
Una vez se seleccione un aislamiento con alta capacidad de virulencia sobre un insecto
de interés económico se deben realizar pruebas que permitan conocer las
características de cada hongo y así determinar las condiciones de producción mas
adecuadas para mantener sus carateristicas de virulencia en campo.
Para futuras pruebas de tolerancia a UV-B se debe evaluar la germinación después de
tiempos de incubación mayor, debido a que el efecto de la exposición a UV-B no
necesariamente es la muerte de las conidias, sino el aumento en los tiempos de
germinación de las mismas después de ser expuestas. Además se debe evaluar la
patogenicidad de las conidias después de ser expuestas a simuladores de radiación
solar.
La Hidrofobicidad hallada en los aislamientos evaluados sugiere que es necesario el uso
de surfactantes para garantizar la dispersión en una solución para ser aplicados en
campo, de otra forma el inóculo queda adherido a las superficies de los recipientes que
los contienen, reduciendo su potencial eficiencia en campo.
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
ANEXOS
Anexo 1. Aislamientos de Beauveria y Metarhizium colectados en diferentes zonas palmeras procedentes de insectos
plaga del cultivo de palma de aceite.
N.D.: No Disponible, a
Temperatura, b
Humedad Relativa,
c Tolerancia a Radiación UV-B,
d Crecimiento Radial,
e Hidrofobicidad.
Código Aislamiento Localidad Departamento Coordenada
s Hospedero
Estadío
Orden: familia Msnm
Precipitación promedio
(mm anuales)
Temperatura promedio
anual (°c)
Humedad relativa promedio anual
(%)
UV-B
C.R. HF.
B001 Beauveria spp. San Carlos de Guaroa
Meta 3°42'40''N 73°14'33''O
Episibine spp. Larva Lepidoptera:Limacodidae 250 2.700 25,9 80,5 X X X
B002 Beauveria San Carlos de Guaroa
Meta 3°42'40''N 73°14'33''O
Episibine spp. Larva Lepidoptera:Limacodidae 250 2.700 25,9 80,5 X X X
B005 Beauveria /Cenicafé/ N.D. N.D. Hypothenemus
hampei Larva Coleptera: Curculionidae N.D.* N.D. N.D. N.D.
X X X
B006 Beauveria San Andres de
Tumaco Nariño
1°48'24''N
78°45'53''O Stenoma cecropia Larva Lepidoptera: Elachistidae 40 3.000 26 88
X X X
B007 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Opsiphanes cassina Larva Lepidoptera: Nymphalidae 80 2.850 28 77 X X
B013 Beauveria Cumaral Meta 4°16'11''N 73°29'11'O
Loxotoma elegans Larva Lepidoptera: Elachistidae 260 2.800 26 80,5 X X X
B015 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Antaeotricha sp Larva Lepidoptera:Oecophoridae 80 2.850 28 77 X X X
B016 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X
B017 Beauveria Puerto Wilches Santander 7°20'54''N
73°53'54''O Dirphia gragatus Larva Lepidoptera:Saturniidae 80 2.850 28 77
X X X
B018 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X X
B019 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X X
B021 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X X
B024 Beauveria Puente Sogamoso Santander 7°20'54''N 73°53'54''O
Leptopharsa gibbicarina
Adulto Hemiptera : Tingidae 92 2.850 28 79 X X X
B025 Beauveria Puente Sogamoso Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Pupa Lepidoptera: Elachistidae 92 2.850 28 79 X X X
B027 Beauveria Puente Sogamoso Santander 7°20'54''N 73°53'54''O
Dirphia gragatus Larva Lepidoptera:Saturniidae 92 2.850 28 79 X X X
B028 Beauveria Puente Sogamoso Santander 7°20'54''N 73°53'54''O
Leptopharsa gibbicarina
Adulto Hemiptera : Tingidae 92 2.850 28 79 X X X
B030 Beauveria N.D. N.D. N.D. Producto comercial N.D. N.D. N.D. N.D. N.D. N.D. X X
B032 Beauveria Puente Sogamoso Santander 7°20'54''N 73°53'54''O
Demotispa neivai Adulto Coleptera: Chrysomelidae 92 2.850 28 79 X X X
B035 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X X
B036 Beauveria Tucurinca Magdalena 10°45'51''N 74°09'26''O
Leptopharsa gibbicarina
Adulto Hemiptera : Tingidae 20 1.130 27,6 80 X X
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
89
B037 Beauveria Tucurinca Magdalena 10°45'51''N 74°09'26''O
Brassolis sophorae Larva Lepidoptera: Nymphalidae 20 1.130 27,6 80 X X X
B038 Beauveria Tucurinca Magdalena 10°45'51''N 74°09'26''O
Brassolis sophorae Larva Lepidoptera: Nymphalidae 20 1.130 27,6 80 X X X
B039 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X
B040 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Stenoma cecropia Larva Lepidoptera: Elachistidae 80 2.850 28 77 X X X
B041 Beauveria Barrancabermeja Santander 7°04'03''N 73°50'50''O
Antaeotricha sp Larva Lepidoptera:Oecophoridae 80 2.538 27,1 83 X X X
B042 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Euprosterna elaeasa Larva Lepidoptera: Limacodidae 80 2.850 28 77 X X X
B043 Beauveria Puerto Wilches Santander 7°20'54''N 73°53'54''O
Elaidobius kamerunicus
Adulto Coleoptera:Curculionidae 80 2.850 28 77 X X X
B044 Beauveria San Carlos de Guaroa
Meta 3°42'40''N 73°14'33''O
N.D. Larva N.D. 250 2.700 25,9 80,5 X X X
B045 Beauveria San Carlos de Guaroa
Meta 3°42'40''N 73°14'33''O
Loxotoma elegans Larva Lepidoptera: Elachistidae 250 2.700 25,9 80,5 X X X
Mt001 Metarhizium Puerto Wilches Santander 7°20'54''N 73°53'54''O
N.D. N.D. N.D. 80 2.850 28 77 X X X
Mt002 Metarhizium Puerto Wilches Santander 7°20'54''N 73°53'54''O
Strategus aloeus Larva Coleoptera: Scarabaeidae 80 2.850 28 77 X X X
Mt004 Metarhizium San Andres de Tumaco
Nariño 1°48'24''N 78°45'53''O
Sagalassa valida Larva Lepidoptera: Glyphipterygidae
40 3.000 26 88 X X X
Mt005 Metarhizium Barrancabermeja Santander 7°04'03''N 73°50'50''O
Demotispa neivai Adulto Coleptera: Chrysomelidae 90 2.538 27,1 83 X X X
Mt006 Metarhizium Barrancabermeja Santander 7°04'03''N 73°50'50''O
Demotispa neivai Adulto Coleptera: Chrysomelidae 90 2.538 27,1 83 X X X
Mt009 Metarhizium Barrancabermeja Santander 7°04'03''N 73°50'50''O
Demotispa neivai Adulto Coleptera: Chrysomelidae 90 2.538 27,1 83 X X X
Mt010 Metarhizium Yarima Santander 6°52'55''N 73°24'43''O
Strategus aloeus Larva Coleoptera: Scarabaeidae 90 2.538 27,1 83 X X X
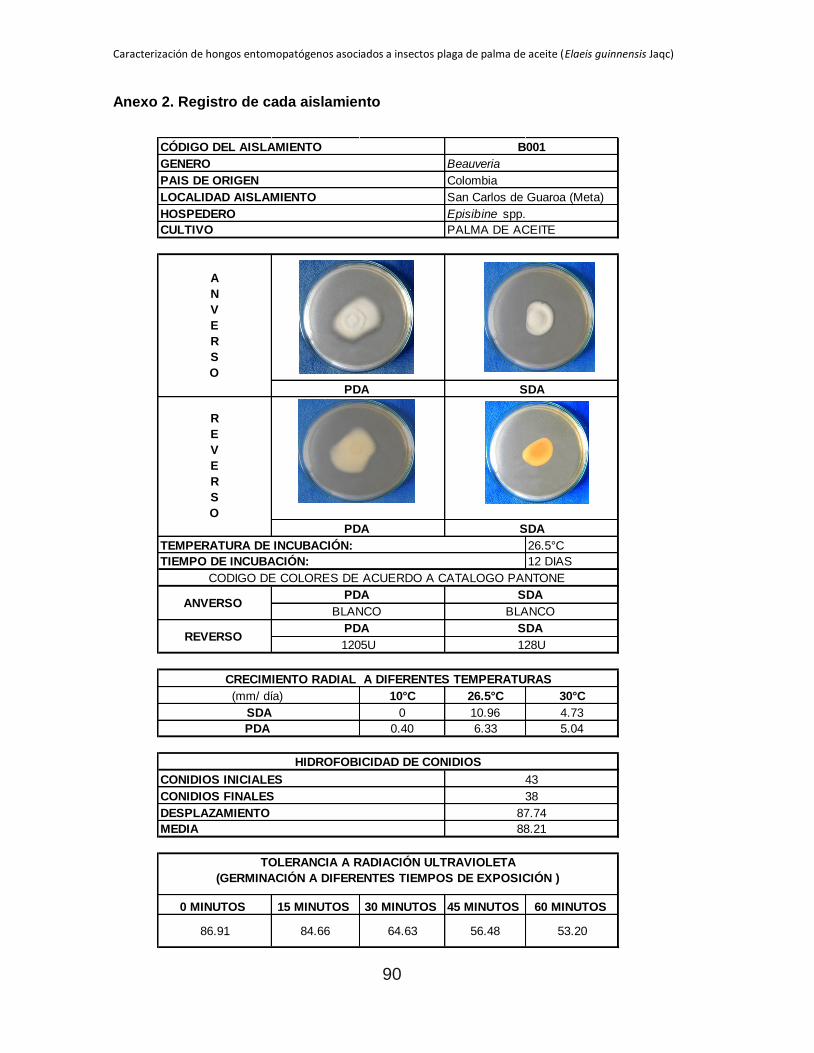
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guinnensis Jaqc)
90
Anexo 2. Registro de cada aislamiento
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 10.96 4.73
0.40 6.33 5.04
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
HIDROFOBICIDAD DE CONIDIOS
MEDIA 88.21
87.74DESPLAZAMIENTO
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN )
CONIDIOS INICIALES 43
CONIDIOS FINALES 38
86.91 84.66 53.2056.4864.63
TIEMPO DE INCUBACIÓN:
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
1205U 128U
PDA SDA
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA SDA
BLANCO BLANCO
TEMPERATURA DE INCUBACIÓN:
HOSPEDERO Episibine spp.
CULTIVO PALMA DE ACEITE
A
N
V
E
R
S
O
PDA SDA
R
E
V
E
R
S
O
PDA SDA
LOCALIDAD AISLAMIENTO San Carlos de Guaroa (Meta)
CÓDIGO DEL AISLAMIENTO B001
GENERO Beauveria
PAIS DE ORIGEN Colombia
PDA
SDA
(mm/ día)
ANVERSO
REVERSO
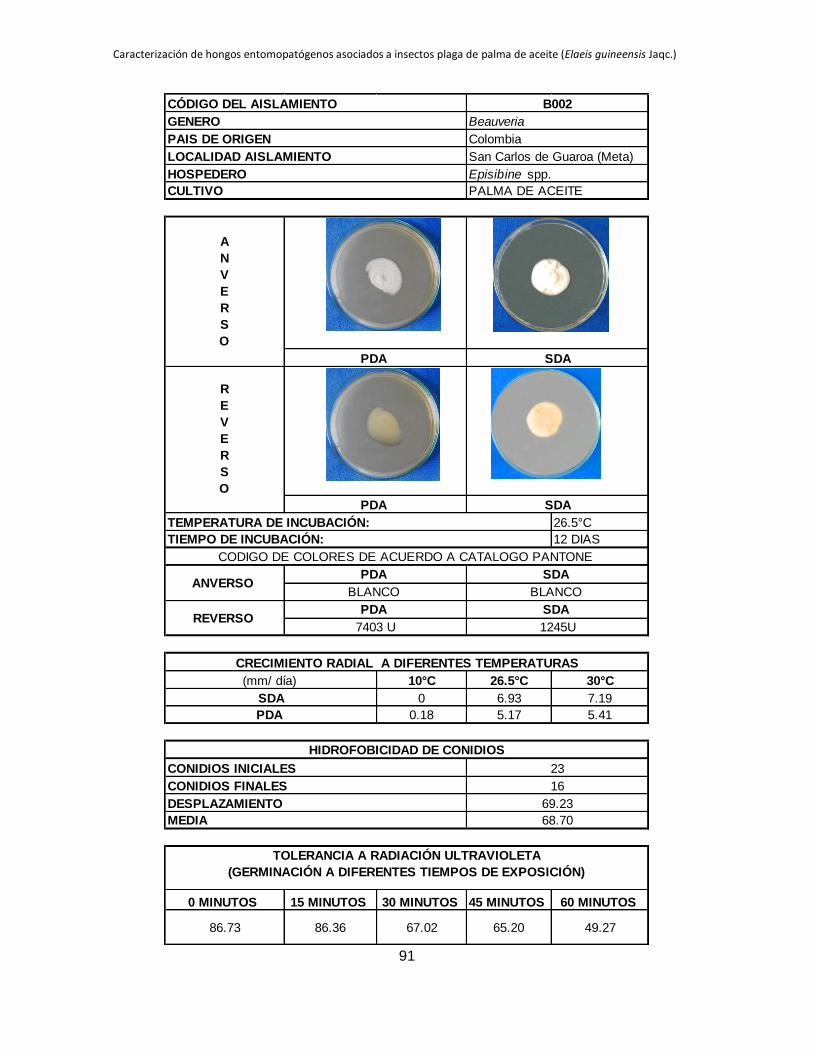
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
91
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 6.93 7.19
0.18 5.17 5.41
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
MEDIA 68.70
23
16
69.23
CONIDIOS INICIALES
CONIDIOS FINALES
DESPLAZAMIENTO
CÓDIGO DEL AISLAMIENTO B002
GENERO Beauveria
PAIS DE ORIGEN Colombia
LOCALIDAD AISLAMIENTO San Carlos de Guaroa (Meta)
HOSPEDERO Episibine spp.
