branching out: origins of the sea urchin larval skeleton in development and evolution
TRANSCRIPT

REVIEW
Branching Out: Origins of the Sea Urchin Larval Skeleton inDevelopment and Evolution
Daniel C. McIntyre, Deirdre C. Lyons, Megan Martik, and David R. McClay*
Department of Biology, Duke University, Durham, North Carolina
Received 24 December 2013; Revised 14 February 2014; Accepted 14 February 2014
Summary: It is a challenge to understand how the infor-mation encoded in DNA is used to build a three-dimensional structure. To explore how this works theassembly of a relatively simple skeleton has beenexamined at multiple control levels. The skeleton of thesea urchin embryo consists of a number of calcite rodsproduced by 64 skeletogenic cells. The ectoderm sup-plies spatial cues for patterning, essentially telling theskeletogenic cells where to position themselves andproviding the factors for skeletal growth. Here, wedescribe the information known about how this works.First the ectoderm must be patterned so that the sig-naling cues are released from precise positions. Theskeletogenic cells respond by initiating skeletogenesisimmediately beneath two regions (one on the right andthe other on the left side). Growth of the skeletal rodsrequires additional signaling from defined ectodermallocations, and the skeletogenic cells respond to pro-duce a membrane-bound template in which the calcitecrystal grows. Important in this process are threesignals, fibroblast growth factor, vascular endothelialgrowth factor, and Wnt5. Each is necessary for explicittasks in skeleton production. genesis 52:173–185. VC 2014 Wiley Periodicals, Inc.
Key words: VEGF; FGF; Wnt5; calcite; skeleton patterning
The sea urchin larva produces a beautiful, intricatelypatterned, calcareous endoskeleton. Enclosed withinthe transparent epithelium of the embryo, the skeletongives the larva its characteristic three-dimensionalshape. Because each half of the skeleton is formed as asingle crystal of calcite, the skeleton is refractive to lightand prominently visible under a light microscope (Fig.1). The form of the skeleton generally resembles anartist’s easel and for this the larva was named pluteusby Johannes Muller in 1846 (from the Greek for easel,
see H€orstadius, 1973). The skeleton of the early pluteuslarva has four arms extending out ventrally from thelarval body (Fig. 1f). The detailed anatomy of this struc-ture differs widely among the euechinoids (Wray,1992), reflecting changes in the patterning inputs overtime. In all species, the skeleton is produced by a smallnumber of mesenchyme cells. These cells fuse, andshare a common cytoplasm. The membranes of this syn-cytium surround an extracellular pocket in which thecalcite skeleton grows. The final skeletal pattern isstrongly dependent on signaling inputs from the sur-rounding ectodermal epithelium (Armstrong et al.,1993; Duboc et al., 2004; Guss and Ettensohn, 1997;Hardin and Armstrong, 1997; Hardin et al., 1992; McIn-tyre et al., 2013; Rottinger et al., 2008). The nature ofthese patterning signals, how they are localized withinthe ectoderm, and how the mesenchyme cells respondto them are questions considered here. This system is afascinating and relatively simple example of how tissuescollaborate to produce pattern during morphogenesis.All animals pattern the three-dimensional shape of theembryo. Here, we take advantage of molecular informa-tion that has accumulated on specification of ectodermand mesoderm in the sea urchin to understand how theskeletal patterning template emerges.
Current Address for Daniel C. McIntyre: Skirball Institute of Biomolec-
ular Medicine, 540 First Avenue 4th floor, Lab 17, New York, NY
10016.
* Correspondence to: David R. McClay, Department of Biology, DukeUniversity, 124 Science Dr., Box 90338, Durham, NC 27708.
E-mail: [email protected]
Contract grant sponsor: NIH, Contract grant numbers: RO1-HD-14483
and PO1-HD-037105Published online 18 February 2014 in
Wiley Online Library (wileyonlinelibrary.com).
DOI: 10.1002/dvg.22756
VC 2014 Wiley Periodicals, Inc. genesis 52:173–185 (2014)
The echinoderm phylum consists of five extantclasses: Crinoidea (feather stars and sea lillies), Asteroi-dea (sea stars), Holothuroidea (sea cucumbers), Ophiur-oidea (brittle stars and basket stars), and Echinoidea(sea urchins and sand dollars; McClay, 2011). Most echi-noderms employ a biphasic life cycle that includes aplanktonic larval stage followed by a benthic adultstage. The sea urchin larval skeleton is thought to serveseveral functions. Primarily, it protects the digestive
organs and helps orient the arms upward in the watercolumn for effective swimming and feeding (Penning-ton and Strathmann, 1990). A ciliary band a few cellswide runs along the larval arms and functions in loco-motion and orientation, and it produces currents direct-ing food to the mouth (Strathmann, 2000, 2007;Strathmann and Grunbaum, 2006). Echinoid larval skel-etons are diverse between orders and even betweenspecies of the same order (Kinjo et al., 2008; Wray,
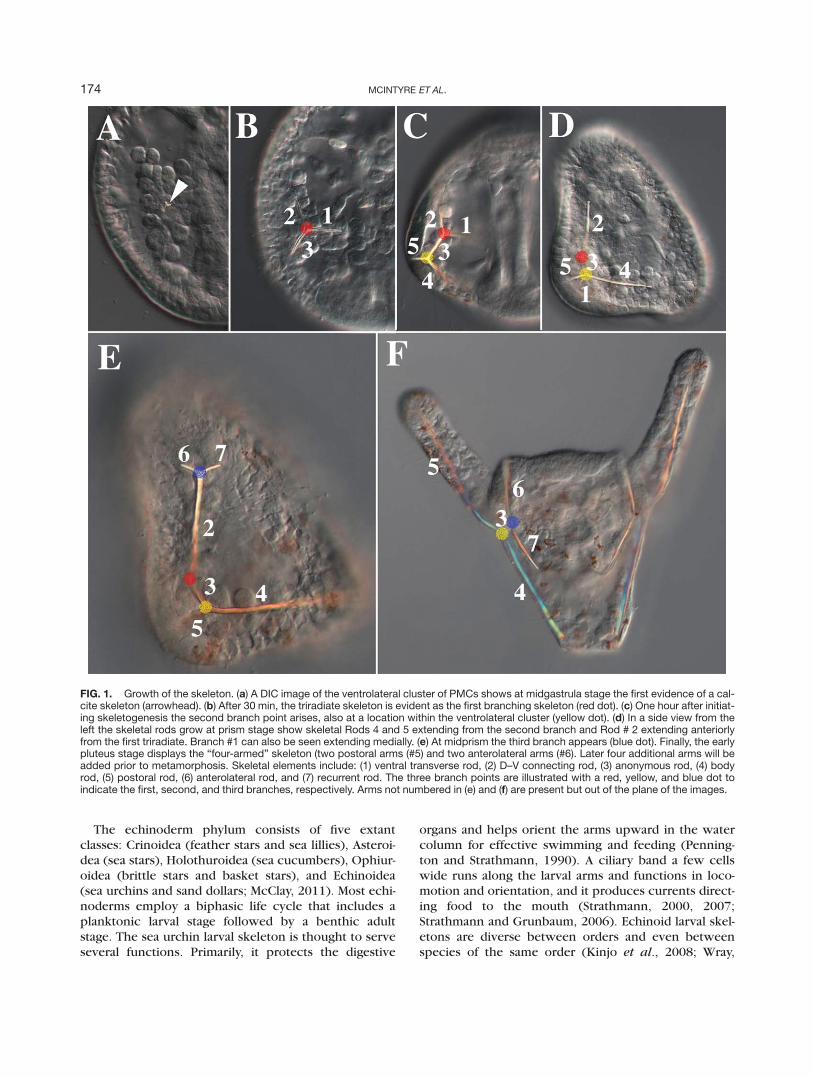
FIG. 1. Growth of the skeleton. (a) A DIC image of the ventrolateral cluster of PMCs shows at midgastrula stage the first evidence of a cal-cite skeleton (arrowhead). (b) After 30 min, the triradiate skeleton is evident as the first branching skeleton (red dot). (c) One hour after initiat-ing skeletogenesis the second branch point arises, also at a location within the ventrolateral cluster (yellow dot). (d) In a side view from theleft the skeletal rods grow at prism stage show skeletal Rods 4 and 5 extending from the second branch and Rod # 2 extending anteriorlyfrom the first triradiate. Branch #1 can also be seen extending medially. (e) At midprism the third branch appears (blue dot). Finally, the earlypluteus stage displays the “four-armed” skeleton (two postoral arms (#5) and two anterolateral arms (#6). Later four additional arms will beadded prior to metamorphosis. Skeletal elements include: (1) ventral transverse rod, (2) D–V connecting rod, (3) anonymous rod, (4) bodyrod, (5) postoral rod, (6) anterolateral rod, and (7) recurrent rod. The three branch points are illustrated with a red, yellow, and blue dot toindicate the first, second, and third branches, respectively. Arms not numbered in (e) and (f) are present but out of the plane of the images.
