bioactivity of plants secondary metabolites...
TRANSCRIPT

BIOACTIVITY OF PLANTS SECONDARY METABOLITES
Estrogenic, cytotoxic and anabolic effects on estrogen
target organs of an extract of Erythrina excelsa and
Ecdysterone
A thesis presented to:
The Institute of Cardiovascular Research and Sports Medicine
Department of Molecular and Cellular Sports Medicine
German Sport University Cologne
In partial fulfillment of the requirements for the degree
PhD in Natural Sciences
by:
Sadrine Tchoukouegno Ngueu
From Bamenyam, Cameroon

Members of the jury:
- Prof. Dr. Patrick Diel (First referee)
- Prof. Dr. Günter Vollmer (Second referee)
- Prof. Dr. Wilhelm Schänzer
- Dr. Frank Hülsemann
Date of defence: 05.09.2013

Gedruckt mit Unterstützung des Deutschen Akademischen Austauschdienstes (DAAD)

Versicherung gem. § 7 Abs. 2 Nr. 4
Hierdurch versichere ich:
Ich habe diese Arbeit selbständig und nur unter Benutzung der angegebenen
Quellen und technischen Hilfen angefertigt; sie hat noch keiner anderen Stelle zur
Prüfung vorgelegen. Wörtlich übernommene Textstellen, auch Einzelsätze oder Teile
davon, sind als Zitate kenntlich gemacht worden.
……………………………………
Sadrine Tchoukouegno Ngueu
Erklärung gem. § 7 Abs. 2 Nr. 5
Hierdurch erkläre ich, dass ich die Leitlinien guter wissenschaftlicher Praxis der
DeutschenSporthochschule Köln in der aktuellen Fassung eingehalten habe.
……………………………………
Sadrine Tchoukouegno Ngueu

I dedicate this work to:
God almighty and to Jesus Christ my Lord and
Savior in whom I trust.
My lovely husband Olivier Ndiapi for his
endless support through happy and difficult
times. These 4 years away from each other
were challenging but your support was always
an encouragement to me.

Acknowledgements
M.Sc. Sadrine Tchoukouegno Ngueu vi
Acknowledgements
I would not have completed this work without the help and support of several
persons. Foremost, special thanks go to my supervisor Prof.Dr. Patrick Diel who did
not only give me his support throughout the completion of my thesis work, but also
supported me with the DAAD scholarship application procedure. I extend my thanks
to Prof. Guenter Vollmer for his helpful pieces of advice regarding my work and who,
during these years, wrote letters of recommandation that helped extend my
scholarship. I am also grateful to Prof. Dr. Maria Paar who guided me through my first
chromatographic separation.
I thank the German academic service exchange (DAAD) for granting me the
scholarship.
I had the opportunity to work in a team composed of persons from different
horizons and experiences which was very helpful not only for my academic
advancement, but also contributed to build certain traits of my character. I think of my
colleagues Jonas Hengevoß, Xiao Juan Hu, Sephanie Mosler, Felix Kluxen, Nora
Zoth, Almut Molzberger, Carmen Weigt, Tina Blei, Dennis Mueller, and Martina
Velders. I specially thank Felix Kluxen not only for the helpful tips in the lab, but also
for his availability and patience towards me during my first days in Germany.
My gratitude also goes to our technical assistant crew composed of Anika
Voß, Ute Laudenbach-Leschowski, Ghilav Mojgan, and Bianca Collins who were
always available to assist me in the pratical aspects of laboratory work. Special
thanks go to Ute Laudenbach for her support in all aspects of animal experiments.
I would also like to acknowledge the support of my former Master degree
supervisor, Prof. Njamen Dieudonne, during my scholarship application process and
during my PhD thesis work.

Acknowledgements
M.Sc. Sadrine Tchoukouegno Ngueu vii
Special thanks to Maria Halabalaki (School of pharmacy, University of Athens)
for her support in my work and for granting me the opportunity to carry out an
internship in the pharmacognosy laboratory of the University of Athens.
I will not forget Job Tchoumtchoua who has been more than a friend to me.
Thank you for the long discussions over the phone and for introducing and
supervising me in your lab in Athens.
I thank Raoul Kempkes of the Biochemistry department for his technical
support in the use of the flow cytometer.
Finally, I thank my family for their continuous support in the course of my
education. I think also of my late parents and my late elder brother who took me as
he his own child and guided me throughout my studies.

Zusammenfassung
M.Sc. Sadrine Tchoukouegno Ngueu viii
Zusammenfassung
Heilpflanzen dienen als primäre Gesundheitsversorgung in den meisten
asiatischen und afrikanischen Ländern. In westlich geprägten Ländern in denen
hauptsächlich auf naturwissenschaftlichen Grundsätzen basierende Medizin
praktiziert wird, werden Stoffe aus pflanzlicher Herkunft entweder direkt als
Kräuterformulierungen oder als Ergänzung zu bestehenden Therapien verwendet.
Daneben können Pflanzen mit aktiven Wirkstoffen auch Teil der Ernährung sein. Es
besteht die allgemeine Überzeugung, dass pflanzliche Heilmittel aufgrund ihres
natürlichen Ursprungs sicher sind. Daher werden sie oft im Rahmen
selbsttherapeutischer Maßnahmen verwendet und sind in Deutschland auch nicht
rezeptpflichtig. Diese Arbeit beschäftigt sich mit der Bioaktivität, Wirkungen aber auch
potentiellen Nebenwirkungen derartiger pflanzlicher Heilmittel. Hierbei stehen
Extrakte aus Erythrina excelsa sowie die Substanz Ecdysterone im Mittelpunkt des
Interesses. Erythrina excelsa ist eine Pflanze die in der kamerunischen traditionellen
Medizin zur Behandlung von Wechseljahrsbeschwerden und gynäkologische
Tumoren Anwendung findet. Ob Erythrina excelsa allerdings wirklich
pharmakologisch aktive Substanzen enthält, die derartige Wirkungen erzielen
können, wurde bisher noch nicht untersucht. Ecdysterone (Ecdy) ist ein Inhaltsstoff
von verschiedenen Pflanzen, die in der chinesischen, ayurvedischen und russischen
traditionellen Medizin zur Behandlung unterschiedlichster Beschwerden verwendet
werden. Für Ecdy konnte eine anabole Wirkung in vitro und in vivo gezeigt werden.
Obwohl die zugrunde liegenden Mechanismen noch nicht aufgeklärt sind, wird es
intensiv als Nahrungsergänzungsmittel zur Steigerung des Muskelaufbaus beworben
und auch von Bodybuildern verwendet. Ein Missbrauch zum Doping in anderen
Sportarten wird befürchtet. Die Ziele dieser Arbeit waren zum einen die Bioaktivität
von Extrakten von Erythrina excelsa, die in Bezug zur traditionellen Anwendung in
der Naturmedizin stehen, sowie die biologisch aktiven Inhaltsstoffe in den Extrakten
näher zu charakterisieren. Zum anderen sollten die Mechanismen der anabolen
Wirkung von Ecdysterone untersucht werden.

Zusammenfassung
M.Sc. Sadrine Tchoukouegno Ngueu ix
Mittels Yeast Estrogen Screen (YES), uterotrophen Assay und des MTT-Assay
konnte gezeigt werden, dass ein Äthanolextrakt von E. excelsa östrogene/anti-
östrogene und zytotoxische Effekte aufweist. Eine dreitägige subkutane
Verabreichung des Extraktes in einer Dosis von 50 mg/kg KG/Tag an
ovariektomierten Wistar Ratten führte zu signifikanten östrogenen Wirkungen auf
morphometrische Parameter der Gebärmutter (Feuchtgewicht und epithelialen
Höhen) und auf die Expression von Östrogen-abhängigen Genen in der Gebärmutter
(C3 und Clusterin) und in der Leber (CaBP9K und IGFBP-1). Bei einer Dosis von 100
mg/kg KG/Tag zeigte der Extrakt unerwartet auf die beschriebenen biologischen
Endpunkte eine anti-östrogene Wirkung. Eine fünftägige Behandlung von
Estrogenrezeptor (ER) -positiven MCF-7 Brustkrebszellen mit dem Extrakt führte zu
einem biphasischen Profil: Bei niedrigen Konzentrationen kam es zu einem leichten
Anstieg der Zellviabilität, bei höheren Konzentrationen dagegen zu einer Induktion
von Zelltod. Dagegen wurde bei der Behandlung von ER negativen HT-29 Zellen
Zytotoxizität bei allen getesteten Konzentrationen beobachtet. Um die Wirkstoffe des
Extraktes näher zu charakterisieren, wurde eine Vakuum Flüssigchromatographie
Fraktionierung durchgeführt. Biologisch aktive Fraktionen wurden mittels des YES
Assays identifiziert und die biologische Wirkung dieser Fraktion auf Proliferation und
Apotose von MCF-7-Brustkrebs-Zellen mittels Durchflusszytometrie,
Genexpressionsanalyse und Immunhistochemie untersucht. Die Dichloromethan-
Methanol (50:50 v/v) Fraktion (F4), die in Bezug auf östrogene und zytotoxische
Wirkung die aktivste Fraktion war, zeigte eine nicht-monotone Konzentrations-
Wirkungs-Kurve auf den Zellzyklus, die Proliferation und Apoptose von MCF-7
Brustkrebszellen. Bei niedrigen Konzentrationen und in Abwesenheit von E2, förderte
F4 Östrogenrezenptor- abhängig die Proliferation von MCF-7 Zellen (Hochregulation
von Cyclin D1 und Herunterregulation des Bax/Bcl-2-mRNA Verhältnisses).
Interessanterweise wirkte F4 in den gleichen Konzentrationen bei Anwesenheit von
E2, als Antagonist und verhinderte ein E2-induziertes Zellwachstum. Bei höheren
Konzentrationen modulierte F4 den Zellzyklus zugunsten einer gesteigerten
Apoptose (Erhöhung des Caspase-gespaltenen Zytokeratin 18, Hochregulation des
Bax/Bcl-2 Verhältnisses und Herunterregulation von Cyclin-D1-mRNA). In
Zusammenarbeit mit der Arbeitsgruppe von Dr. Halabalaki von der Abteilung für
Pharmakognosie und Naturstoffchemie der Universität von Athen wurde eine weitere

Zusammenfassung
M.Sc. Sadrine Tchoukouegno Ngueu x
Charakterisierung des Extraktes mittels LC-HRMS und LC-HRMS/MS Analyse
durchgeführt um die im Extrakt enthaltenen Isoflavonoide zu identifizieren. Diese
Analyse zeigte, dass Pyran- und Prenyl-Isoflavonoidderivate die charakteristischen
chemischen Stoffe des Extrakts sind was auch die beobachtete Kombination der
biologischen Eigenschaften des Extraktes erklären könnte.
Im zweiten Teil dieser Arbeit wurden die anabolen Wirkmechanismen von
Ecdysterone, Erythrina excelsa Extrakt und Genistein in differenzierten C2C12
Myoblasten, einem Zellkulturmodell für Skelettmuskelhypertrophie untersucht. Als
untersuchter biologischer Endpunkt diente die Zunahme des Durchmessers von
C2C12 Myotuben. Für die nähere Charakterisierung der involvierten molekularen
Mechanismen wurden Studien mit Vergleichssubstanzen wie
Androgenrezeptoragonisten (DHT), Androgenrezeptorantagonisten (Flutamid),
anabol wirkende Peptidhormone (IGF-1) und verschiedene Östrogenrezeptor
bindende Substanzen mit agonistischen (E2), antagonistischen sowie ER alpha und
ER beta selektiven Bindungseigenschaften verwendet. Mit diesem Modell konnten
die wachstumsfördernden Wirkungen von IGF1, DHT, E2, Ecdy und des
Phytoestrogens Genistein auf die Skelettmuskulatur bestätigt werden. Co-
Behandlung von Myotuben mit Ecdy und ZK führte zu einer Hemmung der Ecdy-
induzierten Hypertrophie. Darüber hinaus führte auch eine Co-Behandlung von
Myotuben mit E2 oder Ecdy und dem selektiven ER beta Antagonisten zu einer
Antagonisierung der hypertrophen Wirkung von beiden Substanzen, was ein Zeichen
von ERβ-abhängigem Effekt ist.
Zusammenfassend zeigen die Befunde dieser Arbeit einerseits, dass der
Extrakt von Erythrina excelsa biologische Effekte aufweist, die in engem
Zusammenhang zu seiner traditionellen Verwendung stehen. Sie weisen auch darauf
hin, dass diese Pflanze eine Quelle pharmakologisch aktiver Stoffe ist, die von
potentiellem Interesse für weitere Untersuchungen sind. Darüber hinaus zeigen die
Ergebnisse dieser Arbeit, dass Stoffe mit ER beta Bindungseigenschaften das
Skelettmuskelwachstum fördern. Diese Erkenntnis ist von Interesse für die
Behandlung von atropischen Erkrankungen des Skelettmuskels wie z. B der
altersbedingten Abnahme der Muskelmasse (Sarkopenie). Allerdings kann dieser

Zusammenfassung
M.Sc. Sadrine Tchoukouegno Ngueu xi
Effekt auch zum Zwecke des Dopings missbraucht werden. Daher sind diese
Befunde auch für die Ausgestaltung von Anti-Doping-Bestimmungen von Bedeutung.

Summary
M.Sc. Sadrine Tchoukouegno Ngueu xii
Summary
Medicinal plants serve as primary health care in most Asian and African
countries. In developed countries where allopathic medicine is the most practiced
form of medicine, chemicals of plant origin are increasingly consumed either as
herbal formulations, diet, or complement to existing therapies in order to prevent or
treat diseases. There is a general belief that because of their natural origin, herbal
remedies are safe. Therefore they are commonly used in self therapeutic measures
and are not prescribed in Germany. This work deals with the bioactivity and potential
side effects of such herbal remedies. Namely the extract from Erythrina excelsa and
the pure compound Ecdysterone were the focus of our attention. Erythrina excelsa is
a plant used in the Cameroonian traditional medicine to alleviate menopausal
symptoms and to treat female gynecological tumors. However, no test for biological
activities evidencing these health benefit claims has been made so far. Ecdysterone
(Ecdy) is the principal constituent of several plants used in the Chinese, Ayurvedic,
and Russian traditional medicines to treat various ailments. Ecdy has been shown to
exert anabolic properties in vitro and in vivo. Although the mechanisms underlying
these effects are not yet elucidated, it is advertised as a food supplement to increase
muscle mass and physical performance and is used by bodybuilders. A misuse for
doping purpose in sports is suspected. The aims of this thesis were on the one hand
to investigate the biological activities likely to explain the traditional use of Erythrina
excelsa and to characterize its biologically active ingredients. On the other hand, this
work also aims at examining the mechanisms underlying the anabolic effect of
ecdysterone
Using the yeast estrogen screen, the uterotrophic assay, and the MTT assay,
we showed that an ethanol extract of E. excelsa exhibits estrogenic/anti-estrogenic
and cytotoxic effects. A three-day subcutaneous administration of the extract at a
dose of 50 mg/kg BW/day to ovariectomized Wistar rats produced significant
estrogenic effects on uterine morphometric parameters (wet weight and epithelial
height) and on the gene expression of estrogen responsive genes in the uterus (C3

Summary
M.Sc. Sadrine Tchoukouegno Ngueu xiii
and Clusterin) and in the liver (CaBP9K and IGFBP-1). Unexpectedly, at a dose of
100 mg/kg BW/day the extract significantly decreased the same parameters below
the control level indicating anti-estrogenic effects. The extract exhibited a biphasic
profile on the growth of ER positive MCF-7 cells with a slight increase of cell viability
at low concentrations and induction of cell death at higher concentrations. But on ER
negative HT-29, cytotoxicity was observed at all concentrations tested. To get more
insights into the active principles of the extract, a bioactivity-guided fractionation was
performed and the bioactive fractions were identified by means of both YES and XTT
assays. The effect of the most active fraction [Dichloromethane-Methanol (50:50, v/v)
(F4)] on the proliferation and apoptosis of MCF-7 breast cancer cell line was
assessed by flow cytometry, gene expression analysis and immunohistochemistry.
F4 exhibited a non-monotonic concentration-response effect on the proliferation and
apoptosis of MCF-7 breast cells. At low concentrations and in the absence of E2, F4
promoted cell cycle progression through ER signaling and modulated gene
expression in favor of cell survival (upregulation of cyclin D1 and and downregulation
of the Bax/Bcl-2 ratio mRNA). Interestingly, when E2 was present, F4 at the same
concentrations acted as an antagonist and prevented E2-induced cell growth. At
higher concentrations, F4 modulated the cell cycle in favor of increased apoptotic cell
death (increase of caspase cleaved cytokeratin 18, upregulation of Bax/Bcl-2 ratio
and downregulation of cyclin D1 mRNA). In collaboration with the group of Dr.
Halabalaki from the Department of Pharmacognosy and Natural Products Chemistry
of the Unviversity of Athens, a LC-HRMS and LC-HRMS/MS analysis was performed
to identify the isoflavonoids contained in the extract. This analysis showed that pyran
and prenyl derivatives of isoflavonoids are the most characteristic chemical groups of
the extract and may explain the observed biological activities.
In the second part of this work, the mechanisms of the anabolic effect of
ecdysterone, Erythrina excelsa extract and other plant secondary metabolites such
as genistein were studied in a cell culture model of differentiated C2C12 myoblasts.
This test system is a well established model to study the ability of anabolic
substances to induce skeletal muscle hypertrophy as measured by the increase in
the diameter of myotubes. Several comparative substances such as androgen
receptor agonist (DHT) and antagonist (flutamide), anabolic peptide hormone (IGF-1),

Summary
M.Sc. Sadrine Tchoukouegno Ngueu xiv
and different estrogen receptor binding compounds with agonistic (E2), antagonistic
(ZK), and ERα and ERβ selective binding properties were used for the
characterization of the involved molecular mechanisms. With this model, we
confirmed the skeletal muscle growth-promoting effects of IGF1, DHT, E2, Ecdy, and
the phytoestrogen genistein. Co-treatment of myotubes with ZK and Ecdy led to an
inhibition of Ecdy-induced hypertrophy. In addition, a co-treatment of myotubes with
E2 or Ecdy and the selective ERβ antagonist prevented both substances to induce
myotube hypertrophy, what unequivocally shows an ERβ-mediated effect.
In general, the results of this work show that the extract of Erythrina excelsa
has biological activities that are in close agreement with its reported traditional use to
manage menopausal complaints and to treat female gynecological tumors. They also
highlight the fact that this plant is a source of substances of potential interest for
further pharmacological investigations. In addition, our findings also show that
substances with ERβ binding affinity promote skeletal muscle growth and may
provide new insights into therapeutic strategies for the treatment of skeletal muscle
atrophic conditions such as the age-related decline in muscle mass (sarcopenia).
However, this effect may be misused for doping purpose and is therefore also of
importance for Anti-doping regulation.

