capture-elisa based on recombinant pr3 is sensitive for pr3–anca testing and allows detection of...
TRANSCRIPT

Ž .Journal of Immunological Methods 211 1998 111–123
Capture-ELISA based on recombinant PR3 is sensitive forPR3–ANCA testing and allows detection of PR3 and
PR3–ANCArPR3 immunecomplexes
Jiayan Sun a,c, David N. Fass b, Jay A. Hudson a, Margaret A. Viss a,Jorgen Wieslander d, Henry A. Homburger c, Ulrich Specks a,)¨
a Thoracic Diseases Research Unit, Mayo Clinic and Foundation, Rochester, MN, USAb Hematology Research Unit, Mayo Clinic and Foundation, Rochester, MN, USA
c DiÕision of Clinical Biochemistry and Immunology, Mayo Clinic and Foundation, Rochester, MN, USAd Wieslab, IDEON, S-223 70 Lund, Sweden
Received 15 August 1997; revised 31 October 1997; accepted 14 November 1997
Abstract
Ž . Ž .Proteinase 3 PR3 , a constituent of azurophil granules of neutrophils polymorphonuclear cells, PMNs , is the targetŽ . Ž .antigen for most anti-neutrophil cytoplasmic antibodies c-ANCA in Wegener’s granulomatosis WG . We have recentlyŽ .developed an expression system for recombinant PR3 rPR3 that is recognized by c-ANCA. Here, we report on the
Ž .development and characterization of two monoclonal antibodies moABs and a rabbit polyclonal antiserum generatedagainst this rPR3. Epitope competition analysis indicates that the moABs MCPR3-1 and MCPR3-2 recognize overlappingepitopes on the PR3 molecule that are distinct from the ones recognized by moABs 4A5 and 6A6 developed by others. SinceMCPR3-2 does not appear to compete for epitopes recognized by a sizable proportion of PR3–ANCA, we used it to develop
Ž .a sensitive capture enzyme linked immunosorbent assay ELISA for clinical PR3–ANCA testing. Both purified PMN–PR3Ž .and crude human mast cell line HMC-1 rPR3-S176A cell lysates were used as sources of PR3 target antigen in this assay
Ž . Ž .with equal analytical sensitivity and specificity. Of 109 patients with ANCA-associated disease, 91 83.5% and 90 82.6%were PR3–ANCA positive by capture ELISA when PMN–PR3 and HMC-1rPR3-S176A cell lysates were used as antigen,
Ž .respectively. When HMC-1rPR3 and HMC-1rPR3-S176A cells were used as indirect immunofluorescence IIF substrate,Ž . Ž .88 80.7% and 92 84.4% were PR3–ANCA positive, respectively. These differences were not statistically significant.
Only 1 of 151 controls without defined ANCA-associated disease tested positive by capture ELISA with either target antigenŽ .both negative by PR3–ANCA specific IIF . The capture ELISA can also be used to detect of PR3–ANCA immunecom-
Abbreviations: a1-PI, a1-protease inhibitor; ANCA, anti-neutrophil cytoplasmic antibodies; c-ANCA, cytoplasmic fluorescence patternANCA; ELISA, enzyme linked immunosorbent assay; GN, glomerulonephritis; IIF, indirect immunofluorescence; moAB, monoclonalantibody; NGS, normal goat serum; PBS, phosphate-buffered saline; PMN, polymorphonuclear cells; PR3, proteinase 3; rPR3, recombinantproteinase 3; RF, rheumatoid factor; RT, room temperature; WG, Wegener’s granulomatosis
) Corresponding author. Thoracic Diseases Research Unit, Guggenheim Bldg. 642 A, Mayo Clinic and Foundation, 200 First Street SW,Rochester, MN 55905, USA. Tel.: q1-507-284-2301; fax: q1-507-284-4521; e-mail: [email protected].
0022-1759r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0022-1759 97 00203-2

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123112
plexes and, in combination with the rabbit antiserum, for the quantitative measurement of PR3 in biological fluids. q 1998Elsevier Science B.V.
Keywords: ANCA; Capture ELISA; Human; Mast cell; Proteinase 3; Wegener’s granulomatosis
1. Introduction
Ž .Anti-neutrophil cytoplasmic antibodies ANCAwere first described in association with small vessel
Ž .vasculitis and Wegener’s granulomatosis WGŽDavies et al., 1982; Hall et al., 1984; van der
.Woude et al., 1985 . Based on the fluorescencepattern induced by ANCA on ethanol fixed neu-trophil cytospin preparations, they are currently cate-gorized in two broad groups: cytoplasmic staining
Ž .ANCA c-ANCA and perinuclear staining ANCAŽ . Žp-ANCA Falk and Jennette, 1988; Wiik and van
.der Woude, 1990 . A multitude of target antigens forANCA have been identified with various clinical
Ž .associations Kallenberg et al., 1994 . While newtarget antigens continue to be identified, proteinase 3Ž .PR3 and myeloperoxidase are the most clinicallyuseful because of their strong associations with WG
Ž .and microscopic polyangitis Kallenberg et al., 1994 .The neutrophil serine protease, PR3, has been identi-fied as the principal target antigen for c-ANCAŽJenne et al., 1990; Ludemann et al., 1990; Gupta et¨
.al., 1990 . Because of its high specificity for WG,the finding of a positive c-ANCA with PR3–ANCAspecificity has significant therapeutic implicationsŽ .Cohen Tervaert et al., 1989; Nolle et al., 1989 .¨
The search for simple PR3–ANCA detection sys-tems with high analytical sensitivity and specificityhas been hampered by properties of PR3–ANCA aswell as by properties of the target antigen: the major-ity of PR3–ANCA recognize conformational epi-
Ž .topes on PR3 Bini et al., 1992 and the purificationŽ .of PR3 from polymorphonuclear cell PMN gran-
Žules for use in solid-phase assays is tedious Kao et.al., 1988; Zhao and Lockwood, 1996 and prone to
result in partial loss of recognizability by at leastsome PR3–ANCA. The availability of an expression
Ž .system for recombinant PR3 rPR3 that can be usedas substrate for PR3–ANCA detection might offer analternative solution to some of these problems. Wehave recently reported on the development of anexpression system for conformationally intact, enzy-
matically active rPR3 expressed in the human mastŽ .cell line HMC-1 Specks et al., 1996 and we have
shown that these HMC-1rPR3 cells represent a sub-strate for PR3–ANCA detection by indirect immuno-
Ž .fluorescence IIF that appears superior to the use ofŽ .PMN as substrate Specks et al., 1997 .
Ž .The studies presented here were performed: 1 toŽ .characterize two monoclonal antibodies moABs ,
MCPR3-1 and MCPR3-2, and a polyclonal rabbitantiserum generated against rPR3 expressed in
Ž .HMC-1 cells; 2 to evaluate their application in acapture enzyme linked immunosorbent assayŽ .ELISA for the detection of PR3–ANCA andPR3–ANCArPR3 immune complexes, as well as
Ž .for the quantitative measurement of PR3; and 3 tocompare the use of lysates from HMC-1rPR3-S176A
Ž .cells and purified PR3 from neutrophil PMN gran-ules as antigens in the capture ELISA for PR3–ANCA detection.
