calpain is a mediator of preservation-reperfusion injury in rat liver transplantation
TRANSCRIPT

Calpain is a Mediator of Preservation-Reperfusion Injury in Rat Liver TransplantationAuthor(s): Vivek Kohli, Wenshi Gao, Carlos A. Camargo and Pierre Alain ClavienSource: Proceedings of the National Academy of Sciences of the United States of America,Vol. 94, No. 17 (Aug. 19, 1997), pp. 9354-9359Published by: National Academy of SciencesStable URL: http://www.jstor.org/stable/42608 .
Accessed: 05/05/2014 11:17
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
National Academy of Sciences is collaborating with JSTOR to digitize, preserve and extend access toProceedings of the National Academy of Sciences of the United States of America.
http://www.jstor.org
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions

Proc. Natl. Acad. Sci. USA Vol. 94, pp. 9354-9359, August 1997 Medical Sciences
Calpain is a mediator of preserv liver transplantation
(calcium/organ preservation/warm ischemia/cold ischemia/pi
VIVEK KOHLI, WENSHI GAO, CARLOS A. CAMARGO, JR.,
Hepatobiliary and Liver Transplantation Laboratory, Department of Surgery, I
Communicated by Gertrude B. Elion, Glaxo Wellcome, Inc., Research 7
ABSTRACT Proteases as well as alterations in intracel- lular calcium have important roles in hepatic preservation- reperfusion injury, and increased calpain activity recently has been demonstrated in liver allografts. Experiments were designed to evaluate (i) hepatic cytosolic calpain activity during different periods of cold ischemia (CI), rewarming, or reperfusion, and (ii) effects of inhibition of calpain on liver graft function using the isolated perfused rat liver and arte- rialized orthotopic liver transplantation models. Calpain ac- tivity was assayed using the fluorogenic substrate Suc-Leu- Leu-Val-Tyr-7-amino-4-methyl coumarin (AMC) and ex- pressed as mean ? SD pmol AMC released/min per mg of cytosolic protein. Calpain activity rose significantly after 24 hr of CI in University of Wisconsin solution and further increased with longer preservation. Activity also increased within 30 min of rewarming, peaking at 120 min. Increased durations of CI preceding rewarming resulted in significantly higher activity (P < 0.01). Calpain activity increased rapidly upon reperfusion and was significantly enhanced by previous CI (P < 0.01). Calpain inhibition with Cbz-Val-Phe methyl ester significantly decreased aspartate aminotransferase re- leased in the isolated perfused rat liver perfusate (P < 0.05). Duration of survival after orthotopic liver transplantation using livers cold-preserved for 40 hr was also significantly increased (P < 0.05) with calpain inhibitor. In conclusion, calpain proteases are activated during each phase of trans- plantation and are likely to play an important role in the mechanisms of preservation-reperfusion injury.
The mechanisms and mediators involved in preservation- reperfusion (P/Rp) injury of liver allografts remain largely unknown (1). Proteases (2-5) as well as alteration in intracel- lular calcium concentration (2, 6, 7) have been found to play an important role in hepatic P/Rp injury. Recent evidence suggests that calpain proteases are involved in the pathogenesis of anoxia/reoxygenation or ischemia/reperfusion injury in several animal models, including the brain (8), myocardium (9), and renal tubular cells (10). Calpain activation also was noted in liver tissue after reperfusion of transplanted human livers, and interestingly, the degree of calpain activity corre- lated with postoperative serum levels of transaminases (11).
Injuries to parenchymal and nonparenchymal cells of the liver occur during each phase of liver transplantation, which includes the period of cold ischemia (CI), anaerobic rewarm- ing, and reperfusion. Cold preservation typically causes sinu- soidal lining cell injury with alterations in cytoskeleton and detachment of endothelial cells from the hepatocyte plate (12, 13). Rewarming ischemia occurs during the implantation phase of orthotopic liver transplantation (OLT), and mainly
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. ?1734 solely to indicate this fact. ? 1997 by The National Academy of Sciences 0027-8424/97/949354-6$2.00/0 PNAS is available online at http://www.pnas.org.
t
ation-reperfusion injury in rat
'otease)
\ND PIERRE ALAIN CLAVIEN*
4uke University Medical Center, Durham, NC 27710
'riangle Park, NC, June 18, 1997 (received for review January 8, 1997)
causes anoxic hepatocyte injury with anaerobic glycolysis, ATP depletion, and intracellular acidosis (14, 15). Reperfu- sion injury is dependent on the degree of injury occurring during the previous phases (1). It involves complex and unclear intracellular mechanisms and mediators as well as kupffer cell activation along with platelet and polymorphonuclear cell adhesion (1).
Alteration in intracellular calcium concentrations with dra- matic increase of cytosolic calcium due to release from cellular organelles is a common feature of anoxia/reoxygenation injury (16), including hepatocytes (17), endothelial cells (18-20), and kupffer cells (20). As a result, calcium-dependent intracellular pathways and proteases are activated, culminating in cell injury and eventually death (21). For instance, in hepatocytes in- creased cytosolic calcium activates nonlysosomal proteases, leading to cell membrane bleb formation (17, 22), a typical feature of hepatocyte P/Rp injury (13). Furthermore, calcium channel blockers (2, 6, 23) and inhibition of proteases (2, 3, 24) have beneficial effects on hepatic P/Rp injury. Nisoldipine, a calcium channel blocker, added to a mixture of antiproteases significantly improved survival in a rat OLT model (2). Sim- ilarly, improved liver functions were seen in the isolated perfused rat liver (IPRL) model after addition of antiproteases in the preservation solution (3). However, the specific pro- teases involved in P/Rp of liver allograft remain unknown.