174 MCINTYRE ET AL.

1992). The number, length, shape and decoration (e.g.,spines) of the skeletal rods supporting the arms canvary widely. Likewise, the elements and basketlikeshape of the body skeleton are different betweenspecies.
Figure 1 shows a sequence of skeletal growth in Lyte-
chinus variegatus. For orientation, we will refer to thedorsal-ventral sides of the embryo with the mouth inthe center of the ventral side. We will also use the termsanterior–posterior (A–P) to define the orthogonal axiswith the anus or blastopore being the most posteriorstructure. We use this terminology throughout basedon recent conventions in the literature (Range et al.,2013; Wei et al., 2012), and because the nomenclatureof the skeletal elements, established early in the lastcentury, is largely based on those terms. On the rightand left side of the embryo the skeletogenic cells, alsocalled primary mesenchyme cells (PMCs) enter the blas-tocoel, migrate, and form a ring around the future arch-enteron at the posterior end of the embryo. Cells thenmigrate within the ring so that more than half the 64PMCs accumulate as the right and left ventrolateral clus-ters (Peterson and McClay, 2003). As will be seen subse-quently, the migratory cues that localize PMCs to theventrolateral clusters are provided through ectodermalsignaling. In response to local signals each ventrolateralcluster of PMCs produces a triradiate skeletal primor-dium (Fig. 1a,b). Shortly after the first branch (red dotin Fig. 1), the skeleton branches again (yellow dot).Later a third branch occurs in the anterior ectodermjust inside the overlying dorsal–ventral margin (DVM;blue dot).
The basic relationship underlying skeletal pattern-ing—that the mesenchymal cells produce skeletonunder the direction of ectodermal signaling—wasinferred more than 100 years ago. Both Herbst (1893)and Driesch (1896) (in H€orstadius, 1973) suggested thatthe ectoderm contributed inputs to patterning of theskeleton. They based their idea on the observation thatthe apex of the skeleton was always at the mid-dorsalectoderm. However, for about 100 years there was littleexperimental evidence to support this hypothesis ofnonautonomous input. H€orstadius (1973) credits Boveri(1901a, 1901b) as the first to report that the micro-meres become the skeletogenic cells. H€orstadius (1935)later confirmed this lineage relationship by vitally stain-ing micromeres and transplanting them to unstainedmicromereless embryos. In this manner, embryologistsin the first half of the 20th century made significant pro-gress in understanding the source of mesenchyme cellsin the early embryo. With this knowledge of cell fates inhand, other researchers turned to studying the behav-iors of differentiating mesenchymal cells. Production ofthe calcite crystal rods was studied in detail by Okazaki(1960, 1975). A article by Okazaki and Inoue (1976)was especially important in showing that while the
skeleton produced in vivo was birefringent, it neverthe-less was smooth [perhaps suggesting a partial amor-phous calcium carbonate (ACC) structure]. However, ifone isolated the skeleton and grew calcite crystals on it,the crystals grew in a single orientation, thus showingthat despite its smooth appearance the skeleton was asingle crystal. Later it was shown that the calcite crystalgrows by addition of ACC granule secretion from theskeletogenic cells followed by rapid conversion of thatACC into the single calcite crystal (Beniash et al.,1999). Gustafson and Wolpert (1961) described thebehavior of skeletogenic cells as they migrated insidethe blastocoel in preparation for skeletogenesis, andOkazaki (1975) showed that many of those same behav-iors occurred when PMCs were placed in vitro. TheOkazaki work and earlier work by von Ubisch (1933,1939) and Baltzer et al. (1959) supported the hypothe-sis that autonomous patterning mechanisms were pro-grammed into the skeletogenic cells. As a consequencethey showed that details of the skeletal elements suchas whether or not the skeleton rods were fenestrated,smooth, or contained sharp branches, was a function ofthe autonomous skeletogenic program. All of theseearly studies, and many others, set the stage for molecu-lar analyses of specification, migration of PMCs, pattern-ing of the ectoderm, signaling from the ectoderm topattern the skeleton shape, and production of anatomi-cally correct skeletal elements. That complex processprovides a model for how multiple tissues in an embryocontribute to emergence of pattern (reviewed by Etten-sohn, 2009, Lyons et al., 2012).
In the last two decades much work has been dedi-cated to building a detailed gene regulatory network forsea urchin embryogenesis, and the skeletogenic lineageis the best understood of all cell types (McClay, 2011;Oliveri et al., 2008). In parallel, the biogenesis of theskeleton itself has been studied in detail from structural,proteomic and genomic perspectives (Killian and Wilt,2008; Livingston et al., 2006; Wilt et al., 2003). One ofthe most significant advances came in 1992, when itwas finally demonstrated experimentally that the ecto-derm is a source of nonautonomous patterning informa-tion for the mesenchyme cells (Hardin et al., 1992).Hardin et al. showed that if ectodermal patterning wasperturbed there was a dramatic effect on skeletal pat-terning, even when the skeletogenic cells were unper-turbed. Shortly thereafter, a series of chimeraexperiments again demonstrated that ectodermalinputs were crucial for patterning (Armstrong et al.,1993; Armstrong and McClay, 1994; Hardin and Arm-strong, 1997). By transplanting mesenchymal cellsbetween species with different skeletal patterns, it wasshown that the overall shape of the skeleton dependson inputs from the host ectoderm. The skeletal rodsgrew then with a combination of nonautonomousinputs from ectoderm plus the autonomous patterning
LARVAL SKELETON PATTERNING 175

based on the genotype of the skeletogenic cells. Thefact that skeletogenic cells of one species correctly pat-terned a larval skeleton even when surrounded by ecto-derm of another species also suggested that many ofthe patterning cues were remarkably conserved.
The molecular nature of the signals that pattern themesenchyme cells has begun to be revealed over thelast several years. Recent work has shown that twoorthogonal bands of ectoderm are uniquely specified atthe molecular level to produce growth factors and pat-terning cues to the mesenchyme cells. In this review,we highlight the recent advances in the understandingof ectodermal patterning, explore how the various sig-nals direct skeleton formation, and discuss how this sys-tem has changed during evolution. Many questionsremain unresolved. In particular, while we have a grossunderstanding of the ways mesenchyme cells respondto signaling, many of the molecular mechanisms under-lying these processes are not known. How do the skele-togenic cells move to their correct locations? How dothey organize into a syncytium? How do they interpretthe signals and secrete the skeleton crystal in the cor-rect shape? And how does the protein matrix functionas it is incorporated into the crystal? To address thosequestions it is first necessary to understand the informa-tion already known about skeletal patterning. That isthe purpose of this article.
ECTODERMAL PATTERNING FORESHADOWSTHE EMBRYONIC SKELETON
Ectoderm patterning begins with early subdivisions ofcell specification along the A–P and dorso-ventral (D–V)axes. This process results in two bands of ectoderm. TheDVM of the ectoderm separates the dorsal and ventral
ectoderm (Fig. 2a, pink band). The border ectoderm(BE) encircles the embryo immediately adjacent to theendoderm. (Fig. 2a, blue band; McIntyre et al., 2013).The DVM and ventral half of the BE eventually becomesthe ciliary band that surrounds the ventral ectoderm.Once specified, these territories will participate in mor-phogenesis in several ways: (1) The DVM and the BE pro-vide the ectodermal patterning inputs for skeletogenesis;(2) the ciliary band will later form along the DVM andthe ventral portion of the BE. (3) the ciliary band willalso later contain many of the neurons of the larva(Angerer et al., 2011; Burke et al., 2006; Nakajima et al.,2004). The early distribution of skeletogenic mesen-chyme cells parallels the BE and then the portion thatgrows anterior parallels the DVM band. The right andleft BE–DVM intersections coincide with the ventrolat-eral clusters of PMCs (Fig. 2a). We now know that cellsin the BE and DVM produce growth factors that directmigration of the mesenchymal cells. Vascular endothelialgrowth factor (VEGF) and fibroblast growth factor (FGF)are initially produced throughout the BE (at the time thePMCs initially form into a ring immediately beneath theBE), and then VEGF is upregulated at the BE–DVM inter-section, coincident with the clustering of the PMCs justbeneath. Shortly thereafter skeletogenesis begins inthose ventrolateral clusters (Adomako-Ankomah andEttensohn., 2013; Duloquin et al., 2007; McIntyre et al.,2013; Rottinger et al., 2008). Thus, to understand howskeletogeneic patterning works it is necessary to under-stand how the BE, DVM, and BE–DVM intersection areprecisely located in the ectoderm.