2. Materials and methods
2.1. Materials
Unless specified otherwise, all materials werefrom Sigma, St. Louis, MO. Purified human neu-trophil PR3, elastase and cathepsin G were pur-chased from Athens Research and Technology,Athens, GA. HMC-1rPR3-S176A cells were cul-
Ž .tured as described elsewhere Specks et al., 1996 .These cells express an enzymatically inactive mutantof rPR3 with a substitution of the active site serine at
Ž .position 176 Campanelli et al., 1990 by alanineŽidentical to the designation PR3-S195A which isbased on the bovine chymotrypsinogen A numberingŽ ..Jenne et al., 1997 .
2.2. Indirect immunofluorescence
Ethanol fixed cytospin preparations of humanPMN, HMC-1rPR3, HMC-1rVEC and HMC-

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123 113
1rPR3-S176A cells were prepared as described else-Ž .where Specks et al., 1997 . Cytospin preparations
were incubated with primary antibodies for 30 min,washed three times in 1% normal goat serumrphos-
Ž .phate-buffered saline NGSrPBS for 5 min each,incubated with secondary antibodies for 30 min,washed again three times in 1% NGSrPBS andcovered with cover slips in 90% glycerolrPBS. Theentire procedure was performed at room temperatureŽ .RT .
2.3. Cyto-ELISA
A cyto-ELISA was developed to screen for anti-PR3 reactivity of hybridoma supernatants using the
Ž .moAB 4A3 Sommarin et al., 1995 as prototype.1=106 HMC-1rPR3 or HMC-1rVEC cells werewashed and suspended in 20 mM Tris–HCl, 0.5 M
Ž .NaCl, pH 7.5 wash buffer 1 and placed in mi-crotiter wells. After centrifugation of the plates at900=g, cells were fixed in 95% ethanol at 48C for 5min and air dried. Supernatants from clones to betested for reactivity with rPR3 were incubated inwells containing HMC-1rPR3 cells or HMC-1rVECcontrol cells for 60 min at RT. After washing threetimes with wash buffer 1, HRP-conjugated goat
Žanti-mouse IgG BioRad, Hercules, CA, cat a172-1011, diluted 1:400 in 20 mM Tris–HCl, 0.5 M
.NaCl, 0.5% BSA, pH 7.4 was incubated in the wellsfor 60 min at RT. After washing, 100 ml of 0.1%5-aminosalicylic acid, 0.01% H O solution were2 2
added as substrate for the color reaction which wasstopped after 4 min by addition of an equal volumeof 1 M NaOH. Absorbance was measured at 490 nmwavelength.
2.4. Generation of antibodies
For the generation of the moABs MCPR3-1 andMCPR3-2, mice were immunized with HMC-1rPR3cell granule extract. HMC-1rPR3 cells were dis-
Ž .rupted by nitrogen cavitation 350 psi=20 min asŽ .described for neutrophils Borregaard et al., 1983 .
The slow spin supernatant was layered over a discon-tinuous Percoll gradient and centrifuged at 20,000=
Žg for 15 min. Of the resulting three bands the
.lowest band was not detectable consistently , theupper band tested positive for PR3 by dot blot using
Ža rabbit PR3 anti-serum and the moAB 4A3 Som-.marin et al., 1995 . The PR3 containing granule
fraction was collected, the granules were disruptedby repeated freeze–thawing and emulsified with Fre-und’s adjuvant and used for immunization of themice. Mouse sera, the fusions, and the clones weretested for reactivity against rPR3 by comparingHMC-1rPR3 cells to sham-transfected HMC-1rVEC cells by IIF and by cyto-ELISA.
For the generation of the rabbit antiserum, HMC-1rPR3-S176A cells were lysed. After batch adsorp-tion of the lysate with the anion exchange resin
Ž .Accell-QMA Waters, Milford, MA , the supernatantwas further purified using cation exchange chro-
Žmatography on a Mono S column Pharmacia, Pis-.cataway, NJ . The bound proteins were eluted with a
ŽNaCl gradient 0 to 1 M, 0.1% Triton X-100, 0.05 M.acetate, pH 5.5 . The eluted fractions, showing PR3
reactivity by capture ELISA, were pooled and furtherpurified by reverse phase HPLC using a C3 columneluted with acetonitrile. The immunoreactive peakfractions were pooled, lyophilized, resuspended inPBS, emulsified with Freund’s adjuvant, and used asimmunogen. Rabbit serum samples were evaluatedfor reactivity with rPR3 between booster injectionsby IIF using HMC-1rPR3-S176A cells in compari-son to HMC-1rVEC cells. The IgG fraction of therabbit serum was prepared by adsorption to proteinA. Handling and care of the animals required forantibody production were in accordance with institu-tional guidelines.
2.5. Immunoblotting
Five million cells were lysed in 500 ml of lysisŽbuffer 50 mM Tris–HCl, 0.15 M NaCl, 0.5% Non-
idet-P 40, 0.5 mM EDTA, 75 mgrml PMSF, 2.mgrml aprotinin, 0.5 mgrml leupeptin, pH 7.2 .
Proteins were resolved on 12% SDS-PAGE gelsunder non-reducing conditions, transferred to Im-mun-Litee membranes, and analyzed using the Im-mun-Litee chemiluminescent assay according to the
Ž .manufacturer’s instructions BioRad . The mem-branes were incubated overnight at 48C with the

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123114
Ž .moAB MCPR3-2 0.7 mgrml diluted 1:100 in 1%non-fat dry milk, 20 mM Tris, 500 mM NaCl, 0.05%Tween-20, pH 7.5.
2.6. Capture ELISA
For PR3–ANCA testing, the following assay pro-Žcedure was used. Microtiter wells Immulon 1, Dy-.natech Laboratories, Chantilly, VA were incubatedŽwith 200 ml of MCPR3-2 solution 4 mgrml in Na
.bicarbonate buffer, pH 9.5 at 48C overnight. Afterwashing three times with 20 mM Tris–HCl, 0.5 M
Ž .NaCl, pH 7.5, 0.05% Tween-20 wash buffer 2 , 200Žml of purified neutrophil PR3 solution 0.0625
mgrml in 50 mM Tris–HCl, 0.1 M NaCl, 0.1%.BSA, pH 7.4 were incubated in the wells for 1 h at
RT. Control wells were incubated in parallel withbuffer alone. Cell lysates from HMC-1rPR3-S176Acells were used as an alternative antigen source.5=106 cells were lysed by incubation in 500 ml oflysis buffer for 30 min on ice. After careful shearingof the DNA, 200-ml aliquots of a 1:8 dilution of thecell lysate in lysis buffer were incubated in MCPR3-2coated wells for 1 h at RT. Control wells wereincubated in parallel with lysis buffer alone. Afterwashing three times with wash buffer 2, 200-mlaliquots of serum dilution were incubated for 1 h atRT, followed by three washes and incubation ofalkaline phosphatase conjugated goat anti-human IgGŽ .Sigma, A-9544 diluted 1:10,000 in 20 mM Tris–
ŽHCl, 0.5 M NaCl, 0.5% BSA, pH 7.5 dilution.buffer 1 . For the color reaction the phosphatase
substrate, p-nitrophenyl phosphate was used at aconcentration of 1 mgrml in 0.1 M Tris–HCl, 5 mMMgCl , 0.1 M NaCl, pH 9.5. The absorbance was2
determined after 30 min at a wavelength of 405 nm.For PR3–ANCA testing, serum samples were
Ž .tested at dilutions of 1:20 in dilution buffer 1 induplicate on wells coated with MCPR3-2 and anti-
Žgen neutrophil PR3 or HMC-1rPR3-S176A cell.lysate , and in parallel on control wells coated with
Ž .moAB alone containing no antigen background .Unless specified otherwise, net absorbance valuescalculated by subtraction of the background valuefrom the value obtained from wells containing cap-tured antigen are reported. Serum samples yieldingnet absorbance values of 0.100 or greater were con-sidered PR3–ANCA positive.