Calpain proteases are nonlysosomal calcium-dependent in- tracellular proteases (25, 26). They are ubiquitous in verte- brates and have been isolated from various cells, including hepatocytes (27) and endothelial cells (19). Evidence has been given that calpain proteases play a central role in cytoskeletal and cell membrane metabolism (25) and can be activated by anoxic or ischemia/reperfusion injury (9, 10, 28, 29). Struc- tural proteins and other microtubular proteins undergo partial degradation by calpain protease (25, 26). This could lead to structural instability of the cell manifested as cellular swelling, bleb formation, and cell death (17, 30), all features of P/Rp injury of liver allograft.
In this study we have evaluated calpain activity during each phase of P/Rp injury. Calpain activity was also evaluated after different combinations of cold, rewarming, and reperfusion injury with particular emphasis to relevant combinations as- sociated with graft survival. The involvement of calpain in the pathogenesis of P/Rp injury was assessed by using a specific cell permeable inhibitor in the IPRL and the arterialized rat OLT models.
MATERIALS AND METHODS
Experimental Design. Experiments were designed to deter- mine calpain protease activity in cytosol from livers preserved
Abbreviations: AMC, 7-amino 4-methyl coumarin; AST, aspartate aminotransferase; CI, cold ischemia; IPRL, isolated perfused rat liver; OLT, orthotopic liver transplantation; P/Rp, preservation- reperfusion; UW, University of Wisconsin solution. *To whom reprint requests should be addressed at: Department of Surgery, Duke University Medical Center, Box No. 3247, Durham NC 27710. e-mail: [email protected].
}354
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions

Medical Sciences: Kohli et al.
for various durations of CI, rewarming, or reperfusion as well as a combination of different durations of CI followed by rewarming or reperfusion. Preliminary studies were per- formed to determine calpain inhibition using specific inhibi- tors. Effects of calpain inhibition on liver injury and animal survival after prolonged preservation then were evaluated using the IPRL and arterialized rat OLT models.
Materials. Tris-HCI, EDTA, CaC12, Cbz-Val-Phe-methyl ester, dimethyl sulfoxide, 7 amino-4 methyl coumarin (AMC), Suc-Leu-Leu-Val-Tyr-AMC, taurocholic acid, and BSA were obtained from Sigma. Z-Leu-Leu-Tyr-CHN2 was a gracious gift from John Anagli, Institute of Biochemistry, Basel, Swit- zerland. Krebs-Henseleit buffer was freshly prepared.
Preparation of Cytosol Extract. Cytosol was obtained by a modification of the method described by Hargrove (31). Tris-buffered saline (100 mM Tris-HCI, 145 mM NaCl, pH 7.3) was used. EDTA (10 mM) was added to chelate Ca2+. Liver biopsies were performed at varying preservation or reperfu- sion periods, suspended in the buffer and homogenized (Tis- sumizer, Cincinnati, OH). The homogenized tissue was cen- trifuged at 12,000 x g for 30 min. The supernatant was again spun at 150,000 g for 90 min (rotor 70.1Ti, Beckman). The extracted cytosol was immediately frozen in liquid nitrogen and stored at -70?C.
Protein Assay. Cytosolic protein was assayed in microtiter plates with 50 /1 of diluted sample plus 150 /.l of Bio-Rad- Bradford reagent and optical density read at 562 nm using EL Biokinetics Reader (Bio-Tek, Winooski, VT). BSA was used as the standard for protein assay.
Calpain Assay. Suc-Leu-Leu-Val-Tyr-7-amino-4-methyl coumarin was used as the calpain substrate. Forty microliters of cytosol (1 mg protein/ml) was added to 160 /1 of 50 /M Suc-Leu-Leu-Val-Tyr-AMC (dissolved in dimethyl sulfoxide and buffer 100 mM Tris HCI, 145 mM NaCl at pH 7.3). Three sets of assays were conducted for each sample: (i) reconsti- tuting Ca2+ at 10 mM thus enabling calpain I and II activity; (ii) without reconstituting Ca2+; and (iii) in presence of Ca2+ and 400 nM of z-Leu-Leu-Tyr-CHN2, an inhibitor of calpain proteases and cathepsin L (32). AMC released was measured by fluorometry (Denley Wellflor II, Cambridge Technology, Watertown, MA) using 360-nm excitation and 430-nm emis- sion filters. To simulate the preservation conditions, all assays were conducted at the temperature at which the liver was preserved, i.e., assays involving cold preserved livers were read at 1?C and those involving rewarming or reperfusion were read at room temperature (25?C). Comparisons in calpain activity were made only among calpain activities read at the same temperature. Standard curves were generated for each exper- iment using AMC of known concentrations. Calpain specificity of the assay was determined by: (i) conducting the assay in presence of z-Leu-Leu-Tyr-CHN2, a specific inhibitor of cal- pain proteases, which also inhibits cathepsin L; (ii) defining the calpain activity as Ca2+-dependent activity, thus excluding cathepsin activity, which is strictly calcium independent; and (iii) conducting the assay at pH 7.3 at which the cathepsins are known to be inactivated (33). Calcium-dependent calpain activity was expressed as pmol AMC released/mg cytosolic protein at 10 mM Ca2+. The calcium-independent activity in the different experimental groups was comparable.