CULTIVO PALMA DE ACEITE
PDA
HIDROFOBICIDAD DE CONIDIOS
BLANCO BLANCO
PDA
SDA
SDA
7403 U 1245U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
REVERSO
A
N
V
E
R
S
O
PDA SDA
R
E
V
E
R
S
O
PDA SDA
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
PDA SDA ANVERSO
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
86.73 86.36 67.02 65.20 49.27
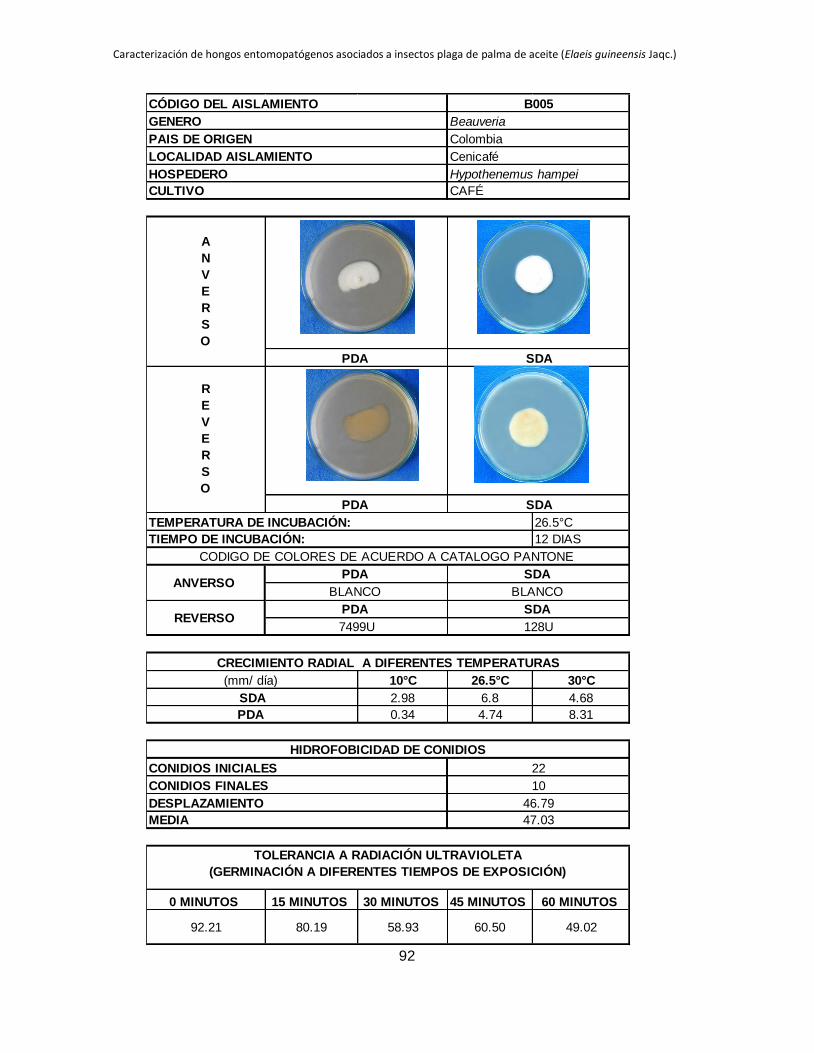
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
92
26.5°C
12 DIAS
10°C 26.5°C 30°C
2.98 6.8 4.68
0.34 4.74 8.31
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
92.21 80.19 49.0260.5058.93
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B005
Beauveria
Colombia
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
Cenicafé
Hypothenemus hampei
CAFÉ
PDA SDA
SDA
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
BLANCO BLANCO
CONIDIOS FINALES
ANVERSO
REVERSO
10
PDA SDA
7499U 128U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
46.79
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
DESPLAZAMIENTO
MEDIA 47.03
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 22
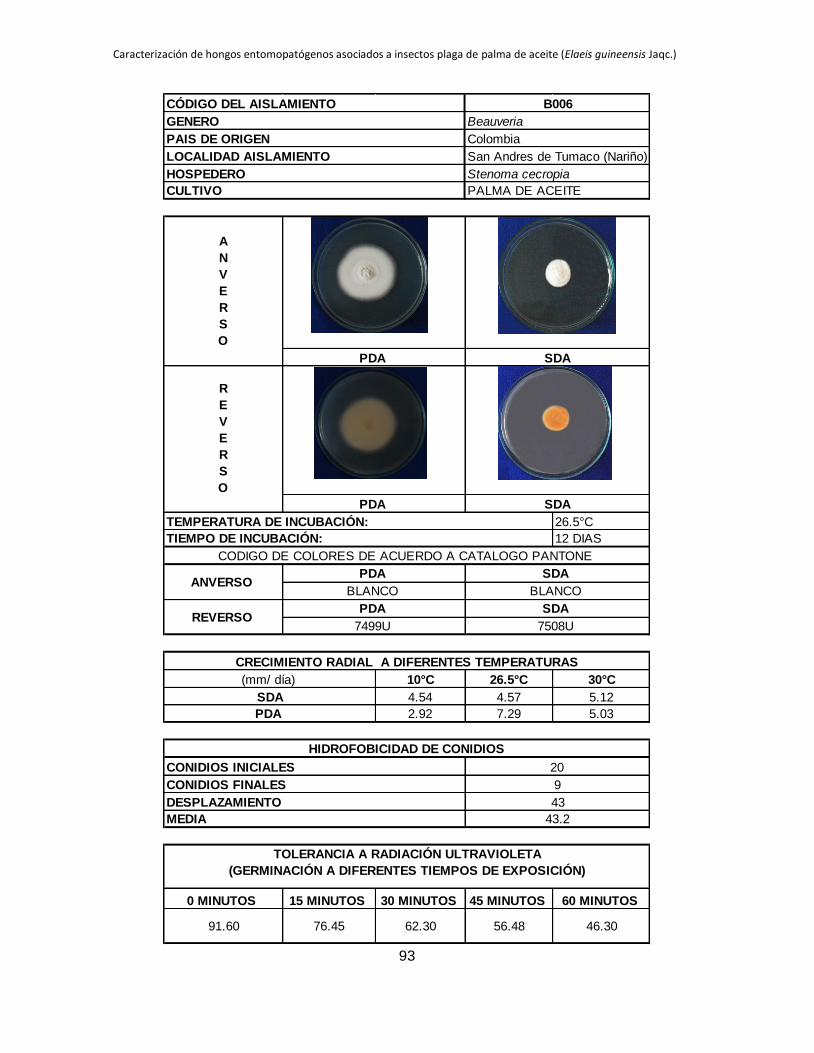
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
93
26.5°C
12 DIAS
10°C 26.5°C 30°C
4.54 4.57 5.12
2.92 7.29 5.03
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
91.60 76.45 62.30 56.48 46.30
CONIDIOS FINALES 9
DESPLAZAMIENTO 43
MEDIA 43.2
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 20
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
ANVERSO PDA SDA
BLANCO BLANCO
REVERSO PDA SDA
7499U 7508U
A
N
V
E
R
S
O
PDA SDA
R
E
V
E
R
S
O
PDA SDA
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
San Andres de Tumaco (Nariño)
Stenoma cecropia
PALMA DE ACEITE
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B006
Beauveria
Colombia
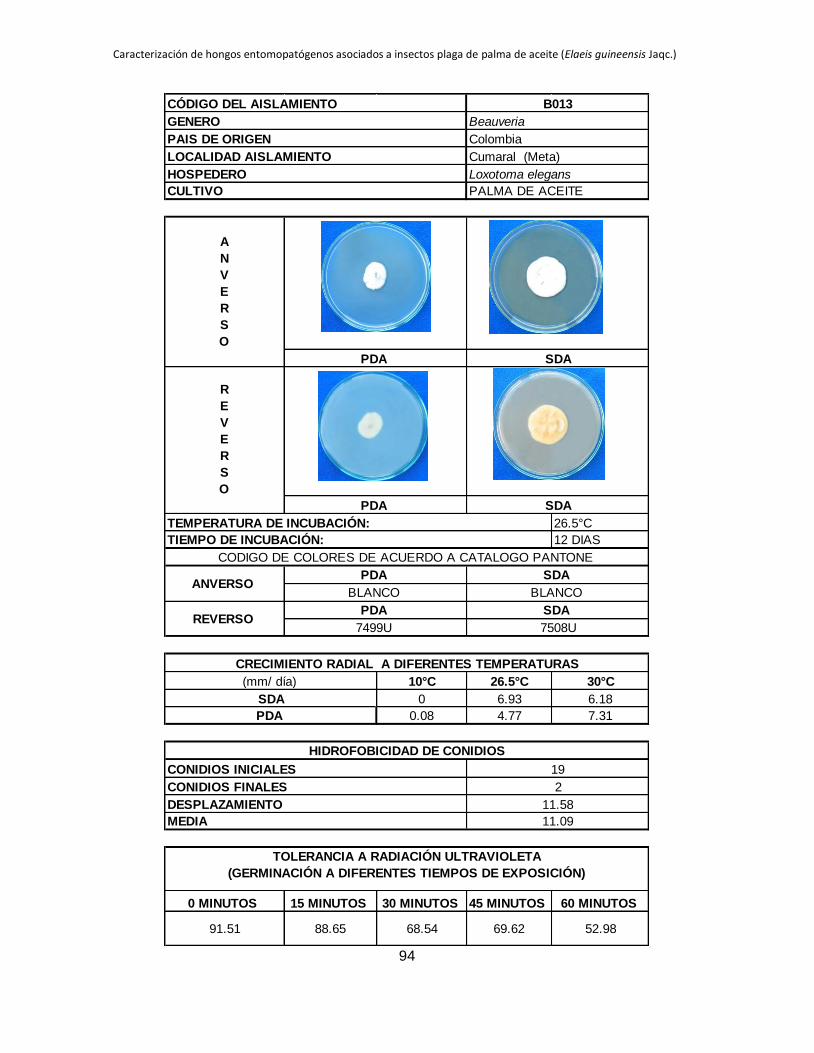
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
94
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 6.93 6.18
0.08 4.77 7.31
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
91.51 52.9869.6268.5488.65
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B013
Beauveria
Colombia
Cumaral (Meta)
Loxotoma elegans
PALMA DE ACEITE
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
CONIDIOS INICIALES 19
PDA SDA
7499U 7508U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
MEDIA 11.09
CONIDIOS FINALES 2
ANVERSO
REVERSO
HIDROFOBICIDAD DE CONIDIOS
BLANCO BLANCO
DESPLAZAMIENTO 11.58
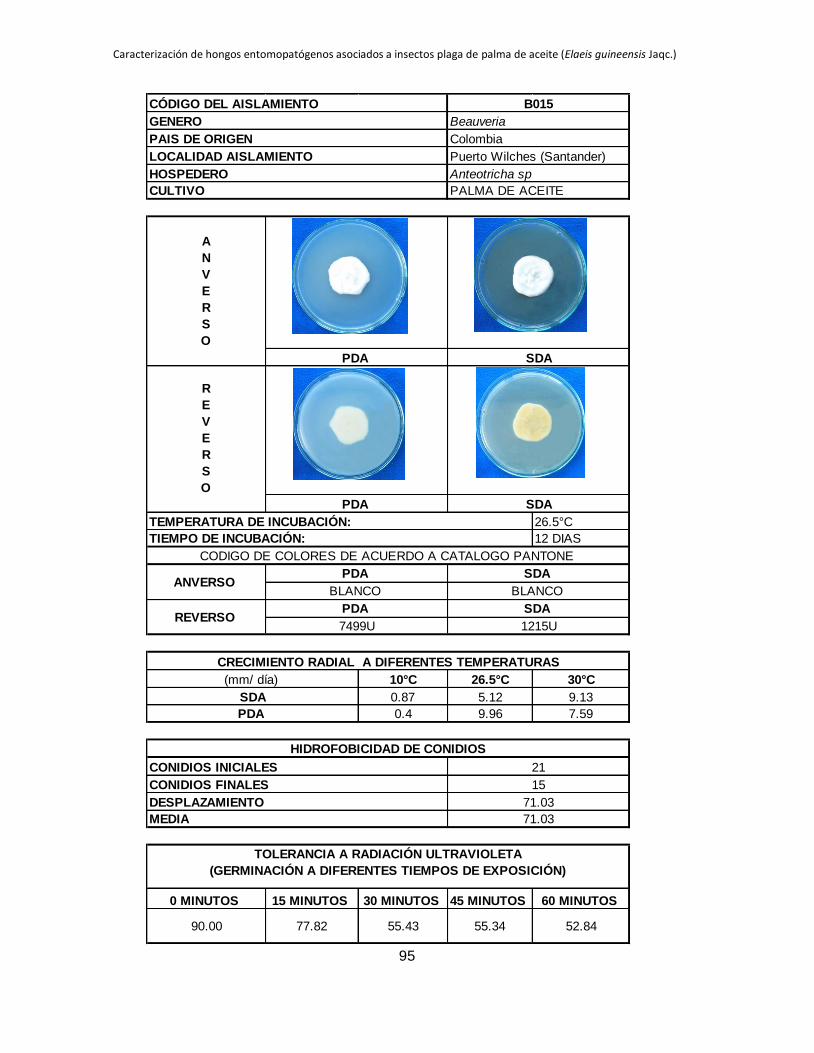
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
95
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.87 5.12 9.13
0.4 9.96 7.59
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
Puerto Wilches (Santander)
Anteotricha sp
PALMA DE ACEITE
90.00 77.82 55.43 52.8455.34
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B015