To identify the possible causes of high back-ground readings and ways to avoid them, the follow-ing measures were evaluated. Blocking of the wellswith 2% BSA, 5% milk, or 0.1% Tween-20 for 1 hat RT prior to incubation of the serum sample had noeffect on background. Consequently, a blocking stepwas not made part of the procedure. Preadsorption ofserum samples with mouse immunoglobulin or hu-man immunoglobulin also did not result in signifi-cant background reduction.
For the detection of PR3–ANCArPR3 immunecomplexes, serum samples were preadsorbed by in-cubation in MCPR3-2 coated wells for 1 h at RT.The supernatants were then reassayed in duplicate inantigen containing wells as described.
For the quantitative determination of PR3, thefollowing modification of the above protocol wasused. A total of 50 mM Tris–HCl, 0.1 M NaCl,0.1% BSA, pH 7.4 was used as wash buffer and forthe dilutions of PR3 and primary and secondaryantibodies. As primary antibody, the rabbit PR3-anti-serum was used at a dilution of 1:50. HRP-con-
Žjugated anti-rabbit IgG BioRad, cat a172–1013,.diluted 1:400 was used as secondary antibody. The
color reaction was obtained and quantified as de-scribed for the cyto-ELISA. Each data point wasassayed in duplicate.
2.7. Patient serum samples
In this study, we used aliquots of the same serumsamples from patients with ANCA-associated dis-ease and from control patients which had been previ-ously tested for c-ANCA using PMN cytospin prepa-rations and for PR3–ANCA by IIF using HMC-1rPR3 cell cytospin preparations and different com-
Ž . Ž .mercial ELISA kits Table 1 Specks et al., 1997 .Ž .Since the presence of rheumatoid factor RF can
cause problems in solid phase assays, we have alsoincluded 20 serum samples containing RF in thisstudy. In addition, 40 serum samples containinganti-ds-DNA antibodies were evaluated.
2.8. Statistical analysis
Pairwise comparisons of the agreement of testresults were performed using the McNemar’s test.

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123 115
Table 1Serum samples and PR3–ANCA test results in the various assays
Ž .Sample group n HMC-1rPR3-IIF Capture ELISA Capture ELISA HMC-1rPR3-S176AaŽ . Ž .positivern PMN–PR3 lysate positivern
Ž .positivern
Ž .Healthy volunteers 20 0r20 0r20 1r20Ž .Anti-MPO-positive 20 1r20 1r20 1r20
Ž .ANA-positive 20 0r20 0r20 0r20Ž .RF-positive 20 0r20 0r20 0r20
Ž .Anti-ds-DNA-positive 40 nd 0r40 0r40Ž .Other antibodies 20 0r20 0r20 0r20
Ž .AMA 7Ž .SMA 3Ž .LKM-1 3
Ž .Jo-1 3Ž .PCA 3
Ž .Reticulin 1Cytoplasmic staining on commercial 1r19 2r19 1r19
Ž .PMN slides 19Previously reported as 88r101 92r101 91r101
Ž .c-ANCA-positive 101
a Except for the RF containing samples the data listed in this column have been previously reported.AMA: anti-mitochondrial antibodies.LKM: anti-liver, kidney microsome antibodies.MPO: myeloperoxidase.PKA: anti-parietal cell antibodies.RF: rheumatoid factor.SMA: anti-smooth muscle antibodies.nd: Not done.
Significance of correlations were analyzed using theF-test for linear regression. In all cases, two-sidedp-valuesF0.05 were considered significant.
3. Results
3.1. Characterization of antibodies
Two moABs designated as MCPR3-1 andMCPR3-2 were selected based on their ability tobind to granule content of HMC-1rPR3 cells but notof cells transfected with the expression vector with-
Ž .out insert HMC-1rVEC cells . IgG was purifiedfrom mouse ascites and MCPR3-1 and MCPR3-2were identified to be of IgG1 subclass. Both antibod-ies react with PMN–PR3 as determined by IIF gen-erating the characteristic c-ANCA staining pattern on
Ž .ethanol fixed PMN not shown and by immunoblotswith purified PMN–PR3 and lysates from HMC-
Ž .1rPR3-S176A cells Fig. 1 . No cross-reactivity withpurified neutrophil elastase or cathepsin G was de-
Ž .tected by immunoblot or direct ELISA not shown .MCPR3-2 has been used to purify native PMN–PR3
Fig. 1. Western blot of purified PMN–PR3 and HMC-1rPR3-S176cell lysate. 5=106 HMC-1rPR3-S176A cells were lysed, aliquots
Ž 5of the cell lysates equivalent to 2.5=10 cells per lane, lanes 1. Ž .and 3 and purified PMN–PR3 1 mg, lane 2 were separated on a
12% polyacrylamide gel under non-reducing conditions. Proteinswere then transferred to filter membranes and probed with the
Ž .moAB MCPR3-2 lanes 1 and 2 and the rabbit polyclonalŽ .antiserum lane 3 .

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123116
from azurophil granule extracts by affinity chro-matography.
Neither antibody interfered with enzymatic activ-ity of rPR3 against the substrates MeSucc-Ala-Ala-Pro-Val-pNa or elastin-FITC, or with its complexa-
Ž . Ž .tion with a1-protease inhibitor a1-PI not shown .Epitope competition analysis was performed as
Ž .described Kwak and Yoon, 1996 using the cyto-ELISA. We compared our moABs raised against
rPR3 to the moABs 4A5, 6A6 and 4A3 raised againstŽ .PMN–PR3 Sommarin et al., 1995 . The results
indicate that MCPR3-1 and MCPR3-2 compete forŽ .binding to PR3 Fig. 2A , whereas the moABs 4A5
Ž .and 6A6 do not compete with MCPR3-1 not shownŽ .or MCPR3-2 Fig. 2B and C . The moAB 4A3
appears to bind to a larger epitope that partiallyŽoverlaps with those recognized by MCPR3-1 not
. Ž .shown and MCPR3-2 Fig. 2D .