Animal Experiments. Wistar rats (Harlan Breeders, India- napolis, IN) weighing between 175 and 225 gm were used for all preservation experiments, including the IPRL. Female Lewis rats (Harlan) were used for the OLT experiments. Rats were allowed free access to food and water before procure- ment of the liver. Phenobarbitone (30 mg/kg) was injected intraperitoneally for anesthesia. Experiments were conducted in compliance with the Duke University Institutional Animal Care and Use Committee.
CI and Rewarming. Livers were procured as previously described (34). Briefly, the portal vein was cannulated and
Proc. Natl. Acad. Sci. USA 94 (1997) 9355
flushed in situ with 12 cc of cold University of Wisconsin solution (UW) allowed to run with gravity at 12 cm above the level of the animal. Livers were either cold-preserved for up to 60 hr in UW at I?C or allowed to rewarm at room temperature (25?C) for up to 120 min. Sequential biopsies were taken from each liver.
IPRL. An IPRL model was used as described by Miller et al. (35) with later modifications (36). The system was a closed circuit using a small amount of perfusate (110 ml) consisting of fresh rat blood diluted with Krebs-Henseleit buffer (pH 7.4; hematocrit of 11%) to which cefazolin (17 mg/l) was added. The perfusion was conducted at 37?C in a temperature- controlled box, and the liver was kept moist in Ringer's solution. Sodium taurocholate (30 gmol/hr) and heparin (500 units/hr) were continuously infused. The portal vein was perfused by gravity at a constant pressure of 12 cm. Venous return from the liver was collected through cannulas in the supra and infra hepatic vena cava and recirculated after oxygenation. A mixture of 95% 02 and 5% CO2 through a silastic tube lung with adjustable gas flows was used. The pO2 was kept above 150 mmHg, pCO2 at 40 mmHg and pH between 7.35 to 7.45. Each liver was perfused for 3 hr.
Liver Biopsy of the Perfused Rat Liver. Sequential biopsies were performed during reperfusion as previously described (37). Biopsies were taken from the inferior and superior parts of the caudate lobe, inferior part of the right lobe, and the superior part of the right lateral lobe. Hemostasis was ensured by ligating the pedicles of these lobes with preplaced 5.0 silk sutures.
Orthotopic Arterialized Rat Liver Transplantation. Arte- rialized OLT were performed between female Lewis rats weighing 150-175 gm as previously described (34, 38). The donor livers were flushed in situ with 5 ml of cold UW (1?C). Total time required for explantation was less than 5 min. Before implantation, livers were rinsed with cold Ringer's solution. A hand-sewn upper caval anastamosis was followed by anastamosis of infrahepatic vena cava and portal vein using the cuff technique. After reestablishing perfusion in the liver through the portal vein, arterial and biliary anastamosis were performed using silastic stents. The portal vein clamping time was less than 20 min in all experiments. To eliminate operator- related discrepancy, all transplant procedures were performed by a single investigator (W.G.) experienced in rat OLT who was blinded to the treatment and control groups. In our hands, the upper limit of CI time associated with no graft loss is 24 hr, and the minimal CI time associated with complete nonsurvival is 40 hr.
Inhibition of Calpain Activity. Preliminary experiments were performed to determine the inhibitory effects of Cbz- Val-Phe methyl ester, a specific and cell permeable calpain protease inhibitor (10, 33). A dose of 60 mg/kg dissolved in 0.2 ml of dimethyl sulfoxide given i.p. 2 hr before retrieval of the liver was found to dramatically reduce calpain activity during cold preservation and to a lesser degree after reperfusion. For instance, after 40 hr of cold preservation calpain activity was 41.3 + 3.4 (mean ?+ SD) pmol AMC/min per mg of cytosolic protein in rats pretreated with the solvent only and 3.2 ? 2.1 in those pretreated with the inhibitor (n = 3 in each group; P <
0.001). Two hours after reperfusion in an OLT model, calpain activity was 67.5 ? 4.2 pmol AMC/min per mg of cytosolic protein in the control group vs 48.5 + 4.7 pmol AMC/min per mg of cytosolic rats pretreated with the inhibitor (n = 3 each group; P = 0.01). No further decrease in calpain protease activity was seen using either repeated or higher doses of the inhibitor (data not shown).