Beauveria
Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
CONIDIOS FINALES 15
PDA SDA
7499U 1215U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
MEDIA 71.03
CONIDIOS INICIALES 21
ANVERSO
REVERSO
HIDROFOBICIDAD DE CONIDIOS
BLANCO BLANCO
DESPLAZAMIENTO 71.03
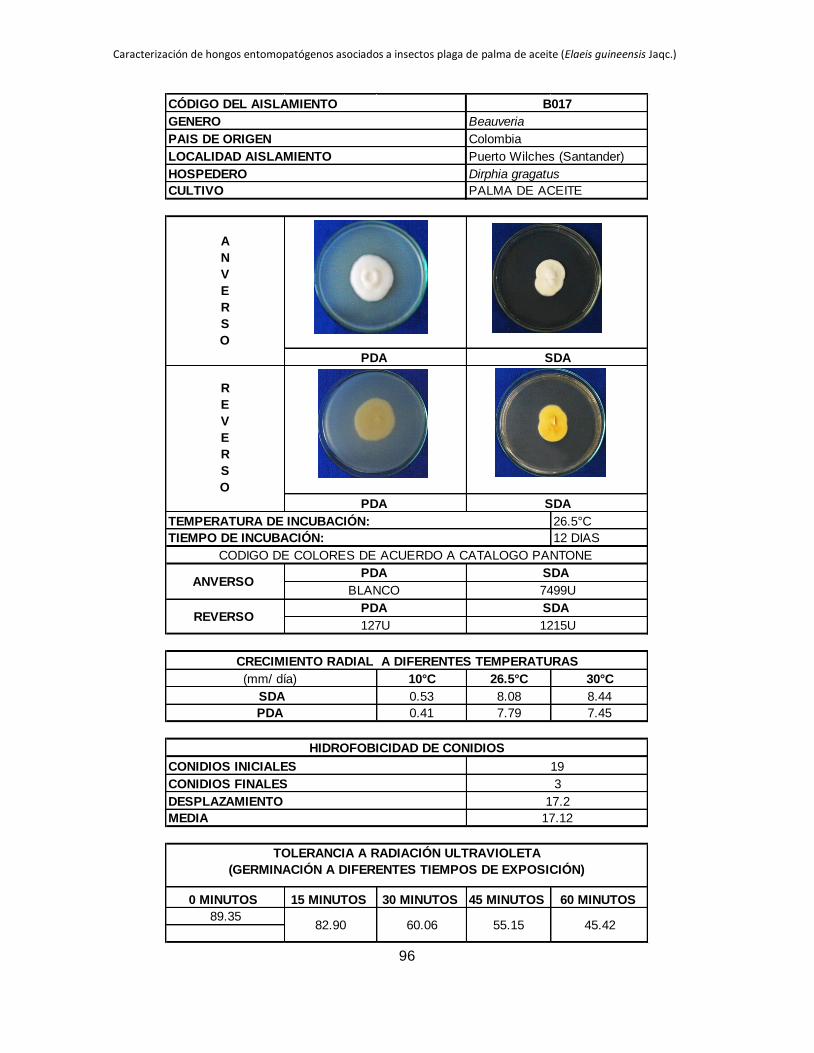
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
96
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.53 8.08 8.44
0.41 7.79 7.45
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
89.35
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
Puerto Wilches (Santander)
Dirphia gragatus
PALMA DE ACEITE
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B017
Beauveria
Colombia
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES
CONIDIOS FINALES 3
DESPLAZAMIENTO 17.2
55.15 45.4260.0682.90
19
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
MEDIA 17.12
A
N
V
E
R
S
O
PDA SDA
R
E
V
E
R
S
O
PDA SDA
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
SDA
PDA SDA
PDA
PDA
BLANCO 7499UANVERSO
REVERSO 127U 1215U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
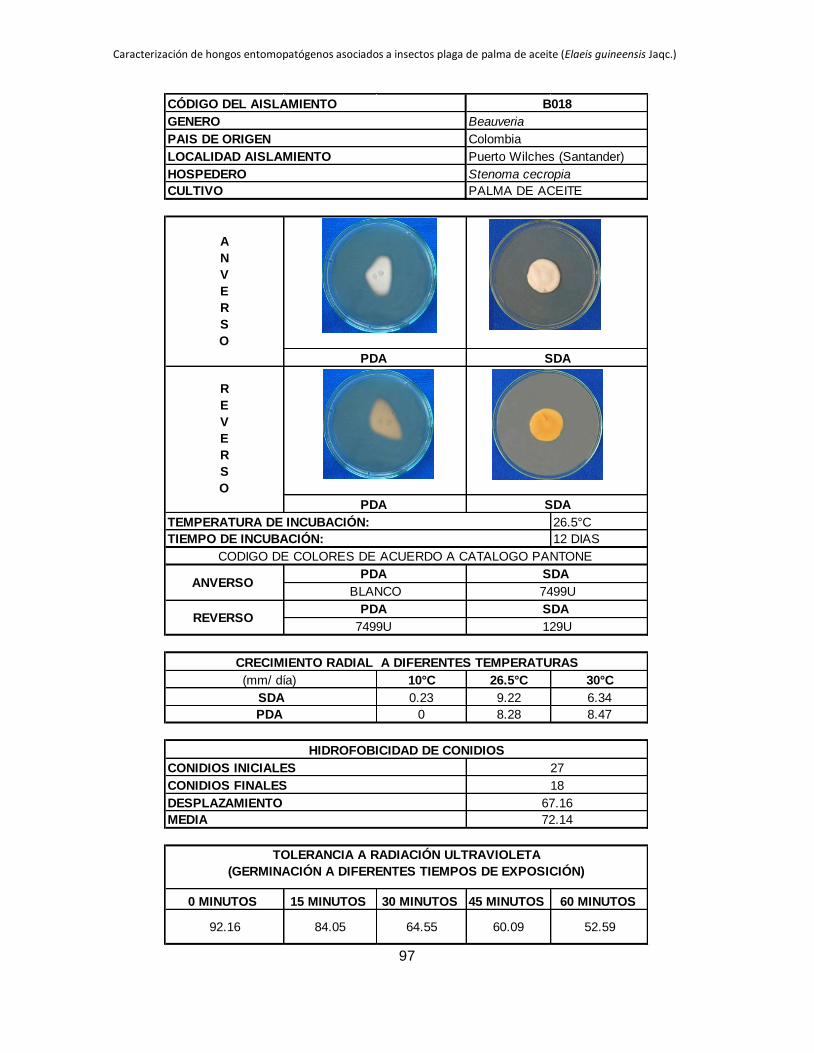
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
97
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.23 9.22 6.34
0 8.28 8.47
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
52.5960.0964.5584.0592.16
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B018
Beauveria
Colombia
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
Puerto Wilches (Santander)
Stenoma cecropia
PALMA DE ACEITE
PDA SDA
SDA
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 27
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
ANVERSO
REVERSO
BLANCO 7499U
MEDIA 72.14
DESPLAZAMIENTO 67.16
CONIDIOS FINALES 18
PDA SDA
7499U 129U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
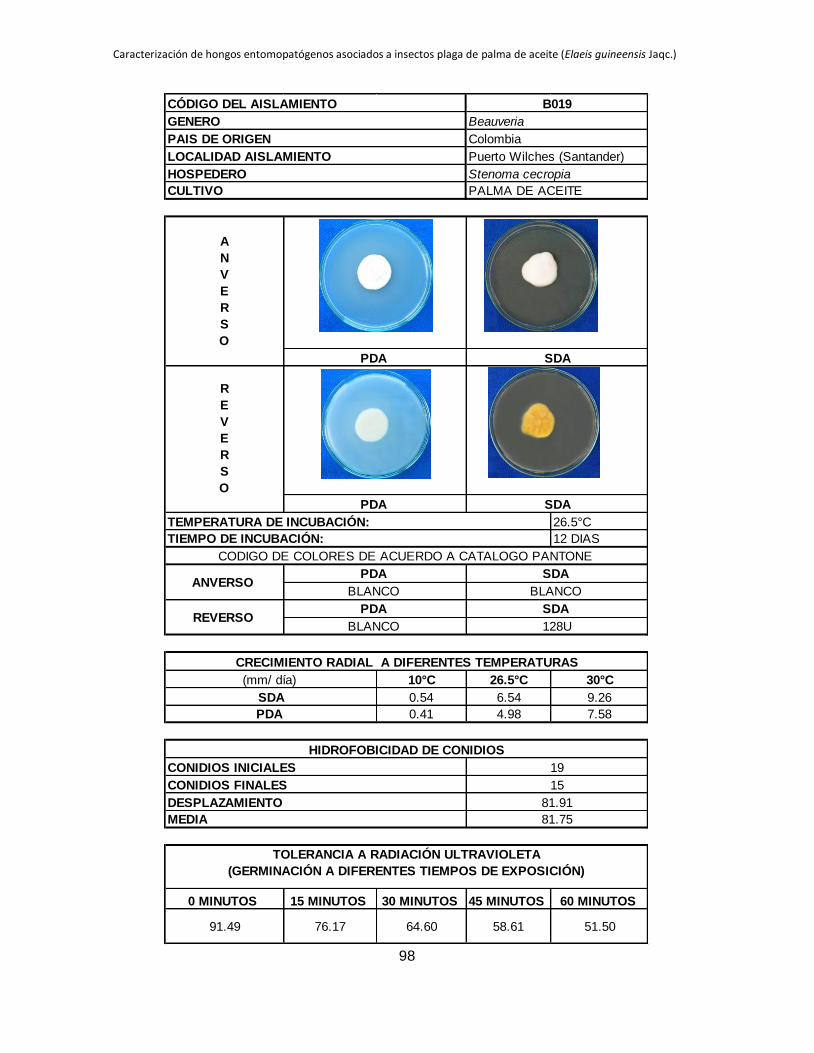
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
98
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.54 6.54 9.26
0.41 4.98 7.58
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
PALMA DE ACEITE
B019
Beauveria
Colombia
Puerto Wilches (Santander)
Stenoma cecropia
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
51.5058.6164.6076.1791.49
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
BLANCO
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA SDA
BLANCO BLANCO
128U
ANVERSO
REVERSO
MEDIA 81.75
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
DESPLAZAMIENTO 81.91
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 19
CONIDIOS FINALES 15
PDA
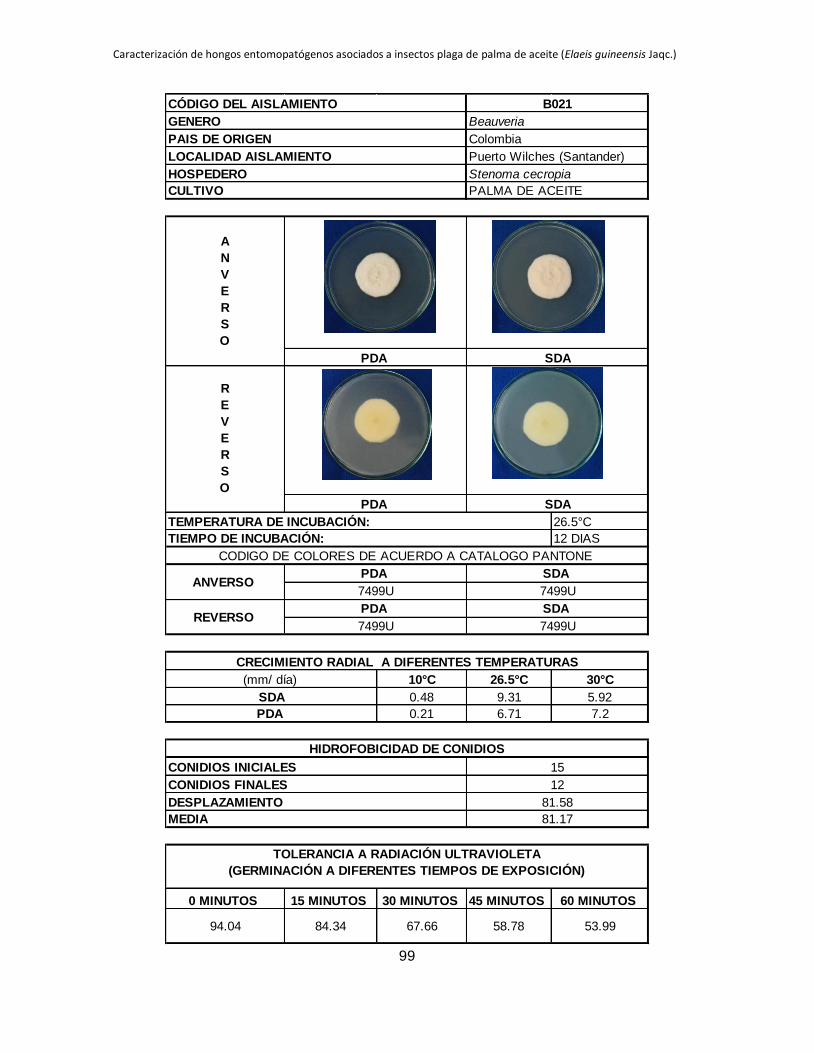
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
99
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.48 9.31 5.92
0.21 6.71 7.2
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
Puerto Wilches (Santander)