Table of contents
M.Sc. Sadrine Tchoukouegno Ngueu xv
Table of contents
Acknowledgements ................................................................................................................. vi
Zusammenfassung ................................................................................................................. viii
Summary .................................................................................................................................. xii
Table of contents ..................................................................................................................... xv
List of figures .......................................................................................................................... xix
List of Tables ........................................................................................................................ xxiii
List of abbreviations ............................................................................................................ xxiv
CHAPTER 1: INTRODUCTION ................................................................................................. 1
1.1 Description of the research problem ............................................................................... 2
1.2 Aims and objectives of the thesis .................................................................................... 4
CHAPTER 2: LITTERATURE REVIEW .................................................................................... 6
Section 2.1: Estrogens ............................................................................................................ 7
2.1.1 Synthesis and endogenous sources ............................................................................ 7
2.1.2 Transport and metabolism ............................................................................................. 8
2.1.3 Mechanism of action ....................................................................................................... 9
2.1.3.1 Structure of estrogen receptors ......................................................................................... 9
2.1.3.2 Tissue distribution.............................................................................................................. 11
2.1.3.3 Mechanism of estrogen action (Figure 5) ...................................................................... 12
2.1.3.3.1 Genomic actions ......................................................................................................... 12
2.1.3.3.2 Non-genomic actions ................................................................................................. 14
2.1.4 Estrogens and health.................................................................................................... 15
2.1.4.1 Estrogens and hormone-dependent cancers ................................................................ 15
2.1.4.5 Estrogen and skeletal muscle .......................................................................................... 17

Table of contents
M.Sc. Sadrine Tchoukouegno Ngueu xvi
Section 2.2: Phytoestrogens ................................................................................................. 19
2.2.1 Definition and sources ................................................................................................. 19
2.2.2 Absorption and metabolism ........................................................................................ 20
2.2.3 Mode and mechanisms of action ................................................................................ 21
2.2.4 Phytoestrogens and health .......................................................................................... 22
2.2.4.1 Phytoestrogens and the reproductive health ................................................................. 22
2.2.4.2 Phytoestrogens and gynecological cancers .................................................................. 22
2.2.4.3 Phytoestrogens and menopausal symptoms ................................................................ 24
Section 2.3: Erythrina excelsa Baker ................................................................................... 26
2.3.1 Taxonomy ...................................................................................................................... 26
2.3.2 Description and geographical distribution ................................................................ 27
2.3.3 Active constituents of plants of the genus Erythrina ............................................... 27
2.3.4 Traditional uses of Erythrina excelsa Baker .............................................................. 27
Section 2.4: Ecdysterone ...................................................................................................... 29
2.4.1 Origin and sources ....................................................................................................... 29
2.4.2 Biological effects .......................................................................................................... 30
Section 2.5: Marker genes used ........................................................................................... 33
2.5.1 Complement component 3 (C3) ................................................................................... 33
2.5.2 Clusterin ......................................................................................................................... 33
2.5.3 Insulin-like growth factor binding protein 1 (IGFBP-1)............................................. 33
2.5.5 Cyclin D1 (CCND1) ........................................................................................................ 34
2.5.6 Bax .................................................................................................................................. 34
2.5.7 Bcl-2 ................................................................................................................................ 35
CHAPTER 3: MATERIAL AND METHODS ............................................................................ 36
3.1 Material .............................................................................................................................. 37
3.1.1 Substances ............................................................................................................................ 37
3.1.2 Chemicals, consumables and lab equipments ................................................................. 39

Table of contents
M.Sc. Sadrine Tchoukouegno Ngueu xvii
3.1.3 Test organisms ...................................................................................................................... 44
3.2 Methods ............................................................................................................................. 45
3.2.1 Preparation of cell culture media and buffers ................................................................... 45
3.2.3 LC-HRMS and LC-HRMS/MS analysis ............................................................................. 49
3.2.4 Vacuum liquid chromatography (VLC)............................................................................... 50
3.2.5 HPLC ...................................................................................................................................... 50
3.2.6 Yeast estrogen screen ......................................................................................................... 51
3.2.7 MTT/XTT assays ................................................................................................................... 54
3.2.8 Myotubes diameter assay .................................................................................................... 54
3.2.9 Western blot ........................................................................................................................... 56
3.2.10 Uterotrophic assay.............................................................................................................. 57
3.2.11 Histological analysis of tissues ......................................................................................... 58
3.2.12 Gene expression analysis ................................................................................................. 60
3.2.13 Cell cycle analysis .............................................................................................................. 63
3.2.14 Immunocytochemistry ........................................................................................................ 65
3.2.15 Statistical analysis .............................................................................................................. 66
CHAPTER 4: RESULTS .......................................................................................................... 67
4.1 Biological activities and chemical characterization of Erythrina excelsa ................ 68
4.1.1 Estrogenic and Cytotoxic properties of E.excela whole extract ............................ 68
4.1.1.1 Yeast estrogen screen ...................................................................................................... 68
4.1.1.2 Uterotrophic assay ............................................................................................................ 70
4.1.1.2.1 Effect of E. excelsa whole extract on the uterus ................................................... 70
4.1.1.2.2 Effect of E. excelsa whole extract on the vagina ................................................... 72
4.1.1.2.3 Effect of E. excelsa whole extract on the liver IGFBP-1 and CaBP9k mRNA
expression .................................................................................................................................... 73
4.1.1.3 MTT assay .......................................................................................................................... 74
4.1.2 Effect of a phytoestrogenic fraction of E. excelsa on the growth of the human
breast cancer line MCF-7. ...................................................................................................... 75

Table of contents
M.Sc. Sadrine Tchoukouegno Ngueu xviii
4.1.2.1 Yeast estrogen screen ...................................................................................................... 75
4.1.2.2 XTT ...................................................................................................................................... 76
4.1.2.3 HPLC fingerprints of F4 and E. excelsa whole extract ................................................ 78
4.1.2.4 Effect of F4 on MCF-7 cell cycle distribution ................................................................. 78
4.1.2.4.1 Antagonisation with ZK .............................................................................................. 80
4.1.2.4.2 Cotreatment with E2 .................................................................................................. 81
4.1.2.5 M30 staining ....................................................................................................................... 83
4.1.2.6 Effect of F4 on the mRNA expression of CCND1, Bax and Bcl-2 in MCF-7 cells ... 83
4.1.3 Chemical characterisation of E. excelsa extract ....................................................... 84
4.2 Effects of estrogens and chemicals of plant origin on skeletal muscle growth ...... 92
4.2.1 E2, genistein, and Ecdysone induce hypertrophy of C2C12 murine skeletal muscle
cells ................................................................................................................................................... 92
4.2.2 Role of ER and AR on ecdysone induced myotube hypertrophy .................................. 92
4.2.3 Role of ERα and ERβ in skeletal muscle growth ............................................................. 93
CHAPTER 5: DISCUSSION .................................................................................................... 98
5.1 Biological activities of Erythrina excelsa ...................................................................... 99
5.1.1 Estrogenic/anti-estrogenic and cytotoxic properties of E. excelsa ................................ 99
5.1.2 Effect of a phytoestrogenic faction of E. excelsa on the growth of the human breast
cancer line MCF-7. ....................................................................................................................... 103
5.2 Effect of estrogens and chemicals of plant origin on C2C12 murine skeletal muscle
cells growth. ...........................................................................................................................105
CHAPTER 6: CONCLUSION AND PERSPECTIVES ...........................................................108
Bibliography ...........................................................................................................................111
Publications and presentations ...........................................................................................141
Curriculum vitae ....................................................................................................................142

List of figures
M.Sc. Sadrine Tchoukouegno Ngueu xix
List of figures
Figure 1: Source of drugs (Adapted from (Cragg and Newman, 2013)). .................................... 3
Figure 2: Biosynthesis of estrogens in ovaries (Campbell reproductive biology July 2012:
http://kcampbell.bio.umb.edu). SCC=P450scc= cholesterol side-chain cleavage enzyme; 3β-
HSD= 3β-hydroxysteroid dehydrogenase. The conversion of androstenedione to testosterone
is mediated by the enzyme 17β- hydroxysteroid dehydrogenase (17β-HSD). ........................... 8
Figure 3: (A) Modular structure of nuclear receptors (Adapted from (Nilsson et al., 2001) and
(B) protein sequence homology of the two ERs (Adapted from (Shao and Brown, 2004);
percentages of homology are from Faulds et al. (2012). .............................................................. 10
Figure 4: ERs distribution in male and female organs (Nilsson and Gustafsson, 2011) ......... 11
Figure 5: Molecular pathways of ERs actions (Heldring et al., 2007). TF= transcription factor;
SM= second messenger. ................................................................................................................... 14
Figure 6: Mechanisms of estrogens-induced cancer .................................................................... 17
Figure 7: Major classes of phytoestrogens ..................................................................................... 20
Figure 8: Overview of possible actions of phytoestrogens in relation to cancer (adapted from
(Mense et al., 2008)). ......................................................................................................................... 24
Figure 9: Erythrina Excelsa (April 2011 in Kumba-Cameroon by Stephane Zingue) ............... 26
Figure 10: 20-hydroxyecdysone (20 HE) or Ecdysterone ............................................................ 29
Figure 11: Pathways of Ecdysterone-induced protein synthesis in skeletal muscle (Gorelick-
Feldman et al., 2010). 20HE: 20-hydroxyescdysone; PIP2: phosphatidylinositol-4,5-
diphosphate; PLC: phospholipase C; IP3: Inositol-1,4,5-triphosphate; IP3R: IP3 receptor; SR:
sarcoplasmic reticulum. ..................................................................................................................... 31
Figure 12: Estrogen-inducible expression in yeast (Routledge and Sumpter, 1996). .............. 52
Figure13: Cell culture procedure for cell cycle analysis. .............................................................. 64
Figure 14: Effect of E. excelsa whole extract as compared to genistein and E2 on the
expression of β-galactosidase in yeast cells. The graph depicts the concentration of 17β-
estradiol, genistein, ZK and E. excelsa ethanolic extract plotted against the CAUs of the
medium after 3 days of incubation (A) estrogenicity; (B) antagonism of E. excelsa with ZK
and (C) anti-estrogenicity. ................................................................................................................. 69
Figure 15: (A) Uterine wet weight (mg/kg BW) and (B) uterine epithelial height (µm) after 3-
day subcutaneous treatment of ovariectomized Wistar rats with either E. excelsa ethanolic

List of figures
M.Sc. Sadrine Tchoukouegno Ngueu xx
extract (50 and 100 mg/kg BW/day) or E2 (4 µg/kg/BW/day). (C) 100X Microphotographs of
uterine tissue following a Hematoxylin-Eosin (HE) staining. Mean values ± S.D. are given for
n = 6 animals per dose group ** p≤ 0.01;*p≤0.05, significantly different from the control group
(Ovex), Kruskal-Wallis H-test followed by Mann–Whitney U-test. .............................................. 71
Figure 16: (A) Relative mRNA expression of C3 and (B) clusterin (normalized to the mRNA
amount of 1A and compared to the vehicle control group) in the uterus of ovariectomized
Wistar rats treated subcutaneously for 3 consecutive days with 50 and 100 mg/kg BW/day E.
excelsa ethanolic extract and 4 µg/kg/BW/day E2. * P≤0.05 significantly different from the
control group (Ovex), Kruskal-Wallis H-test followed by Mann-Withney U-test. ....................... 72
Figure 17: (A) Vaginal epithelial height (µm) after 3-day subcutaneous treatment of
ovariectomized Wistar rats with either E. excelsa ethanolic extract (50 and 100 mg/kg
BW/day) or E2 (4 µg/kg/BW/day). (B) 100X Microphotographs of vagina tissue following a
Hematoxylin-Eosin (HE) staining. Mean values ± S.D. are given for n = 6 animals per dose
group ** p≤ 0.01;*p≤0.05, significantly different from the control group (Ovex), Kruskal-Wallis
H-test followed by Mann–Whitney U-test. ....................................................................................... 73
Figure 18: Relative mRNA expression of (A) IGFBP-1 and (B) CaBP9k (normalized to the
mRNA amount of 1A and compared to the vehicle control group) in the liver of
ovariectomized Wistar rats after 3-day subcutaneous administration of 50 and 100 mg/kg
BW/day E. excelsa ethanolic extract and 4 µg/kg/BW/day E2. *P≤0.05, Kruskal-Wallis H-test
followed by Mann-Withney U-test. ................................................................................................... 74
Figure 19: Effects of E. excelsa whole extract on the growth of (A) MCF-7 and (B) HT-29
cells. Results present the average and standard deviation of n≥3 independent experiments
performed in 6 replicates for each concentration. ......................................................................... 75
Figure 20: Effect of fractions of E. excelsa on the expression of β-galactosidase in yeast
cells. (A) VLC fractions, (B) active fractions compared to the whole extract. The graphs depict
the concentration of test chemicals plotted against the CAUs of the medium after 3 days of
incubation. ............................................................................................................................................ 76
Figure 21: (A) Effect of estrogenic fractions of E. excelsa ethanol extract on MCF-7 cell„s
viability. (B) F4 compared to the whole extract. MCF-7 cells were incubated for 72 h with
increasing concentrations of fractions or whole extract and the cell viability examined by
XTT.* (p≤0.05) and *** (p<0.001) vs. control analyzed by Kruskal Wallis H-test followed by
Mann Whitney U-test. ......................................................................................................................... 77
Figure 22: HPLC chromatograms of E. excelsa whole extract and F4 fraction ........................ 78
Figure 23: Effects of F4 on the cycle distribution of MCF-7 cells. MCF-7 cells were incubated
for 48 h with increasing concentrations of F4 and the cell cycle distribution assessed by flow
cytometry. (A) Representative Histogramms and (B) mean of n≥ 3 independent experiments.
a (p≤0.05), b (p<0.01), c (p<0.001) vs. control analyzed by Kruskal Wallis H-test followed by
Mann Whitney U-test. ......................................................................................................................... 79
Figure 24: Role of ER in F4-mediated effects on the cycle distribution of MCF-cells. MCF-7
cells were cotreated for 48 h with F4 and the pure anti-estrogen ZK and the cell cycle