Statistical Methods. The results are expressed as mean + SD of AMC released/min per mg of cytosolic protein at 10 mMol Ca2+. Paired and unpaired Student's t test for small size and ANOVA were used for analysis using the Statistical
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions
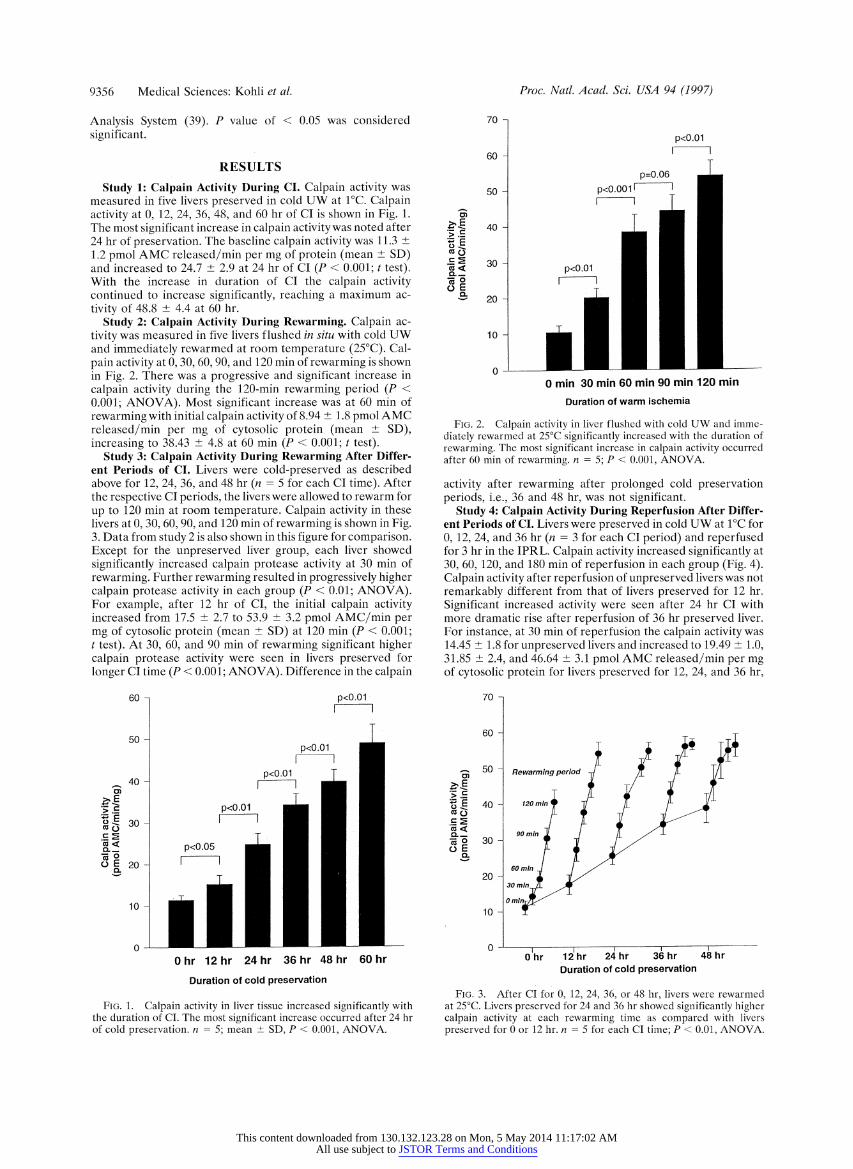
9356 Medical Sciences: Kohli et al.
Analysis System (39). P value of < 0.05 was considered significant.
RESULTS
Study 1: Calpain Activity During CI. Calpain activity was measured in five livers preserved in cold UW at 1?C. Calpain activity at 0, 12, 24, 36, 48, and 60 hr of CI is shown in Fig. 1. The most significant increase in calpain activity was noted after 24 hr of preservation. The baseline calpain activity was 11.3 ? 1.2,pmol AMC released/min per mg of protein (mean ? SD) and increased to 24.7 ? 2.9 at 24 hr of CI (P < 0.001; t test). With the increase in duration of CI the calpain activity continued to increase significantly, reaching a maximum ac- tivity of 48.8 ? 4.4 at 60 hr.
Study 2: Calpain Activity During Rewarming. Calpain ac- tivity was measured in five livers flushed in situ with cold UW and immediately rewarmed at room temperature (25?C). Cal- pain activity at 0, 30, 60, 90, and 120 min of rewarming is shown in Fig. 2. There was a progressive and significant increase in calpain activity during the 120-min rewarming period (P < 0.001; ANOVA). Most significant increase was at 60 min of rewarming with initial calpain activity of 8.94 ? 1.8 pmol AMC released/min per mg of cytosolic protein (mean ? SD), increasing to 38.43 ? 4.8 at 60 min (P < 0.001; t test).
Study 3: Calpain Activity During Rewarming After Differ- ent Periods of CI. Livers were cold-preserved as described above for 12, 24, 36, and 48 hr (n = 5 for each CI time). After the respective CI periods, the livers were allowed to rewarm for up to 120 min at room temperature. Calpain activity in these livers at 0, 30, 60, 90, and 120 min of rewarming is shown in Fig. 3. Data from study 2 is also shown in this figure for comparison. Except for the unpreserved liver group, each liver showed significantly increased calpain protease activity at 30 min of rewarming. Further rewarming resulted in progressively higher calpain protease activity in each group (P < 0.01; ANOVA). For example, after 12 hr of CI, the initial calpain activity increased from 17.5 ? 2.7 to 53.9 ? 3.2 pmol AMC/min per mg of cytosolic protein (mean ? SD) at 120 min (P < 0.001; t test). At 30, 60, and 90 min of rewarming significant higher calpain protease activity were seen in livers preserved for longer CI time (P < 0.001; ANOVA). Difference in the calpain
60 - p<O.01
50 - p<O.01
p<O.01
0 L
o 20 -
10-
Ohr 12hr 24hr 36hr 48hr 60 hr
Duration of cold preservation
FIG. 1. Calpain activity in liver tissue increased significantly with the duration of CI. The most significant increase occurred after 24 hr of cold preservation. n = 5; mean + SD, P < 0.001, ANOVA.
Proc. Natl. Acad. Sci. USA 94 (1997)
70 -
p<0.01
60- I I
p=0.06 50 P < 0.001, ANOVA.
40 - E_ .=-
> 6
ar o 30 -mn
0.O
oE 20 -2
10 -
0
0 min 30 min 60 min 90 min 120 min
Duration of warm pschemia
FIG. 2. Calpain activity in liver flushed with cold UW and imme- diately rewarmed at 25C4 and 36 hr showed significantly increased with the duration of
rewarming. The most significant increase in calpain activity occurred after 60 mfor of rewarming. n = 5 ; P < 0.001 , ANOVA.
activity after rewarming after prolonged cold preservation periods, i.e., 36 and 48 hr, was not significant.