The BE is established by short-range signaling fromthe endoderm.
Recent work demonstrated that the BE is uniquelyspecified via signaling from the endoderm (McIntyre
FIG. 2. Relationship between dorsal–ventral margin (DVM), border ectoderm (BE), and skeleton. (a) Diagram of the mesenchyme blastulastage from the left side showing the DVM and BE. The BE–DVM intersection is diagrammed and will later produce VEGF to initiate skeletalgrowth of the PMC ventrolateral cluster just inside the ectoderm. At that location the triradiate skeletal element initiates. The anonymousrod branches shortly thereafter, also beneath the region of the BE–DVM intersection. The third branch of the skeleton later occurs along theDVM in the oral hood. (b) A diagram of the skeleton with the three branch points shown on each side. On each side, five of the seven rodsgrow in the blastocoel immediately inside either the DVM or BE. The two rods in the anterior/oral hood produce arms that initiate at theDVM branch point (blue dot) and extend ventral and dorsal.
176 MCINTYRE ET AL.

et al., 2013). This narrow band of ectodermal cells,about 4–5 cells wide, forms immediately adjacent to theendoderm prior to gastrulation (Fig. 2a). A number oftranscription factors and signaling molecules mark theBE and BE–DVM intersection; these include Lim1, Nk1,Pax2/5/8, IrxA, Msx, Otp, Trim1, and FoxJ1 (Fig. 3;Czerny et al., 1997; Di Bernardo et al., 1999; McIntyreet al., 2013; Su et al., 2009; Saudemont et al., 2010).These markers are expressed dynamically such that bythe beginning of gastrulation each is localized to a sub-region of the BE. Even though the pattern of the larvalskeleton is species-specific, the transcription factorsexpressed in the BE are similar across several species ofurchins including L. variegatus, Strongylocentrotus
purpuratus, and Paracentrotus lividus.The BE is induced by a short-range Wnt5 signal from
neighboring endodermal cells (McIntyre et al., 2013).The evidence supporting this signal includes knock-down and misexpression of Wnt5, as well as perturba-tions to endoderm specification upstream of Wnt5expression. The initial observation was that many per-turbations to endoderm specification also disrupt for-mation of the skeleton (for example, foxa, bra, Hox11/13: Gross and McClay, 2001; Oliveri et al., 2006; Peterand Davidson, 2010). These data suggested that endo-derm played some role inducing formation of the skele-ton. Since VEGF and FGF were identified as ectodermaldrivers of skeleton, if the endoderm functioned in skele-togenesis, it had to operate indirectly through the ecto-
derm. Indeed, expression of VEGF, FGF, and other BEmarkers were lost in embryos where Hox11/13bexpression was blocked by MASO (McIntyre et al.,2013).
Wnt5 was identified as the endodermal signalbecause it is first expressed downstream of Hox11/13bin the endoderm, is necessary for BE marker expres-sion, and it activates BE markers when ectopicallyexpressed. Embryos injected with a Wnt5 MASO fail toactivate BE markers and produce no skeleton. Whenectopic Wnt5 messenger RNA (mRNA) was injected, BEmarkers were activated without restriction along the A–P axis (see subsequently), showing that Wnt5 is capableof activating BE fates in all ectodermal cells. This resultwas confirmed by transplanting single blastomeres mis-expressing Wnt5 mRNA; the presence of the Wnt5 pro-tein in an ectopic cell set up a developmentalsequence: these cells attracted PMCs, and the PMCsproduced ectopic skeleton. Wnt5 later is expressed inthe dorsal BE after gastrulation has begun. This laterexpression is dependent on the earlier endodermalWnt5 activity, and is not seen in embryos injected withthe Wnt5 MASO (McIntyre et al., 2013).
Several important conclusions emerged from theseexperiments. First, it was clear that BE specificationwas necessary for skeleton formation. In Wnt5 MASOembryos, no skeleton formed and the growth factorVEGF was not expressed at the BE–DVM intersection.Second, although Wnt5 has been shown to be requiredfor convergent extension movements during gastrula-tion in Xenopus and Zebrafish (Kilian et al., 2003;Yamanaka et al., 2002), sea urchin embryos lackingWnt5 function form a gut normally, indicating thatWnt5 probably functions primarily as a signal to theectoderm and not as a regulator of gut morphogenesis.Finally, endodermal Wnt5 acts as a short-range signal.All cells are able to respond to ectopic Wnt5 signal, butin vivo, BE fates are only activated in a band 4–5 cellswide immediately adjacent to the endoderm border.This is consistent with Wnt5 activity in cultured MEFcells, which appear not to release Wnt5 into the culturemedia (Ho et al., 2012). The use of a short-range signalto specify ectodermal fate is consistent with the use oftiered, short-range signals to specify endomesoderm inthe sea urchin embryo (Davidson, 1989) and suggests itmay be the major mechanism used to assign fates alongthe A–P axis.
The above conclusion is at odds with classicalembryological thinking, which hypothesized thatsweeping, long-range gradients of factors within theembryo were responsible for determining fates alongthe A–P axis. As cleavage proceeded it was thoughtthat a cell’s position within the gradient resulted indifferential inheritance of cytoplasm, different com-plements of these factors and, ultimately, differentfates. It was in this intellectual tradition that
FIG. 3. The BE and DVM are special domains of gene expres-sion. Three signals are produced in the BE–DVM intersection. Anumber of transcription factors are expressed in one or more of theseveral domains.
LARVAL SKELETON PATTERNING 177

H€orstadius (1939, 1973), interpreting his extensiveresults on sea urchin blastomere recombinations, pos-ited that dual gradients of secreted factors emanatingfrom the animal and vegetal poles of the embryoprovided the information necessary to specify cell lin-eage. Later it was argued that an equally plausibleexplanation could be a series of local inductive inter-actions between tiered blastomeres to explain theexperimental results obtained by H€orstadius andothers (Davidson, 1989).
In the more than 20 years since Davidson first pro-posed it, most of the accumulated molecular data indi-cate that the short-range tier-to-tier signaling hypothesisis better supported than the long-range gradienthypothesis of earlier thinking. Maternal Wnt signalingautonomously activates specification of the large micro-meres, fated to produce the embryonic skeleton (Emily-Fenouil et al., 1998; Logan et al., 1999; Wikramanayakeet al., 1998). Micromeres signal through Delta-Notchand an unknown signal to their macromere neighborsto regulate endomesoderm specification (Oliveri et al.,2003; Sherwood and McClay, 1997, 1999; Sweet et al.,2002). Shortly thereafter the macromeres equatoriallycleave into the Veg2 and Veg1 layers, the Veg2 layerthen signals to the Veg1 layer above it (Peter and David-son, 2011). The Wnt5 evidence above extends thatshort-range tier to tier signaling relationship into theectoderm since the Veg1 layer induces the BE via short-range Wnt5 (McIntyre et al., 2013), and further signal-ing to the anterior end of the embryo (animal pole) hasbeen suggested by additional Wnt signaling (Rangeet al., 2013). Thus most cells along the A–P axis likelyare specified with the expression and necessary contri-bution of a series of short-range signals.