Stenoma cecropia
PALMA DE ACEITE
94.04 84.34 53.9958.7867.66
CÓDIGO DEL AISLAMIENTO
GENERO
PAIS DE ORIGEN
B021
Beauveria
Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
PDA SDA
7499U 7499U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES
MEDIA 81.17
CONIDIOS FINALES 12
ANVERSO
REVERSO
15
7499U 7499U
DESPLAZAMIENTO 81.58
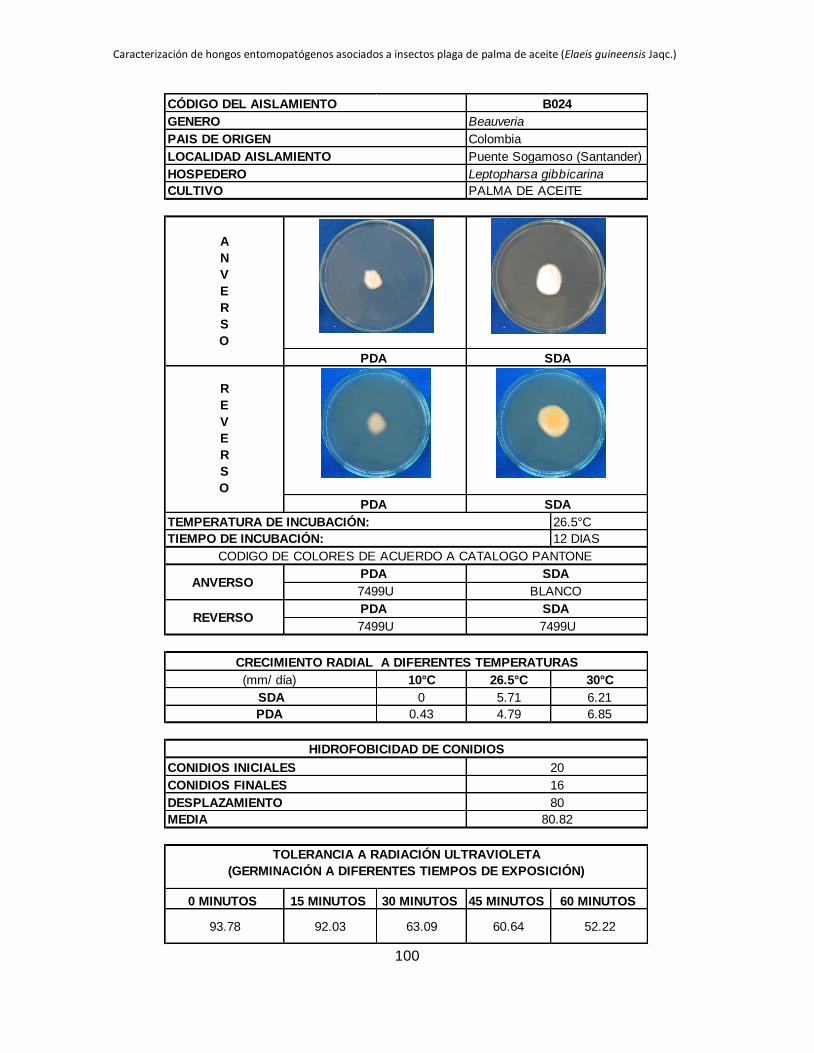
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
100
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 5.71 6.21
0.43 4.79 6.85
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
93.78 52.2260.6463.0992.03
CÓDIGO DEL AISLAMIENTO B024
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puente Sogamoso (Santander)
HOSPEDERO Leptopharsa gibbicarina
CULTIVO PALMA DE ACEITE
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
CONIDIOS FINALES 16
PDA SDA
7499U 7499U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
MEDIA 80.82
CONIDIOS INICIALES 20
ANVERSO
REVERSO
HIDROFOBICIDAD DE CONIDIOS
7499U BLANCO
DESPLAZAMIENTO 80
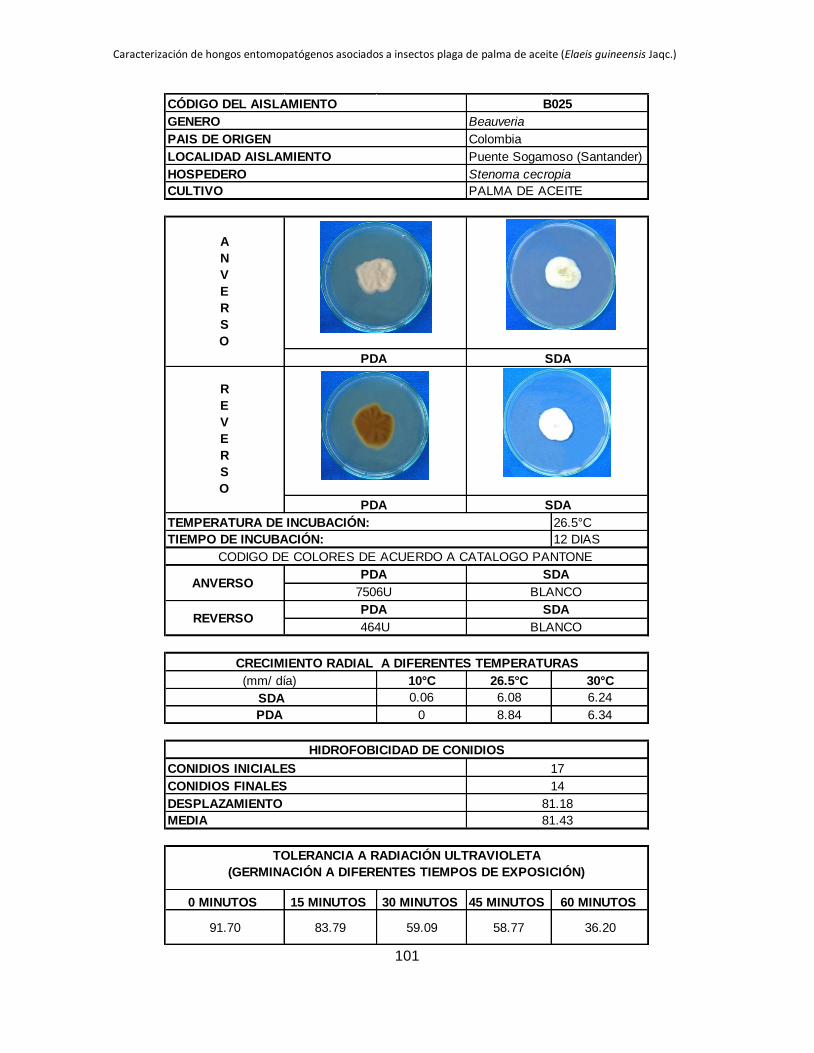
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
101
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.06 6.08 6.24
0 8.84 6.34
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
91.70 36.2058.7759.0983.79
CÓDIGO DEL AISLAMIENTO B025
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puente Sogamoso (Santander)
HOSPEDERO Stenoma cecropia
CULTIVO PALMA DE ACEITE
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
PDA SDA
464U BLANCO
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES
MEDIA 81.43
CONIDIOS FINALES 14
ANVERSO
REVERSO
17
7506U BLANCO
DESPLAZAMIENTO 81.18
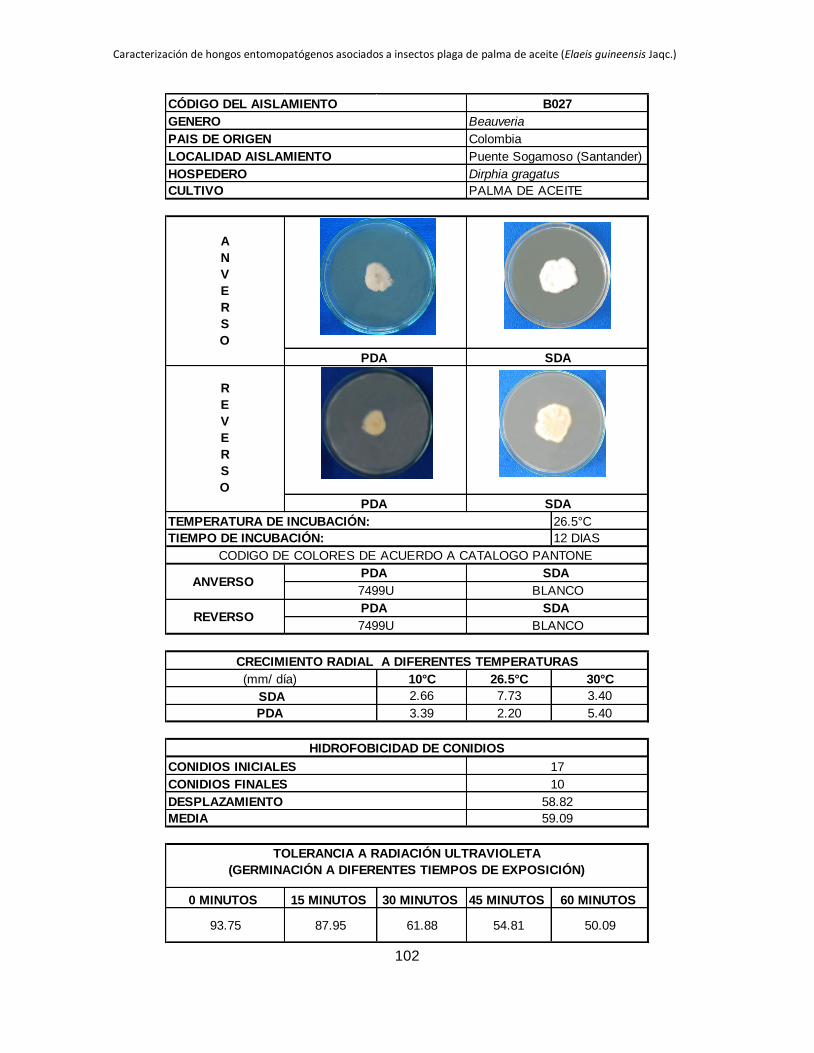
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
102
Puente Sogamoso (Santander)
26.5°C
12 DIAS
10°C 26.5°C 30°C
2.66 7.73 3.40
3.39 2.20 5.40
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B027
GENERO Beauveria
PAIS DE ORIGEN Colombia
LOCALIDAD AISLAMIENTO
HOSPEDERO Dirphia gragatus
CULTIVO PALMA DE ACEITE
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
PDA SDA
7499U BLANCO
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 17
7499U BLANCO
REVERSO
93.75 87.95 61.88 54.81 50.09
DESPLAZAMIENTO 58.82
MEDIA 59.09
CONIDIOS FINALES 10
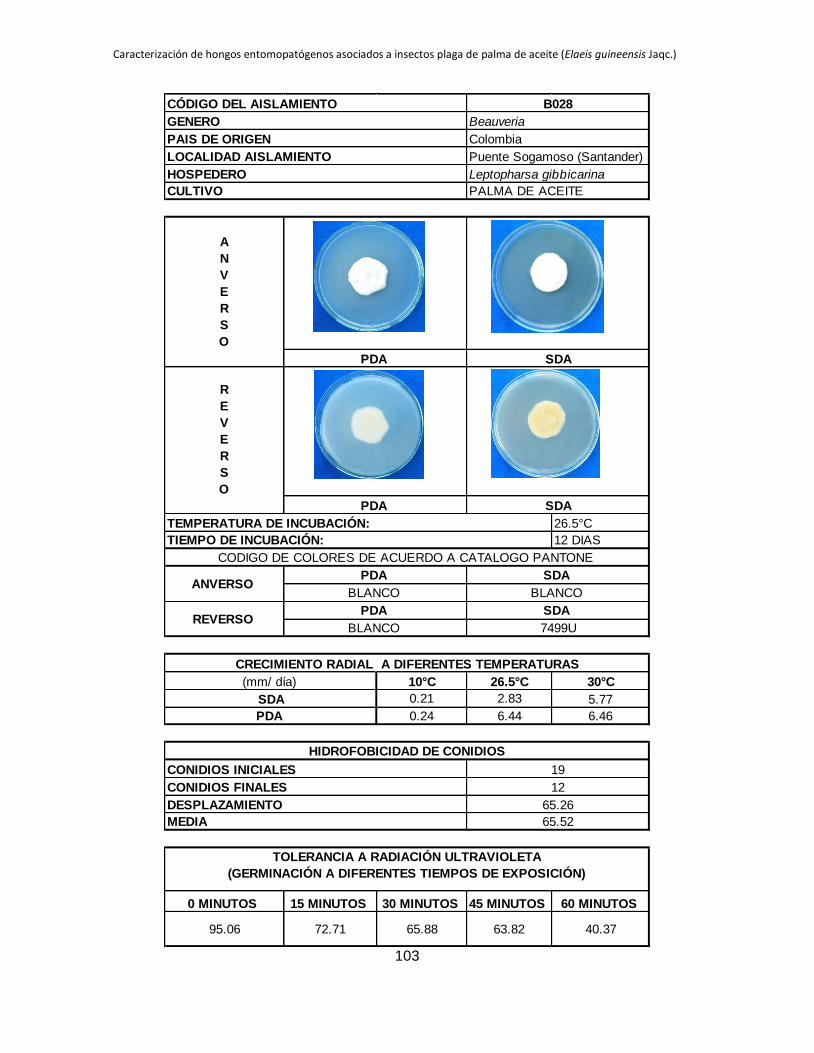
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
103
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.21 2.83 5.77
0.24 6.44 6.46
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B028
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puente Sogamoso (Santander)
HOSPEDERO Leptopharsa gibbicarina
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 12
PDA SDA
BLANCO 7499U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 19
BLANCO BLANCO
REVERSO
MEDIA 65.52
95.06 40.3763.8265.8872.71