Study 4: Calpain Activity During Reperfusion After Differ- ent Periods of CI. Livers were preserved in cold UW at i?C for 0, 12, 24, and 36 hr (n = 3 for each CI period) and reperfused for 3 hr in the IPRL. Calpain activity increased significantly at 30, 60, 120, and 180 min of reperfusion in each group (Fig. 4). Calpain activity after reperfusion of unpreserved livers was not remarkably different from that of livers preserved for 12 hr. Significant increased activity were seen after 24 hr CI with more dramatic rise after reperfusion of 36 hr preserved liver. For instance, at 30 min of reperfusion the calpain activity was 14.45 ? 1.8 for unpreserved livers and increased to 19.49 _+ 1.0, 31.85 2.4, and 46.64 ?+ 3.1 pmol AMC released/min per mg of cytosolic protein for livers preserved for 12, 24, and 36 hr,
70-
60-
50- Rewarming period
1 40 120 mi
60 min
Duration of cold preservation
preserved for 0 or 12 hr. n -S for each CI time; P < 0.01, ANOVA.
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions
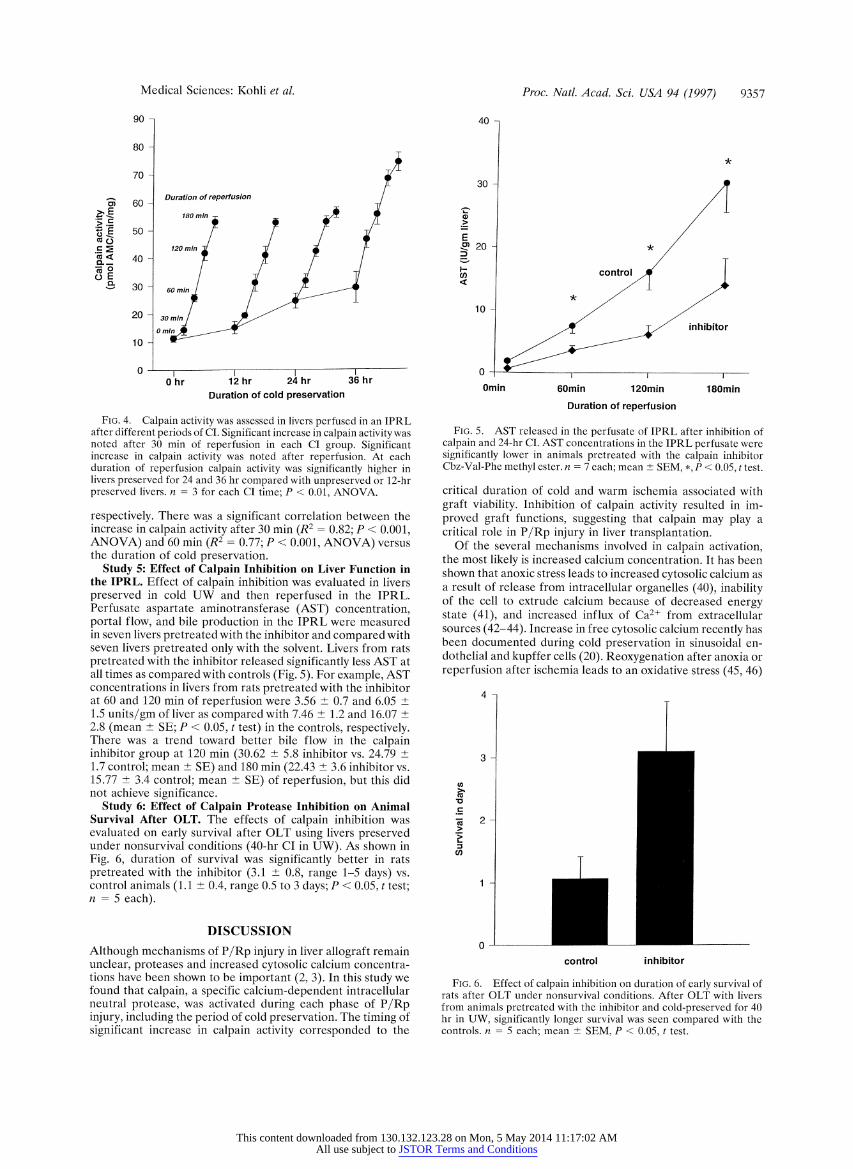
Medical Sciences: Kohli et al.
90-
80-
70-
mi 60 Duration of reperfusion
i ~. E 180 minr18 fm
E 50 *
(80- 40 - 120 mi
a< 40
' 30- - 60 min
20 30 mn
1 0 m
0 i I 0 hr 12 hr 24 hr 36 hr
Duration of cold preservation
FIG. 4. Calpain activity was assessed in livers perfused in an IPRL after different periods of CI. Significant increase in calpain activity was noted after 30 min of reperfusion in each CI group. Significant increase in calpain activity was noted after reperfusion. At each duration of reperfusion calpain activity was significantly higher in livers preserved for 24 and 36 hr compared with unpreserved or 12-hr preserved livers. n = 3 for each CI time; P < 0.01, ANOVA.
respectively. There was a significant correlation between the increase in calpain activity after 30 min (R2 = 0.82; P < 0.001, ANOVA) and 60 min (R2 = 0.77; P < 0.001, ANOVA) versus the duration of cold preservation.