List of figures
M.Sc. Sadrine Tchoukouegno Ngueu xxi
analyzed by flow cytometry. * (p≤0.05), ** (p<0.01), *** (p<0.001) vs. Control; + (p≤0.05), ++
(p<0.01), +++ (p<0.001) vs. single treatment analyzed by Kruskal Wallis H-test followed by
Mann Whitney U-test. ......................................................................................................................... 80
Figure 25: Effect of the cotreatment with E2 and lower (A) and higher (B) concentrations of
F4 on the cycle distribution of MCF-7 cells. MCF-7 cells were treated for 48h with various
concentrations of F4 together with 0.1 nM E2 and the cell cycle distribution analyzed by flow
cytometry. * (p≤0.05), ** (p<0.01), *** (p<0.001) vs. Control; + (p≤0.05), ++ (p<0.01), +++
(p<0.001) vs. single treatment analyzed by Kruskal Wallis H-test followed by Mann Whitney
U-test. ................................................................................................................................................... 82
Figure 26: Effect of F4 on caspase-cleaved cytokeratin 18 in MCF-7 cells. MCF-cells were
exposed for 6h to either ZK (10-6
M) or increasing concentrations of F4 and the protein
expression of caspase-cleaved cytokeratin 18 measured by indirect immunofluorescence
using the M30 CytoDEATH antibody as described in material and methods. (A)
Representative fluorescence microscopy images (100 X magnitude), (B) quantitative analysis
of M30 positive cells. .......................................................................................................................... 83
Figure 27: Effects of F4 on the mRNA expression of Bax (A), Bcl-2 (B), Bax/Bcl-2 ratio (C),
and Cyclin D1 (D) in MCF-7 cells. MCF-7 cells were treated for 48h with the tests chemicals
and the expression level of target genes assessed by real-time RT-PCR. ** (p<0.01)
analyzed by Kruskal-Wallis H-test followed by Mann Whitney U-test. ....................................... 84
Figure 28: Base Peak chromatogram of Erythrina excelsa ethanol extract and Extracted Ion
Chromatograms (XICs) of representative isoflavonoids detected............................................... 85
Figure 29: HRMS/MS spectrum of compound 1. ........................................................................... 87
Figure 30: HRMS/MS spectrum of compound 3. ........................................................................... 88
Figure 31: HRMS/MS spectrum of compound 5. ........................................................................... 89
Figure 32: HRMS/MS spectrum of compound 9. ........................................................................... 89
Figure 33: HRMS/MS spectrum of compound 4. ........................................................................... 90
Figure 34: HRMS/MS spectrum of compound 6. ........................................................................... 90
Figure 35: HRMS/MS spectrum of compound 7. ........................................................................... 91
Figure 36: HRMS/MS spectrum of compound 8. ........................................................................... 91
Figure 37: Effect of E2, PEs, and the phytoecdysteroid ecdysone on C2C12 myotubes
hypertrophy. 4-6 days old myotubes incubated for 48 h with test chemicals were fixed and
photographied by glutaraldehyde induced autofluorescence. Depicted are mean of n≥2
independent experiments. * p≤0.05 vs. Control by Kruskal Wallis H-test and Mann Withney
U-test. ................................................................................................................................................... 92
Figure 38: Role of AR (A) and ER (B) on ecdysterone induced C2C12 myotubes hypertrophy.
4-6 days old myotubes incubated for 48 h with test chemicals were fixed and photographied

List of figures
M.Sc. Sadrine Tchoukouegno Ngueu xxii
by glutaraldehyde induced autofluorescence. Represented are mean± SD of n=2
independent experiments. * p≤0.05 vs. Control, + p≤0.05 vs. Single treated Kruskal Wallis H-
test and Mann Withney U-test. ......................................................................................................... 93
Figure 39: Effect of ER subtype selective agonists on C2C12 myotubes hypertrophy. 4-6
days old myotubes incubated for 48 h with test chemicals were fixed and photographied by
glutaraldehyde induced autofluorescence. Depicted are mean of n=3 independent
experiments. ........................................................................................................................................ 94
Figure 40: Effect of ER subtype selective agonists on C2C12 myotubes hypertrophy. 4-6
days old myotubes incubated for 48 h with test chemicals were fixed and photographied by
glutaraldehyde induced autofluorescence. Depicted are mean of n=3 independent
experiments. * p≤0.05 vs. Control, + p≤0.05 vs. Single treated Kruskal Wallis H-test and
Mann Withney U-test. ......................................................................................................................... 95
Figure 41: Ecdysterone induced myotube hypertrophy is ERβ-mediated. 4-6 days old
myotubes incubated for 48 h with test chemicals were fixed and photographied by
glutaraldehyde induced autofluorescence. * p≤0.05 vs. Control Kruskal Wallis H-test and
Mann Withney U-test. ......................................................................................................................... 96
Figure 42: ERα mRNA expression (A) and ERβ protein expression (B) in C2C12 myotubes.97
Figure 43: Transactivation studies with ERα agonist (16α-LE2) and ERβ agonist (8β-VE2) in
comparison to E2 (Hillisch et al., 2004) ......................................................................................... 107

List of tables
M.Sc. Sadrine Tchoukouegno Ngueu xxiii
List of Tables
Table 1: List of chemicals .................................................................................................................. 39
Table 2: List of antibodies ................................................................................................................ 42
Table 3: Consumables and equipement. ........................................................................................ 42
Table 4: Yeast culture medium components .................................................................................. 45
Table 5: C2C12, HT-29 and MCF-7 cells growth media ............................................................... 46
Table 6: VLC fractions. ..................................................................................................................... 50
Table 7: Experimental groups myotube diameter assay. ............................................................. 56
Table 8: Groups treatment uterotrophic assay ............................................................................... 58
Table 9: Primer Sequences ............................................................................................................... 61
Table 10: PCR reaction mix. ............................................................................................................. 62
Table 11: PCR cycling conditions. .................................................................................................. 62
Table 12: Experimental groups for the cell cycle analysis. .......................................................... 63
Table 13: Yeast assay results with E. excelsa whole extract. ..................................................... 69
Table 14: Yeast assay results with VLC fractions. ........................................................................ 76
Table 15: Representative Isoflavonoids of Erythrina excelsa ethanol extract using LC-PDA-
ESI (+)-LTQ-Orbitrap platform. ......................................................................................................... 86

List of abbreviations
M.Sc. Sadrine Tchoukouegno Ngueu xxiv
List of abbreviations
ACN Acetonitrile
AF-1 Activation function 1
AF-2 Activation function 2
ALPHA ERα-selective agonist (16α-LE2)
AP-1 Activator protein 1
ApoJ Apolipoprotein J
AR Androgen receptor
ATCC American Type Culture Collection
BAX BCL-2 associated protein X
BCL-2 B-cell lymphoma 2
DBD DNA-binding domain
BETA ERβ-selective Agonist (8β-VE2)
BSA Bovine serum albumin
BW Body weight
CAM Complementary and alternative medicine
CAU Corrected absorbance units
C3 Complement component 3
CaBP9k Vitamin D dependent calcium binding protein 9kDa
cAMP Cylic adenosine monophosphate
CCND1 Cyclin D1
CDNA complementary desoxyribonucleic acid
CID Collision-induced dissociation
CPRG Chlorophenol red-β-D-galactopyranoside
DAD Diode-array detector
DAPI 4',6-diamidino-2-phenylindole
DCM Dichloromethane
ddH2O Double distilled water
DEPC Diethyl pyrocarbonate
DHT Dihydrotestosterone
DM Differentiation medium
DMEM Dubelcco's modified eagle medium
DMSO dimethylsulfoxide
DNA Desoxyribonucleic acid
dNTPs Deoxynucleotide triphosphates
E1 Estrone
E2 17β)-estra-1,3,5(10)-triene-3,17-diol, 17β-estradiol
E3 16α-Hydroxy-17β-estradiol, Estriol
Ecdy 20-Hydroxyecdysone, Ecdysterone

List of abbreviations
M.Sc. Sadrine Tchoukouegno Ngueu xxv
EDL Extensor digotorum longus
EDTA Ethylenediaminetetraacetic acid
EGF Epidermal growth factor
EICCAM European information centre for alternative and complementary medicine
ER Estrogen receptor
ERE Estrogen responsive elements
ERα Estrogen receptor alpha
ERβ Estrogen receptor beta
ESI Electrospray ionization
EtOH Ethanol
EU European Union
FACS Fluorescence activated cell sorting
FBS Foetal bovine sserum
Fe2(S04)3 Iron(III) sulfate hydrate, ferric sulfate hydrate
FMD Flow mediated vasodilatation
FSH Follicle-stimulating hormone
GnRH gonadotropin releasing hormone
GPCR G protein-coupled receptor
HCl Hydrogen chloride
HDL high density lipoprotein
hERα Human ERα
HPLC High performance liquid chromatography
IGF1 Insulin-like growth factor 1
IGFBP-1 Insulin-like growth factor binding protein 1
IGF-II Insulin-like growth factor II
ISO Isoflavones
KH2PO4 Potassium dihydrogen phosphate
KOH Potassium hydroxide
LBD ligand binding domain
LC-HRMS Liquid chromatography-high resolution mass spectrometry
LDL Low density lipoprotein
LH Luteinizing Hormone
MAPK mitogen-activated protein kinase
MCF-7 Michigan Cancer Foundation - 7
MeOH Methanol
MgCl2 Magnesium chloride
MgSO4 Magnesium sulfate
mRNA Messenger ribonucleic acid
MS Mass spectrometry
MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)
MW Molecular weight
Na2HPO4 *2H2O Disodium phosphate dihydrate
NaCl Sodium chloride

List of abbreviations
M.Sc. Sadrine Tchoukouegno Ngueu xxvi
NaH2PO4* H2O Disodium phosphate hydrate
NaOH Sodium hydroxide
NF-κB Nuclear factor-kappa B
(NH4)2S04 Ammonium sulfate
NTD N-terminal domin
PBS Phosphate buffered saline
PCR Polymerase chain reaction
PE Phytoestrogen
PI Propidium iodide
PI3k Phosphatidylinositide 3-kinase
PKA Protein kinase A
PKC Protein kinase c
PM Proliferation medium
PMSF Phenylmethylsulfonyl fluoride
Rb Retinoblastoma protein
RCT Randomized controlled trial
RNA Ribonucleic acid
RPMI Roswell Park Memorial Institute medium
RT-qPCR Real time quantitative polymerase chain reaction
SCC Side-chain cleavage
SDS sodium dodecyl sulfate
SERMS Selective estrogen recptor modulators
SGP-2 Sulphated glycoprotein 2
SHBG Sex-hormone binding globulin
SR Sarcoplasmic reticulum
SP-1 Specificity protein 1
TBS Tris buffered saline
TGF-α Transforming growth factor alpha
TM Traditional medicine
TRIS Tris (hydroxymethyl) aminoethane
TRPM-2 Testosterone-repressed protein message 2
VLC Vacumm liquid chromatography
WHO World health organization
XTT 2,3-Bis-(2-Methoxy-4-Nitro-5-Sulfophenyl)-2H-Tetrazolium-5-Carboxanilide
YES yeast estrogen screen
YES yeast estrogen screen

Introduction
M.Sc. Sadrine Tchoukouegno Ngueu 1
CHAPTER 1: INTRODUCTION

Introduction
M.Sc. Sadrine Tchoukouegno Ngueu 2
1.1 Description of the research problem
Most African and Asian countries have a long history of use of traditional
medicine (TM) for health care. A World Health Organization (WHO) report shows that
40-80% of the population in these countries rely on TM for primary health care
(WHO, 2008). Traditional medicine is defined by the WHO as “diverse health
practices, approaches, knowledge and beliefs incorporating plant, animal, and/or
mineral based medicines, spiritual therapies, manual techniques and exercises
applied singularly or in combination to maintain well-being, as well as to treat,
diagnose or prevent illness“. In developed countries, TM is referred to as
complementary and alternative medicine (CAM), non-conventional or parallel
medicines. The use of TM/CAM has increased worldwide over the last decades.
Surveys done in recent decades show that 20% of the EU-citizens have a clear
preference for CAM healthcare, another 20% are regular users of CAM and more
than 100 million citizens in the EU make use of CAM (EICCAM, 2009). The
increasing use of TM/CAM raises some challenging issues concerning evidences on
their quality, safety, and efficiency. The strategy of the WHO is to integrate TM/CAM
into national health care systems. For this to occur, sufficient knowledge should be
available on their efficiency or effectiveness and safety allowing the identification of
safest and most effective therapies. TM also plays an important role in guiding drug
discovery and development. Recent studies (Newman and Cragg, 2010; Cragg and
Newman, 2013) analyzed the sources of new drugs over the period 01/1981-12/2010
and reported that only 36% of 1073 new chemical compounds considered were
devoid of natural inspiration as represented in Figure 1.

Introduction
M.Sc. Sadrine Tchoukouegno Ngueu 3
Figure 1: Source of drugs (Adapted from (Cragg and Newman, 2013)).
N: unmodified natural product; ND: Modified natural product; S: synthetic compound with no
natural product conception; S*, S*/NM: Synthetic compound with a natural product
pharmacophore, N/M indicating competitive inhibition; S/NM: synthetic compound showing
competitive inhibition of the natural product substrate.
Herbal medicine is one of the most used form of alternative therapy and is
used either to prevent or to treat a wide range of health conditions (menopausal
symtoms, fatigue, cardiovascular diseases, diabetes, cancer, mental disorder…).
Erythrina excelsa Baker (Fabaceae/Leguminoseae) extract and Ecdysterone
containing plants extracts such as Rhaponticum carthamoides, Achyranthes bidenta,
and Trianthema portulacastrum are examples of herbal preparations that are used in
several tradiotional medicine systems. E. excelsa is a plant belonging to the genus
Erythrina that is known for its traditional use by several communities to treat a wide
range of ailments (reviewed in Majinda et al., 2005). E. excelsa stem bark is reported
to have been used by some village communities in the SouthWest region of
Cameroon to alleviate menopausal complaints and to treat growths (uterine fibroids
and ovarian cysts being the most frequent) in the female genital tract. Uterine fibroids
are begnin growths that develop from the muscle tissue of the uterus. Normally they
are not harmful but may under certain circumstances negatively affect fertility

Introduction
M.Sc. Sadrine Tchoukouegno Ngueu 4
outcomes (Pritts et al., 2009). It is recognized that 4-25 % of white women over 30
years old are carriers of uterine fibroids, while this frequency increases to 50% in
African women (Lefebvre, 1993). This shows that the use of this medicinal plant may
include a population of both pre- and postmenopausal women. Like most herbal
preparations used in the traditional medicine, testing for biological activity evidencing
these health benefit claims has not been performed. Ecdysterone (Ecdy) also known
as ecdisten, ecdysone, isoinokosterone, 20-hydroxyecdysone or β-ecdysterone
belongs to the class of ecdysteroids that are steroids mainly found in Arthropods but
that are also present in large amounts in certain plants including spinach (Spinacia
oleracea). In vitro and in vivo studies have reported anabolic effects of Ecdy
(Chermnykh et al., 1988; Gorelick-Feldman et al., 2008; Gorelick-Feldman et al.,
2010). However, despite these findings, the mechanisms by which Ecdy may exert
this effect are not yet elucided. As a consequence of this lack of evidence, Ecdy is
not subjected to regulation and is incorporated in many commercially available
supplements for body-builders and sportsmen and may be misused for doping
purpose.
1.2 Aims and objectives of the thesis
The aims and associated objectives of this thesis are :
Aim 1: To evaluate the biological activities of E. excelsa in relation to its traditionally
reported use for the management of menopausal complaints and for the treatment of
female gynecological tumors.
To evaluate the estrogenic activity of an ethanol extract of E.excelsa;
To investigate the cytotoxic effect of E.excelsa ethanol extract.