Ž . Ž .Fig. 2. Epitope competition analysis of moABs by cyto-ELISA. A When MCPR3-1 small filled circles is bound to its epitope atŽ .saturation no signal enhancement is obtained by addition of MCPR3-2 large open circles indicating that the two antibodies compete for
Ž .overlapping binding sites. When MCPR3-2 large open squares is bound to its epitope at saturation only a small signal enhancement can beŽ .achieved by the addition of MCPR3-1 small full squares suggesting that the epitope recognized by MCPR3-1 may be slightly larger thanŽ . Ž .the epitope recognized by MCPR3-2. B When 4A5 large open triangles is bound to its target at saturation a significant enhancement of
Ž . Ž .absorbance can be achieved by addition of MCPR3-2 full small triangles . If MCPR3-2 is bound at saturation first large open squares anŽ .equivalent gain in absorbance can be obtained if 4A5 small full squares is added, indicating that the epitopes recognized by MCPR3-2 and
Ž . Ž .4A5 do not overlap. C When 6A6 is used at saturation crosses significant signal enhancement can be obtained by addition of MCPR3-2Ž . Ž .x and vice versa, indicating that MCPR3-2 and 6A6 also do not compete for the same epitope. D When MCPR3-2 is used at saturationŽ . Ž . Žlarge open squares , the addition of 4A3 results in signal enhancement small full squares . When 4A3 is used at saturation reached at
. Ž .dilution of 1:30; black bars the addition of MCPR3-2 white bars does not result in significant signal enhancement, indicating that 4A3Ž . Ž .occupies a larger overlapping epitope. Shown are representative examples of two D or three repeat experiments A–C .

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123 117
3.2. Detection of PR3–ANCA
The experiments of this study were designed toŽ .address the following questions: 1 Can the moAB
MCPR3-2 be used as capture antibody for PR3–ANCA testing, or does it compete for epitopes rec-
Ž .ognized by a sizable proportion of PR3–ANCA? 2Can HMC-1rPR3 cell lysates be used directly assource of antigen in this assay, avoiding the need forthe tedious purification procedure of PR3?
Because PR3 has been shown to cleave immuno-Ž .globulins and ANCA Dolman et al., 1995 , it is
possible that PR3–ANCA-test results obtained bycapture ELISA could be affected by the enzymaticactivity of PR3, particularly since the moABMCPR3-2 does not inhibit the enzymatic activity ofPR3. Therefore, we first determined whether theinactive mutant PR3-S176A expressed by HMC-
Ž .1rPR3-S176A cells Specks et al., 1996 is recog-nized by PR3–ANCA. Ethanol fixed cytospin prepa-rations of HMC-1rPR3, HMC-1rPR3-S176A andHMC-1rVEC control cells were prepared as de-
Ž .scribed Specks et al., 1997 . Sera from all 109patients with WG and MPA from our previously
Ž .reported patient cohort Specks et al., 1997 weretested by IIF at serum dilutions of 1:4 and 1:16 inparallel on all three substrates. As shown in Table 2,all PR3–ANCA sera that recognized wild type rPR3expressed in HMC-1rPR3 cells also recognized theinactivated mutant rPR3-S176A. In addition, rPR3-S176A was recognized by the remaining three serafrom patients with biopsy proven WG and one addi-
tional serum from a patient with clinically diagnosedWG, indicating that rPR3-S176A represents anequivalent substrate for PR3–ANCA testing to thewild type rPR3 expressed by HMC-1rPR3 cells.While these data might suggest a higher analyticalsensitivity of IIF using HMC-1rPR3-S176A cells assubstrate, the difference was not statistically signifi-
Ž .cant ps0.125 .Subsequently, we assayed all 200 serum samples
Ž .from the reported cohort Specks et al., 1997 , 20rheumatoid factor containing sera, and 40 anti-ds-DNA antibody containing sera in the capture ELISAusing both, purified PMN–PR3 and HMC-1rPR3-S176A cell lysates, as captured target antigen. Bothantigens captured by MCPR3-2 represent sensitive
Ž .substrates for PR3–ANCA testing Table 2 that isstatistically not significantly different from IIF test-
Žing using HMC-1 cells expressing rPR3 wild type.or inactive mutant . Only one serum sample from a
patient with biopsy proven WG was negative in thecapture ELISA when HMC-1rPR3-S176A cell lysate
Ž .was used as antigen background was low at 0.153 .This sample was positive in the capture ELISA withPMN–PR3 as antigen and by IIF using HMC-1rPR3and HMC-1rPR3-S176A cells as substrate. Onesample from a patient with the clinical diagnosis ofWG that had previously been positive by IIF usingHMC-1rPR3, HMC-1rPR3-S176A cells and PMN,tested PR3–ANCA negative in the capture ELISAwith either substrate.
With PMN–PR3 as antigen, one positive resultwas found in a patient with Q-fever and renal insuf-
Table 2PR3–ANCA testing by IIF using HMC-1rPR3 and HMC-1rPR3-S176A cell cytospin preparations and by capture ELISA with PMN–PR3and HMC-1rPR3-S176A cell lysate as captured antigen
Diagnosis n Positive on Positive by capture ELISA with
HMC-1rPR3 HMC-1rPR3-S176 PMN–PR3 HMC-1rPR3-S176 cell lysate
Biopsy proven WG 66 63 66 66 65Clinical WG 14 12 13 12 12MPA 29 13 13 13 13Total, n 109 88 92 91 90% positive 80.7 84.4 83.5 82.6
ap-value 0.125 1.0 0.5
aCalculated using McNemar’s test based on 2=2 cross-tabulation of test results.WG: Wegener’s granulomatosis.MPA: microscopic polyangiitis.GN: glomerulonephritis.
( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123118
Table 3Ž .Absorbance readings obtained from antigen containing wells, wells lacking antigen background and net absorbance values
Ž . Ž .Antigen PR3–ANCA status n Wells with antigen Background median range Net absorbance
PMN–PR3Ž . Ž . Ž . Ž .positive 95 0.752 0.264–1.413 0.221 0.086–0.851 0.424 0.123–1.179Ž . Ž . Ž . Ž .negative 125 0.257 0.082–1.740 0.317 0.090–1.764 y0.022 y0.409–0.094
HMC-1rPR3-S176A lysateŽ . Ž . Ž . Ž .positive 94 0.802 0.269–1.673 0.214 0.099–0.910 0.550 0.148–1.425Ž . Ž . Ž . Ž .negative 126 0.185 0.101–1.170 0.159 0.094–1.151 0.008 –0.148–0.094
ficiency of indeterminate etiology, whereas anotherserum from a healthy volunteer tested positive withHMC-1rPR3-S176A cell lysate as antigen. Both ofthese samples were ANCA negative by IIF on allsubstrates.