Study 5: Effect of Calpain Inhibition on Liver Function in the IPRL. Effect of calpain inhibition was evaluated in livers preserved in cold UW and then reperfused in the IPRL. Perfusate aspartate aminotransferase (AST) concentration, portal flow, and bile production in the IPRL were measured in seven livers pretreated with the inhibitor and compared with seven livers pretreated only with the solvent. Livers from rats pretreated with the inhibitor released significantly less AST at all times as compared with controls (Fig. 5). For example, AST concentrations in livers from rats pretreated with the inhibitor at 60 and 120 min of reperfusion were 3.56 + 0.7 and 6.05 ? 1.5 units/gm of liver as compared with 7.46 ? 1.2 and 16.07 ? 2.8 (mean ? SE; P < 0.05, t test) in the controls, respectively. There was a trend toward better bile flow in the calpain inhibitor group at 120 min (30.62 ? 5.8 inhibitor vs. 24.79 + 1.7 control; mean + SE) and 180 min (22.43 ? 3.6 inhibitor vs. 15.77 ? 3.4 control; mean ? SE) of reperfusion, but this did not achieve significance.
Study 6: Effect of Calpain Protease Inhibition on Animal Survival After OLT. The effects of calpain inhibition was evaluated on early survival after OLT using livers preserved under nonsurvival conditions (40-hr CI in UW). As shown in Fig. 6, duration of survival was significantly better in rats pretreated with the inhibitor (3.1 ? 0.8, range 1-5 days) vs. control animals (1.1 ? 0.4, range 0.5 to 3 days; P < 0.05, t test; n = 5 each).
DISCUSSION
Although mechanisms of P/Rp injury in liver allograft remain unclear, proteases and increased cytosolic calcium concentra- tions have been shown to be important (2, 3). In this study we found that calpain, a specific calcium-dependent intracellular neutral protease, was activated during each phase of P/Rp injury, including the period of cold preservation. The timing of significant increase in calpain activity corresponded to the
Proc. Natl. Acad. Sci. USA 94 (1997) 9357
40-
30-
E m 20- -k /
n control
10- /
~ 1~ T/ inhibitor
0- I Omin 60min 120min 180min
Duration of reperfusion
FIG. 5. AST released in the perfusate of IPRL after inhibition of calpain and 24-hr CI. AST concentrations in the IPRL perfusate were significantly lower in animals pretreated with the calpain inhibitor Cbz-Val-Phe methyl ester. n = 7 each; mean + SEM, *, P < 0.05, t test.
critical duration of cold and warm ischemia associated with graft viability. Inhibition of calpain activity resulted in im- proved graft functions, suggesting that calpain may play a critical role in P/Rp injury in liver transplantation.
Of the several mechanisms involved in calpain activation, the most likely is increased calcium concentration. It has been shown that anoxic stress leads to increased cytosolic calcium as a result of release from intracellular organelles (40), inability of the cell to extrude calcium because of decreased energy state (41), and increased influx of Ca2+ from extracellular sources (42-44). Increase in free cytosolic calcium recently has been documented during cold preservation in sinusoidal en- dothelial and kupffer cells (20). Reoxygenation after anoxia or reperfusion after ischemia leads to an oxidative stress (45, 46)
4
3 -
2 2
control inhibitor
FIG. 6. Effect of calpain inhibition on duration of early survival of rats after OLT under nonsurvival conditions. After OLT with livers from animals pretreated with the inhibitor and cold-preserved for 40 hr in UW, significantly longer survival was seen compared with the controls. n = 5 each; mean ?+ SEM, P < 0.05, t test.
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions

9358 Medical Sciences: Kohli et al.
associated with increased free cytosolic calcium concentration (42), and as recently shown, oxidative stress in hepatocytes leads to calpain activation (30). The initial rapid rise in calpain activation after reperfusion in our model support oxidative- related mechanisms.
Whereas increased cytosolic Ca2+ remains the most likely cause for calpain activation, other mechanisms might be involved. Increased synthesis of calpain, increased calpain activity by inhibition of calpastatin, and increased phospho- lipase activity may contribute to calpain activation (25). Al- though increased calpain synthesis is unlikely during cold and rewarming ischemia, it may occur within a few hours of reperfusion and may explain our failure to completely inhibit calpain activity after 2 hr of reperfusion. Calpain activation also may occur with minimal or no increase in calcium concentration as a result of increase in phospholipases and phospholipid degradation process (47, 48). Phospholipases have been shown to activate calpains during anoxic injury to hepatocytes (49) and may similarly be involved in injury during P/Rp.
Calpain has been shown to be an important mediator of cellular injury. Several observations in our study support the role of calpain proteases as a mediator of P/Rp injury in liver transplantation. First, the degree of increase in calpain activity corresponded to the critical durations of cold preservation and rewarming times associated with graft viability. The most significant increase in calpain activity was seen after 24 hr or more of cold preservation, the period after which the liver allograft viability in the rat begins to decline (38). After rewarming the most significant rise in calpain activity was noted at 60 min. This is similar to the observations that increased rewarming time beyond 60 min result in increased primary graft dysfunction in humans (49). Second, we found additive effects of prolonged CI, rewarming, and reperfusion on calpain activity. Longer durations of CI before rewarming or reperfusion resulted in more rapid and higher increase in calpain activity. This is consistent with previous clinical and experimental data showing cumulative effects of CI with rewarming and reperfusion injury (50, 51). Finally, inhibition of calpain activation in the liver resulted in significantly reduced injury as evaluated by the release of AST in the IPRL and improved early survival after OLT.