D–V AXIS FORMATION AND SUBDIVISION OFTHE BE BY TRANSFORMING GROWTH FACTOR bSIGNALING
The D–V (aboral-oral) axis in the sea urchin is activatedby asymmetric transforming growth factor b (TGFb)signaling in response to what is proposed to be an ear-lier mitochondrial asymmetry (Bradham and McClay,2006; Coffman and Davidson, 2001; Duboc et al.,2004). The two principle signals in this process areNodal and Bmp2/4 (Duboc et al., 2004). Both mole-cules are produced on the ventral side of the embryo.Nodal activates exclusively ventral fates, whereasBMP2/4 diffuses to the opposite side of the embryo andactivates dorsal fates. Several inhibitors of TGFb signal-ing limit the extent of Nodal and BMP signaling, ensur-ing that cells in the D–V margin receive neither signaland therefore have a unique gene regulatory networkstate. In most ectodermal cells, Nodal and BMP2/4serve as activators of the gene regulatory network.Without them, D–V fates are lost and the ectoderm
defaults to a ciliated epithelium. Surprisingly, the BE isnot activated by TGFb; instead gene expressiondomains within that band are limited by Nodal andBMP. Dorsal markers in the BE are repressed on the ven-tral side by Nodal signaling; ventral markers of the BEare repressed in the dorsal side by BMP signaling (McIn-tyre et al., 2013).
Perturbations to the Nodal-BMP system in the seaurchin lead to specific defects in skeletal patterning(Hardin et al., 1992; Duboc et al., 2004). Those defectsgenerally include loss of anterior structures includingthe oral hood, and radialization of posterior skeletal ele-ments. These contrasting effects mirror the dual activ-ities of TGFb signaling in the border versus non-BE.Absent Nodal signaling, triradiates are nucleated in aradial pattern and these elongate into unstructuredrods. When BMP is knocked down, embryos have onlya partially radialized vegetal skeleton—extraspiculesform only around the dorsal aspect of the BE. Theseresults indicate a model where normally, growth factorsignaling is initially activated at the intersection of theBE and DVM (Fig. 2). Signaling from Nodal and BMPcontribute to the refinement of the BE subcompart-ments so that only the BE–DVM intersection containsthe correct information for VEGF and FGF production,thereby providing the site for skeletogenesis immedi-ately adjacent to that patch of cells.
A prediction from this model is that when Nodaland BMP are misexpressed throughout the ectoderm,BE gene expression and skeleton would be lost(because both Nodal and BMP repress BE genes).Indeed this is the case when BMP mRNA is injectedinto embryos (McIntyre and McClay, unpublisheddata). Interestingly, this is only partially true for Nodal.At high doses, Nodal does indeed block skeleton for-mation. However, at low doses embryos have robust,radialized skeletons. In these embryos endodermalWnt5 apparently is able to induce the BE and thatinduction is sufficient for signaling to initiate skeleto-genesis. The ability of Wnt5 to modify ventral ecto-derm specification explains the longstandingobservation that NiCl2 treated embryos have both aciliary band surrounding the vegetal plate of theembryo and radialized triradiate skeletal elements justinside the ciliary band (Hardin et al., 1992). Normally,the CB passes through the DVM, separating the ventralfrom dorsal ectoderm, but it also overlaps with theventral BE. In NiCl2-treated embryos Nodal isexpressed throughout the Ventral and Dorsal ecto-derm. The BE is specified, apparently because the levelof Nodal expression is not high enough to block theBE induction by Wnt5. However, in that situation thereis no BMP produced to separate Dorsal from Ventral,hence no DVM. Nevertheless, the BE is sufficient toallow triradiate skeletal elements to form in a radialpattern surrounding the embryo.
178 MCINTYRE ET AL.

ECTODERMAL SIGNALING TO THESKELETOGENIC CELLS
Over the past 10 years, a number of ectodermal geneshave been implicated in the process of skeleton forma-tion. These include the signaling ligands VEGF, FGF, andrecently, Wnt5 (Adomako-Ankomah and Ettensohn,2013; Duloquin et al., 2007; Knapp et al., 2012; McIn-tyre et al., 2013; Rottinger et al., 2008), as well as twotranscription factors, Otp and Pax 2/5/8 (Cavalieriet al., 2003; Saudemont et al., 2010; Rottinger et al.,2008) and the gene Trim1 (Cavalieri et al., 2011). Acommon feature of these genes is that they are allexpressed at the BE–DVM intersection, immediatelyadjacent to the ventrolateral skeletogenic mesodermclusters. Loss-of-function experiments indicate thateach gene is required for the skeleton to form.
The first molecular data explaining how the ecto-derm contributes to skeletogenesis was the cloning andfunctional analysis of VEGF (Duloquin et al., 2007).VEGF is expressed in the lateral BE–DVM intersection,and its receptor is exclusively expressed by the PMC’s;and VEGFR is particularly enriched in the PMCs foundwithin the ventrolateral clusters. Absent VEGF signal-ing, skeletal formation does not occur, and the PMCsappear disorganized within the blastocoel. EctopicVEGF mRNA resulted in extra skeletal branches, consist-ent with a role for this molecule in skeleton branching.However, based on these data alone, it was not possibleto determine the exact nature of the patterning infor-mation imparted to the PMCs by VEGF alone.
Soon after this, FGF was discovered to play a role inskeletogenesis as well. But unlike VEGF, FGF isexpressed in multiple territories as the embryo devel-ops (Rottinger et al., 2008). After an early period ofexpression at the vegetal pole, FGF is expressed in theBE–DVM intersection, and shortly thereafter isexpressed in the PMCs themselves especially at the dis-tal tips of the arms as the postoral rods elongate. Thiscomplex expression sequence has complicated analysisof its exact function because it is difficult to selectivelyperturb FGF signaling either in the DVM intersection orin the PMCs. However, when FGF expression wasblocked with a MASO, no skeleton was formed andexpression of Pax2/5/8 and Pea3 was lost (Rottingeret al., 2008). By contrast, a recent article suggests thatthe role of FGF in skeletogenesis is more limited andmay contribute little signaling information to the PMCsto initiate the triradiate (Adomako-Ankomah and Etten-sohn, 2013).
Wnt5, discussed above as an activator of BE fates,also has a phase of expression in the BE (McIntyreet al., 2013). After gastrulation Wnt5 expression movesfrom the endoderm to the dorsal and lateral BE. As gas-trulation proceeds, it is further restricted to two lateralpatches of the BE corresponding to the BE–DVM inter-
section where VEGF and FGF are expressed. This laterexpression of Wnt5 depends on earlier Wnt5 signalingfrom the endoderm (McIntyre et al., 2013). As such ithas been difficult to precisely distinguish this laterWnt5 function from the earlier inductive activity. It ispossible that at this stage, Wnt5 signals directly to thePMCs though current evidence suggests that the actionof Wnt5 on skeletogenesis is indirect via activation ofVEGF.
Although we have referred to the BE–DVM junctionas if it is a uniform patch of cells, that may not be true.VEGF, FGF, and Wnt5 are each expressed in that regionbut it is possible that the BE–DVM intersection actuallydoes not uniformly express the three signals. Each isexpressed in this region, but because there are smallembryo-to-embryo variations, it is difficult to distinguishbetween the BE–DVM as a uniform patch of cells, orwhether there are real differences between each sig-nal’s expression domain, and the additional possibilityexists that the patch undergoes dynamic changes. Forexample, if VEGF were expressed slightly more ven-trally than FGF, or if Wnt5 were shifted dorsally by twoor three cells, they may be able to provide moredetailed positional information than if they were per-fectly coincident. In other words the possibility remainsthat the BE–DVM intersection contains subdomains thathave functional significance. Recently, Adomako-Ankomah and Ettensohn (2013) examined expressionpatterns of VEGF and FGF mRNA using double fluores-cent WMISH. At mesenchyme blastula both mRNAswere restricted to the ectoderm over the ventrolateralclusters. During gastrula stages VEGF and FGF expres-sion overlapped briefly in the ectoderm over the ventro-lateral clusters (in the BE–DVM intersection), but bylate gastrula the expression of FGF ceased to beobserved in the ectoderm over the ventrolateral clus-ters and switched to being expressed in PMCs of theventrolateral cluster themselves (Adomako-Ankomahand Ettensohn, 2013). As often happens it is the detailsthat matter. At the site of the ventro-lateral clusters twosuccessive skeletal branching events occurs. The extentto which signaling from the BE–DVM controls that pre-cise patterning remains to be clarified.
An important question yet to be addressed is howeach of the signals is uniquely activated in the BE–DVMintersection. What are the immediate upstream tran-scriptional activators? Do different signals have thesame or different activators? The logic underlying acti-vation is of interest because it determines the timingand intensity of each signal, which in turn are neededfor proper patterning. Moreover, as the skeletal patternhas changed through evolution, it is likely that the acti-vation logic has been one target of selection.