The medians and ranges of absolute, backgroundand net absorbance values obtained with the twoantigen preparations for PR3–ANCA positive andnegative serum samples are listed in Table 3. Back-ground absorbance of )0.500 occurred significantlymore frequently with PMN–PR3 than with HMC-1rPR3-S176A lysates as antigen: 42% vs. 8.4% ofPR3–ANCA negative samples and 8.4% vs. 6.4% ofPR3–ANCA positive samples yielded such values.While high background in PR3–ANCA positivesamples was mostly caused by the presence ofPR3–ANCArPR3 immunecomplexes, the cause ofhigh backgrounds in PR3–ANCA negative samplesremained unclear.
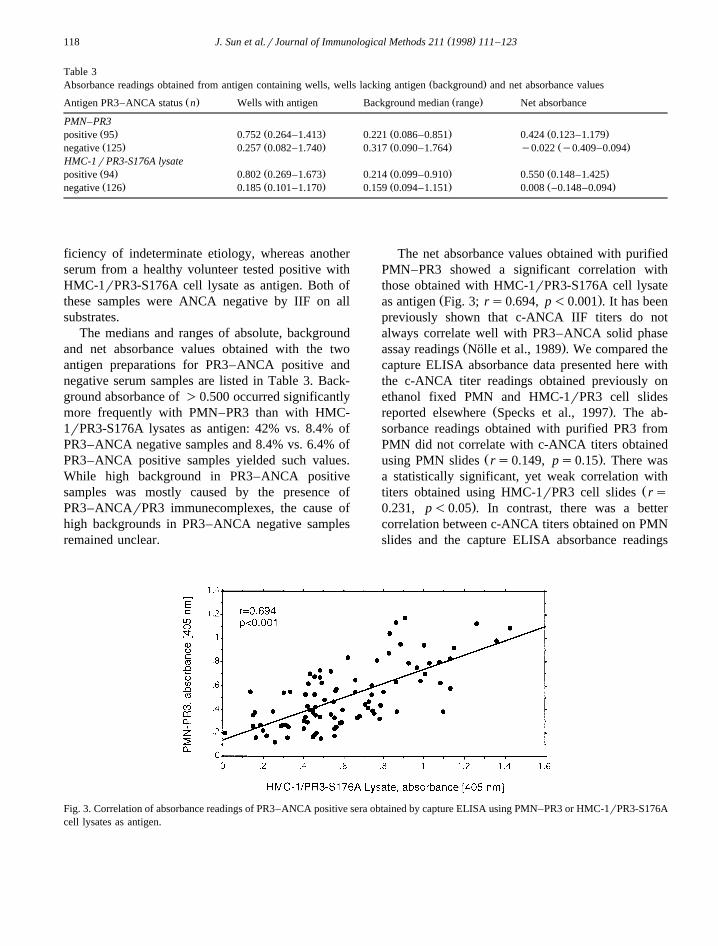
The net absorbance values obtained with purifiedPMN–PR3 showed a significant correlation withthose obtained with HMC-1rPR3-S176A cell lysate
Ž .as antigen Fig. 3; rs0.694, p-0.001 . It has beenpreviously shown that c-ANCA IIF titers do notalways correlate well with PR3–ANCA solid phase
Ž .assay readings Nolle et al., 1989 . We compared the¨capture ELISA absorbance data presented here withthe c-ANCA titer readings obtained previously onethanol fixed PMN and HMC-1rPR3 cell slides
Ž .reported elsewhere Specks et al., 1997 . The ab-sorbance readings obtained with purified PR3 fromPMN did not correlate with c-ANCA titers obtained
Ž .using PMN slides rs0.149, ps0.15 . There wasa statistically significant, yet weak correlation with
Žtiters obtained using HMC-1rPR3 cell slides rs.0.231, p-0.05 . In contrast, there was a better
correlation between c-ANCA titers obtained on PMNslides and the capture ELISA absorbance readings
Fig. 3. Correlation of absorbance readings of PR3–ANCA positive sera obtained by capture ELISA using PMN–PR3 or HMC-1rPR3-S176Acell lysates as antigen.
( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123 119
obtained when HMC-1rPR3-S176A cell lysates wereŽ .used as antigen rs0.472, p-0.001 . The strongest
correlation was observed between PR3–ANCA titersobtained by IIF on HMC-1rPR3 cells and the cap-ture ELISA absorbance readings obtained with
ŽHMC-1rPR3-S176A cell lysates rs0.576, p-.0.001 .
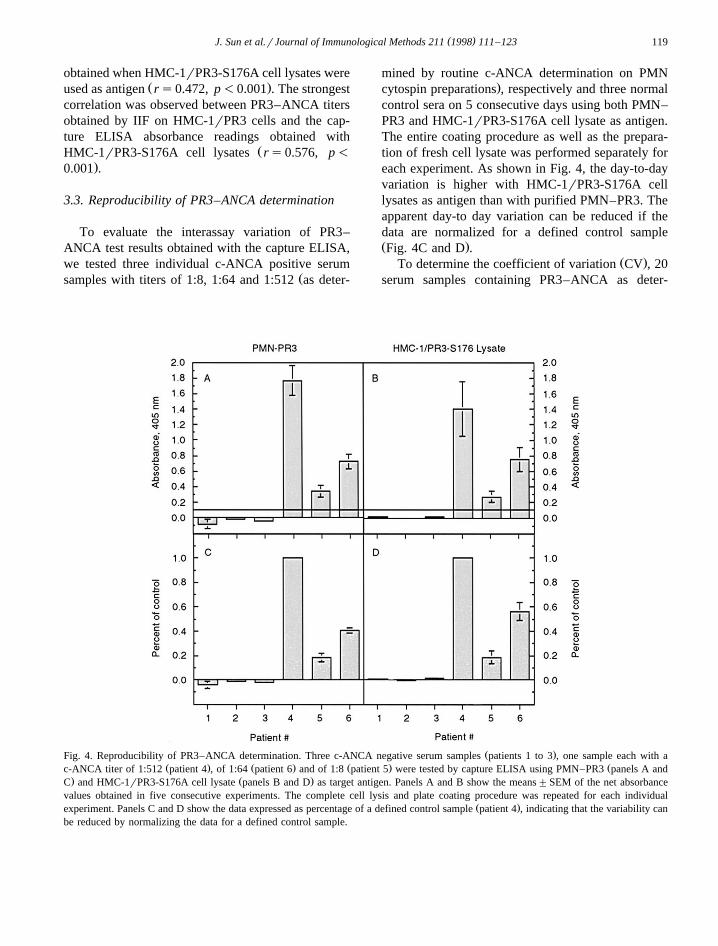
3.3. Reproducibility of PR3–ANCA determination
To evaluate the interassay variation of PR3–ANCA test results obtained with the capture ELISA,we tested three individual c-ANCA positive serum
Žsamples with titers of 1:8, 1:64 and 1:512 as deter-
mined by routine c-ANCA determination on PMN.cytospin preparations , respectively and three normal
control sera on 5 consecutive days using both PMN–PR3 and HMC-1rPR3-S176A cell lysate as antigen.The entire coating procedure as well as the prepara-tion of fresh cell lysate was performed separately foreach experiment. As shown in Fig. 4, the day-to-dayvariation is higher with HMC-1rPR3-S176A celllysates as antigen than with purified PMN–PR3. Theapparent day-to day variation can be reduced if thedata are normalized for a defined control sampleŽ .Fig. 4C and D .