Calpain proteases are likely to cause cellular injury by several mechanisms. They initiate proteolysis of vital cell membrane and cytoskeletal structural proteins (25, 26). Ini- tiation of proteolytic cleavage of these proteins may lead to progressive cell injury and death. Hepatocyte bleb formation, typically seen during cold preservation (13), is a common manifestation of hepatocyte injury (17, 52) and may result from cell membrane and cytoskeletal damage as a result of increased calpain activity. Calpain proteases also behave as signaling proteases, for example, by activating protein kinase C (26, 53) and may result in activation of several pathways, leading to cell injury. Recent evidence suggests activation of apoptosis during P/Rp injury (54, 55). Calpains are one of the effector arms of apoptotic cell death (56, 57). Increased calpain activity may represent activation of such processes within the liver, including the typical sinusoidal lining cell injury related to cold preservation (13). The increase in calpain activity during P/Rp as well as the beneficial effects of calpain inhibition in this study were seen with cold preservation using UW, the accepted preserva tive solution for liver transplanta- tion. Thus, it is likely that inhibition of calpain activity during clinical liver transplantation may decrease P/Rp injury.
In conclusion, calpain protease activation occurs during each phase of P/Rp during liver transplantation and inhibition of calpain activation improves the graft functions. These changes were observed using UW, the standard preservative solution for abdominal organ transplantation. Thus, it is likely that inhibition of calpain may have important clinical appli-
Proc. Natl. Acad. Sci. USA 94 (1997)
cations. Future studies should aim at developing calpain protease inhibitors with improved solubility and specificity. Further experiments with such inhibitors will help determine optimum calpain inhibition and their possible therapeutic implications in the prevention of P/Rp injury during liver transplantation.
1. Clavien, P. A., Harvey, P. R. C. & Strasberg, S. M. (1992) Transplantation 53, 957-978.
2. Takei, Y., Marzi, I., Kauffman, F. C., Currin, R. T., Lemasters, J. J. & Thurman, R. G. (1990) Transplantation 50, 14-20.
3. Clavien, P. A., Sanabria, J. S., Upadhaya, A., Cywes, R., Harvey, P. R. C. & Strasberg, S. M. (1993) Transplantation 56, 44-53.
4. Nichols, J. C., Bronk, S. F., Mellgren, R. L. & Gores, G. J. (1994) Gastroenterology 106, 168-176.
5. Ferguson, D. M., Gores, G. J., Bronk, S. F. & Krom, R. A. F. (1993) Transplantation 55, 627-633.
6. Nauta, R. J., Tsimoyiannis, E., Uribe, M., Walsh, D. B., Miller, D. & Butterfield, A. (1991) Ann. Surg. 213, 137-142.
7. Thomas, C. E. & Reed, D. J. (1989) Hepatology 10, 375-384. 8. Lee, K. S., Frank, S., Vanderklish, P., Arai, A. & Lynch, G. (1991)
Proc. Natl. Acad. Sci. USA 88, 7233-7237. 9. Tolnai, S. & Korecky, B. (1986) Can. J. Cardiol. 2, 42-47.
10. Edelstein, C. L., Wieder, E. D., Yaqoob, M. M., Gengaro, P. E., Burke, T. J., Nemenoff, R. A. &. Schrier, R. W. (1995) Proc. Natl. Acad. Sci. USA 92, 7662-7666.
11. Aguilar, H. I., Steers, J. L., Weisner, R. H., Krom, R. A. F. & Gores, G. J. (1997) Transplantation 63, 612-614.
12. Caldwell-Kenkel, J. C., Currin, R. T., Tanaka, Y., Thurman, R. G. & Lemasters, J. J. (1989) Hepatology 10, 292-295.
13. McKeown, C. M. B., Edwards, V., Phillips, M. J., Harvey, P. R. C., Petrunka, C. N. & Strasberg, S. M. (1988) Transplantation 46, 178-191.
14. Harvey, P. R. C., Iu, S., McKeown, C. M. B, Ilson, R. G. & Strasberg, S. M. (1988) Transplantation 45, 1016-1119.
15. Kamiike, W., Wanatabe, F., Hashimoto, T., Tagawa, K., Ikeda, Y., Nakao, K. & Kawashima, Y. (1982) J. Biochem. 91, 1349- 1356.
16. Snowdowne, K. W., Freudenrich, C. C. & Borle, A. B. (1985) J. Biochem. 260, 11619-11626.
17. Nieminen, A. L., Gores, G. J., Wray, B. E., Tanaka, Y., Herman, B. & Lemasters, J. J. (1988) Cell Calcium 9, 237-246.
18. Nicotera, P., McConkey, D. J., Dypbukt, J. M., Jones, D. P. & Orrenius, S. (1989) Drug Metab. Rev. 20, 193-201.
19. Geeraerts, M. D., Ronveanoux-Dupal, M. F., Lemasters, J. J. & Herman, B. (1991) Am. J. Physiol. 261, C889-C896.
20. Cabrillac, J. C., Haddad, P., Piche, D., Musallam, D. L. & Huet, P. M. Hepatology (1996) 24, 306 (abstr.).
21. Cheung, T., Bonventre, J., Malis, C. & Leaf, A. (1986) N. Engl. J. Med. 314, 1670-1676.
22. Nicotera, P., Hartzell, P., Davis, G. & Orrenius, S. (1986) FEBS Lett. 209, 139-144.
23. Cheng, S., Ragsdale, J. R., Sasaki, A. W., Lee, R. G., Denecey, C. W. & Pinson, W. (1991) J. Surg. Res. 50, 560-564.
24. Lie, T. S., Seger, R., Hong, G. S., Preissinger, H. & Ogawa, K. (1989) Transplantation 48, 396-399.
25. Croall, D. E. & Demartino, G. N. (1991) Physiol. Rev. 71, 813-847.
26. Saido, T. C., Sorimachi, H. & Suzuki, K. (1994) FASEB J. 8, 814-822.
27. Pontremoli, S., Melloni, E., Salamino, F., Sparatore, B., Michette, M. & Horecker, B. L. (1984) Proc. Natl. Acad. Sci. USA 81, 53-56.