Introduction
M.Sc. Sadrine Tchoukouegno Ngueu 5
Aim 2: To get more insights into the “active principles” of E. excelsa
To get a fraction containing the secondary metabolites accounting for the
biological activities of interest (estrogenic and cytotoxic effects);
To evaluate the effect of this fraction on the growth of MCF-7 breast cancer
cell line;
To identify the main chemical components of the extract.
Aim 3: To elucidate the mechanisms by which the phytoecdysteroid Ecdysterone
affect mammalian skeletal muscle homeostasis.
To evaluate the ability of Ecdysterone and other chamicals of plant origin to
induce skeletal muscle hypertrophy;
To characterize the mechanisms invoved in the anabolic activity of
Ecdysterone.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 6
CHAPTER 2: LITTERATURE REVIEW

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 7
Section 2.1: Estrogens
There are three major naturally occurring estrogens (structures see below) in
women. 17β-estradiol (E2) is the most potent and main estrogen found in
premenopausal and non-pregnant women. After menopause estrone (E1) is
predominant. Estriol (E3) is known as the pregnancy hormone.
2.1.1 Synthesis and endogenous sources
In premenopausal women, the ovaries are the primary site of production and
source of circulating E2. The steroidogenic cells of the ovary are the granulosa cells
that are the cellular compartment surrounding the oocyte and the theca cells that
reside in the ovarian stroma (Havelock et al., 2004). The biosynthesis of estrogens in
the ovaries is under the influence of the hypothalamic-pituitary axis and consists of
the production of androgens from cholesterol in theca cells, followed by their
conversion to estrogens in granulosa cells (Figure 2). The primary source of estrogen
in postmenopausal women is from the conversion of androstenedione to estrone in
adipose tissue. The placenta is the major source of estrogens during pregnancy.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 8
Figure 2: Biosynthesis of estrogens in ovaries (Campbell reproductive biology July 2012:
http://kcampbell.bio.umb.edu). SCC=P450scc= cholesterol side-chain cleavage enzyme; 3β-
HSD= 3β-hydroxysteroid dehydrogenase. The conversion of androstenedione to testosterone
is mediated by the enzyme 17β- hydroxysteroid dehydrogenase (17β-HSD).
2.1.2 Transport and metabolism
In the circulation, most of the estrogen is reversibly bound to and transported
by the sex hormone-binding globulin (SHBG). SHBG is a protein mainly secreted by
the liver and it influences the action of the steroids in target tissue by regulating their
bioavailability. Estrogens can also bind to albumin but with less affinity as compared
to SHBG (17alpha-ethinylestradiol, the E2 derivate used for hormonal contraception,
is almost exclusively bound to albumin). A small amount of estrogens circulate bound
to HDL and LDL (Shwaery et al., 1997; Tang et al., 1997).
Estrogens are mainly metabolized in the liver and the gastro-intestinal tract.
They are conjugated in the liver as glucuronides, sulfates and thioether (Fishman et
al., 1960; Tseng et al., 1972). The conjugated forms of estrogens are water soluble
and are readily excreted via different routes (Eriksson and Gustafsson, 1971). The
glucuronide conjugates are more rapidly excreted in urine than the sulfates that have

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 9
a higher chance to be hydrolyzed in tissues to biologically active estrogens
(Sandberg and Slaunwhite, 1957). The estrogen metabolites that are excreted in the
bile pass into the intestine, are hydrolyzed by intestinal bacteria (Adlercreutz et al.,
1976), and are either reabsorbed into the portal circulation or excreted in the feces.
2.1.3 Mechanism of action
2.1.3.1 Structure of estrogen receptors
The biological effects of estrogens are mediated through two distinct estrogen
receptors (ER) subtypes; ERα and ERβ that are members of the large superfamily of
nuclear receptors. Like all other members of this family, ERs are composed of five
independent but interacting functional domains (A/B-F). These domains are the N-
terminal domain (NTD or A/B), the DNA-binding domain (DBD or C), the hinge or D
domain, the ligand-binding domain (LBD or E), and the F domain (Figure 3).
The NTD or A/B encodes the transcriptional activation function (AF-1) (Kraus
et al., 1995; McInerney and Katzenellenbogen, 1996) that is a region involved in the
interaction with coregulator proteins. It can also be phosphorylated and thus is
important for autonomous ligand-independent activation of transcriptional activity of
the ER (Onate et al., 1998; Webb et al., 1998). This domain also comprises amino
acids that are targets for post-translational modifications by kinases of growth factor
pathways that stimulate the activation function 1 (AF-1) activity (Nilsson and
Gustafsson, 2011)
The DBD contains a zinc finger structure that plays an important role in the
binding of the receptor to specific DNA sequences called estrogen response
elements (ERE) (Beato, 1989; Schwabe et al., 1993).
The D domain contains amino acid sequences that promote nuclear
localization and those that are targets for post-translational modifications that affect
ER activity and degradation (Nilsson and Gustafsson, 2011). It also serves as a
flexible region connecting DBD and LBD (Kumar et al., 2011).
The COOH-terminal or ligand-binding domain (LBD) is involved in ligand
binding, receptor dimerization, and contains the ligand-dependent activation function
2 (AF-2) that mediates the interaction of ERs with coregulator proteins and the

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 10
transactivation of target gene expression (Giguere et al., 1988; Danielian et al., 1992;
Tsai and O'Malley, 1994; Henttu et al., 1997; Darimont et al., 1998; Feng et al., 1998;
Shiau et al., 1998).
The role of the F-domain in ER activity is not well defined, but some data
suggest that this domain may affect the agonist/antagonist activity of selective ER
modulators (SERMs), ER dimerization, and ER-coregulator interactions (Nilsson and
Gustafsson, 2011).
The two ERs share a quite high degree of sequence homology in their DBD
(96% amino acid identity) and LBD (59% amino acid identity) (Nilsson and
Gustafsson, 2011). However, the NTD of ERβ is shorter than that of ERα with a very
poor sequence homology (Kumar et al., 2011). Moreover, the AF-1 domain of ERα is
very active in the stimulation of reporter-gene expression from a variety of estrogen
response element (ERE)-reporter constructs in different cell lines; while the
transcriptional activity of the AF-1 of ERβ under the same conditions is negligible
(Cowley and Parker, 1999).
Figure 3: (A) Modular structure of nuclear receptors (Adapted from (Nilsson et al., 2001) and
(B) protein sequence homology of the two ERs (Adapted from (Shao and Brown, 2004);
percentages of homology are from Faulds et al. (2012).
H2
N-
-COOH
< 20% 95% 30%
59%
< 20%
A)
B)

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 11
2.1.3.2 Tissue distribution
The two ER subtypes are expressed in both male and female reproductive and
non-reproductive organs but show some differences in their tissue distribution (Figure
4). In some organs, ERα and ERβ are co-expressed at similar levels, whereas in
other tissues, one or the other subtype predominates (Nilsson and Gustafsson,
2011). ERα is predominantly expressed in the uterus, prostate (stroma), ovary (theca
cells), testes (Leydig cells), epididymis, bone, breast, liver, kidney, white adipose
tissue, and various regions of the brain. ERβ is predominantly expressed in the
intestine, prostate (epithelium), testis, ovary (granulosa cells), bone marrow, salivary
gland, vascular endothelium, lung, bladder and certain regions of the brain (Ascenzi
et al., 2006; Nilsson and Gustafsson, 2010).
Figure 4: ERs distribution in male and female organs (Nilsson and Gustafsson, 2011)

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 12
2.1.3.3 Mechanism of estrogen action (Figure 5)
Most of the effects of estrogens in target organs are exerted through the action
of ERs on the gene expression (genomic actions), but a number of other effects are
so rapid (within seconds or minutes) that they cannot depend on DNA transcription
and protein synthesis (non-genomic actions).
2.1.3.3.1 Genomic actions
Ligand-dependent mechanism
ERE-dependent pathway
The ERE-dependent pathway is the classic/main mechanism through which
the ERs modulate expression of target genes. In the absence of ligands, ERs are
largely located in the cytosol where they are associated with intracytoplasmic
chaperones such as heat-shock protein 70 and 90 (Smith et al., 1993). The binding of
ligand causes the dissociation of the heat-shock proteins and the migration of the
receptor to the nucleus where it dimerizes to either homodimers (ERα:ERα or
ERβ:ERβ) or heterodimers (ERα:ERβ) and binds to ERE located in the promoter
region of target genes (Nilsson et al., 2001). The binding of the ligand also induces a
conformational change of the LBD that allows the recruitment of transcriptional
cofactors (Rosenfeld and Glass, 2001). Two main categories of transcriptional
cofactors can be distinguished: coactivators that activate target gene transcription,
negative coregulators or corepressors that inhibit gene activation and possibly turn
off activated target genes (Nilsson et al., 2001).
ERE-independent pathway
The fact that around one third of the genes in humans that are regulated by
ERs do not contain ERE-like sequences (O'Lone et al., 2004) has led to the
subsequent discovery of signaling pathways that deviate from the classical model. In
the ERE-independent signaling, ligand-bound ERs interact with other transcription
factor complexes like fos/jun (AP-1-responsive elements) (Kushner et al., 2000) and
SP-1 (GC-rich SP-I motifs) (Porter et al., 1997; Saville et al., 2000) that are already
bound to their specific sites on DNA and influence the transcription of target genes
whose promoters do not harbor EREs. The actions of ERs at SP-1 or AP-1 binding

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 13
sites (also called “tethering”) depend on the ligand, the cell type and the receptor
subtype involved.
Ligand-independent mechanism
In the absence of ligand, other signaling pathways can influence ER
transcriptional activity either by directly targeting the receptor (through
phosphorylation) or by regulating the activity of receptor coregulators. These
signaling pathways include:
- Regulators of the general cellular phosphorylation state such as protein
kinases A (PKA) (Aronica and Katzenellenbogen, 1993; Ince et al., 1994) and
C (PKC) (Migliaccio et al., 1993; Lahooti et al., 1998);
- Peptides Growth factors such as epidermal growth factor (EGF) (Ignar-
Trowbridge et al., 1992; Bunone et al., 1996), insulin-like growth factor (IGF-
1), and transforming growth factor alpha (TGF-α) (Ignar-Trowbridge et al.,
1996);
- Neurotransmitters (dopamine) (Power et al., 1991; Smith et al., 1993);
- Cell cycle regulators such as cyclin D1 (Neuman et al., 1997; Prall et al., 1998)
and cyclin A (Trowbridge et al., 1997);
- NF-κB (Chu et al., 2004; Mahmoodzadeh et al., 2009).

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 14
2.1.3.3.2 Non-genomic actions
Apart from intracellular cellular ERs, membrane-associated ERs have been
described and are thought to rapidly alter cellular physiology by activating G-protein
coupled receptor (GPCR)-associated pathways. These non-genomic effects include:
i) regulation of intracellular calcium homeostasis and mitogen-activated protein
kinase (MAPK) pathway in different cell lines (Chen et al., 1999; Improta-Brears et
al., 1999); ii) stimulation of adenylate cyclase activity and cyclic adenosine
monophosphate (cAMP) production (Aronica et al., 1994); and iii) activation of the
phosphoinositol 3-kinase (PI3k) signaling pathway (Chen et al., 1999; Castoria et al.,
2001; Marino et al., 2002).
Figure 5: Molecular pathways of ERs actions (Heldring et al., 2007). TF= transcription factor;
SM= second messenger.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 15
2.1.4 Estrogens and health
Estrogens play an important role in the growth and differentiation of the female
reproductive organs that are also known as its classical targets. They are also
involved in the function of several other tissues and organs including the
cardiovascular and nervous systems, bone, skeletal muscle…
2.1.4.1 Estrogens and hormone-dependent cancers
Estrogens are crucial for the normal growth and development of reproductive
organs.
Ovaries: estrogens have direct and indirect (through the pituitary gland)
stimulatory effects on follicle growth.
Uterus: estrogens increase uterine weight through stimulation of fluid retention
and hyperplasia of the cells of the endometrium, effects that prepare the
uterus for implantation. They also induce the secretion of cervical mucus that
facilitates the penetration of spermatozoa in the uterus.
Mammary tissue: estrogens are required for complete ductal elongation and
branching, and the subsequent development of the lobulo-alveolar end buds
(Brisken and Rajaram, 2006).
Vagina: estrogens stimulate the thickening, the stratification and keratinization
of epithelial cells of the vagina. They also play an important role in the
secretion of mucus that lubricates the vagina.
However, animal studies using rodents have shown that estrogens can induce
and promote mammary (Shull et al., 1997; Cavalieri et al., 2006), endometrial
(Newbold et al., 1990) and other cancers (Kirkman, 1959; Bhat et al., 1993).
Similarly, estrogens have been shown to promote tumor growth and the proliferation
of cancer cells in vivo and in vitro. In humans, parameters such as early menarche,
late menopause, short breastfeeding periods, and low number of pregnancies that
maximize the number of ovulatory cycles and therefore the lifetime exposure to
estrogens are associated with an increased risk of breast-cancer. Estrogens

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 16
supplementation in postmenopausal women is associated with an increased risk of
breast and endometrial cancers.
Two main mechanisms (Figure 6) are proposed to explain the carcinogenic effect
of natural and synthetic estrogens:
Estrogens affect the rate of cell division and stimulate the proliferation of target
cells via ER-mediated processes. This increased and abnormal cell
proliferation can result in increased DNA damage leading to cancer if not
repaired. It is also proposed that estrogens may act indirectly by enhancing
the replication of cells carrying genetic errors (Nandi et al., 1995; Feigelson
and Henderson, 1996; Dickson and Stancel, 2000; Hahn and Weinberg,
2002).
Specific oxidative metabolites (catechol estrogen metabolites) of estrogens
can react with DNA to cause mutations that may initiate cancer (Cavalieri et
al., 1997; Cavalieri et al., 2000; Chakravarti et al., 2001; Cavalieri et al., 2004;
Li et al., 2004; Zhao et al., 2006).

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 17
Figure 6: Mechanisms of estrogens-induced cancer
2.1.4.5 Estrogen and skeletal muscle
Estrogens are involved in the regulation of glucose metabolism in several
tissues including skeletal muscles where they increase glucose uptake. Increasing
evidences also show that estrogens modulate skeletal muscle mass and strength.
Aging is associated with a decline in muscle strength and mass known as
sarcopenia. However, there are gender differences in the age and rate at which
Sarcopenia occurs. The prevalence of sarcopenia is higher among women than men
and women exhibit an accelerated loss of muscle mass and strength at an earlier age
(around the time of menopause)(Calmels et al., 1995; Dionne et al., 2000; Lemoine
et al., 2003; Wiik et al., 2003). Imbalance between muscle protein synthesis and
muscle protein breakdown is the major factor leading to sarcopenia (Short and Nair,
2000). Increased oxidative stress and inflammation (Morley et al., 2001) and
menopause-associated decline in hormonal levels (Maltais et al., 2009) have also
been shown to be implicated. Estrogens positively modulate skeletal muscle mass
and strength (Lowe et al., 2010; Pollanen et al., 2011), decrease oxidative stress in

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 18
muscle (Lowe et al., 2010), and affect myosin function and composition (Kadi et al.,
2002; Moran et al., 2007). A recent study by Enns and Tiidus (Enns and Tiidus, 2010)
suggested a gender and E2-specific influence on membrane and exercise-induced
muscle damage. In the same line, Velders et al (2012) demonstrated that estrogens
were able to regulate skeletal muscle growth and regeneration by stimulating
anabolic pathways, activating satellite cells and modulating immune response.
Both functional ERs isoforms are expressed in human skeletal muscle
(Lemoine et al., 2003; Wiik et al., 2003; Wiik et al., 2009) and despite the recognized
role of estrogens on muscle homeostasis, the mechanisms by which they exert their
effects are not fully understood. Recent studies from our lab show that ERβ is the
major ER isoform mediating estrogens effects on skeletal muscle growth and repair
(Velders et al., 2012; Weigt et al., 2012).

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 19
Section 2.2: Phytoestrogens
2.2.1 Definition and sources
Phytoestrogens (PEs) are naturally-occurring plant compounds that are
structurally and/or functionally similar to mammalian estrogens and their active
metabolites. They are commonly divided into 4 groups: isoflavones (e.g. genistein,
daidzein, glycitein, and formonetin), lignans (e.g. secoisolariciresinol, matairesinol,
pinoresinol, and lariciresinol), coumestans (e.g. coumestrol), and stilbens (e.g.
resveratrol) as presented in Figure 7. Lignans are found in many fiber-rich foods such
as berries, seeds (particularly flaxseeds), grains, nuts and fruits (Nakamura et al.,
2007; Martin-Millan et al., 2010; Patisaul and Jefferson, 2010). Isoflavones are
components of berries, wine, grains and nuts but are most abundant in soybeans and
other legumes (Kurzer and Xu, 1997). Coumestans are found in many plants but
especially in clover and alfalfa sprouts, and stilbens in grape-containing products,
particularly red wine (Jefferson et al., 2012).