DESPLAZAMIENTO 65.26
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
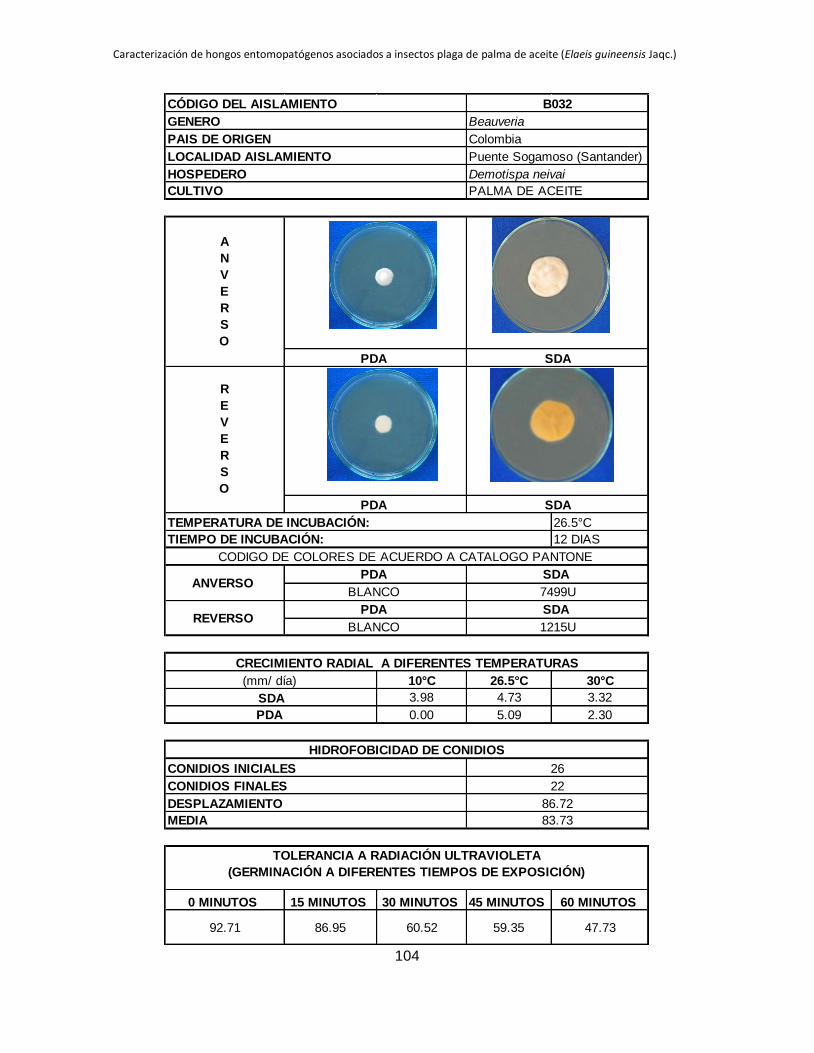
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
104
26.5°C
12 DIAS
10°C 26.5°C 30°C
3.98 4.73 3.32
0.00 5.09 2.30
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B032
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puente Sogamoso (Santander)
HOSPEDERO Demotispa neivai
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 22
PDA SDA
BLANCO 1215U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 26
BLANCO 7499U
REVERSO
MEDIA 83.73
47.7359.3560.5286.9592.71
DESPLAZAMIENTO 86.72
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
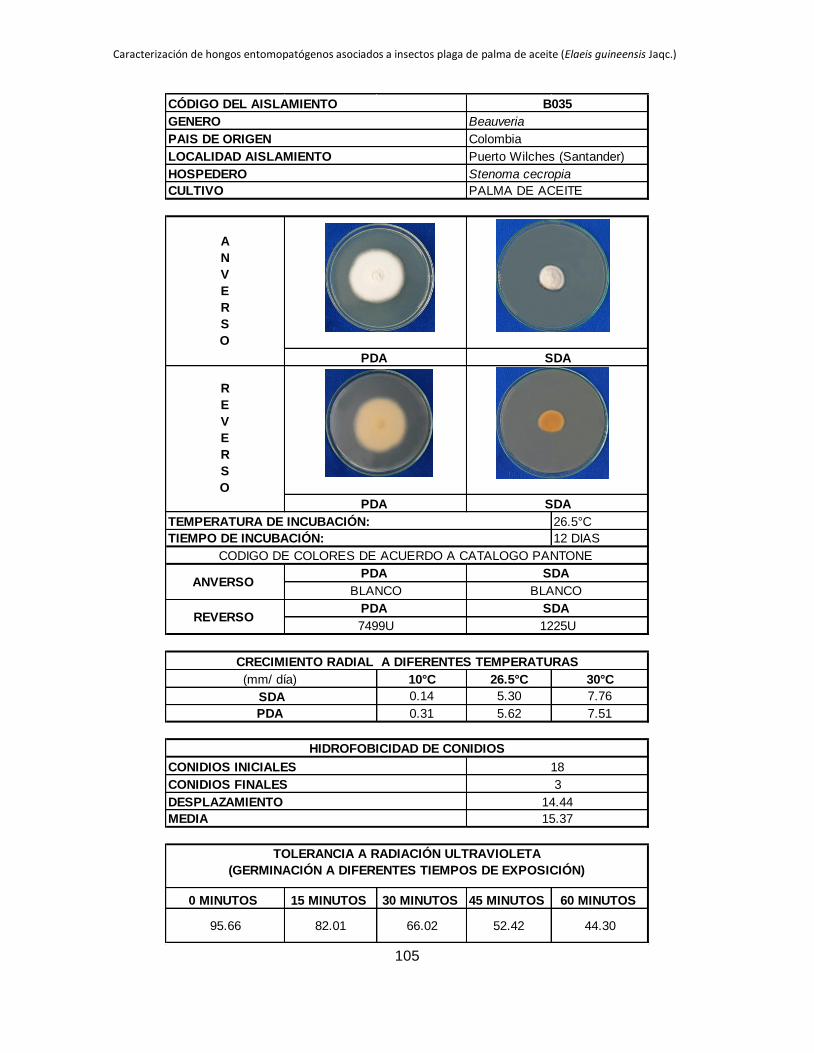
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
105
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.14 5.30 7.76
0.31 5.62 7.51
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B035
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puerto Wilches (Santander)
HOSPEDERO Stenoma cecropia
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 3
PDA SDA
7499U 1225U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 18
BLANCO BLANCO
REVERSO
MEDIA 15.37
44.3052.4266.0282.0195.66
DESPLAZAMIENTO 14.44
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
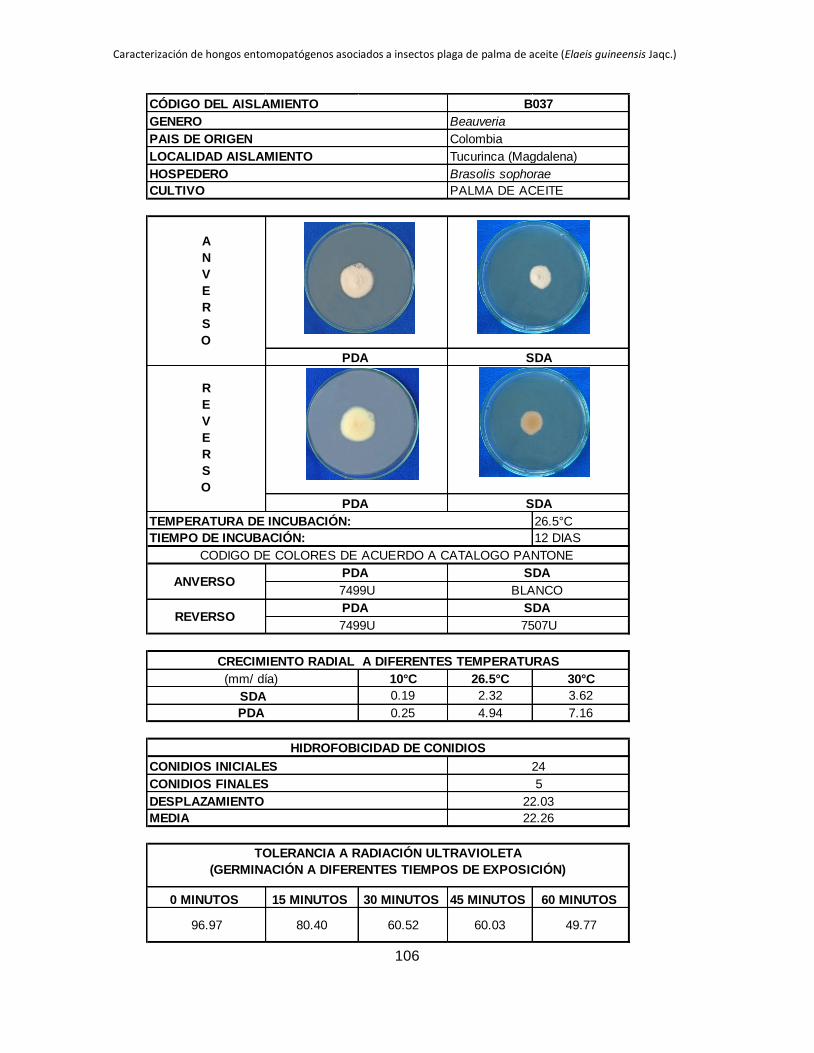
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
106
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.19 2.32 3.62
0.25 4.94 7.16
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B037
GENERO Beauveria
PAIS DE ORIGEN Colombia
LOCALIDAD AISLAMIENTO Tucurinca (Magdalena)
HOSPEDERO Brasolis sophorae
CULTIVO PALMA DE ACEITE
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
PDA SDA
7499U 7507U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 24
7499U BLANCO
REVERSO
49.7760.0360.5280.4096.97
DESPLAZAMIENTO 22.03
MEDIA 22.26
CONIDIOS FINALES 5
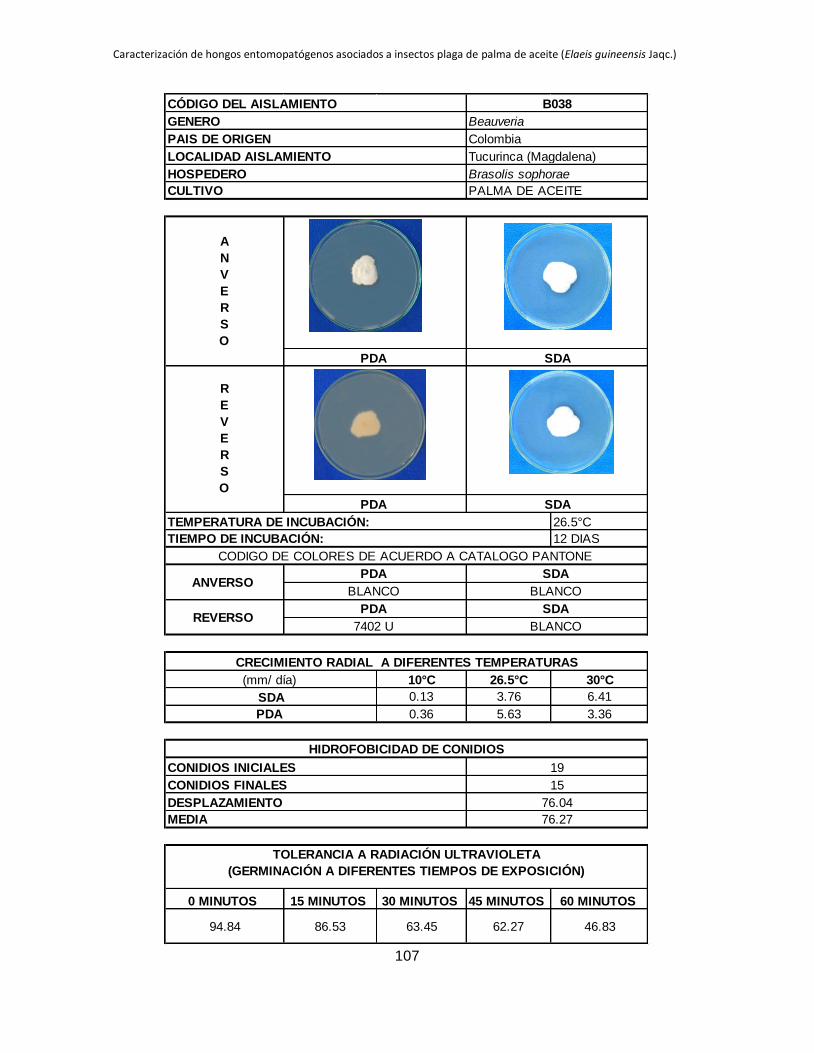
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
107
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.13 3.76 6.41
0.36 5.63 3.36
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B038
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Tucurinca (Magdalena)
HOSPEDERO Brasolis sophorae
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 15
PDA SDA
7402 U BLANCO
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 19
BLANCO BLANCO
REVERSO
MEDIA 76.27
46.8362.2763.4586.5394.84
DESPLAZAMIENTO 76.04
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
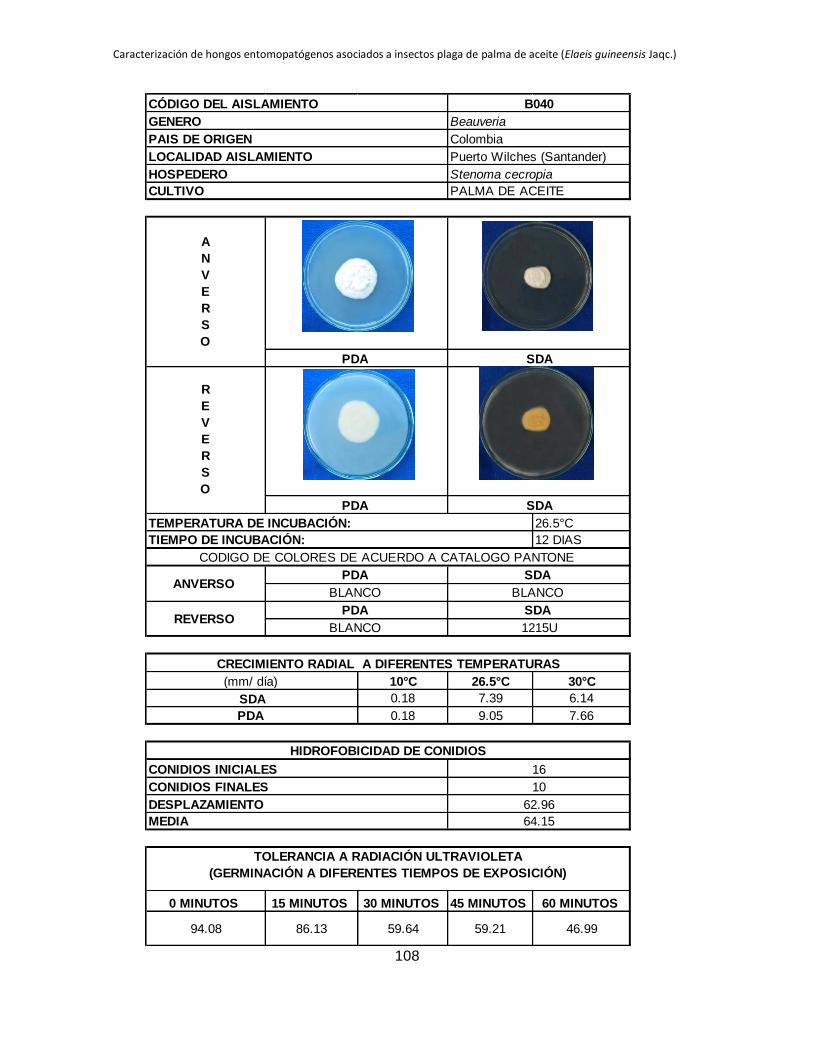
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
108
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.18 7.39 6.14
0.18 9.05 7.66
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B040
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puerto Wilches (Santander)