Ž .To determine the coefficient of variation CV , 20serum samples containing PR3–ANCA as deter-
Ž .Fig. 4. Reproducibility of PR3–ANCA determination. Three c-ANCA negative serum samples patients 1 to 3 , one sample each with aŽ . Ž . Ž . Žc-ANCA titer of 1:512 patient 4 , of 1:64 patient 6 and of 1:8 patient 5 were tested by capture ELISA using PMN–PR3 panels A and
. Ž .C and HMC-1rPR3-S176A cell lysate panels B and D as target antigen. Panels A and B show the means"SEM of the net absorbancevalues obtained in five consecutive experiments. The complete cell lysis and plate coating procedure was repeated for each individual
Ž .experiment. Panels C and D show the data expressed as percentage of a defined control sample patient 4 , indicating that the variability canbe reduced by normalizing the data for a defined control sample.

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123120
mined by IIF using HMC-1rPR3-S176A cells werepooled. These sera had titers ranging from 1:8 to 1:512 as previously determined by standard IIF usingPMN cytospin preparations. This pooled sample wasassayed in 10 consecutive experiments including theentire coating and cell lysis procedure. For PMN–PR3 as target antigen the mean net absorbance value"SEM was 1.36"0.05, for HMC-1rPR3-S176Acell lysates it was 0.69"0.07. The interassay CVwas 11.99% when PMN–PR3 and 31.01% whenHMC-1rPR3-S176A cell lysate was used. Theintra-assay CV calculated based on experiments per-formed in 10 repeat wells within the same assay was5.61% for PMN–PR3 and 13.03% for HMC-1rPR3-S176A cell lysate as antigen.
3.4. Detection of PR3–ANCArPR3 immunecom-plexes
Sera from patients with WG may contain freePR3–ANCA as well as PR3–ANCArPR3 im-
Ž .munecomplexes Baslund et al., 1994 . The clinicalsignificance of the presence of such immunecom-plexes remains unclear. In capture ELISA systems,such immunecomplexes may be the source of signifi-cant background that, rarely, may obscure the pres-ence of free PR3–ANCA. Fig. 5 shows two PR3–
ŽANCA positive sera not from the series of 200.patients that generated background values in the
capture ELISA that were as high as the valuesgenerated in wells containing the antigen. Removalof free PR3 and PR3–ANCArPR3 immunecom-plexes from the samples by preadsorption of thesesera with MCPR3-2 resulted in substantial back-ground reduction. The presence of free PR3–ANCAin these samples was confirmed by incubating theMCPR3-2-preadsorbed sample with added exoge-nous purified PMN–PR3. The addition of this mix-ture to MCPR3-2 coated wells resulted in doublingof the signal, thereby proving the presence of freePR3–ANCA in these samples which would havebeen overlooked otherwise. While these data confirmthat the capture ELISA results can be confounded bythe presence of PR3–ANCArPR3 immunecom-plexes, they also show that, when combined withpreadsorption of the serum sample with MCPR3-2,this capture ELISA can be used for the detection ofsuch immunecomplexes.
Fig. 5. The presence of circulating PR3–ANCArPR3-complexescan mask the presence of free PR3–ANCA. Shown are two serumsamples that yielded background absorbance values on MCPR3-2
Ž .coated wells not containing antigen B that were as high as theŽ .absorbance obtained in antigen containing wells A . Subtraction
of background would have falsely resulted in a negative PR3–ANCA result. After solid phase preadsorption with MCPR3-2Žremoving PR3 and PR3–ANCArPR3 immunecomplexes from
.the serum samples the supernatant was retested in wells withoutŽ .antigen, resulting in reduction of background absorbance C .
When free PR3 was added to the supernatant and the mixture wasŽ .assayed in parallel in wells without antigen D , the absorbance
almost doubled, clearly confirming the presence of free PR3-ANCA in these serum samples.
3.5. QuantitatiÕe determination of PR3
Microtiter wells were coated with the moABMCPR3-2 as described in Section 2. To generate astandard curve for PR3, serial dilutions of purifiedenzymatically active PMN–PR3 were prepared and
Fig. 6. Application of the capture ELISA for quantitative PR3measurements. Shown is the dose response curve for concentra-tions of purified PMN–PR3 ranging from 1.5 to 200 ngrml. Eachdata point represents the mean"SEM of 15 consecutive experi-ments performed by three different operators. The dilutions ofPR3 were prepared freshly for each of the experiments.

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123 121
incubated in the wells for 1 h at RT. The assay wasperformed as described in Section 2. Fig. 6 showsthe resulting standard curve which indicates that thedetection limit is about 2 ngrml and that it is almostlinear over a 20-fold concentration range. The curveshown resulted from 15 experiments performed onconsecutive days by three different operators. Addi-tional experiments indicated that the absorbancereadings are not affected by the presence of
Ž .PR3ra1-PI complexes not shown .
4. Discussion
Solid phase assays for PR3–ANCA testing havebeen developed in order to allow antigen-specific,reader-independent, semiquantitative PR3–ANCAtesting. Various methods of antigen-preparation havebeen reported and the thorough standardization andclinical evaluation of several of these methods is still
Ž .in progress Hagen et al., 1996 . Previous studieshave suggested that the sensitivity of most PR3–ANCA ELISA assays based on the direct coating ofthe purified antigen to the plastic plate is inferior tothe standard IIF method of c-ANCA detection using
Žneutrophil cytospin preparations Specks et al., 1997;.Wieslander, 1991 . One possible explanation is that
immobilization may result in partial denaturation ofthe antigen with alteration of conformational epi-topes. This problem might be circumventable byimmobilizing the antigen via capturing moABs. In-deed, a recently described capture ELISA method,using a combination of three different moABs againstPR3 to capture the antigen, promises a sensitivityequivalent to the standard c-ANCA detection method
Ž .by IIF Baslund et al., 1995 . Detailed data about theanalytical sensitivity of another capture ELISA usedfor PR3–ANCA detection are currently not availableŽ .Merkel et al., 1997 .
The moABs MCPR3-1 and MCPR3-2 describedhere were raised using granule extracts of HMC-1rPR3 cells as immunogen and were shown to reactspecifically with rPR3 and native PMN–PR3. Ourepitope competition analysis would suggest that theyrecognize overlapping epitopes on PR3. They do notcompete for the epitopes recognized by the moABs
Ž .4A5 and 6A6 developed by Sommarin et al. 1995 ,but their moAB 4A3 seems to recognize a larger
epitope that partially overlaps with the binding sitesof MCPR3-1 and MCPR3-2. These findings are con-sistent with the data indicating that MCPR3-2 doesnot compete for a sizable number of PR3–ANCAbinding sites. This is in contrast to 4A5 and 6A6which inhibit the binding of 61% and 11% of PR3–
Ž .ANCA sera, respectively Sommarin et al., 1995 .The antibody 4A3 only partially inhibited the bind-ing of about 22% of PR3–ANCA sera, which wereall also inhibited by 4A5.