Three GRN models of ectoderm specification in seaurchins have been published (Li et al., 2012; Saude-mont et al., 2010; Su et al., 2009). These network
LARVAL SKELETON PATTERNING 179

models do not include the D–V margin and BE as partof their models, and connections to VEGF, FGF, andWnt5 are either incomplete or missing. Knowledge ofexpression of transcription factors and the signals inthe BE and DVM help complete this analysis, thoughthe number of transcription factors involved may yet beincomplete, and perturbation analyses establishing epi-static relationships have yet to be published.
EVOLUTION OF SKELETAL PATTERNING
Once the GRN operating at the intersection of the BEand DVM is understood, and once the position and pat-tern of the skeletal rudiment is known in greater detail,it will become possible to ask how evolution of differ-ent patterns occurred. It has been suggested thatbecause the echinoderm crown group ancestor pos-sessed an auricularian-like larva with no skeleton(Nakano et al., 2003), that the larval skeleton in echi-noids is the result of a heterochronic shift that activatesthe adult skeletogenic program early in development(Ettensohn, 2009; Gao and Davidson, 2008). Somehowthis skeletogenic program was integrated with existingGRNs that govern the placements of the DVM territoryand the BE territory. To appreciate how this happenedit would be of great value to know the expression pat-terns of genes in these territories in echinoidoutgroups.
As yet the expression patterns of several key playersincluding Pax2/5/8, Otp and FGF have not been exam-ined in other echinoderms. But, the expression of VEGFand VEGFR has been examined in sea stars (class Aster-oidea), which do not have larval skeletons (Morinoet al., 2012). Transcripts were not detected by in situ
hybridization or quantitative polymerase chain reactionin sea stars before the bipinnaria larval stage. At the bra-chiolaria stage VEGF (expressed in the ectoderm) andVEGFR (expressed in the mesoderm) were associatedwith the rudiments of the adult skeleton. Thus, we canhypothesize that the heterochronic activation of VEGFand VEGFR in the euechinoid ancestor was an equallyessential step in the evolution of the larval skeleton aswas the heterochronic activation of the skeletogenicprogram itself.
Brittle stars (class Ophiuroidea) also have a larvalskeleton that is considered to have emerged independ-ently from that in euechinoids (Williamson, 2003),which if true, makes them an excellent additional testcase for asking how the adult skeletogenic program andectoderm/mesoderm communication system evolved.In contrast to echinoids, the brittle star skeletogenic lin-eage does not arise from an asymmetric cell division at4th cleavage. Instead, the skeletogenic cells, also calledPMCs, become obvious inside the blastocoel before gas-trulation and form bilateral rudiments of the larval skel-eton (Yamashita, 1985). Brittle star homologs of VEGF
are expressed in the ectoderm in bilateral patches verymuch like the pattern in sea urchins, and VEGFR is like-wise expressed in PMC ventrolateral clusters (Morinoet al., 2012). As in sea urchins, the expression of brittlestar VEGF is initially expressed in a ring in the ectodermbefore gastrulation, but then expression is restricted tothe lateral patches. During prism stages VEGF andVEGFR continue to be expressed at the tips of thegrowing arms, in the ectoderm and mesoderm, respec-tively. This remarkable similarity in expression patternsof VEGF and VEGFR between sea urchins and brittlestars suggests that either the conventional wisdom thatthe larval skeletons are independently derived in thesetwo groups is wrong, or suggests that heterochronicactivation of this pathway was essential during inde-pendent innovations of the larval skeleton is bothclades.
As more genes are examined in brittle stars, and inmore species, a clearer picture will emerge. Ideally, itwould be of value to assess how the whole network sur-rounding VEGF signaling operates relative to seaurchins. Recently, Vaughn et al. (2012) generated a tran-scriptome of the gastrula stage of a brittle star andexamined its complement of “network” genes and skel-etogenic genes. A comparison of territory-specific GRNcomponents expressed in brittle star gastrulae showedthat the highest conservation between sea urchin andbrittle star were those genes in the skeletogenic GRN,suggesting that convergent evolution of the larval skele-ton in brittle stars may have occurred. They detectedVEGF transcripts in the gastrula, but not VEGFR. Also, anumber of genes that are present in the BE in seaurchins, such as Irx, Msx, Lim, and Nk1, were not repre-sented in their gastrula transcriptome. Nodal was notpresent in the transcriptome, but BMP2/4, Hnf6, andFoxj1 were. These differences could indicate large dif-ferences in the way the BE is established in brittle starsbut until a detailed analysis of brittle star skeletogenesisand ectodermal signaling is conducted evolutionaryinsights will remain rudimentary.
While the phylogenetic relationship of brittle starsand sea urchins might still be under debate, it is wellestablished that the sea cucumbers (class Holothuroi-dea) branch closest to the euechinoids (Janies et al.,2011; Pisani et al., 2012). Their larval form does nothave an asymmetric early cleavage resulting in micro-meres, nor a well-developed larval skeleton. However,in some species the auricularian larvae possess poste-rior spicules, which are thought to act as ballast forproper orientation in the water column (Penningtonand Strathmann, 1990). McCauley et al. (2010) investi-gated the developmental and molecular origin of spi-cules in the larva of the sea cucumber Parastichopus.This species makes a very small spicule granule in theblastocoel underneath the posterior dorsal ectoderm.They showed a group of Alx1-positive cells ingress into
180 MCINTYRE ET AL.

the blastocoel, migrate away from the archenteron,form a dorsal chain of cells and then coalesce into acluster of approximately 8 cells that corresponds to theposition of the spicule. Knockdown of Parastichopus
Alx1 with an antisense morpholino abolished this dor-sal cluster and the spicule, suggesting that like seaurchins, Alx1 is involved in the skeletogenic fate. Iftrue, one might expect holothurian skeletogenesis touse additional orthologs of genes used by sea urchins.
GOING FORWARD: BUILDING THE 3D SKELETON
Once the triradiate skeleton is established, the skeletalrods elongate to form the three-dimensional frameworkof the larva. The rods either grow parallel to the DVMor BE, or, the postoral and anterolateral rods grow outventrally, in each case distorting a patch of one of thesebands, which expands in parallel with rod growth.Growth of each type of rod is remarkably stereotypicalbetween embryos, but differs predictably between thevarious types of rods. Guss and Ettensohn (1997) meas-ured rates of skeletal elongation and showed that mostrods have a basal elongation rate of �6 mm/h at 23�C.However, certain rods, notably the postoral and antero-lateral rods, accelerate their rate of growth later indevelopment. In addition, the ventral transverse rodceases to elongate well before the end of embryogene-sis. The authors concluded that mesenchyme cells usean autonomous program for rod production but extrin-sic cues modify the basal activity.
Once the skeleton is initiated in the BE–DVM inter-section it then appears to use both bands for much ofits patterning since all skeletal elements except those inthe anterior/oral hood grow parallel to these bands.How might that work? If we take all the information onlocalized cues (Armstrong et al., 1993; Duloquin et al.,2007; Guss and Ettensohn, 1997; McIntyre et al., 2013;Rottinger et al., 2008), and combine that informationwith data from studies on biomineralization (Illies et al.,2002; Killian and Wilt, 2008; Knapp et al., 2012; Living-ston et al., 2006; Urry et al., 2000; Wilt, 2005), and alsoadd in a number of earlier studies using experimentalembryology (Armstrong et al., 1993; Armstrong andMcClay, 1994; Guss and Ettensohn, 1997; Hardin andArmstrong, 1997; Hardin et al., 1992; Hodor and Etten-sohn, 1998; Malinda and Ettensohn, 1994; Malindaet al., 1995; Peterson and McClay, 2003), we can beginto outline mechanisms used to build a skeleton. The fol-lowing model is preliminary and probably misses manycomponents of skeletal growth, but will be useful indirecting future studies on how skeletal patterning mayfunction in this embryo.