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 20
2.2.2 Absorption and metabolism
Isoflavones and lignans are the two major classes of PEs in food and are the
most studied and the best characterized with regard to PE metabolism. Genistein and
daidzein, the predominant isoflavones, occur in soy mainly as biologically inactive
glycoside conjugates (genistin and daidzin) (Patisaul and Jefferson, 2010). After oral
ingestion, the glycoside conjugates are either hydrolyzed into their corresponding
aglycones bioactive form (genistein and daidzein) by intestinal glucosidases or
absorbed as conjugates. Genistein and daidzein can also be obtained from the
intestinal hydrolysis of their respective precursor biochanin A and formonetin that are
found in high amounts in red clover (Axelson et al., 1984). The aglycone metabolites
Genistein Daidzein
Enterolactone Enterodiol
Coumestrol
Resveratrol
Isoflavones
Lignans
Coumestans
Stilbens
Figure 7: Major classes of phytoestrogens

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 21
are further metabolized to more specific metabolites (p-ethylphenol for genistein,
equol or O-desmethylangolensin for daidzein) (Axelson et al., 1984; Bannwart et al.,
1984; Kelly et al., 1993; Joannou et al., 1995). Once in circulation, PE undergo in the
liver several other metabolic pathways including hydroxylation, glucuronidation, and
sulfation (D'Alessandro et al., 2005). Then they are excreted in the urine or in the
bile.
Lignans exist in food as glycosides and are converted to enterolactone and
enterodiol by the gut microflora (D'Alessandro et al., 2005). It is worth to note that the
extent of PE metabolism and therefore the bioavailability of the active metabolites are
highly variable among individuals and are influenced by many factors including the
other components of the diet, gut microflora, use of antimicrobials, intestinal transit
time, and genetic polymorphisms (Setchell et al., 1984; Messina et al., 2006).
2.2.3 Mode and mechanisms of action
Most PEs bind to ERα and ERβ and activate ER-dependent gene transcription
through both ERs. However, unlike E2 that binds with equal affinity to both ERs, they
bind predominantly to ERβ (Kuiper et al., 1997) and may exert both agonistic and
antagonistic effects depending on the cellular context. Therefore, most PEs are
termed as selective estrogen receptor modulators (SERMs). They have been
reported to produce their effects either via direct binding to ER or by acting indirectly
to modulate the concentration of endogenous estrogens. PEs modulate the brain-
pituitary-gonad axis which is the principal endocrine system to regulate mammalian
reproduction. They induce changes in the concentration of circulating steroid
hormones by inhibiting the secretion and activity of gonadotropin releasing hormone
(GnRH) (Bowe et al., 2003; Christoffel et al., 2006; Losa et al., 2011), by increasing
sex hormone binding globulin (SHBG) synthesis (Adlercreutz et al., 1987; Berrino et
al., 2001), and through direct inhibition of some steroidogenic enzymes (Adlercreutz
et al., 1993; Evans et al., 1995; Kao et al., 1998; Brooks and Thompson, 2005) .
PEs may also induce effects that may not be mediated through ERs. For
instance, the isoflavone genistein that is one of the most studied PE inhibited
topoisomerase II (an enzyme involved in replication and transcription of DNA) in NIH
3T3 cells (Okura et al., 1988). Several in vitro studies have also shown that genistein

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 22
inhibits the activity of tyrosine kinase which is essential for cellular growth and
differentiation (Akiyama et al., 1987; Pagliacci et al., 1994; Dalu et al., 1998).
Mutagenic (Kulling and Metzler, 1997; Mitchell et al., 2000; Misra et al., 2002) and
antioxidant properties (Wei et al., 1993) of PEs have been described. Additionally,
PEs exert non-genomic rapid effects for example through the binding of early
intermediate genes such as jun and fos (Kushner et al., 2000).
2.2.4 Phytoestrogens and health
2.2.4.1 Phytoestrogens and the reproductive health
The effects of PEs on female reproductive health depend on applied dose,
route of exposure, and hormonal or developmental status of the exposed individual:
different tissues have species-specific windows of sensitivity to morphological and
functional disruption (Jefferson et al., 2012). Since Bennetts et al (1946) reported that
sheep that grazed on red clover were infertile due to the estrogenic content in clover,
several others have also reported on the impact of dietary PEs on the reproductive
function in adults. There are limited available literature answering to this question
with regard to the situation in humans but considerable studies have been performed
in rodents. Perinatal PEs exposure in rodents is associated with an altered timing of
puberty onset and estrous cyclicity as well as an abnormal development of the
female reproductive tract (Kouki et al., 2003; Nikaido et al., 2004; Jefferson et al.,
2005; Bateman and Patisaul, 2008; Jefferson et al., 2012).
2.2.4.2 Phytoestrogens and gynecological cancers
Although weaker than natural estrogens, PEs mimic estrogenic effects on the
reproductive organs. This raises some concerns because estrogens are implicated in
the pathology of hormone-dependent cancers. However, epidemiologic studies show
that Asiatic women have a significant lower incidence of hormone-dependent cancers
compared to their age-matched Caucasian women and that this lower risk is
positively correlated to PE consumption (Adlercreutz et al., 1992; Knight and Eden,
1996; Setchell and Cassidy, 1999; Messina and Wood, 2008). This observation has
motivated the scientific community to conduct intensive research that has
unfortunately yielded heterogenic and contradictory results (probably because of the
critical time-window of exposure). The impact of PEs on the incidence and

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 23
progression of reproductive cancer is controversially discussed and this controversy
is fueled by the fact that apart from binding to ER and mimic estrogen effects, PEs
have a plethora of other effects which are not yet fully understood. Moreover, as
previously mentioned with regard to the impact on reproductive health, studies of our
lab and other have revealed that the time of exposure to PEs is an important
parameter to consider when evaluating breast cancer outcomes (Whitsett and
Lamartiniere, 2006; Lee et al., 2009; Molzberger et al., 2012). Nevertheless, recent
studies report beneficial effects of PE consumption on gynecological cancer risk. In a
3 years randomized controlled trial (RCT), Palacios et al (2010) assessed the effects
of an oral soy isoflavone extract (Phytosoya 70 mg daily dose) on the endometrium
and breast of 395 postmenopausal women and found no increase in endometrial and
breast cancers. Steinberg et al (2011) demonstrated that 403 healthy
postmenopausal women that received a daily isoflavone dose of 80-120 mg for one
year showed no significant difference in endometrial thickness or fibroids. In the
same line, Ollberding et al (2012) found a reduced risk of endometrial cancer
associated with total isoflavone, daidzein or genistein intake after 13.6 years follow
up of 46027 non hysterectomized postmenopausal women. Myung and coworkers
(Myung et al., 2009) also showed a protective effect of soy intake on the risk of
ovarian cancer. The possible actions of phytoestrogens in relation to cancer are
presented on Figure 8.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 24
Figure 8: Overview of possible actions of phytoestrogens in relation to cancer (adapted from
(Mense et al., 2008)).
2.2.4.3 Phytoestrogens and menopausal symptoms
Despite intensive investigations on the effects of PE on the vasomotor
symptoms of menopause (mainly hot flushes), it remains unclear whether PEs are
clinically relevant for the management of these disorders. Bolanos and coworkers
(2010) recently performed a systematic review and meta-analysis of 90 randomized
controlled trials (RCT) to determine whether soy intervention either as diet, extract or
concentrate reduces the incidence of hot flashes in climacteric women. They found a
significant tendency in favor of soy ISO but were not able to establish conclusive
results due to the high heterogeneity found in the studies. Following a similar
exercise, Howes et al (2006) concluded that soy supplementation may produce a
slight to modest reduction of the number of daily flushes.
The impact of PEs, particularly ISO, on the cardiovascular health is also
controversially discussed with some studies reporting beneficial effects and others no

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 25
effect. Hypercholesterolemia is a well-documented and established risk factor for
coronary heart disease and soy proteins but not purified isoflavones have been
shown to lower serum cholesterol levels (Nagata et al., 1998; Dewell et al., 2006).
Several randomized placebo-controlled trials have been performed to
determine the effect of oral ISO supplementation on endothelial function as
measured by flow-mediated dilatation (FMD) in postmenopausal women. In a meta-
analysis of 9 of such trials, Li et al (2010) found that oral ISO supplementation does
not improve endothelial function in postmenopausal women with high baseline FMD
levels but leads to significant improvement in women with low baseline FMD levels.
Bone cell culture and the rat model of postmenopausal osteoporosis have
been extensively used to show that PE can prevent postmenopausal bone loss (Lee
et al., 2004; Hertrampf et al., 2007). Setchell and lydeking-Olsen (2003) compared
data from 17 studies on cultured bone cells, 24 studies of animal model for
postmenopausal osteoporosis, 15 human observational/epidemiological studies, 17
dietary intervention studies and found that diets rich in PE have bone-sparing effects
in the long-term. Yamori et al (2002) came to the same conclusion after an
intervention study performed with immigrants Japanese women in Brazil.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 26
Section 2.3: Erythrina excelsa Baker
2.3.1 Taxonomy
Kingdom: Plantae
Phylum: Magnoliophyta
Class: Magnoliopsida
Order: Fabales
Family: Fabaceae, Leguminosae,
Genus: Erythrina
Botanical names: Erythrina bagshawei Baker F., Erythrina seretii De Wild, Erythrina
excelsa Baker
Figure 9: Erythrina Excelsa (April 2011 in Kumba-Cameroon by Stephane Zingue)

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 27
2.3.2 Description and geographical distribution
The genus Erythrina comprises around 110 species of orange or red flowered
trees, shrubs and herbaceous plants found throughout tropical and semi-topical
regions of the World. These species are distinguished based on their morphology
mainly the color and shapes of the flowers and fruits, and the inflorescence
characteristics (Majinda et al., 2005). E. excelsa is a deciduous medium sized tree of
up to 30m tall that grows very fast and usually flowers when leafless. The
inflorescence of E. excelsa is compact, axillary or terminal. The flowers are bisexual,
papilionaceous with a spathe-like, glabrous or slightly hairy calyx that is splitted at
one side. The corolla is orange-red to scarlet or coral-red. The fruit is a linear-oblong
spirally twisted pod markedly constricted between the orange-red with whitish scar
seeds (up to 10/fruit) (Lemmens, 2008). E. excelsa is mainly found in the Democratic
Republic of Congo, Cameroon, Ivory Coast, Mali, Southern Nigeria, Sudan, and
throughout East Africa except Zanzibar.
2.3.3 Active constituents of plants of the genus Erythrina
Several secondary metabolites have been reported : alkaloids, flavanones,
flavonols, chalcones, cinnamoyl phenols, stilbenoids, isoflavones, isoflavans,
isoflavanones, pterocarpans, isoflav-3-enes, 3-phenoxychromones, coumastans, 3-
phenylcoumarins, lignans, cinnamate esters, simple phenolics, triterpenes,
sesquiterpenes, squalene, C-18 unsaturated hydroxycarboxylic acid, and
octacosanol (Majinda et al., 2005). Previous phytochemical studies reported the
isolation of a long chain ester of ferulic acid (Wandji et al., 1990) and a prenylated
isoflavonoid (Tanee Fomum et al., 1986) from E. excelsa.
2.3.4 Traditional uses of Erythrina excelsa Baker
The wood of E. excelsa is popular in Uganda for making drums, stools,
shields, mortars and carving for the tourist industry. It is also used as fuel wood and
for making charcoal (Lemmens, 2008). As in Uganda, E. excelsa is planted in
Cameroon as an ornamental and roadside tree.
Some village communities in the South West region of Cameroon reported the
use of E. excelsa for the management of menopausal complaints and for the

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 28
treatment of growths in the female genital tract (ovarian cysts and uterine fibroids).
Others have also reported the use of E. excelsa as an antidote for snakebites (Owuor
and Kisangau, 2006; Lemmens, 2008; Samy et al., 2008) and for arthritis and catarrh
(Jiofack et al., 2010).

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 29
Section 2.4: Ecdysterone
2.4.1 Origin and sources
Ecdysterone (Ecdy) (Figure 10) belongs to the large family of ecdysteroids that
are polyhydroxylated hormones found in arthropods. Several plant species
synthesize ecdysteroids and there were more than 300 reported phytoecdysteroids in
2008 (Bathori et al., 2008). Some of these ecdysteroids containing plants are
included in the human diet (spinach) and are used in several traditional medicine
systems for the treatment of various ailments. For example:
- Rhaponticum carthamoides is used in the Siberian traditional medicine
in cases of overstrain and common weakness after illness (Petkov et
al., 1984);
- Achyranthes bidentata is used in Chinese traditional medicine for the
treatment of osteoporosis (Zhang et al., 2012);
- Trianthema portulacastrum is used in Ayurvedic traditional medicine as
analgesic, laxative and for the treatment of blood diseases, anemia,
inflammation, and night blindness (Shivhare et al., 2012).
Figure 10: 20-hydroxyecdysone (20 HE) or Ecdysterone

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 30
2.4.2 Biological effects
In plants, phytoecdysteroids act as protective agents against predatory insects
(Le Bizec et al., 2002). In insects, ecdysteroids exert trophic effects on muscles and
play an important role in metamorphosis (hence the name molting hormone)
(Schwartz and Truman, 1984). This muscle growth stimulatory effect of ecdysteroids
has inspired several research activities to test whether they could produce the same
effect in vertebrates and mammals in particular.
In vitro and in vivo experimental models have shown that Ecdy exhibits
anabolic effect on mammalian skeletal muscles. Ecdy has been demonstrated to
increase protein synthesis in skeletal muscle (Chermnykh et al., 1988) and in other
organs (Otaka et al., 1968; Chaudhary et al., 1969; Otaka et al., 1969; Hikino and
Takemoto, 1972; Syrov and Kurmukov, 1976). Gorelick et al (2008; 2010) using
cultured murine C2C12 myotubes showed that this increased protein synthesis was a
result of direct or indirect stimulation of the PI3K/Akt signaling pathway as indicated
on Figure 11.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 31
Figure 11: Pathways of Ecdysterone-induced protein synthesis in skeletal muscle (Gorelick-
Feldman et al., 2010). 20HE: 20-hydroxyescdysone; PIP2: phosphatidylinositol-4,5-
diphosphate; PLC: phospholipase C; IP3: Inositol-1,4,5-triphosphate; IP3R: IP3 receptor; SR:
sarcoplasmic reticulum.
Toth et al (2008) showed that treatment of adult non-castrated male Wistar
rats for 7 consecutive days with 5 mg/kg BW Ecdy leads to an increase of the fiber
size of both soleus and extensor digotorum longus (EDL) muscles. They proposed
that this increase may involve the activation of satellite cells as they found that the
number of nuclei also increased in Ecdy treated animals.
To date, it is not clear whether Ecdy exerts the same effects in humans
although ecdysteroids were reported as being used by athletes in Asian countries
(Bathori et al., 2008) and by Russian Olympic athletes (Le Bizec et al., 2002). Schoch
et al (2006) investigated the effect of 8 weeks Ecdy supplementation on body
composition, markers of catabolism (creatinine, creatine kinase, lactate
deshydrogenase...), and on training adaptation of 45 resistance-trained males and
found no effect on the parameters considered.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 32
Conversely to anabolic-androgenic steroids (AAS) that increase muscle mass
mainly through their binding to androgen receptor (AR), no nuclear receptor that is
homologous to the ecdysone nuclear receptor (EcR) found in insects have yet been
described in mammals (Gorelick-Feldman et al., 2010). Ecdysterone has been
characterized as devoid of binding ability to either AR (Bathori et al., 2008; Gorelick-
Feldman et al., 2008), ER (Bathori et al., 2008; Seidlova-Wuttke et al., 2010), or
glucocorticoid receptor (Bathori et al., 2008). Therefore, the mechanisms underlying
the action of Ecdy in mammals are still to be clarified.

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 33
Section 2.5: Marker genes used
The molecular markers for estrogenicity/cytotoxicity used in this work are
described in the following paragraphs.
2.5.1 Complement component 3 (C3)
The C3 gene encodes for a protein called complement component 3 that plays
a key role in a part of the body´s immune system known as complement system.
Together with other proteins of the complement system, C3 triggers inflammation,
serve as a first line of defense against infection, and acts to bridge the innate and the
adaptive immune system (Carroll and Prodeus, 1998; Barrington et al., 2001). The
C3 gene is constitutively expressed in the liver and its up-regulation in the uterine
endometrium following estrogen administration has been extensively demonstrated
(Diel et al., 2004; Hofer et al., 2009; Kluxen et al., 2012).
2.5.2 Clusterin
Clusterin, also known as testosterone-repressed protein message 2 (TRPM2),
Apolipoprotein J (ApoJ), or sulphated glycoprotein 2 (SGP-2), is a 75-80 kDa
heterodimeric glycoprotein produced by a wide array of tissues and found in most
biological fluids. Based on its distribution and in vitro properties, several physiologic
functions have been proposed and include complement regulation, lipid transport,
sperm maturation, initiation of apoptosis, endocrine secretion, membrane protection,
and promotion of cell interactions (Rosenberg and Silkensen, 1995). Estrogen-
dependent regulation of the clusterin mRNA-expression has been reported in rat
uterus (Diel et al., 2000) and in endometrial estrogen-dependent carcinoma
(Wunsche et al., 1998).
2.5.3 Insulin-like growth factor binding protein 1 (IGFBP-1)
IGFBP-1 mRNA and protein expression have been identified in human liver, in
uterine decidua, and in nonhuman kidney. It is one of the six structurally homologous
proteins that specifically bind and modulate the mitogenic and metabolic actions of

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 34
insulin-like growth factor (IGF)-I and IGF-II (Lee et al., 1993). It has been shown in
vitro (Diel et al., 1995), in vivo (Geis et al., 2005), and in humans (Isotton et al., 2012)
that estrogens up-regulate IGFBP-1 expression.
2.5.4 Vitamin D dependent calcium binding protein 9kDa (CaBP9K)
CaBP9k or calbindin-9k is a member of a large family of intracellular calcium
binding proteins that have high affinity for calcium (Kim et al., 2012). It plays an
important role in calcium transport and re-asorbtion in many tissues (Wasserman and
Taylor, 1966; Bruns et al., 1988; Balmain, 1991; Krisinger et al., 1994). Diel et al
(1995) identified CaBP9k as an estrogen regulated gene in Fe33 rat hepatoma cells.
The CaPB9k promoter contains several EREs (Lee et al., 2003).
2.5.5 Cyclin D1 (CCND1)
Cyclin D1 together with other members of cyclins family regulate progression
through the cell cycle leading to mammalian cell division. Upon binding to cyclin
dependent kinases Cdk4 and Cdk6, CCND1 phosphorylates the retinoblastoma
protein (Rb) and promotes the transition from G1 to S phase. CCND1 gene is
amplified in 15% of primary ER positive breast cancers and is overexpressed in up to
50% of cases (Tobin and Bergh, 2012). Although there is no classical ERE in the
CCND1 promoter (Liu et al., 2002), estrogens when bound to ER (ERα) are able to
upregulate CCND1 mRNA expression (Liu et al., 2002; Tobin and Bergh, 2012). On
the other hand, CCND1 has been shown to directly interact with the LBD of ER and
to stimulate ER transactivation in a ligand-independent fashion (Zwijsen et al., 1997)
2.5.6 Bax
Bax belongs to the proapoptotic members of the Bcl-2 family of proteins whose
role in the regulation of apoptotic cell death is well characterized. It is a soluble
monomeric protein that is localized in the cytosol or is attached at the periphery of
intracellular membranes in unstimulated cells (mitochondria, endoplasmic reticulum,
and nucleus) (Goping et al., 1998; Danial, 2007). Upon a death stimulus, Bax is
activated and translocates to the outer mitochondrial membrane where it promotes
the release of cytochrome C from the mitochondria. Once released, cytochrome C

Literature review
M.Sc. Sadrine Tchoukouegno Ngueu 35
activates a caspase cascade leading to increased mitochondrial mediated apoptotic
cell death (Adams and Cory, 1998).
2.5.7 Bcl-2
Bcl-2 is the prototype protein of the BCL-2 family and was first discovered as a
proto-oncogene in follicular lymphoma (Tsujimoto et al., 1984). It codes for an
antiapoptotic protein that acts by binding and antagonizing the proapoptotic effectors
Bax and Bak (Carpinelli et al., 2012). Bcl-2 protein is overexpressed in a variety of
solid organ malignancies including breast cancer (Dawson et al., 2010). Estrogens
have been shown to prevent apoptosis of normal and breast tumor cells through the
activation of Bcl-2 gene expression (Somai et al., 2003).