One of our goals was to identify an easily accessi-ble abundant source of antigen. The analytical speci-ficity of PR3–ANCA ELISA systems for PR3–ANCA depends on the purity of the immobilizedantigen. Described purification procedures for PR3are tedious, require large numbers of neutrophils asstarting material and expose personnel to the risk ofinfection. The principal of the capture ELISA cir-cumvents the antigen purity issue, provided that thecapturing antibodies used have no cross-reactivitywith other relevant antigens. Unfortunately, PR3 hasbeen shown to cleave immunoglobulins including
Ž .ANCA Dolman et al., 1995 . Consequently, theproteolytic activity of PR3 might interfere with thestability of the capturing antibody, particularly sinceMCPR3-2 does not inhibit the enzymatic activity ofPR3. This reasoning contributed to the design of theenzymatically inactive rPR3-S176A mutant ex-
Žpressed in HMC-1rPR3-S176A cells Specks et al.,.1996 . The direct comparison of HMC-1rPR3-
S176A cell lysates to purified enzymatically activePMN–PR3 showing equivalent analytical sensitivity
Ž .for PR3–ANCA indicates: 1 proteolytic degrada-tion of immobilized MCPR3-2 by active PR3 does
Ž .not represent a technical problem, and 2 HMC-1rPR3-S176A cell lysates represent a convenientsource for specific target antigen, obviating the te-dious purification of PR3.
The PR3–ANCA values obtained for a pooledcontrol serum with the two antigen preparations weremore variable with the HMC-1rPR3-S176A celllysate. This was expected as the coating procedurecontains more variables when cell lysates are used.Both the cell counting procedure as well as thepreparation of a homogeneous lysate solution areoperator-dependent procedures at this stage of assaydevelopment. As suggested by the lower intra-assayCV, the inter-assay CV is likely to be significantly

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123122
improved when large batches of plates are preparedand the assay is performed on plates from the samebatch. Our data also indicate that expression of thesample data as percentage of a standardized controlserum will further reduce the variability of the re-sults.
Some patients with PR3–ANCA also may havecirculating PR3–ANCArPR3 immunecomplexesŽ .Baslund et al., 1994 . Rarely, this may cause highbackgrounds in the capture ELISA to the extent ofmasking the presence of free PR3–ANCA. It wasnot a problem among 101 consecutive c-ANCA posi-tive serum samples and we had to search for suchsamples by screening a large serum bank. Neverthe-less, serum samples yielding high background read-ings should be further tested for the presence of suchimmunecomplexes by repeating the assay afterpreadsorption of the serum sample with MCPR3-2and after addition of free exogenous PR3 to thesample as described above. Future studies will haveto determine the clinical relevance of the relationshipof free PR3–ANCA to PR3–ANCArPR3 im-munecomplexes. This assay may represent a conve-nient tool for such studies.
Finally, this assay allows the quantitative determi-nation of PR3. Since the assay results are not af-fected by the presence of PR3–a1-PI complexes, theprimary reading does not allow a distinction betweenfree PR3 and PR3–a1-PI complexes. However, incombination with antibodies against a1-PI this assaycan be used for the detection of such complexes andpreadsorption of the sample with antibodies againsta1-PI allows the measurement of free PR3. Theavailability of sensitive assays that allow the specificdistinction between PR3 and elastase are of crucialimportance for the study of neutrophil serine pro-
Žtease mediated inflammatory conditions Vender,.1996 , particularly since there are significant differ-
ences in substrate and inhibitor spectrum betweenelastase and PR3 despite their structural homology
Žand functional similarities Padrines et al., 1994; Raoet al., 1993; Robache-Gallea et al., 1995; Spector et
.al., 1995 .
Acknowledgements
The authors want to thank Mr. Robert D. Litwiller,Mr. Randall S. Miller and Ms. Elaine M. Wiegert for
excellent technical assistance with the antigen purifi-cation, antibody production and immunofluorescenceassays, respectively; Ms. Amber M. Hummel formaintenance of the cell lines; Dr. Steven V. Bittorfand Ms. Hua Tang for performing some of theexperiments resulting in Fig. 2d and Fig. 6; Mr.Darrell Schroeder from the Department of Biostatis-tics for help with the statistical analysis of the data;and Ms. Kathryn L. Stanke for help with the prepara-tion of the manuscript. The authors are also indebtedto Dr. Joseph H. Butterfield for the kind gift of theHMC-1 parent cell line. This work was supported by
Ž .a grant-in-aid AHA96008260 from the AmericanHeart Association, a Research Grant from the Ameri-
Ž .can Lung Association of Minnesota both to US andfunds from the Mayo Foundation. It was performedduring the tenure of a Clinician-Scientist AwardŽ .AHA94004360 from the American Heart Associa-tion to US.
References
Baslund, B., Petersen, J., Permin, H., Wiik, A., Wieslander, J.,1994. Measurements of proteinase 3 and its complexes witha1-proteinase inhibitor and anti-neutrophil cytoplasm antibod-
Ž .ies ANCA in plasma. J. Immunol. Meth. 175, 215.Baslund, B., Segelmark, M., Wiik, A., Szpirt, W., Petersen, J.,
Wieslander, J., 1995. Screening for anti-neutrophil cytoplas-Ž .mic antibodies ANCA : is indirect immunofluorescence the
method of choice?. Clin. Exp. Immunol. 99, 486.Bini, P., Gabay, J.E., Teitel, A., Melchior, M., Zhou, J.-L., Elkon,
K.B., 1992. Antineutrophil cytoplasmic autoantibodies in We-gener’s granulomatosis recognize conformational epitopes onproteinase 3. J. Immunol. 149, 1409.
Borregaard, N., Heiple, J.M., Simons, E.R., Clark, R.A., 1983.Subcellular localization of the b-cytochrome component of thehuman neutrophil microbicidal oxidase: translocation duringactivation. J. Cell Biol. 97, 52.
Campanelli, D., Melchior, M., Fu, Y., Nakata, M., Shuman, H.,Nathan, C., Gabay, J.E., 1990. Cloning of cDNA for pro-teinase 3: a serine protease, antibiotic and autoantigen fromhuman neutrophils. J. Exp. Med. 172, 1709.
Cohen Tervaert, J.W., van der Woude, F.J., Fauci, A.S., Ambrus,J.L., Velosa, J., Keane, W.F., Meijer, S., van der Giessen, M.,The, T.H., van der Hem, G.K., Kallenberg, C.G.M., 1989.Association between active Wegener’s granulomatosis andanticytoplasmic antibodies. Arch. Intern. Med. 149, 2461.