Since the pattern of the calcite crystal depositeddepends on the shape of the syncytial cavity of PMCcell extensions, much of the patterning model hingeson how PMCs organize the syncitium. PMCs initially
form as a ring at the posterior end of the blastocoel. For-mation of the PMC ring likely is guided by VEGF, whichat an early stage is expressed in a matching ring allaround the BE (Adomako-Ankomah and Ettensohn,2013; McIntyre et al., 2013). When that ring first formsthe PMCs distribute themselves roughly equally aroundthe circumference of the posterior ectoderm. It is dur-ing this stage that the PMCs extend long thin filopodiato touch each other and to touch the ectoderm beneath(the BE). They then initiate fusion to form the syncy-tium (Hodor and Ettensohn, 1998; Miller et al., 1995).The ventrolateral clusters form by PMCs migrating later-ally from both the dorsal and ventral sides of the ring(Peterson and McClay, 2003). Formation of the ventro-lateral clusters is directed by secretion of VEGF and/orFGF from the BE–DVM intersection (Duloquin et al.,2007; McIntyre et al., 2013; Rottinger et al., 2008). Ifeither signal, or loss of the receptor for either signal ismissing, the PMCs fail to form the ventrolateral clusters.The next step is formation of the initial triradiate spi-cule. This process appears to be tightly controlled bysignaling, though the mechanism by which this hap-pens is less clear. VEGF appears to promote nucleationand branching (Knapp et al., 2012; McIntyre et al.,2013); if VEGF is knocked down embryos do not makeskeletons, and overexpressed VEGF results in largenumbers of ectopic, branched skeletal elements. FGFknockdown embryos produce triradiate skeletons butskeletal rods do not elongate (Rho and McClay, 2011),though in Paracentrotus FGF knockdown appears toeliminate even the triradiate (Rottinger et al., 2008).
The final embryonic skeleton emerges as the rodselongate and branch once more as the ventral/oralhood forms. The result is a bilaterally symmetric struc-ture, each half of which is formed from a single crystalof calcite. The work of Okazaki (1960, 1975) and Oka-zaki and Inoue (1976) demonstrated that each half ofthe skeleton was a single crystal of calcite whichaccounted for the unique birefringence of the skeletalrods. The optical axis of the crystal is always aligned inthe same manner, with the c-axis parallel to the postoral/body rods and with the three arms of the initial trir-adiate extending in the plane perpendicular to the c-axis. The precise, reproducible orientation of the crys-tal suggests the process is closely regulated, and it willbe crucial to explain how the polarity of this crystal isestablished (especially given that its scale is so muchsmaller than that of the cluster of mesenchyme cellbodies which deposit it. Important for biomineraliza-tion, once the initial axis is formed, it is retained as thecrystal grows because when added, ACC reaches thepre-existing crystal, and joins the crystal entrained bythe initial crystal axes.
It is thought that the skeleton is deposited in a mold,the PMC syncytium. When the skeletal rods are observedwith an electron microscope, the surface of each rod is
LARVAL SKELETON PATTERNING 181

visibly smooth, showing no edges characteristic of grow-ing crystals (Okazaki, 1960, 1975; Okazaki and Inoue,1976). However, when such a rod is used as a nucleusfor artificial growth, crystal edges readily appear as thein vitro crystal grows and the crystal growth is entirelyin parallel planes indicating that the smooth rod never-theless is a single crystal. When rods are fractured, thecore of the rod appears to have concentric growthlayers, much like rings on a tree. These observations sug-gested that the crystals are deposited in several stages.The ACC is inherently unstable, and in vitro will rapidly(within minutes) crystallize into calcite (Brecevic andNielsen, 1989). Sea urchins and other marine inverte-brates stabilize ACC with the addition of proteins, storethe ACC intracellularly, and then deliver it to the growingskeleton though intracellular vesicles where it is inducedto crystallize (Beniash et al., 1999; Decker et al., 1987;Gong et al., 2012; Wilt, 2005). The process of inductionis not understood, but the ACC is added to the existingrod and may require changes to the proteins incorpo-rated in the ACC. There are up to around 50 proteinsincorporated into the ACC (Adomako-Ankomah andEttensohn, 2011; Rafiq et al., 2012), so there is plenty ofroom for regulative activities in skeletal growth. Maha-mid et al. (2011) speculate that perhaps the same mole-cules could be responsible for both stabilizing ACC andinducing calcite crystal formation after some type ofstructural change. Because of the use of ACC, the crystalcan take the shape of whatever structure contains it atthe time the ACC is induced to crystallize. All this sug-gests the crystals are NOT grown in a particular shape,but rather deposited in a mold, the PMC syncytial cavity.The PMCs appear, therefore, to act as a biological tem-plate, which is consistent with how Calcium carbonateis deposited in mollusks (Addadi et al., 2006; Wilt,2005). In fact Addadi et al. conclude that template-directed spicule nucleation occurs in both sea urchinsand mollusks. The question becomes what governs theplacement of PMC chains, and tells which cells shouldfuse together? That question returns focus to the BE andDVM.
Important clues of function come from interspeciesexperiments (Armstrong et al., 1993; Armstrong andMcClay, 1994). Lytechinus PMCs were inserted into theblastocoel of Tripneustes esculentus in place of Trip-
neustes PMCs, or the reciprocal recombination wasdone. In both combinations the PMCs were able tointerpret the ectoderm signals of the other species toproduce the three-dimensional shape of the skeleton,i.e., the ventrolateral clusters formed at the correctlocation, and the skeleton grew along the BE and DVM.Thus, the ectodermal signaling inputs were well con-served over an evolutionary distance of more than 10million years. Normally Tripneustes forms fenestratedpostoral skeleton rods while all Lytechinus postoralskeletal rods are simple. In the recombinants the Trip-
neustes PMCs produced fenestrated postoral rods whilethe Lytechinus PMCs produced simple rods. Thus theactual rod type is determined by the genotype of thePMCs. Also determined by the genotype of the PMCswas the extent of the rods produced. As evidence, Trip-
neustes forms long recurrent rods while the recurrentrods in the Lytechinus skeleton are bent and stunted. Inthe recombinants the Tripneustes recurrent rodsformed at the correct location and were correctly pat-terned (requiring ectoderm input for placement), whilethe Lytechinus recurrent rods were stunted whengrown in Tripneustes, reflecting the autonomous read-out of Lytechinus genotypic information in response toTripneustes ectodermal positional cues. Of note, theLytechinus ectoderm provided spatial signals to Trip-
neustes even though the Tripneustes skeleton grew inan area where no Lytechinus skeleton grows. This sug-gests that the ectoderm provides spatial signals essen-tially telling the skeletogenic cells where their locationin the embryo (positional information), and it providesgrowth factors that promote skeletal growth. The PMCsgenotypically provide the template for the actual skele-tal element by virtue of the syncitial folds in which theskeletal rods grow: a simple template for simple rods, afenestrated template for fenestrated rods. Thus, thethree-dimensional skeleton is a combination of spatialcues and growth factor provision from the ectodermplus a built in response mechanism that allows the PMCsyncytium and the proteins produced by the PMCs tomold the correct skeletal element in any position.
This then has been the thesis of this article: signalingcues from the ectoderm provide the spatial informationfor the PMC syncytium template using two belts of ecto-derm, the DVM and the BE. That means ectodermpatterning ultimately is responsible for the three-dimensional shape of the larval skeleton. The challengeis to understand the many levels of control that sets upthe BE and D–V margin as specific territories in the ecto-derm, then to understand how the DVM and BE providethe correct spatial cues to the PMCs, at the correct time,and finally, how the PMCs read, interpret, and then formthe template for the skeletal element they produce.
ACKNOWLEDGMENTS
The authors appreciate the input provided by membersof the McClay laboratory.
LITERATURE CITED
Addadi L, Joester D, Nudelman F, Weiner S. 2006. Mol-lusk shell formation: A source of new concepts forunderstanding biomineralization processes. Chemis-try 12:980–987.
Adomako-Ankomah A, Ettensohn CA. 2011. P58-A andP58-B: Novel proteins that mediate skeletogenesisin the sea urchin embryo. Dev Biol 353:81–93.
182 MCINTYRE ET AL.

Adomako-Ankomah A, Ettensohn CA. 2013. Growthfactor-mediated mesodermal cell guidance and skel-etogenesis during sea urchin gastrulation. Develop-ment 140:4214–4225.
Angerer LM, Yaguchi S, Angerer RC, Burke RD. 2011.The evolution of nervous system patterning:Insights from sea urchin development. Develop-ment 138:3613–3623.
Armstrong N, Hardin J, McClay DR. 1993. Cell–cellinteractions regulate skeleton formation in the seaurchin embryo. Development 119:833–840.
Armstrong N, McClay DR. 1994. Skeletal pattern is speci-fied autonomously by the primary mesenchyme cellsin sea urchin embryos. Dev Biol 162:329–338.