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 36
CHAPTER 3: MATERIAL AND METHODS

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 37
3.1 Material
3.1.1 Substances
The following substances were used either as test substance or positive
control:
Substances Chemical structure Manufacturer
1. 17β-Estradiol or E2 (Estra-1,3,5(10)-trien-3,16α,17 β-diol)
Sigma-aldrich (Steinheim, Germany)
2. Genistein (Gen) (4´,5,7-Trihydroxyisoflavon)
LC Laboratories (Wolburn, USA)
3. ERα-selective agonist or ALPHA [(16α-LE2), 3,17-dihydroxy-19-nor-17α-pregna-1,3,5 (10)-triene-21,16α-lactone;)]
Bayer HealthCare Pharmaceuticals (Berlin-Germany)
4. ERβ-selective agonist or BETA [(8β-VE2), 8-vinylestra-1,3,5 (10)-triene-3,17ß-diol]
Bayer HealthCare Pharmaceuticals (Berlin-Germany)

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 38
5. ZK 283361 (ERβ antagonist or Anti-BETA)
Bayer HealthCare Pharmaceuticals (Berlin-Germany)
6. 20-Hydroxyecdysone or Ecdy ( 2β,3β,14α,20β,22,25-Hexahydroxy-7-cholesten-6-one)
Sigma-aldrich (Steinheim, Germany)
7. Cisplatin (cisplatinum, or cis-diamminedichloroplatinum)
Millipore (Darmstadt, Germany)
8. ZK 191703 or ZK (pure antiestrogen )
Bayer HealthCare Pharmaceuticals (Berlin-Germany
9. Dihydrotestosterone (DHT)
Sigma-aldrich (Steinheim, Germany)
10. Flutamide (Flu)
Sigma-aldrich (Steinheim, Germany)
11. LONG®R3 IGF-1, human recombinant analog (IGF-1)
Sigma-aldrich (Steinheim, Germany)

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 39
3.1.2 Chemicals, consumables and lab equipments
Table 1: List of chemicals
Chemicals Manufacturer
Yeast growth and assay media components
Adenine Sigma-aldrich (Steinheim, Germany)
CuSO4 _
CPRG Roche diagnostics (Mannheim, Germany)
Fe2(S04)3 Merck (Darmstadt, Germany)
KH2PO4 _
KOH _
L-arginine-HCI Sigma-aldrich (Steinheim, Germany)
L-aspartic acid _
L-glutamic acid _
L-histidine _
L-Ieucine _
L-isoleucine _
L-lysine-HCl _
L-methionine _
L-phenylalanine _
L-serine _
L-threonine _
L-tyrosine _
L-valine _
MgSO4 _
(NH4)2S04 Merck (Darmstadt, Germany)
Pantothemic acid Sigma-aldrich (Steinheim, Germany)
Pryridoxine _
Thiamine _
Inositol _
Biotin _
D-(+)-glucose Merck (Darmstadt, Germany)
Buffers components
Na2HPO4 *2H2O Merck (Darmstadt, Germany)
NaCl _
NaH2PO4* H2O _
NaOH _
Tris _
Triton X 100 Sigma-aldrich (Steinheim, Germany)
Tween 20 Sigma-aldrich (Steinheim, Germany)
Sucrose

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 40
Bromophenol blue Merck (Darmstadt, Germany)
Xylene cyanol _
HCl (1M) _
NaOH (1M) _
EDTA Sigma-aldrich (Steinheim, Germany)
Glacial acetic acid Merck (Darmstadt, Germany)
PMSF Sodium deoxycholate
Protein analysis
BSA PAA Laboratories (Pasching, Austria)
Milk powder Reformhaus Bacher (Cologne, Germany)
SDS-loading buffer (Laemmli 1970) BIO-RAD (Munich, Germany)
NuPAGE Novex 4–12% Bis–Tris Midi Gel Invitrogen, life technologies
DC protein assay reagent A BIO-RAD (Munich, Germany)
DC protein assay reagent B _
Precision Plus Protein™Dual color Standards _
Cell culture media components
DMEM Gibco, Invitrogen (Karlsruhe, Germany)
Glutamin _
FBS _
Horse serum _
Human insulin _
PBS cell culture grade _
Penicillin-Steptomycin _
Trypsin-EDTA (0.05%) _
RPMI 1640 _
Sodium bicarbonate _
Sodium pyruvate _
Freezing medium _
PCR reaction components
dNTPs Invitrogen, life technologies (Karlsruhe, Germany)
PCR buffer _
MgCl2 for PCR _
Taq polymerase _
Ampuwa _
Primers _
RNA preparation
TRIzol ®reagent Invitrogen, life technologies (Karlsruhe, Germany)
Chloroform Merck (Darmstadt, Germany)

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 41
Isopropanol _
EtOH for molecular biology _
DEPC
Nucleic acids electrophoresis and staining dyes
DAPI Sigma-aldrich (Steinheim, Germany)
Propidium iodide (PI) _
Ethidium bromide _
Sybr Green BIO-RAD (Munich, Germany)
Agarose Biozym (Hess. Oldendorf, Germany)
100 bp DNA ladder Invitrogen, life technologies (Karlsruhe, Germany)
Kits
MTT Roche diagnostics (Mannheim, Germany)
XTT Roche diagnostics (Mannheim, Germany)
QuantiTect® Reverse transcription Qiagen
Other Organic solvents
EtOH for histology
MeOH Merck (Darmstadt, Germany)
MeOH HPLC grade VWR (Leuven,Belgium)
Cyclohexane HPLC grade _
Dichloromethane HPLC grade _
Xylol Quadflieg (Geselkirchen, Germany)
DMSO Merck (Darmstadt, Germany)
Formaldehyd 37% _
Others
Paraffin Sasol wax (Hamburg, Germany)
Corn oil Sigma-aldrich (Steinheim, Germany)
Glutardialdehyde Merck (Darmstadt, Germany)
Activated carbon _
Meliseptol B. Braun (Melsungen, Germany)
Liquid nitrogen Air Liquide (Düsseldorf, Germany)
Eosine VWR (Leuven,Belgium)
Mounting medium (Entellan®) Merck (Darmstadt, Germany)
Mayer Hamalun _
Silicagel (60 Å, 70-230 mesh) Sigma-aldrich (Steinheim, Germany)

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 42
Antibodies
Table 2: List of antibodies
Antibody Host Dilution Manufacturer
Primary antibodies
Anti-β-actin(clone AC-74) mouse 1:5000 Sigma-aldrich
Anti-ERβ (H-150) rabbit 1:200 Santa Cruz Biotechnology,
Inc
M30 CytoDEATH mouse 1:10 Roche Applied Science
secondary antibodies
Anti-Mouse Ig, HRP* rabbit 1:2000 Dako
Anti-Rabbit Ig, HRP Swine 1:2000 Dako
Anti-Mouse Ig biotinylated Goat 1:400/1:200 Dako
signal detection reagents
chemoluminescent POD-Substrate (LumiLight Plus)
Roche Diagnostics
FluoroLinkTM Cy3 labelled streptavidin 1:1000
Amersham Biosciences
* Horseradisch peroxidase
Table 3: Consumables and equipement.
Consumables and equipments Manufacturer
42ippsipette tips 1 – 5mL _
Eppendorf Tubes Eppendorf (Essling-Berzdorf, Germany) RT-qPCR plates 96-well plates without border QPCR
Agilent Technologies (Waldbronn, Germany)
Optical Cap Mx3000P/Mx3005P 8 x Strip _
Falcon tubes 15 mL BD Biosciences(Heidelberg, Germany)
Falcon tubes 50 mL BD Biosciences(Heidelberg, Germany)
Gloves Comfort Unigloves (Troisdorf, Germany)
Gloves Nitrile VWR (Leuven,Belgium)
Syringes Injekt solo 2-10 mL B. Braun (Melsungen, Germany)
Sterile syringue filter 0.2 μm VWR (Leuven,Belgium)
Cannulas 0.6 x 30 mm HenrySchein (Melville NY, USA)
Towels facial tissues Flirty H.-Kimberly (Mainz, Germany)
Coverslips Coverslips 50x30 mm VWR (Leuven,Belgium)
Polysine slides _
Embedding cassette Histosette _
pH meter Mettler Toledo (Giessen, Germany)
Cell scraper BD Biosciences(Heidelberg, Germany)
Protran®Nitrocellulose Membranes Whatman Inc (USA)
96-well optically flat bottom plates Nunc (Danemark)
96-well optically round bottom plates BD Biosciences(Heidelberg, Germany)
6-well plates _
Chromatography column NS 14/23 Lenz Laborglas GmbH & Co. kG

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 43
(Wertheim, Germany)
Ice Machine, AF 80 Scotsman (Milan, Italy)
Freezer (-20 °C) Liebherr (Bulle, Switzerland)
Freezer (-80 °C) HERAfreeze Heraeus ( Hanau, Germany)
Gel chamber small or medium Biometra (Göttingen, Germany)
Camera and evaluation system for agarose gels MWG-Biotech (Ebersberg, Germany)
Refrigerator Liebherr (Bulle, Switzerland)
Refrigerated Centrifuge Centrifuge 5417 R Eppendorf (Essling-Berzdorf, Germany)
Microwave MS192A LG (Willich, Germany)
PCR GeneAmp PCR system 9600 Applied Biosystems (Foster City, USA)
Pipettes Eppendorf (Essling-Berzdorf, Germany)
RT-qPCR Stratagene Mx3005P Agilent Technologies
Spectrophotometer NanoDrop 1000 Thermo Fisher Scientific (Schwerte, Germany)
Benchtop centrifuge Biofuge 13 Heraeus (Hanau, Germany)
Vortex Genie 2 Scientific Industries (Bohemia NY, USA)
Libra Scaltec SBA52 Denver Instrument (Göttingen, Germany)
Inverted microscope Axiovert 200M Zeiss (Jena, Germany)
Confocal microscope Axiophot KS300 Zeiss (Jena, Germany)
Microscope Camera (3 CCD colr video camera) Sony (Tokio, Japan)
Microscope Camera B / W HAL 100 Zeiss (Jena, Germany)
Microtome Autocut Mod 1140 Reichert-Jung (Vienna, Austria)
Oven T5052 E Heraeus ( Hanau, Germany)
Embedding centre MPS/P2 Slee (Mainz, Germany)
Microplate Reader MWG-Biotech (Ebersberg, germany) XCell4 SureLock™ Midi-Cell electrophoresis system Invitrogen ( Karlsruhe, Germany)
Trans-Blot®Turbo™ Transfert System BIO-RAD (Munich, Germany)
BD FACSCalibur flow cytometer BD Biosciences (USA)
Elisa reader Thermoscientific (Lagenselbord- Germany)
CO2 incubator C 150 Binger GmbH (Tuttligen, Germany)
Waterbath Kottermann (Vienna, Austria)
Ultrospec 2000 (UV/visible spectrophotometer) Pharmacia Biotech
SANYO Incubator MIR 152 SANYO Electric Co.Ltd (Japan)
Microplate shaker VWR (Leuven,Belgium)
Autoclave Systec GmbH (Wettenberg, Germany)
HeraSafe biological safety cabinet Heraeus ( Hanau, Germany)
HPLC system Thermo Finnigan, San Jose, CA
UHPLC system Thermoscientific (Lagenselbord- Germany)
LTQ-Orbitrap Mass Spectrophotometer _
LTQ-Orbitrap Mass Spectrophotometer _ Hypersil Gold column (100 x 2.1 mm id, 3µm particle size) _ Discovery Supelco HS C-18 column (25 x 4.6 mm, 5µm particle size) Sigma-aldrich (Steinheim, Germany)

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 44
3.1.3 Test organisms
3.1.3.1 Animals
Juvenile female Wistar rats aged 5 weeks and weighing 101-124 g were
obtained from Janvier (Le Genest St Isle, France). They were maintained under
controlled temperature (20±1°C), humidity (50-80%), and illumination (12h light, 12h
dark) conditions. The rats had free access to standard rat soy free diet (Sniff R10-
Diet, Ssniff GmbH, Soest Germany) and water. All animal handling and experimental
conditions were carried out according to the Institutional Animal Care and Use
Committee guidelines regulated by the German federal law for animal welfare.
3.1.3.2 Yeast strain
Yeast cells (saccharomyces cerevisiae) originated from Glaxo. They are stably
transfected with the human estrogen receptor alpha (hERα) gene, together with
expression plasmids containing estrogen-responsive elements and the lac-Z reporter
gene encoding the enzyme β-galactosidase.
3.1.3.3 Cell lines
MCF-7
MCF-7 cells are immortalized cells from human breast adenocarcinoma that
were isolated in 1970 from a 69 year-old Caucasian woman. They grow as an
adherent monolayer in cell culture flasks and have retained several characteristics
particular to the mammary epithelium. Moreover, estrogens can activate the ER
found in their cytoplasm and they are known as ER positive cell line. MCF-7 cells are
useful for in vitro breast cancer studies and were kindly provided by Bayer
HealthCare Pharmaceuticals (Berlin, Germany).
HT-29
HT-29 cells were isolated in 1964 from a primary adenocarcinoma of a 44
year-old Caucasian woman. They grow as adherent monolayer and are able to
express differentiation features that are characteristic of mature intestinal cells. Apart
from their particular use in glucose metabolism and hormones receptors studies, they

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 45
are like all other cancer cell lines used in the area of cancer research. They were also
provided by Bayer HealthCare Pharmaceuticals.
C2C12
The C2C12 cell line is an immortal line of mouse skeletal myoblasts derived
from the thigh muscle of a C3H mouse after a crush injury. C2C12 cells grow as
adherent monolayer and exhibit characteristic features of satellite cells. They are
used to study the process of satellite cells activation, proliferation and differentiation.
Because they are able to differentiate in contractile myotubes producing
characteristic muscle protein, C2C12 are also used to study several aspects of
skeletal muscle function and adaptation. They were purchased from ATCC (American
Type Culture Collection, CRL-1772).
3.2 Methods
3.2.1 Preparation of cell culture media and buffers
Table 4: Yeast culture medium components
Stock solutions Components/concentration
Minimal medium
13.61 g KH2PO4, 1.98 g (NH4)2S04, 4.2 g KOH pellets, 0.2 g MgSO4, 1 ml Fe2(S04)3 solution (40 mg/50 ml H20), 50 mg L-Ieucine, 50 mg L-histidine, 50 mg adenine, 20 mg L-arginine-HCI, 20 mg L-methionine, 30 mg L-tyrosine, 30 mg L-isoleucine, 30 mg L-lysine-HCI, 25 mg L-phenylalanine, 100 mg L-glutamic acid, 150 mg L-valine, and 375 mg L-serine to 1 L double-distilled water pH 7.1.
Vitamin Solution
8 mg thiamine, 8 mg pyridoxine, 8 mg pantothenic acid, 40 mg inositol, and 20 mL biotin solution (2 mg/100 mL H20) to 180 mL doulble-distilled water.
L-aspartic Acid 4 mg/mL
D-( + )-glucose 20% w/v
L-threonine 24 mg/mL
Copper Sulfate 20 mM
*CPRG 10 mg/mL
* Chlorophenol red-β-D-galactopyranoside
The minimal medium and all other stock solutions were sterile filtered through
0.2 μm filter and stored at 4°C in glass bottles. Growth medium was prepared by
mixing 5 mL of glucose solution, 1.25 mL of L-aspartic acid solution, 0.5 mL of
vitamin solution, 0.4 mL of L-threonine solution, 125μL of copper sulfate solution to