Davies, D.J., Moran, J.E., Niall, J.F., Ryan, G.B., 1982. Segmen-tal necrotising glomerulonephritis with antineutrophil anti-body: possible arbovirus aetiology. Br. Med. J. 285, 606.

( )J. Sun et al.rJournal of Immunological Methods 211 1998 111–123 123
Dolman, K.M., Jager, A., Sonnenberg, A., von dem Borne, A.E.,Goldschmeding, R., 1995. Proteolysis of classic anti-neu-
Ž .trophil cytoplasmic autoantibodies c-ANCA by neutrophilproteinase 3. Clin. Exp. Immunol. 101, 8.
Falk, R.J., Jennette, J.C., 1988. Anti-neutrophil cytoplasmic au-toantibodies with specificity for myeloperoxidase in patientswith systemic vasculitis and idiopathic necrotizing and cres-centic glomerulonephritis. N. Engl. J. Med. 25, 1651.
Gupta, S.K., Niles, J.L., McCluskey, R.T., Arnaout, M.A., 1990.Ž .Identity of Wegener’s autoantigen P29 with proteinase 3 and
myeloblastin. Blood 76, 2162.Hagen, E.C., Andrassy, K., Csernok, E., Daha, M.R., Gaskin, G.,
Gross, W.L., Hansen, B., Heigl, Z., Hermans, J., Jayne, D.,Kallenberg, C.G.M., Lesavre, P., Lockwood, C.M., Lude-¨mann, J., Mascart-Lemone, F., Mirapeix, E., Pusey, C.D.,Rasmussen, N., Sinico, R.A., Tzioufas, A., Wieslander, J.,Wiik, A., van der Woude, F.J., 1996. Development and stan-dardization of solid phase assays for the detection of anti-neu-
Ž .trophil cytoplasmic antibodies ANCA . A report on the sec-ond phase of an international cooperative study on the stan-dardization of ANCA assays. J. Immunol. Meth. 196, 1.
Hall, J.B., McN., W.B., Wood, C.J., Ashton, V., R., A.W., 1984.Vasculitis and glomerulonephritis: a subgroup with an antineu-trophil cytoplasmic antibody. Aust. New Zealand J. Med. 14,277.
Jenne, D.E., Tschopp, J., Ludemann, J., Utecht, B., Gross, W.L.,¨1990. Wegener’s autoantigen decoded. Nature 346, 520.
Jenne, D.E., Frohlich, L., Hummel, A.M., Specks, U., 1997.¨Cloning and functional expression of the murine homologue ofproteinase 3: implications for the design of murine models ofvasculitis. FEBS Lett. 408, 187.
Kallenberg, C.G.M., Brouwer, E., Weening, J.J., Cohen Tervaert,J.W., 1994. Anti-neutrophil cytoplasmic antibodies: currentdiagnostic and pathophysiological potential. Kidney Int. 46, 1.
Kao, R.C., Wehner, N.G., Skubitz, K.M., Gray, B.H., Hoidal,J.R., 1988. A distinct human polymorphonuclear leukocyteproteinase that produces emphysema in hamsters. J. Clin.Invest. 82, 1963.
Kwak, J.-W., Yoon, C.-S., 1996. A convenient method for epitopecompetition analysis of two monoclonal antibodies for theirantigen binding. J. Immunol. Meth. 191, 49.
Ludemann, J., Csernok, E., Ulmer, M., Lemke, H., Utecht, B.,¨Rautmann, A., Gross, W.L., 1990. Anti-neutrophil cytoplasmantibodies in Wegener’s granulomatosis: immunodiagnosticvalue, monoclonal antibodies and characterization of the targetantigen. Neth. J. Med. 36, 157.
Merkel, P.A., Polisson, R.P., Chang, Y., Skates, S.J., Niles, J.L.,1997. Prevalence of antineutrophil cytoplasmic antibodies in a
large inception cohort of patients with connective tissue dis-ease. Ann. Intern. Med. 126, 866.
Nolle, B., Specks, U., Ludemann, J., Rohrbach, M.S., DeRemee,¨ ¨R.A., Gross, W.L., 1989. Anticytoplasmic autoantibodies: theirimmunodiagnostic value. Ann. Int. Med. 111, 28.
Padrines, M., Wolf, M., Walz, A., Baggiolini, M., 1994. Inter-leukin-8 processing by neutrophil elastase, cathepsin G andproteinase-3. FEBS Lett. 352, 231.
Rao, N.V., Marshall, B.C., Gray, B.H., Hoidal, J.R., 1993. Inter-action of secretory leukocyte protease inhibitor with proteinase3. Am. J. Respir. Cell Mol. Biol. 8, 612.
Robache-Gallea, S., Morand, V., Bruneau, J.M., Schoot, B., Tagat,E., Realo, E., Chouaib, S., Roman-Roman, W., 1995. In vitro´processing of human tumor necrosis factor-a. J. Biol. Chem.270, 23688.
Sommarin, Y., Rasmussen, N., Wieslander, J., 1995. Characteriza-tion of monoclonal antibodies to proteinase 3 and applicationin the study of epitopes for classical anti-neutrophil cytoplasmantibodies. Exp. Nephrol. 3, 249.
Specks, U., Fass, D.N., Fautsch, M.P., Hummel, A.M., Viss,M.A., 1996. Recombinant human proteinase 3, the Wegener’sautoantigen, expressed in HMC-1 cells is enzymatically activeand recognized by c-ANCA. FEBS Lett. 390, 265.
Specks, U., Wiegert, E.M., Homburger, H.A., 1997. Human mastŽ .cells expressing recombinant proteinase 3 PR3 as substrate
for clinical testing for anti-neutrophil cytoplasmic antibodiesŽ .ANCA . Clin. Exp. Immunol. 109, 286.
Spector, N.L., Hardy, L., Ryan, C., Miller, W.H., Humes, J.L.,Nadler, L.M., Luedke, E., 1995. 28-kDa mammalian heatshock protein, a novel substrate of a growth regulatory pro-tease involved in differentiation of human leukemia cells. J.Biol. Chem. 270, 1003.
van der Woude, F.J., Rasmussen, N., Lobatto, S., Wiik, A.,Perman, H., van Es, L.A., van der Giessen, M., van der Hem,G.K., The, T.H., 1985. Autoantibodies against neutrophils andmonocytes: tool for diagnosis and marker of disease activity inWegener’s granulomatosis. Lancet 1, 425.
Vender, R.L., 1996. Therapeutic potential of neutrophil-elastaseinhibition in pulmonary disease. J. Invest. Med. 44, 531.
Wieslander, J., 1991. How are antineutrophil cytoplasmic antibod-ies detected?. Am. J. Kidney Dis. 18, 154.
Wiik, A., van der Woude, F.J., 1990. The new ACPArANCAnomenclature. Neth. J. Med. 35, 107.
Zhao, M.-H., Lockwood, C.M., 1996. A comprehensive method topurify three major ANCA antigens: proteinase 3, myeloperoxi-dase and bactericidalrpermeability-increasing protein fromhuman neutrophil granule acid extract. J. Immunol. Meth. 197,121.