Baltzer F, Chen PS, Whiteley AH. 1959. Biochemicalstudies on sea urchin hybrids. Exp Cell Res Suppl 6:192–209.
Beniash E, Addadi L, Weiner S. 1999. Cellular controlover spicule formation in sea urchin embryos: Astructural approach. J Struct Biol 125:50–62.
Boveri T. 1901a. Die Polaritat von oocyte, ei und larvedes Strongylocentrotus lividus. Zool Jahrb Abt AnatOntog Tiere 14:630–653.
Boveri T. 1901b. Uber die Polaritat des Seeigel-Eies.Verh Phys Med Ges Wurzburg 34:145–176.
Bradham CA, McClay DR. 2006. p38 MAPK is essentialfor secondary axis specification and patterning insea urchin embryos. Development 133:21–32.
Brecevic L, Nielsen AE. 1989. Solubility of amorphouscalcium carbonate. J Cryst Growth 98:504–510.
Burke RD, Angerer LM, Elphick MR, Humphrey GW,Yaguchi S, Kiyama T, Liang S, Mu X, Agca C, KleinWH, Brandhorst BP, Rowe M, Wilson K, ChurcherAM, Taylor JS, Chen N, Murray G, Wang D, MellottD, Olinski R, Hallb€o€ok F, Thorndyke MC. 2006. Agenomic view of the sea urchin nervous system.Dev Biol 300:434–460.
Cavalieri V, Guarcello R, Spinelli G. 2011. Specific expres-sion of a TRIM-containing factor in ectoderm cellsaffects the skeletal morphogenetic program of thesea urchin embryo. Development 138:4279–4290.
Cavalieri V, Spinelli G, Di Bernardo M. 2003. ImpairingOtp homeodomain function in oral ectoderm cellsaffects skeletogenesis in sea urchin embryos. DevBiol 262:107–118.
Coffman JA, Davidson EH. 2001. Oral-aboral axis specifi-cation in the sea urchin embryo. I. Axis entrainmentby respiratory asymmetry. Dev Biol 230:18–28.
Czerny T, Bouchard M, Kozmik Z, Busslinger M. 1997.The characterization of novel Pax genes of the seaurchin and Drosophila reveal an ancient evolutionaryorigin of the Pax2/5/8 subfamily. Mech Dev 67:179–192.
Davidson EH. 1989. Lineage-specific gene expression andthe regulative capacities of the sea urchin embryo: Aproposed mechanism. Development 105:421–445.
Di Bernardo M, Castagnetti S, Bellomonte D, Oliveri P,Melfi R, Palla F, Spinelli G. 1999. Spatially restrictedexpression of PlOtp, a Paracentrotus lividus
orthopedia-related homeobox gene, is correlatedwith oral ectodermal patterning and skeletal mor-phogenesis in late-cleavage sea urchin embryos.Development 126:2171–2179.
Decker GL, Morrill JB, Lennarz WJ. 1987. Characteriza-tion of sea urchin primary mesenchyme cells andspicules during biomineralization in vitro. Develop-ment 101:297–312.
Duboc V, Rottinger E, Besnardeau L, Lepage T. 2004.Nodal and BMP2/4 signaling organizes the oral-aboral axis of the sea urchin embryo. Dev Cell 6:397–410.
Duloquin L, Lhomond G, Gache C. 2007. LocalizedVEGF signaling from ectoderm to mesenchyme cellscontrols morphogenesis of the sea urchin embryoskeleton. Development 134:2293–2302.
Emily-Fenouil F, Ghiglione C, Lhomond G, Lepage T,Gache C. 1998. GSK3beta/shaggy mediates pattern-ing along the animal-vegetal axis of the sea urchinembryo. Development 125:2489–2498.
Ettensohn CA. 2009. Lessons from a gene regulatorynetwork: Echinoderm skeletogenesis providesinsights into evolution, plasticity and morphogene-sis. Development 136:11–21.
Gao F, Davidson EH. 2008. Transfer of a large gene regu-latory apparatus to a new developmental address inechinoid evolution. Proc Natl Acad Sci USA 105:6091–6096.
Gong YU, Killian CE, Olson IC, Appathurai NP, AmasinoAL, Martin MC, Holt LJ, Wilt FH, Gilbert PU. 2012.Phase transitions in biogenic amorphous calciumcarbonate. Proc Natl Acad Sci USA 109:6088–6093.
Gross JM, McClay DR. 2001. The role of Brachyury (T)during gastrulation movements in the sea urchinLytechinus variegatus. Dev Biol 239:132–147.
Guss KA, Ettensohn CA. 1997. Skeletal morphogenesisin the sea urchin embryo: Regulation of primarymesenchyme gene expression and skeletal rodgrowth by ectoderm-derived cues. Development124:1899–1908.
Gustafson T, Wolpert L. 1961. Studies on the cellularbasis of morphogenesis in the sea urchin embryo;directed movements of primary mesenchyme cellsin normal and vegetalized larvae. Exp Cell Res 24:64–79.
Hardin J, Armstrong N. 1997. Short-range cell–cell sig-nals control ectodermal patterning in the oralregion of the sea urchin embryo. Dev Biol 182:134–149.
Hardin J, Coffman JA, Black SD, McClay DR. 1992. Com-mitment along the dorsoventral axis of the seaurchin embryo is altered in response to NiCl2.Development 116:671–685.
LARVAL SKELETON PATTERNING 183

Ho HY, Susman MW, Bikoff JB, Ryu YK, Jonas AM, Hu L,Kuruvilla R, Greenberg ME. 2012. Wnt5a-Ror-Dish-evelled signaling constitutes a core developmentalpathway that controls tissue morphogenesis. ProcNatl Acad Sci USA 109:4044–4051.
Hodor PG, Ettensohn CA. 1998. The dynamics and regu-lation of mesenchymal cell fusion in the sea urchinembryo. Dev Biol 199:111–124.
Horstadius S. 1939. The mechanics of sea urchin devel-opment as studied by operative methods. Biol. Rev.14:132–179.
H€orstadius S. 1973. Experimental embryology of echi-noderms. Oxford: Clarendon Press.
H€orstadius S. 1935. €Uber die determination im verlaufeder eiachse bei seeigeln. Pubbl Staz Nool Napoli 14:251–429.
Illies MR, Peeler MT, Dechtiaruk AM, Ettensohn CA.2002. Identification and developmental expressionof new biomineralization proteins in the sea urchinStrongylocentrotus purpuratus. Dev Genes Evol212:419–431.
Janies DA, Voight JR, Daly M. 2011. Echinoderm phylog-eny including Xyloplax, a progenetic asteroid. SystBiol 60:420–438.
Kilian B, Mansukoski H, Barbosa FC, Ulrich F, Tada M,Heisenberg CP. 2003. The role of Ppt/Wnt5 in regu-lating cell shape and movement during zebrafishgastrulation. Mech Dev 120:467–476.
Killian CE, Wilt FH. 2008. Molecular aspects of biomi-neralization of the echinoderm endoskeleton. ChemRev 108:4463–4474.
Kinjo S, Shirayama Y, Wada H. 2008. Evolutionary his-tory of larval skeletal morphology in sea urchinEchinometridae (Echinoidea: Echinodermata) asdeduced from mitochondrial DNA molecular phy-logeny. Evol Dev 10:632–641.
Knapp RT, Wu CH, Mobilia KC, Joester D. 2012.Recombinant sea urchin vascular endothelial growthfactor directs single-crystal growth and branching invitro. J Am Chem Soc 134:17908–17911.
Li E, Materna SC, Davidson EH. 2012. Direct and indi-rect control of oral ectoderm regulatory geneexpression by Nodal signaling in the sea urchinembryo. Dev Biol 369:377–385.
Livingston BT, Killian CE, Wilt F, Cameron A, LandrumMJ, Ermolaeva O, Sapojnikov V, Maglott DR,Buchanan AM, Ettensohn CA. 2006. A genome-wideanalysis of biomineralization-related proteins in thesea urchin Strongylocentrotus purpuratus. DevBiol 300:335–348.
Logan CY, Miller JR, Ferkowicz MJ, McClay DR. 1999.Nuclear beta-catenin is required to specify vegetalcell fates in the sea urchin embryo. Development126:345–357.
Lyons DC, Kaltenbach SL, McClay DR. 2012. Morpho-genesis in sea urchin embryos: Linking cellular
events to gene regulatory network states. WileyInterdiscip Rev Dev Biol 1:231–252.