HOSPEDERO Stenoma cecropia
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 10
PDA SDA
BLANCO 1215U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 16
BLANCO BLANCO
REVERSO
MEDIA 64.15
46.9959.2159.6486.1394.08
DESPLAZAMIENTO 62.96
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
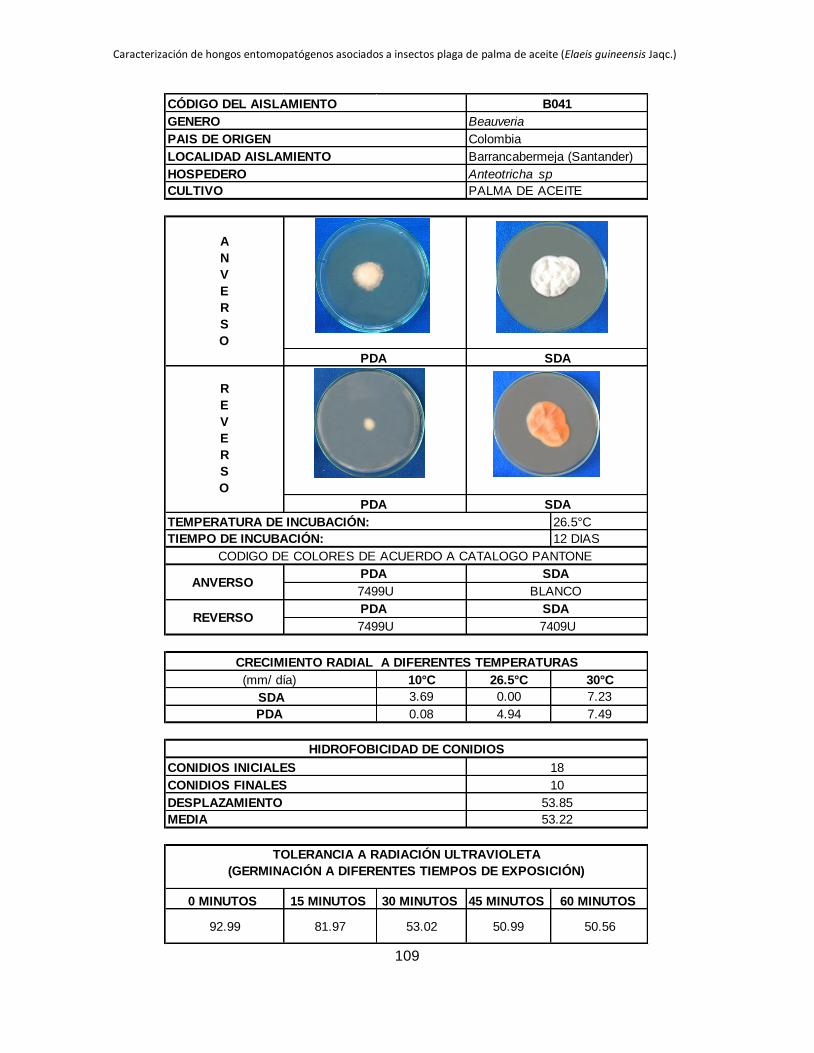
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
109
26.5°C
12 DIAS
10°C 26.5°C 30°C
3.69 0.00 7.23
0.08 4.94 7.49
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B041
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Barrancabermeja (Santander)
HOSPEDERO Anteotricha sp
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 10
PDA SDA
7499U 7409U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 18
7499U BLANCO
REVERSO
MEDIA 53.22
50.5650.9953.0281.9792.99
DESPLAZAMIENTO 53.85
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
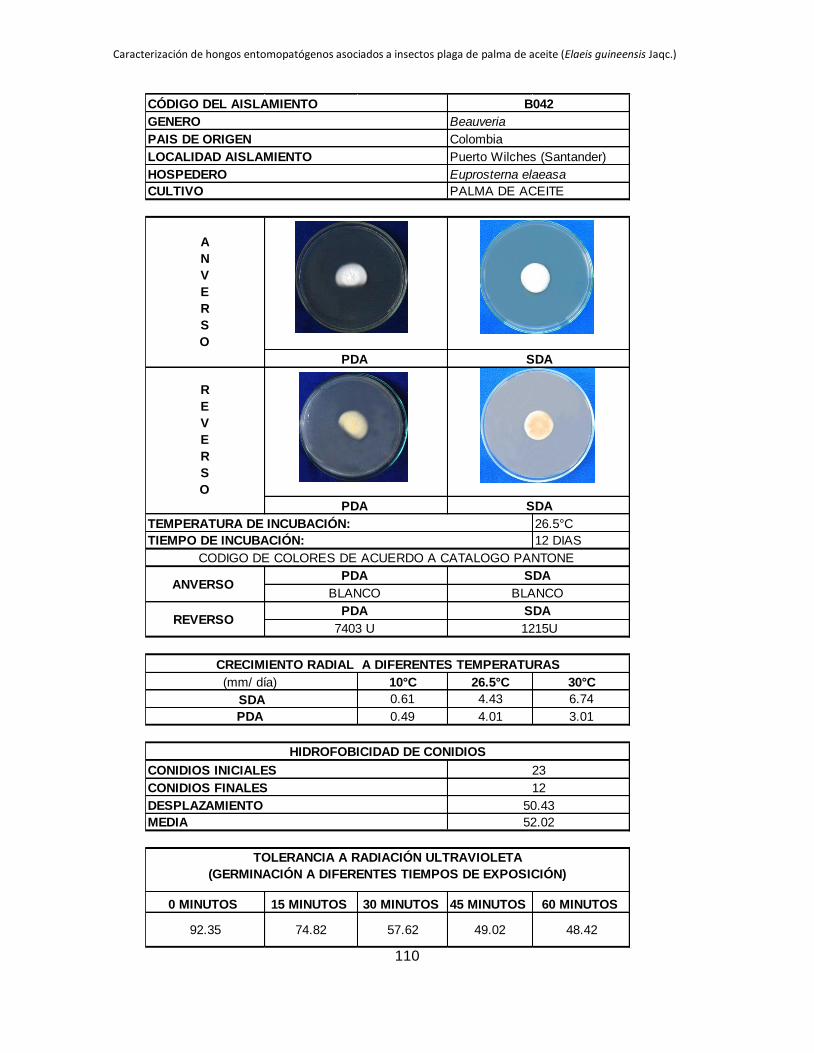
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
110
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.61 4.43 6.74
0.49 4.01 3.01
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B042
GENERO Beauveria
PAIS DE ORIGEN Colombia
LOCALIDAD AISLAMIENTO Puerto Wilches (Santander)
HOSPEDERO Euprosterna elaeasa
CULTIVO PALMA DE ACEITE
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
PDA SDA
7403 U 1215U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 23
BLANCO BLANCO
REVERSO
92.35 48.4249.0257.6274.82
DESPLAZAMIENTO 50.43
MEDIA 52.02
CONIDIOS FINALES 12
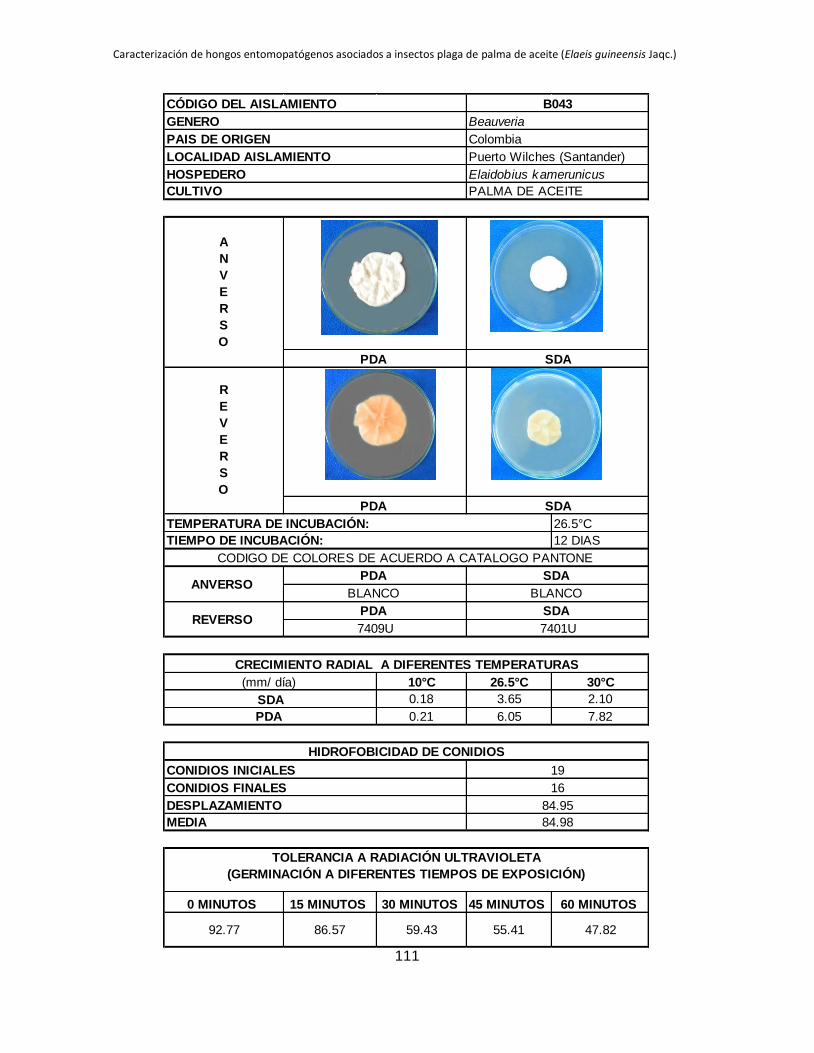
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
111
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.18 3.65 2.10
0.21 6.05 7.82
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
LOCALIDAD AISLAMIENTO Puerto Wilches (Santander)
HOSPEDERO Elaidobius kamerunicus
CÓDIGO DEL AISLAMIENTO B043
GENERO Beauveria
PAIS DE ORIGEN Colombia
CULTIVO PALMA DE ACEITE
PDA SDA
SDA ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
(mm/ día)
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
7409U 7401U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
REVERSO
BLANCO BLANCO
PDA SDA
92.77 86.57 47.8255.4159.43
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 19
CONIDIOS FINALES 16
DESPLAZAMIENTO 84.95
MEDIA 84.98
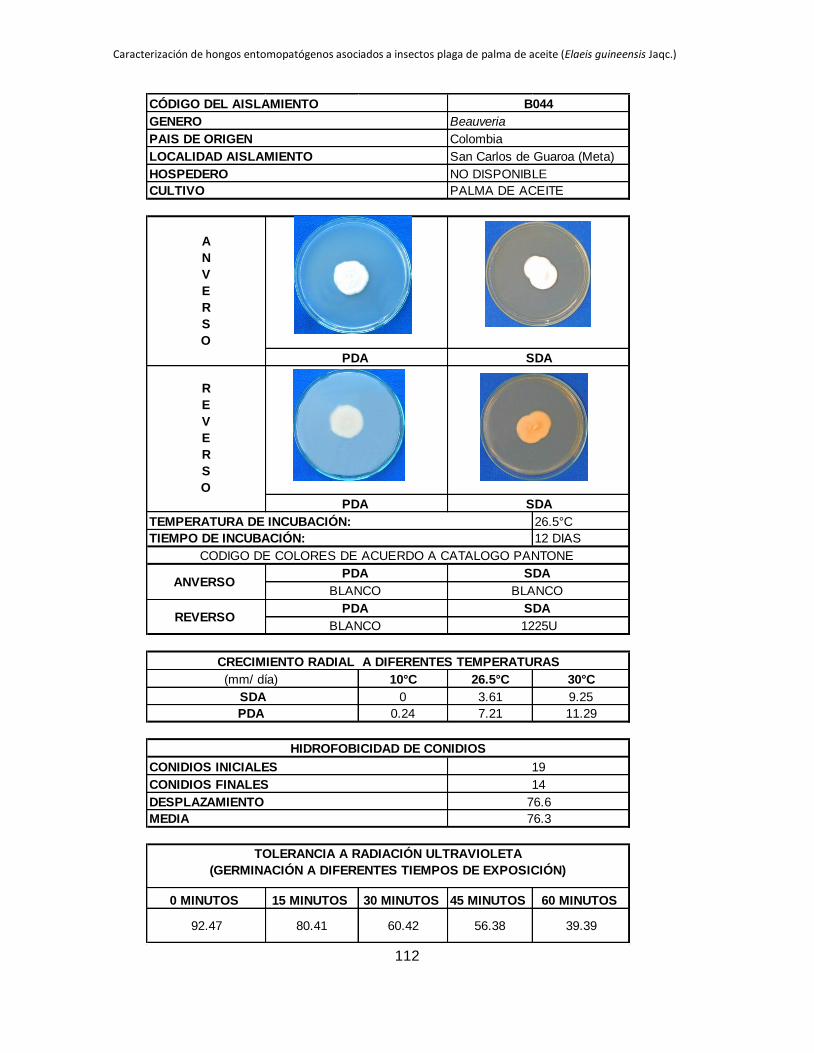
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
112
San Carlos de Guaroa (Meta)
NO DISPONIBLE
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 3.61 9.25
0.24 7.21 11.29
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B044
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO
HOSPEDERO
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 14
PDA SDA
BLANCO 1225U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 19
BLANCO BLANCO