28. Bartus, R. T., Hayword, N. J., Elliot, P. J., Sawyer, S. D., Baker, K. L., Dean, R. L., Akiyama, A., Straub, J. A., Haberson, S. L. & Li, Z. (1994) Stroke 25, 2265-2270.
29. Hong, S. C., Goto, Y. L., Lanzino, G., Soleau, S., Kessell, N. F. & Lee, K. S. (1994) Stroke 25, 663-669.
30. Miyoshi, H., Umeshita, K., Sakon, S., Imajohomi, M. S., Fujitani, K., Gotoh, M., Oiki, E., Kambayashi, J. & Monden, M. (1996) Gastroenterology 110, 1897-1904.
31. Hargrove, J. L., Trotter, J. F., Ashline, H. C. & Krishnamurti, P. V. (1989) Metabolism 38, 666-672.
32. Crawford, C., Mason, R. W., Wilkstorm, P. & Shaw, E. (1988) Biochem. J. 253, 751-758.
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions

Medical Sciences: Kohli et al.
33. Barrett, A. J. & Krishke, H. (1981) Methods Enzymol. 80, 535-561.
34. Kamada, N. & Calne, R. Y. (1979) Transplantation 28, 47-50. 35. Miller, L. L., Gly, C. G., Watson, M. C. & Bale, W. F. (1951) J.
Exp. Med. 94, 431-435. 36. Clavien, P. A., Morgan, G., Sanabria, J. R., Harvey, P. R. C. &
Strasberg, S. M. (1991) Transplantation 52, 412-417. 37. Clavien, P. A., Sanabria, J. R., Cywes, R., Harvey, P. R. C. &
Strasberg, S. M. (1992) Liver 12, 69-72. 38. Gao, W., Lemasters, J. J. & Thurman, R. (1993) Transplantation
56, 19-24. 39. Statistical Analysis System Institute. (1990) SAS User's Guide:
Statistics, Version 6 (Statistical Analysis System Institute, Cary, NC).
40. Hatanaka, N., Kamiike, W., Shimizu, S., Miyata, M., Inoue, T., Yoshida, Y., Takawa, K. & Matsuda, H. (1995) J. Surg. Res. 58, 485-490.
41. Arnould, T., Michiels, C., Alexandre, I. & Remacle, J. (1992) J. Cell Physiol. 152, 215-221.
42. Greene, E. L. & Paller, M. S. (1994)Am. J. Physiol. 266, F13-F20. 43. Salvaterra, C. G. & Goldman, W. F. (1993) Am. J. Physiol. 264,
L323-L328. 44. Gyenes, M. & De Groot, H. (1993) Am. J. Physiol. 264, G353-
G350. 45. Jaeschke, H., Smith, C. V. & Mitchell, J. R. (1988) J. Clin. Invest.
81, 1240-1246.
Proc. Natl. Acad. Sci. USA 94 (1997) 9359
46. Sussman, M. S. & Bulkley, G. B. (1990) Methods Enzymol. 186, 711-766.
47. Sakaida, I., Thomas, A. P. & Farber, J. L. (1992) Am. J. Physiol. 263, C684-C690.
48. Arora, A. A., Groen, P. D., Emori, Y. & Gores, G. J. (1996)Am. J. Physiol. 33, G238-G245.
49. Cisneros, C., Guillen, F., Gomez, R., Gutierrez, J., Vorwald, P., Montero, A. & Moreno, E. (1991) Transplant Proc. 23, 1976.
50. Cywes, R., Greig, P. D., Sanabria, J. R., Clavien, P. A., Levey, G. A., Harvey, P. R. C. & Stasberg, S. M. (1992) Ann. Surg. 216, 235-247.
51. Ploeg, R. J., D'Alessandro, A. M., Knetchle, S. J., Stegall, M. D., Pirsch, J. D., Hoffman, R. M., Sasaki, T., Sollinger, H. W., Belzer, F. 0. & Kalayoglu, M. (1993) Transplantation 55, 807-813.
52. Gores, G. J., Herman, B. & Lemasters, J. J. (1990) Hepatology 11, 690-698.
53. Kishimoto, A., Mikawa, K., Hashimoto, K., Yasuda, I., Tanaka, S., Tominaga, M., Koruda, T. & Nishizuka, Y. (1989) J. Biol. Chem. 264, 4088-4092.
54. Taneja, C., Klien, K. M., Swank, G. M., Deitch, E. A. & Koneru, B. (1996) Hepatology 24, 425 (abstr.).
55. Tox, U., Thies, J. C., Teklote, J., Otto, G., Stremmel, W. & Theilmann, L. (1996) Hepatology 24, 425 (abstr.).
56. Squier, M. K., Miller, A. C., Malkinson, A. M. & Cohen, J. J. (1994) J. Cell Physiol. 159, 229-237.
57. Sarin, A., Clerici, M., Blatt, S. P., Hendrix, C. W., Shearer, G. M. & Henkart P. A. (1994) J. Immunol. 153, 862-872.
This content downloaded from 130.132.123.28 on Mon, 5 May 2014 11:17:02 AMAll use subject to JSTOR Terms and Conditions