Mahamid J, Sharir A, Gur D, Zelzer E, Addadi L, WeinerS. 2011. Bone mineralization proceeds throughintracellular calcium phosphate loaded vesicles: Acryo-electron microscopy study. J Struct Biol 174:527–535.
Malinda KM, Ettensohn CA. 1994. Primary mesenchymecell migration in the sea urchin embryo: Distribu-tion of directional cues. Dev Biol 164:562–578.
Malinda KM, Fisher GW, Ettensohn CA. 1995. Four-dimensional microscopic analysis of the filopodialbehavior of primary mesenchyme cells during gas-trulation in the sea urchin embryo. Dev Biol 172:552–566.
McCauley BS, Weideman EP, Hinman VF. 2010. A con-served gene regulatory network subcircuit drivesdifferent developmental fates in the vegetal pole ofhighly divergent echinoderm embryos. Dev Biol340:200–208.
McClay DR. 2011. Evolutionary crossroads in develop-mental biology: Sea urchins. Development 138:2639–2648.
McIntyre DC, Seay NW, Croce JC, McClay DR. 2013.Short-range Wnt5 signaling initiates specification ofsea urchin posterior ectoderm. Development 140:4881–4889.
Miller J, Fraser SE, McClay D. 1995. Dynamics of thinfilopodia during sea urchin gastrulation. Develop-ment 121:2501–2511.
Morino Y, Koga H, Tachibana K, Shoguchi E, KiyomotoM, Wada H. 2012. Heterochronic activation of VEGFsignaling and the evolution of the skeleton in echi-noderm pluteus larvae. Evol Dev 14:428–436.
Nakajima Y, Kaneko H, Murray G, Burke RD. 2004.Divergent patterns of neural development in larvalechinoids and asteroids. Evol Dev 6:95–104.
Nakano H, Hibino T, Oji T, Hara Y, Amemiya S. 2003.Larval stages of a living sea lily (stalked crinoid echi-noderm). Nature 421:158–160.
Okazaki K. 1960. Skeleton formation of the sea urchinlarvae. II. Organic matrix of the spicule. Embryolog-ica 5:283–320.
Okazaki K. 1975. Spicule formation by isolated micro-meres of the sea urchin embryo. Am Zool 15:567–581.
Okazaki K, Inoue S. 1976. Crystal property of the larvalsea urchin spicule. Dev Growth Differ 18:413–434.
Oliveri P, Davidson EH, McClay DR. 2003. Activation ofpmar1 controls specification of micromeres in thesea urchin embryo. Dev Biol 258:32–43.
Oliveri P, Tu Q, Davidson EH. 2008. Global regulatorylogic for specification of an embryonic cell lineage.Proc Natl Acad Sci USA 105:5955–5962.
Oliveri P, Walton KD, Davidson EH, McClay DR. 2006.Repression of mesodermal fate by foxa, a key
184 MCINTYRE ET AL.

endoderm regulator of the sea urchin embryo.Development 133:4173–4181.
Pennington JT, Strathmann RR. 1990. Consequences ofthe calcite skeletons of planktonic echinoderm lar-vae for orientation, swimming, and shape. Biol Bull179:121–133.
Peter IS, Davidson EH. 2010. The endoderm gene regu-latory network in sea urchin embryos up to mid-blastula stage. Dev Biol 340:188–199.
Peter IS, Davidson EH. 2011. A gene regulatory networkcontrolling the embryonic specification of endo-derm. Nature 474:635–639.
Peterson RE, McClay DR. 2003. Primary mesenchymecell patterning during the early stages followingingression. Dev Biol 254:68–78.
Pisani D, Feuda R, Peterson KJ, Smith AB. 2012. Resolvingphylogenetic signal from noise when divergence israpid: A new look at the old problem of echinodermclass relationships. Mol Phylogenet Evol 62:27–34.
Rafiq K, Cheers MS, Ettensohn CA. 2012. The genomicregulatory control of skeletal morphogenesis in thesea urchin. Development 139:579–590.
Range RC, Angerer RC, Angerer LM. 2013. Integrationof canonical and noncanonical Wnt signaling path-ways patterns the neuroectoderm along the ante-rior–posterior axis of sea urchin embryos. PLoS Biol11:e1001467.
Rho HK, McClay DR. 2011. The control of foxN2/3expression in sea urchin embryos and its functionin the skeletogenic gene regulatory network. Devel-opment 138:937–945.
Rottinger E, Saudemont A, Duboc V, Besnardeau L,McClay D, Lepage T. 2008. FGF signals guide migra-tion of mesenchymal cells, control skeletal morpho-genesis [corrected] and regulate gastrulation duringsea urchin development. Development 135:353–365.
Saudemont A, Haillot E, Mekpoh F, Bessodes N, QuirinM, Lapraz F, Duboc V, Rottinger E, Range R, Oisel A,Besnardeau L, Wincker P, Lepage T. 2010. Ancestralregulatory circuits governing ectoderm patterningdownstream of Nodal and BMP2/4 revealed by generegulatory network analysis in an echinoderm. PLoSGenet 6:e1001259.
Sherwood DR, McClay DR. 1997. Identification andlocalization of a sea urchin Notch homologue:Insights into vegetal plate regionalization andNotch receptor regulation. Development 124:3363–3374.
Sherwood DR, McClay DR. 1999. LvNotch signalingmediates secondary mesenchyme specification in thesea urchin embryo. Development 126:1703–1713.
Strathmann RR. 2000. Functional design in the evolu-tion of embryos and larvae. Semin Cell Dev Biol 11:395–402.
Strathmann RR. 2007. Time and extent of ciliaryresponse to particles in a non-filtering feedingmechanism. Biol Bull 212:93–103.
Strathmann RR, Grunbaum D. 2006. Good eaters, poorswimmers: Compromises in larval form. IntegrComp Biol 46:312–322.
Su YH, Li E, Geiss GK, Longabaugh WJ, Kramer A,Davidson EH. 2009. A perturbation model of thegene regulatory network for oral and aboral ecto-derm specification in the sea urchin embryo. DevBiol 329:410–421.
Sweet HC, Gehring M, Ettensohn CA. 2002. LvDeltais a mesoderm-inducing signal in the sea urchinembryo and can endow blastomeres withorganizer-like properties. Development 129:1945–1955.
Urry LA, Hamilton PC, Killian CE, Wilt FH. 2000.Expression of spicule matrix proteins in the seaurchin embryo during normal and experimentallyaltered spiculogenesis. Dev Biol 225:201–213.
Vaughn R, Garnhart N, Garey JR, Thomas WK,Livingston BT. 2012. Sequencing and analysis of thegastrula transcriptome of the brittle star Ophiocomawendtii. Evo Dev 3:19.
von Ubisch Lv. 1933. Keimblattchimaren. Die Naturwis-senschaften 18:325–329.
von Ubisch Lv. 1939. Keimblattchimarenforschung anSeeigellarven. Biol Rev 14:88–103.
Wei Z, Range R, Angerer R, Angerer L. 2012. Axial pat-terning interactions in the sea urchin embryo: Sup-pression of Nodal by Wnt1 signaling. Development139:1662–1669.
Wikramanayake AH, Huang L, Klein WH. 1998. Beta-cat-enin is essential for patterning the maternally speci-fied animal-vegetal axis in the sea urchin embryo.Proc Natl Acad Sci USA 95:9343–9348.
Williamson DI. 2003. The origins of larvae. Dordrecht,Netherlands: Kluwer Academic Publishers.
Wilt FH. 2005. Developmental biology meets materialsscience: Morphogenesis of biomineralized struc-tures. Dev Biol 280:15–25.
Wilt FH, Killian CE, Livingston BT. 2003. Developmentof calcareous skeletal elements in invertebrates. Dif-ferentiation 71:237–250.
Wray GA. 1992. Rates of evolution in developmentalprocesses. Am Zool 32:123–134.
Yamanaka H, Moriguchi T, Masuyama N, Kusakabe M,Hanafusa H, Takada R, Takada S, Nishida E. 2002.JNK functions in the non-canonical Wnt pathway toregulate convergent extension movements in verte-brates. EMBO Rep 3:69–75.
Yamashita M. 1985. Embryonic development of thebrittle-star Amphipholis kochii in laboratory culture.Biol Bull 169:131–142.
LARVAL SKELETON PATTERNING 185