REVERSO
MEDIA 76.3
DESPLAZAMIENTO 76.6
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
39.3956.3860.4280.4192.47
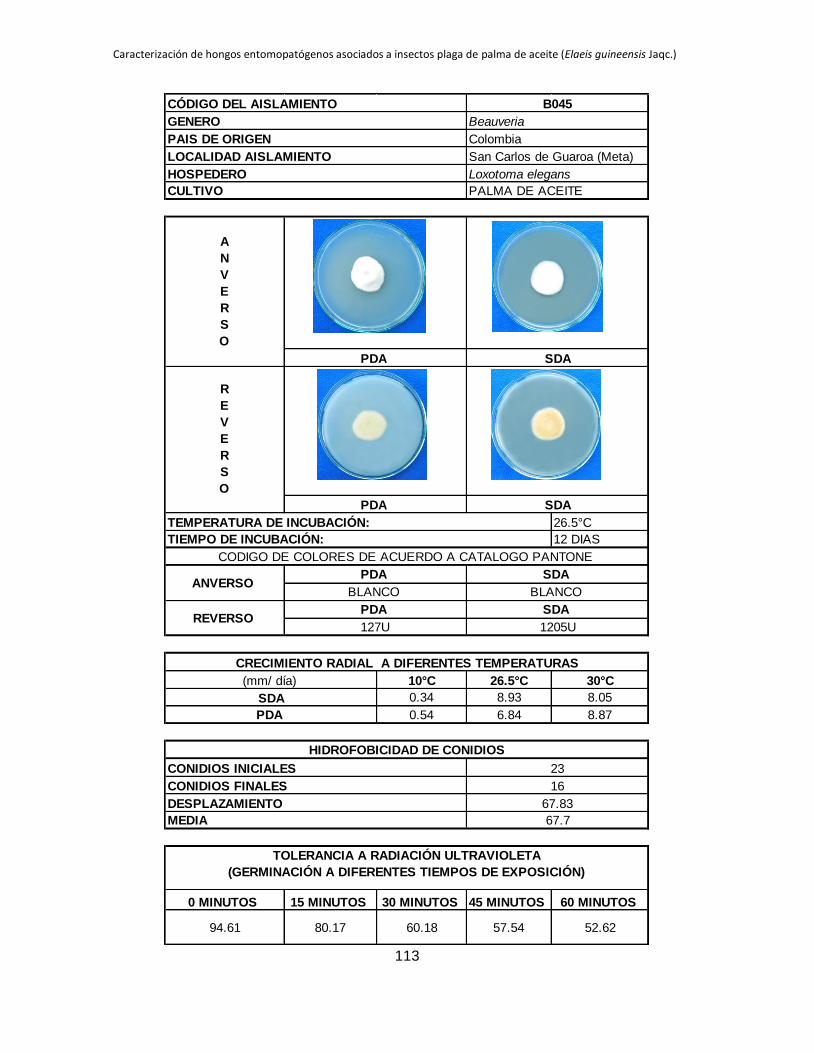
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
113
26.5°C
12 DIAS
10°C 26.5°C 30°C
0.34 8.93 8.05
0.54 6.84 8.87
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO B045
GENERO Beauveria
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO San Carlos de Guaroa (Meta)
HOSPEDERO Loxotoma elegans
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 16
PDA SDA
127U 1205U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 23
BLANCO BLANCO
REVERSO
MEDIA 67.7
52.6257.5460.1880.1794.61
DESPLAZAMIENTO 67.83
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
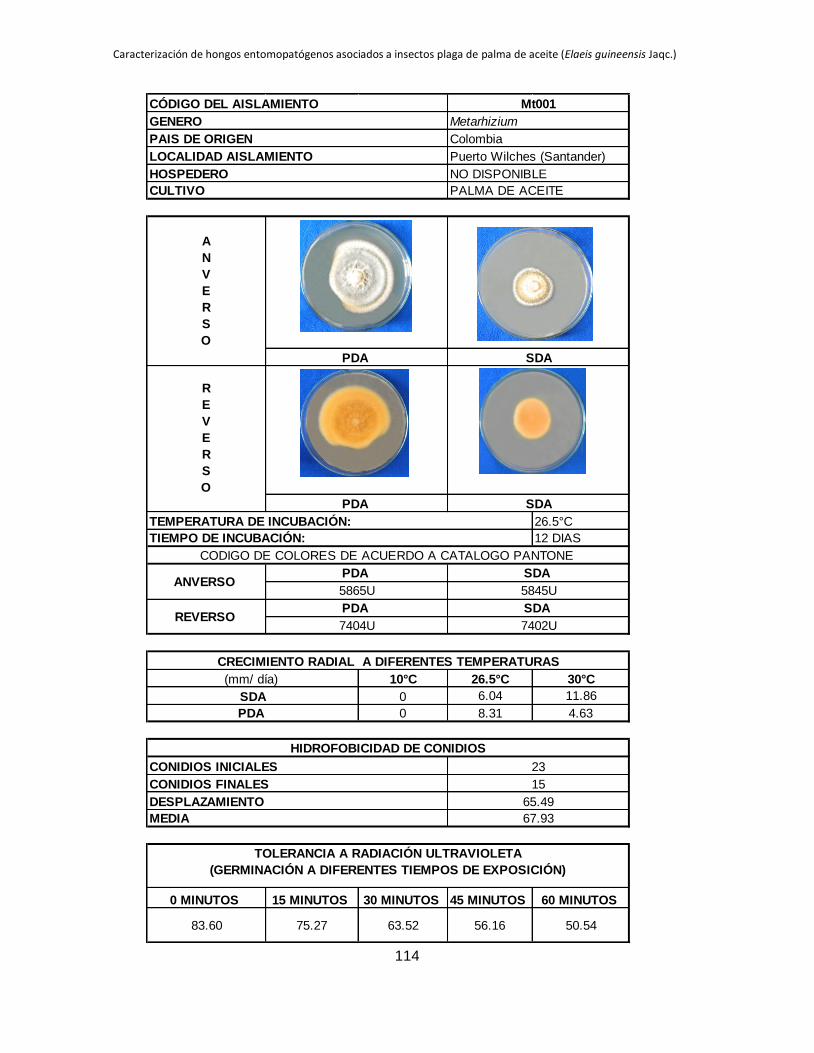
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
114
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 6.04 11.86
0 8.31 4.63
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO Mt001
GENERO Metarhizium
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Puerto Wilches (Santander)
HOSPEDERO NO DISPONIBLE
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 15
PDA SDA
7404U 7402U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 23
5865U 5845U
REVERSO
MEDIA 67.93
50.5456.1663.5275.2783.60
DESPLAZAMIENTO 65.49
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
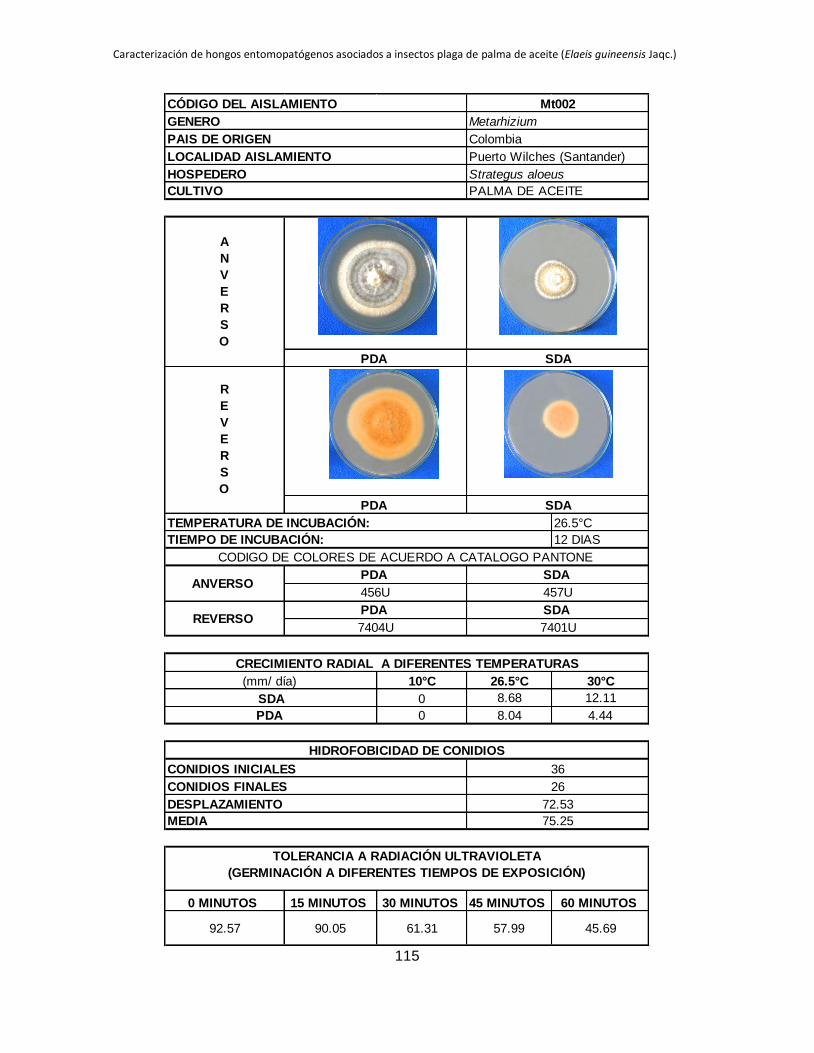
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
115
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 8.68 12.11
0 8.04 4.44
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
SDA
A
N
V
E
R
S
O
ANVERSO 457U
CÓDIGO DEL AISLAMIENTO Mt002
GENERO Metarhizium
PAIS DE ORIGEN Colombia
LOCALIDAD AISLAMIENTO Puerto Wilches (Santander)
HOSPEDERO Strategus aloeus
CULTIVO PALMA DE ACEITE
REVERSO
456U
PDA
PDA SDA
SDA
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
R
E
V
E
R
S
O
PDA
75.25
CONIDIOS FINALES 26
(mm/ día)
SDA
PDA
SDA
7404U 7401U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
92.57 90.05 61.31
HIDROFOBICIDAD DE CONIDIOS
57.99 45.69
CONIDIOS INICIALES 36
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
DESPLAZAMIENTO 72.53
MEDIA
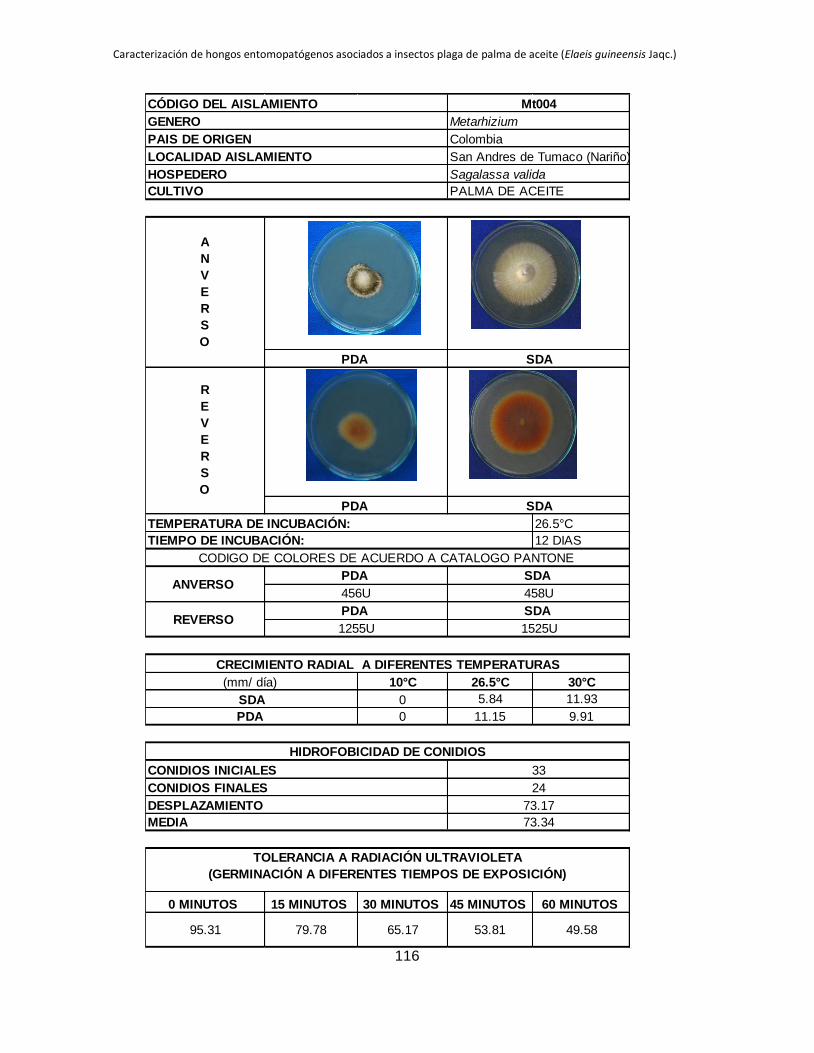
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
116
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 5.84 11.93
0 11.15 9.91
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO Mt004
GENERO Metarhizium
PAIS DE ORIGEN Colombia
LOCALIDAD AISLAMIENTO San Andres de Tumaco (Nariño)
HOSPEDERO Sagalassa valida
CULTIVO PALMA DE ACEITE
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
SDA
CONIDIOS INICIALES 33
PDA SDA
1255U 1525U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
456U 458U
REVERSO
49.5853.8165.1779.7895.31
DESPLAZAMIENTO 73.17
MEDIA 73.34
CONIDIOS FINALES 24