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 46
45 mL minimal medium solution. Assay medium was prepared by adding 0.5 mL of
CPRG solution to fresh growth medium.
Table 5: C2C12, HT-29 and MCF-7 cells growth media
Cell line Medium composition
C2C12 (Proliferation medium)
420 mL DMEM 50 mL FBS 10 mL glutamin (4nM) 5 mL penicillin/streptomycin (100U/L) 5 mL sodium pyruvate (100mM) 10 mL sodium bicarbonate (7.5%)
HT-29
450 mL DMEM 50 mL FBS 5 mL L-glutamin (4nM) 10 mL penicillin/streptomycin (100U/l)
MCF-7 (Normal growth medium)
450 mL RPMI 50 mL FBS 5 mL L-glutamin (4nM) 10 mL penicillin/streptomycin (100U/l) 1.25 mL human Insulin (4mg/ml)
Charcoal-stripped FBS
In experiments with the aim to assess the impact of E2 or PEs on cell
proliferation, the steroids contained in FBS have to be removed in other to avoid any
interference. To achieve this, activated charcoal was added to FBS (10 mg/mL) and
the mixture shook for 1h at RT. The mixture was afterwards placed at 4 °C overnight
to allow the charcoal to sediment. Then it was centrifuged (3500 rpm, 4 °C) and
sterile filtered through a 0.2 μm syringe filter.
Buffers and solutions
Phosphate-buffered saline (PBS 0.1 M)
Adjust the pH to 7.4
Na2HPO4 *2H2O 14.4 g
NaH2PO4* H2O 2.6 g
NaCl 8766 g
ddH2O 1L

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 47
Tris-buffered saline (TBS 0.05 M)
Tris 6.057 g
NaCl 8.76 g
ddH2O 1L
Ajust the pH to 7.6 with 1M HCl
Depending on the purpose of use (antibody dilution, washing steps), both
buffers were sometimes supplemented with 0.1% Tween 20.
TAE (Tris-Acetate-EDTA) buffer
TAE buffer was used in agarose gel electrophoresis for the separation of RNA
and DNA.
For 1 L 50X TAE Stock Solution:
Tris Base (MW=121.1 g/mol) 242 g
Glacial Acetic Acid 57.1 mL
0.5 M EDTA (pH 8.0) 100 mL
ddH2O 1 L
To prepare 1L of 0.5 M EDTA (pH 8.0) solution:
EDTA (MW = 372.24 g/mol) 186.1 g
ddH2O 1 L
Adjust the pH with 10 N NaOH (40 g NaOH in 100 mL ddH2O).
Lysis puffer
This buffer was used for the preparation of proteins from cells. For 100 mL:
Tris (50 mM) 0.605 g
0.25% sodium deoxycholate 0.25g
NaCl (150 mM) 0.8766g
1 mM EDTA 0.0292g

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 48
1% Triton X 100 1mL
Adjust the pH to 7.4 with HCl
Diethylpyrocarbonate (DEPC)-treated water.
DEPC-treated water is suitable for dissolving RNA after its extraction from
cells or tissues. To prepare 1L, 1 mL of DEPC was added in 999 mL ddH2O and
placed overnight in a water bath at 37°C. Afterwards the water was autoclaved for 20
min to remove the DEPC.
6X Sucrose loading buffer
For 10 mL solution:
Sucrose (40 % w/v)
Bromophenol blue 0.25 %
Xylene xyanol 0.25 %
DNA Ladder
The DNA ladder is a solution of DNA molecules of different lengths that is
used in agarose gel electrophoresis as a reference to estimate the size of an
unknown double-stranded DNA molecule. The DNA ladder was also used in total
RNA quality control to estimate the location of the 28 S and 18 S ribosomal RNA
bands.
For 1 mL solution:
100 bp DNA ladder 50 μL
6x sucrose loading buffer 166 μL
1x TAE buffer 784 μL

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 49
3.2.2 Plant material extraction
The stem bark of E. excelsa was collected in Kumba (in the SouthWest region
of Cameroon) in April 2011. The botanical sample was identified and authenticated at
the Cameroon National Herbarium in Yaounde where a voucher specimen (N° 61487
HNC) is deposited. The samples were air-dried (19-29°C), ground and 1.5 kg of the
powder extracted at room temperature (22 ± 5°C) with ethanol (3× 6 l; 72 h each).
After filtration, the solvent was removed using a rotary evaporator under reduced
pressure.
3.2.3 LC-HRMS and LC-HRMS/MS analysis
An Accela Ultra High-Performance Liquid Chromatography (UHPLC) system
hyphenated to a hybrid LTQ-Orbitrap Discovery Mass Spectrometer was employed
for the extract profiling. The UHPLC system was equipped with an Accela pump, an
Accela autosampler and an Accela PDA detector. Stock solution of 1 mg/mL (MeOH:
H2O, 50/50, v/v) of the crude extract was prepared and 10 μL injected on a Hypersil
Gold column (100 x 2.1 mm i.d., 3 μm particle size). The mobile phase used was
aqueous acetic acid 0.1% (v/v) (solvent A) and acetonitrile (solvent B). The initial
conditions were 98% of solvent A and 2% of solvent B adjusting linearly to 98% B in
28 min. This solvent composition was maintained for 0.9 min (98% B) followed by a
return to the initial conditions (in 0.1 min) and a re-equilibration step (1 min) prior to
the next run. The flow rate was set to 500 μL/min and the wavelengths used for
monitoring the chromatograms were 220, 280, and 365 nm (recording window 200 -
700 nm).
The mass spectrometer was equipped with electrospray ionization (ESI)
source and operated in positive mode. A data dependent triple play method involving
three scan events was developed for the profiling of the extract. This method
consisted of a full scan acquisition, a Selective Ion Mode (SIM) and a MS/MS
acquisition employing the Orbitrap analyzer and was designed so that the major ion
detected in each full scan could be selected, scanned in SIM mode and fragmented,
resulting to highly accurate MS and MS/MS spectra. For all measurements, the mass
tolerance was set at 5 ppm. The ESI source was operated at a sheath gas flow rate
of 50 arb, auxiliary gas flow rate of 10 arb, ion spray voltage of 3.5 kV, capillary

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 50
temperature of 300 oC, capillary voltage of 35 V, tube lens of 110 V and collision-
induced dissociation (CID) of 35% (HRMS/MS acquisitions). Xcalibur 2.0.7 software
was used for the pre-and post acquisition of the results.
3.2.4 Vacuum liquid chromatography (VLC)
The column (200x15mm) was packed with 25 g of silica gel. The dry loading
method was applied for sample application. 1g of the crude extract was dissolved in
methanol, the resulting solution adsorbed on 3g of silica and the solvent allowed to
evaporate. The dry adsorbent loaded with the sample was then applied to the column
and eluted using a step gradient of solvent under vacuum. 250 mL fractions were
collected at each polarity step and the column was allowed to run dry after each
fraction collection. The solvent was evaporated to yield five fractions (Table 6) that
were monitored for biological activity.
Table 6: VLC fractions.
Fraction Solvent Weight (mg)
F1 Cyclohexane(100%) 6.7
F2 Cyclohexane - DCM(*) (50:50) 21.9
F3 DCM(100%) 9.8
F4 DCM - MeOH (50:50) 485
F5 MeOH (100%) 24.1
(*) Dichloromethane
3.2.5 HPLC
The HPLC system consisted of a vacuum degasser, a quaternary pump, an
autosampler and a Diode-Array Detector (DAD). A Discovery SUPELCO HS C-18
column (25 × 4.6 mm, 5 μm particle size) was used for separations. A binary mobile
phase consisting of 1% acetic acid (A) and methanol (B) was used. The initial
conditions were 50% A and 50% B. Then a gradient program was applied as follows:
50-98% B and 50-2% A over 60 min, isocratic 98% B and 2% A for 5 min, return to
the initial conditions over 5 min and maintain of these conditions for 10 min prior to
the following injection. The injection volume was set to 20 μL and the flow rate to
1mL/min. DAD conditions: 254, 280, and 365 nm and recording window from 200 to

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 51
700 nm. The control of the system as well as data acquisition and analysis were
performed using ChromQuestTM 4.1 software.
3.2.6 Yeast estrogen screen
3.2.6.1 Principle
The yeast estrogen screen (YES) (Figure 12) is a transactivation assay used
to identify compounds that can interact with the human estrogen receptor (here hER-
α). In this system, yeast cells (saccharomyces cerevisiae) are transfected with hERα
gene, together with expression plasmids containing estrogen-responsive elements
and the lac-Z reporter gene encoding the enzyme β-galactosidase. Upon binding an
active ligand, the receptor interacts with transcription factors and other transcriptional
components to modulate gene transcription. This causes expression of the reporter
gene lac-Z and the enzyme produced is secreted into the medium where it
metabolizes a chromogenic substrate, chlorophenolred-β-D-galactopyranoside
(CPRG) which is normally yellow, into a red product that can be measured by
absorbance at 540nm.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 52
Figure 12: Estrogen-inducible expression in yeast (Routledge and Sumpter, 1996).
3.2.6.2 Assay procedure
Yeast cells were stored at -80°C as 10X concentrated stock in growth medium
supplemented with 15% glycerin. At day zero, growth medium was inoculated with
yeast stock and incubated at 28°C till an OD of 1 at 690nm was attained
(approximately 24h). The following day, chemicals serially diluted in DMSO were
transferred (2μL) to 96-well optically flat bottom plates. Each plate contained one row
(12 wells) of blanks (2μL DMSO only) and each assay had a 17β-estradiol standard
curve (10-12-10-8 M). Assay medium was inoculated with the optimal yeast density of
the 24 h culture prepared at day 0 and 200μL was given in each well. The plates
were then sealed with autoclave tape and incubated at 32°C in a naturally ventilated
heating cabinet on an orbital shaker. After 3 days of incubation, color change of the
medium was quantified by absorbance at 540 nm. Yeast growth (turbidity) was
measured at 620 nm.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 53
3.2.6.3 Calculations
Dose–response curves for β-galactosidase activity were obtained using
corrected absorbance units (CAU):
CAU = (AU 540nm) compound − [(AU 620nm) compound − (AU 620nm)
blank]. The best-fitting curve was calculated with SigmaPlot for Windows, version 11
using a four-parameter logistic regression according to the following formula:
Hillslope x) (logEC50101
min -max min
CAU
min = baseline absorbance
max = maximal absorbance
EC50 = concentration of test compound yielding a half-maximal effect
X= given concentration of test compound.
Other parameters such as the relative induction efficiency (RIE) and the
relative potency (RP) of each chemical were calculated as follows:
100 E2 min)-(max
compound min)-(max RIE
100 Compound (EC50)
E2 (EC50)RP

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 54
3.2.7 MTT/XTT assays
3.2.7.1 Principle
Both assays are based on the cleavage of yellow tetrazolium salts MTT (3-
(4,5-dimethythiazol-2-yl)-2,5-diphenyl tetrazolium bromide) or XTT (2,3-Bis-(2-
Methoxy-4-Nitro-5-Sulfophenyl)-2H-Tetrazolium-5-Carboxanilide) by mitochondrial
succinate dehydrogenases of metabolic active cells. The colored (dark purple for
MTT and orange for XTT) formazan product is then measured spectrophotometrically
and correlates to cell viability since the reduction of tetrazolium salts can occur only
in metabolically active cells. The difference between the two assays is that the XTT is
performed in the presence of an electron- coupling reagent and the formazan product
formed is water-soluble and does not require a further solubilization step as it is the
case for MTT.
3.2.7.2 Assay procedure
Breast cancer (MCF-7) and colon carcinoma (HT-29) cell lines were used. The
cell culture media were prepared as indicated in Table 5. For the assay, the cells
were washed with phosphate buffered saline (PBS) and detached with 0.05 %
trypsin-EDTA. After centrifugation, the pellet was resuspended in medium, the cells
counted and diluted to give the optimal seeding density (104 cells/well). The cells
were further plated into a 96-well plate, allowed to attach for 24-48 h and incubated
for 3 or 5 days with either cisplatin or the plant extract dissolved in DMSO. Each
concentration was added to the plates in six replicates. The final DMSO
concentration was not more than 0.1%, this concentration being used in the solvent
control wells.
3.2.8 Myotubes diameter assay
C2C12 cells were routinely cultured in a proliferation medium (Table 5). They
were passaged 2 to 3 times/week (at 80% confluence) to avoid confluence that would
cause a depletion of the myoblast population. For the assay, myoblasts in PM were
seeded at a density of 105 cells/well in 6-well plates. Differentiation towards
myotubes was induced on the 3rd day (at confluence) by shifting the proliferation
medium (PM= DMEM+10% foetal bovine serum) to a differentiation medium (DM=

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 55
DMEM supplemented with 2-10% horse serum). Myotubes at day 4-6 post-fusion
were incubated for 48 h with the different substances (Table 7). At the end of the
treatment period, cultures were fixed in 2.5% glutardialdehyde and photographed by
glutaraldehyde-induced autofluorescence using an Axiovert 200 M microscope
equipped with a HaL 100 camera. Average diameters of at least 50 myotubes were
measured for each condition at different points separated by 25-50 µm along the
length of the myotube (Stitt et al., 2004) using the Axiovision LE software.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 56
Table 7: Experimental groups myotube diameter assay.
3.2.9 Western blot
3.2.9.1 Protein preparation from C2C12 cells
C2C12 cells were cultured and seeded for this assay as previously mentioned
with the difference that they were allowed to grow to confluence and to differentiate in
normal growth medium. At 10 days old, cells were incubated for 72h in the presence
of either E2 (10-8 M) or vehicle control (DMSO) and the lysate prepared.
Cell culture dishes were placed on ice and the cells washed twice with ice-cold
PBS. Then ice-cold lysis buffer supplemented with protease enzyme inhibitor PMSF
(phenylmethanesulfonylfluoride, dissolved in isopropanol) at final concentration of 1
mM was added directly in the dishes and the cells scraped off using a cold plastic
scraper. The cell suspension was transferred into pre-cooled micro centrifuge tubes
and maintained for 30 min on ice. After 20 min centrifugation at 12000 rpm, the
supernatants were transferred in new tubes and the protein concentration
determined.
3.2.9.2 Determination of protein concentration
The total level of protein in each sample was determined using the BIO-RAD
DC Protein Assay which is based on the Lowry method (Lowry et al., 1951). BSA
(Bovine Serum Albumin) standard protein samples ranging from 0 to 2 mg/mL were

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 57
prepared in lysis buffer and the myotubes protein samples diluted up to 1: 5. Then 5
µL of each sample protein was introduced in each well of a microplate reader
followed by 25 µL Reagent A and 200 µL Reagent B. After 15 min of incubation at
room temperature (RT), the absorption at 650 nm was read using a microplate
reader. The protein samples were stored at -80°C till further use.
3.2.9.3 SDS gel electrophoresis
Sample protein (20 μg) were mixed with equal amounts of SDS-loading buffer
and loaded on a NuPAGE Novex 4–12% Bis–Tris Midi Gel. After migration at 90 mA
for 60 min using the XCell4 SureLock™ Midi-Cell electrophoresis system, proteins
were blotted for 30 min at 25V, 1A (Trans-Blot®Turbo™ Transfert System) onto
nitrocellulose membranes (Protran®Nitrocellulose Membranes). The membranes
were blocked for 1h at room temperature with 5% dry milk powder in Tris-buffered
saline with 0.1% Tween 20 (TBS-T). The membranes were incubated overnight at
4°C with either β-actin or ERβ antibody diluted (Table 2) in 1% dry milk powder in
TBS-T. After four times wash with TBS-T for 10 min each, the membranes were
incubated for 1h at RT with the corresponding secondary antibody solution. After
another four times wash (10 min each) in TBS-T, blot signals were visualized by the
chemoluminescent POD-Substrate and the images analyzed using the Image J 1.40g
software.
3.2.10 Uterotrophic assay
Juvenile female Wistar rats aged 6 weeks were ovariectomized. After 14 days
of endogenous estrogens decline, the animals weighing 200-250g were randomly
distributed into treatment and vehicle groups (Table 8) and treated for three
consecutive days with a daily dose. The animals were sacrificed by decapitation 24 h
after the last dose. Uterine and liver wet weights were recorded. Half of the uteri were
kept together with vaginas in 4% formaldehyde for subsequent histological analysis
and the rest of the uteri as well as the livers were snap frozen in liquid nitrogen for
RNA preparation.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 58
Table 8: Groups treatment uterotrophic assay
Groups Dose Number of animal
Control 1 mL/kg BW career (20% DMSO + 80% corn oil)
7
E2 4 μg/kg BW 7
E.excelsa 50 mg/kg BW 6
100 mg/kg BW 6
3.2.11 Histological analysis of tissues
The standard paraffin process that includes fixation, processing, embedding,
sectioning, and staining was used for historical anlaysis of tissues.
3.2.11.1 Fixation
To prevent the degenerative process that takes place when tissues are
deprived from their blood supply, tissues were put in cassettes carrying details for
their identification and kept in a 4% formalin solution right after their preparation.
3.2.11.2 Dehydration and clearing
To allow cutting, tissues have to be infiltrated with paraffin wax that is however
not soluble in water or alcohol but in a solvent called xylene or xylol which is miscible
with both alcohol and paraffin. The water that is contained in tissues was removed by
passing the tissues through increasing concentrations of ethanol (50-100%). Finally,
the alcohol that replaced the water was replaced by xylol. The specimens were
afterwards transferred in molten wax (60°C) in an embedding centre.
3.2.11.3 Embedding
The specimens were removed from their cassettes and placed in wax filled
molds according to the desired orientation (transversal and/or longitudinal). The
cassettes were then placed on top of the molds and the now called “block” allowed to
solidify on a cold surface.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 59
3.2.11.4 Sectioning
Sections of 5-7 μm were cut on a microtome (Mod. 1140/Autocut), flatted on
the surface of warm water and picked up onto polysine microscope slides.
3.3.11.5 Staining
Hematoxylin-eosin (HE) was used as the staining technique. Because staining
solutions used are aqueous, the wax had to be removed and the sections rehydrated.
The sections were first incubated at 60°C for 1h (Hereaus B5042 incubator) and were
afterwards passed in xylene and decreasing concentrations of ethanol as follows:
Xylol 1 15 min
Xylol 2 10 min
Xylol 3 10 min
100% EtOH 1 5 min
100% EtOH 2 5 min
96% EtOH 1 5 min
96% EtOH 2 5 min
70% EtOH 1 5 min
70% EtOH 2 5 min
Distilled water 5 min
The slides were immersed in different solutions as follows:
1. Mayer‟s hemalun 5 min
2. Tape water 5 min
3. Eosin 2 min
4. Distilled water Briefly
5. EtOH 70 % 2-3 min
6. EtOH 96 % 2-3 min
7. EtOH l 100 % 2-3 min
8. Xylol 2-3 min
9. Mounting with coverslips and mounting medium.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 60
3.2.12 Gene expression analysis
3.2.12.1 RNA preparation from cells and tissues
The total cytoplasmic RNA was extracted from cells and tissues using the
standard TRIzol® method according to the manufacturer‟s instructions (Invitrogen,
Life Technologies). For the animal experiment, tissue samples from the same
treatment group were pooled.
3.2.12.2 RNA concentration and quality control
RNA concentration was obtained by measuring the absorbance at 260nm
using a NanoDropTM 1000 spectrophotometer. The purity of the extracted RNA was
estimated by the ratio between absorbance at 260 and 280nm and its integrity
controlled by agarose gel electrophoresis and ethidium bromide staining.
3.2.12.3 Complementray DNA (cDNA) synthesis
Synthesis of cDNA including the removal of genomic DNA was performed
using the QuantiTect® Reverse Transcription Kit of Qiagen according to the provided
instructions. The incubation steps were performed in a GeneAmp PCR system 9600
thermocycler.
3.2.12.4 Primer design
The genomics sequences available at the University of California Santa Cruz
Genome Bioinformatics site (UCSC genome browser) and the Primer3 software
(Whitehead Institute for Biomedical Research, Cambrigde, USA) were used to design
primers (sequences in Table 9). The specificity of the designed primer was checked
using the BLAST program of the NCBI (National Centre for Biotechnology
Information). The design criteria were as follows:
Melting temperature: between 58°C and 60°C
Length: 18-23 bp
Amplicon length: 150-200 bp
G/C content: 40-60%