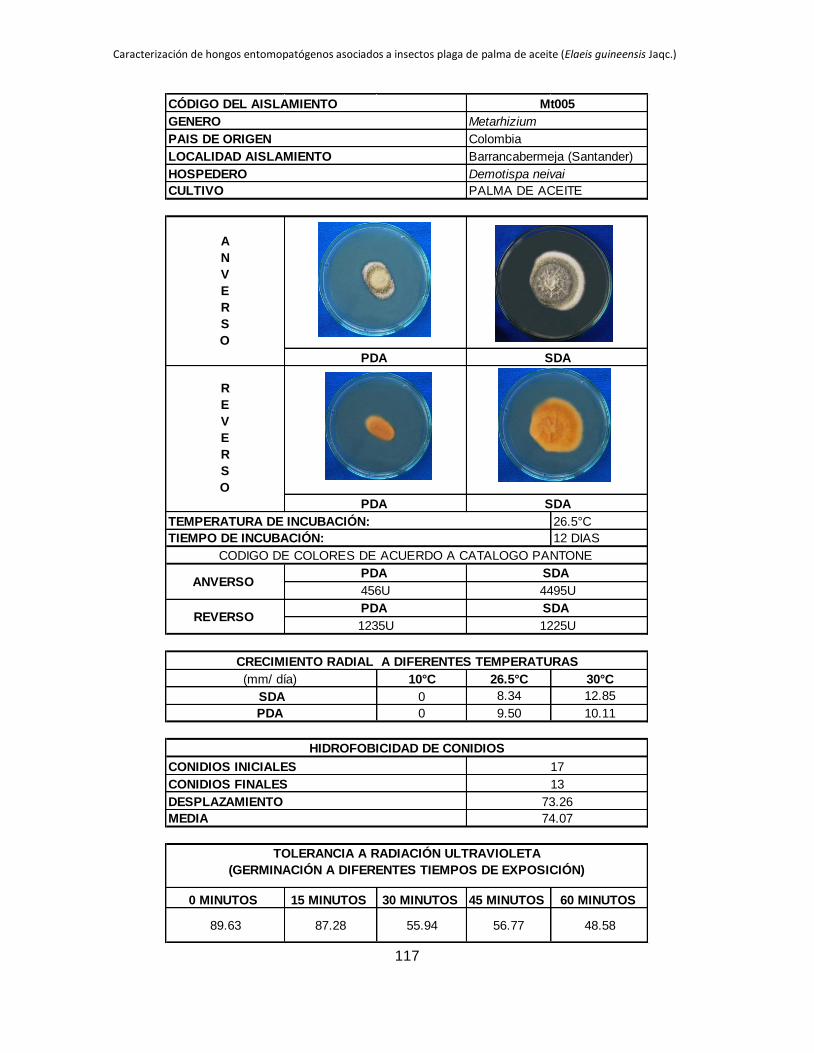
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
117
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 8.34 12.85
0 9.50 10.11
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO Mt005
GENERO Metarhizium
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Barrancabermeja (Santander)
HOSPEDERO Demotispa neivai
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 13
PDA SDA
1225U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 17
456U 4495U
REVERSO
MEDIA 74.07
48.5856.7755.9487.2889.63
DESPLAZAMIENTO 73.26
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
1235U
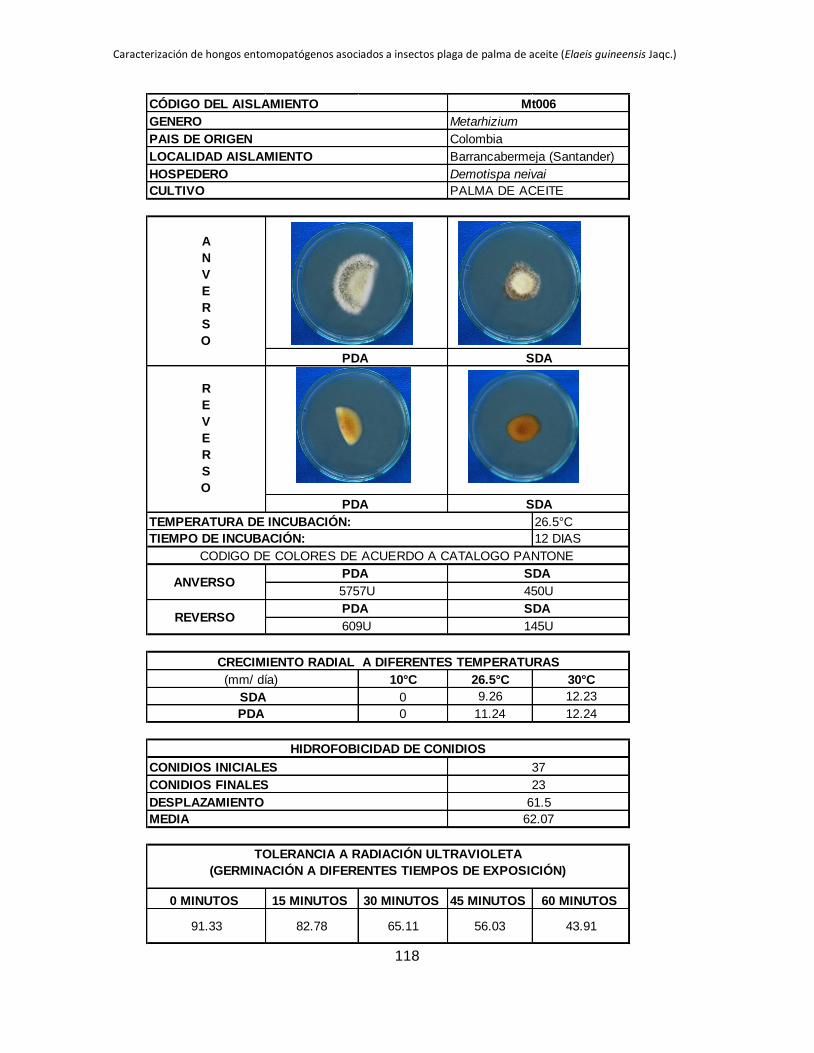
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
118
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 9.26 12.23
0 11.24 12.24
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO Mt006
GENERO Metarhizium
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Barrancabermeja (Santander)
HOSPEDERO Demotispa neivai
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 23
PDA SDA
609U 145U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 37
5757U 450U
REVERSO
MEDIA 62.07
91.33 82.78 65.11 56.03 43.91
DESPLAZAMIENTO 61.5
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
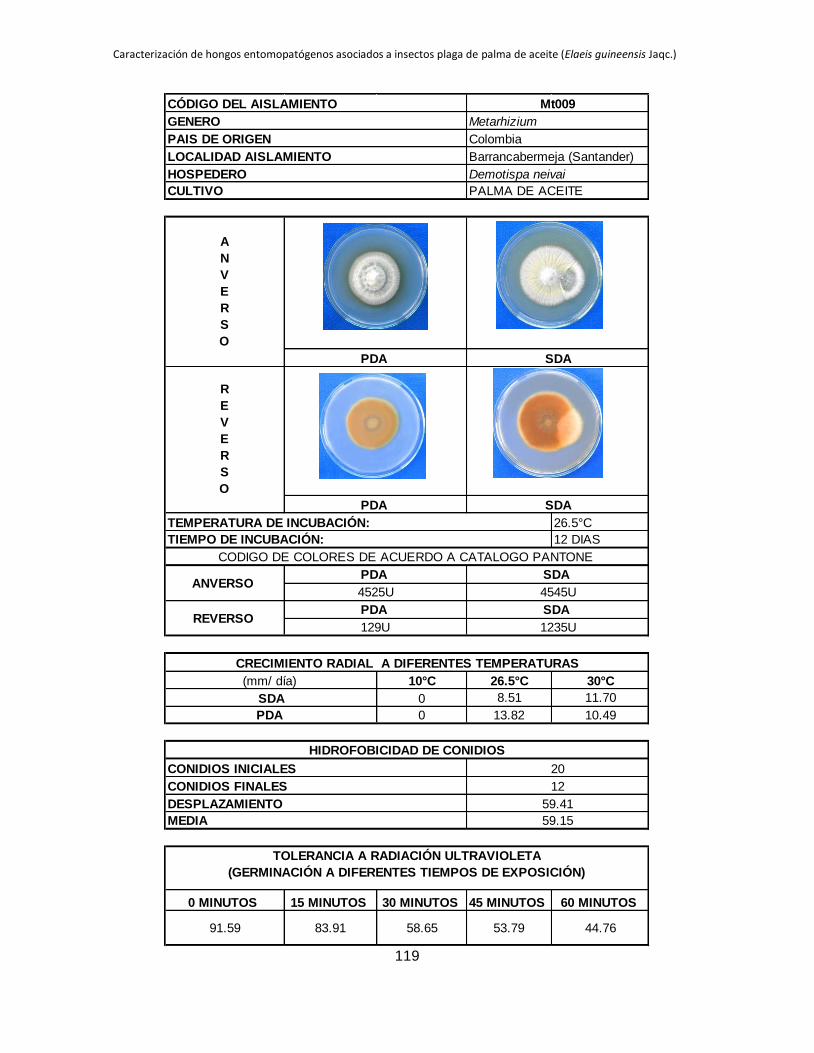
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
119
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 8.51 11.70
0 13.82 10.49
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO Mt009
GENERO Metarhizium
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Barrancabermeja (Santander)
HOSPEDERO Demotispa neivai
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 12
PDA SDA
129U 1235U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 20
4525U 4545U
REVERSO
MEDIA 59.15
44.7658.65 53.7983.9191.59
DESPLAZAMIENTO 59.41
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)
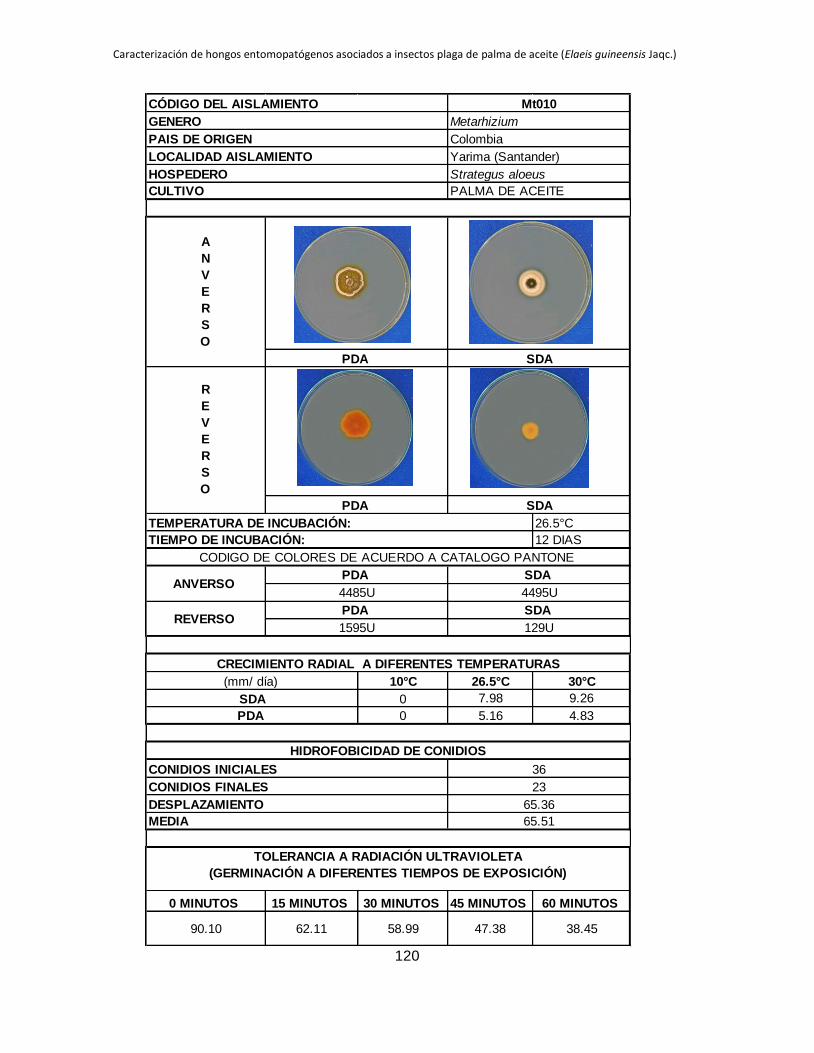
Caracterización de hongos entomopatógenos asociados a insectos plaga de palma de aceite (Elaeis guineensis Jaqc.)
120
26.5°C
12 DIAS
10°C 26.5°C 30°C
0 7.98 9.26
0 5.16 4.83
0 MINUTOS 15 MINUTOS 30 MINUTOS 45 MINUTOS 60 MINUTOS
CÓDIGO DEL AISLAMIENTO Mt010
GENERO Metarhizium
PAIS DE ORIGEN Colombia
TEMPERATURA DE INCUBACIÓN:
TIEMPO DE INCUBACIÓN:
CODIGO DE COLORES DE ACUERDO A CATALOGO PANTONE
PDA
LOCALIDAD AISLAMIENTO Yarima (Santander)
HOSPEDERO Strategus aloeus
CULTIVO PALMA DE ACEITE
ANVERSO
R
E
V
E
R
S
O
PDA SDA
A
N
V
E
R
S
O
PDA SDA
SDA
CONIDIOS FINALES 23
PDA SDA
1595U 129U
CRECIMIENTO RADIAL A DIFERENTES TEMPERATURAS
(mm/ día)
SDA
PDA
HIDROFOBICIDAD DE CONIDIOS
CONIDIOS INICIALES 36
4485U 4495U
REVERSO
MEDIA 65.51
90.10 62.11 38.4547.3858.99
DESPLAZAMIENTO 65.36
TOLERANCIA A RADIACIÓN ULTRAVIOLETA
(GERMINACIÓN A DIFERENTES TIEMPOS DE EXPOSICIÓN)