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 61
Table 9: Primer Sequences
Gene name Forward and reverse primers Amplicon size (bp)
Mouse
Cyclophilin (CYC) GGATTCATGTGCCAGGGTGG
212 CACATGCTTGCCATCCAGCC
ERα TGTTTGCTCCTAACTTGCTCCT
222 GGTGGATGTGGTCCTTCTCTT
Rat
1A CGTCACAGCCCATGCATTCG
212 CTGTTCATCCTGTTCCAGCTC
C3 CAGCCCGCAGAGTGCCAGTAGTC
CCATCCTCCTTTCCATCAACTGC
Clusterin CCCTTCTACTTCTGGATGAA
GAACAGTCCACAGACAAGAT
IGFBP-1 GATTAGCTGCCGCTCAACAG
283 TTCCCACTCCATGGGTAGAC
CaBP9k ATCCAAACCAGCTGTCCAAG
181 TGTCGGAGCTCCTTCTTCTG
Human
1A CTATAGTGGAGGCCGGAGCAGCAA
203 GAGGGGCGTTTGGTATTGGGTTATG
BAX AACATGGAGCTGCAGAGGA
GGAGGAAGTCCAATGTCCAG
BCL-2 GGATGCCTTTGTGGAACTGT
GCCTGCAGCTTTGTTTCAT
CCND1 TCGGTGTCCTACTTCAAATGTGT
233 GGGATGGTCTCCTTCATCTTAG
3.2.12.5 Quantitative realtime RT-PCR
Real-time RT-PCR was performed in a MX3005P thermal cycler. The protocol
consisted of a denaturing cycle at 94-95 °C for 4 min followed by 40 cycles of 20 s at

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 62
94-95 °C, 20-30 s at 58-60 °C, and 30 s at 72°C. The reaction setup and the cycling
conditions are resumed in Tables 10 and 11.
Table 10: PCR reaction mix.
Components 25 μL reaction 50 μL reaction Final []
Ampuwa 16.425 32.85 _
10X PCR buffer 2.5 5 1X
MgCl2 (50 mM) 1.5 3 3 mM
dNTPs (2.5 mM) 1 2 0.1 mM
Primer mix (10 mM each ) 0.5 1 0.2 mM
Template DNA 0.5 1 _
Taq-polymerase (5U/μL) 0.075 0.15 0.75 U
Sybr green (1X) 2.5 5 0.1X
Table 11: PCR cycling conditions.
Cycle Step Temperature Time Description
1 Segment 1 94-95°C 4 min Initial denaturation
40-45 Segment 2
94-95°C 20s Template denaturation, Primer annealing and Extension 58-60°C 20-30s
72 °C 30s
1 Segment 3
95°C 1 min Flourescnece data collection, dissociation (melting) curve analysis
55°C 30s
95°C 30s
3.2.12.6 Analysis of real time RT-PCR results
Expression levels of the analyzed genes were normalized to the housekeeping
gene (cytochrome C oxidase or 1A) and correlated to the untreated control animal
group using the ΔΔCt method (Pfaffl, 2001). This method assumes that the DNA
amplification efficiency is 100% or equal to 1 and proposes the following formula for
the calculation of the amount of target gene:
Expression level target gene (Fold induction) FI = 2 -∆∆Ct where:
Ct = cycle number at which the fluorescence crosses the threshold (threshold cycle)
ΔCt = Ct target-Ct reference
ΔΔCt = ΔCt target- ΔCt control

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 63
3.2.13 Cell cycle analysis
The analysis of cell cycle is based on the ability to stain cellular DNA in a
stoichiometric manner, i.e. the amount of stain is directly proportional to the amount
of DNA within the cell. Because the DNA amount changes between the different
phases during the cell cycle, the measure of DNA content in cells by flow cytometry is
a powerful tool that allows the study of how chemicals affect cell cycle distribution
and progression. For the assay, MCF-7 cells were routinely cultured in the normal
growth medium as indicated in Table 5. To avoid any influence of steroids contained
in the serum, FBS was stripped with 0.5% charcoal and the medium containing
charcoal-treated FBS (assay medium) used for the assay. The cells were plated at a
density of 2x105 cells/well in 6 -well plates and pre-incubated 48 h in assay medium
prior to the treatment. The procedure is represented in Figure 13. All the test
chemicals (see Table 12 for different groups) were dissolved in DMSO and the final
DMSO concentration in each well was not more than 0.1%. The cell cycle distribution
was analyzed using a BD FACSCalibur flow cytometer.
Table 12: Experimental groups for the cell cycle analysis.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 64
Figure13: Cell culture procedure for cell cycle analysis.

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 65
3.2.14 Immunocytochemistry
This method was applied for the detection of apoptosis of MCF-7 cells using
the M30 CytoDEATH antibody that recognizes a specific caspase cleavage site
within cytokeratin 18 (Roche, Mannheim Germany). MCF-7 cells were plated in 24-
well plates (5x104 cells/well), pre-incubated for 48 h with charcoal treated-FBS
medium and exposed for 6h to the test chemicals. The following protocol provided by
Roche was used with some modifications:
1. Washing of cells with PBS
2. 30 min fixation at -20°C in ice-cold pure methanol
3. 2 x 5 min wash with washing buffer (PBS+0.1%
Tween 20)
4.
Incubation with 100 µL/well M30 CytoDEATH antibody
(diluted 1/10 in washing buffer containing 1% BSA) at
4°C overnight
5. 2 x 5 min wash with washing buffer
6. Incubation with polyclonal goat anti-mouse Ig,
biotynylated (diluted 1/200 in PBS) for 1h at RT
7. 2 x 5 min wash with washing buffer
8. Incubation with streptavidin-cy3 (diluted 1/1000 in
PBS) for 1h at RT
9. 2 x 5 min wash with washing buffer
10. Incubation with Dapi (1/1000 in PBS) for 3 min at RT
11. 2 x 5 min wash with washing buffer
12. Acquisition of photos using fluorescence microscope
Photos were taken and analyzed with an Axiovert 200 M inverted fluorescence
microscope equipped with an HL 100 camera. At least 1000 cells/treatment groups
were counted and the apoptotic index calculated according to the following formula:
100cellsof number Total
cells positive M30of Number index Apoptotic

Material and methods
M.Sc. Sadrine Tchoukouegno Ngueu 66
3.2.15 Statistical analysis
All data are expressed as mean±SD and were statistically analyzed with SPSS
17.0-21.0 for Windows. Kruskal-Wallis H-test and the Mann–Whitney U-test were
used and significance level set at P < 0.05.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 67
CHAPTER 4: RESULTS

Results
M.Sc. Sadrine Tchoukouegno Ngueu 68
4.1 Biological activities and chemical characterization of Erythrina
excelsa
4.1.1 Estrogenic and Cytotoxic properties of E.excela whole extract 1
4.1.1.1 Yeast estrogen screen
The ability of E. excelsa whole ethanol extract to interact with the human
estrogen receptor alpha was assessed using the yeast transactivation assay. As
shown in Figure 14A, E. excelsa similarly to E2 and genistein dose-dependently
induced lac-Z reporter gene expression. Yeast growth was inhibited by
concentrations of extract above 10 μg/mL (data not shown). The EC50 values as well
as RIEs and RPs are summarized in Table 13. The response of the yeast to E.
excelsa was antagonized by the pure anti-estrogen ZK (Figure 14B). In the anti-
estrogenic activity assay, only ZK was able to produce a full dose-response effect
(Figure 14C).
1 These results are submitted as original research article in planta medica
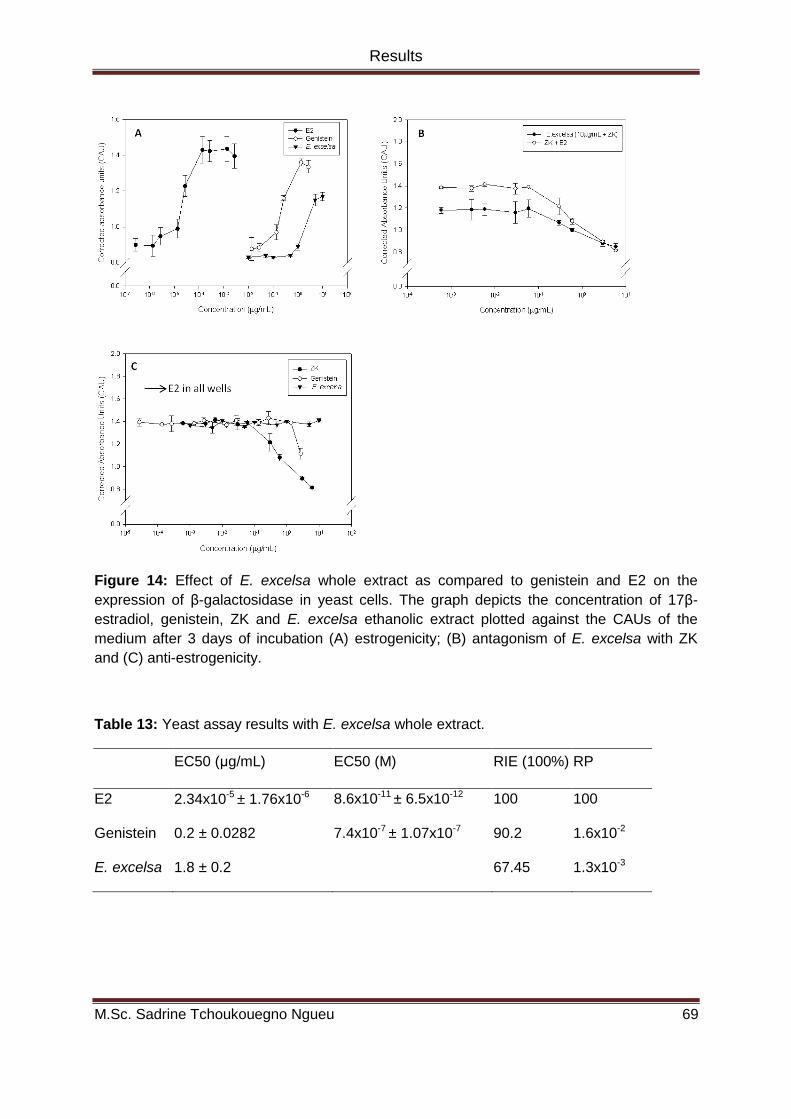
Results
M.Sc. Sadrine Tchoukouegno Ngueu 69
Figure 14: Effect of E. excelsa whole extract as compared to genistein and E2 on the
expression of β-galactosidase in yeast cells. The graph depicts the concentration of 17β-
estradiol, genistein, ZK and E. excelsa ethanolic extract plotted against the CAUs of the
medium after 3 days of incubation (A) estrogenicity; (B) antagonism of E. excelsa with ZK
and (C) anti-estrogenicity.
Table 13: Yeast assay results with E. excelsa whole extract.
EC50 (μg/mL) EC50 (M) RIE (100%) RP
E2 2.34x10-5 ± 1.76x10-6 8.6x10-11 ± 6.5x10-12 100 100
Genistein 0.2 ± 0.0282 7.4x10-7 ± 1.07x10-7 90.2 1.6x10-2
E. excelsa 1.8 ± 0.2 67.45 1.3x10-3

Results
M.Sc. Sadrine Tchoukouegno Ngueu 70
4.1.1.2 Uterotrophic assay
4.1.1.2.1 Effect of E. excelsa whole extract on the uterus
Uterine wet weight and epithelial height
E. excelsa at a dose of 50 mg/kg BW/day like E2 led to a significant increase
of uterine wet weight and uterine epithelial height but was less potent. At 100 mg/kg
BW/day, a significant decrease of uterine wet weight and a tendency to a decrease of
the uterine epithelial height were observed (Figure 15).

Results
M.Sc. Sadrine Tchoukouegno Ngueu 71
Figure 15: (A) Uterine wet weight (mg/kg BW) and (B) uterine epithelial height (µm) after 3-
day subcutaneous treatment of ovariectomized Wistar rats with either E. excelsa ethanolic
extract (50 and 100 mg/kg BW/day) or E2 (4 µg/kg/BW/day). (C) 100X Microphotographs of
uterine tissue following a Hematoxylin-Eosin (HE) staining. Mean values ± S.D. are given for
n = 6 animals per dose group ** p≤ 0.01;*p≤0.05, significantly different from the control group
(Ovex), Kruskal-Wallis H-test followed by Mann–Whitney U-test.
Uterine C3 and clusterin mRNA expression
As expected, Clusterin mRNA expression was down-regulated and C3 up-
regulated following treatment with E2. In response to the treatment with 50 mg/kg
BW/day of E. excelsa ethanolic extract, the same but less pronounced gene
expression pattern was detected. The 100 mg/kg dose of E. excelsa, unlike the 50
mg dose, caused a significant up-regulation of Clusterin and a down-regulation of
C3 (Figure 16).

Results
M.Sc. Sadrine Tchoukouegno Ngueu 72
Figure 16: (A) Relative mRNA expression of C3 and (B) clusterin (normalized to the
mRNA amount of 1A and compared to the vehicle control group) in the uterus of
ovariectomized Wistar rats treated subcutaneously for 3 consecutive days with 50 and
100 mg/kg BW/day E. excelsa ethanolic extract and 4 µg/kg/BW/day E2. * P≤0.05
significantly different from the control group (Ovex), Kruskal-Wallis H-test followed by
Mann-Withney U-test.
4.1.1.2.2 Effect of E. excelsa whole extract on the vagina
E2 increased the vaginal epithelial height by approximately 5-fold. No
significant effects of E. excelsa could be detected at both doses, but clear tendencies
of either increase or decrease were seen (Figure 17).

Results
M.Sc. Sadrine Tchoukouegno Ngueu 73
Figure 17: (A) Vaginal epithelial height (µm) after 3-day subcutaneous treatment of
ovariectomized Wistar rats with either E. excelsa ethanolic extract (50 and 100 mg/kg
BW/day) or E2 (4 µg/kg/BW/day). (B) 100X Microphotographs of vagina tissue following a
Hematoxylin-Eosin (HE) staining. Mean values ± S.D. are given for n = 6 animals per dose
group ** p≤ 0.01;*p≤0.05, significantly different from the control group (Ovex), Kruskal-Wallis
H-test followed by Mann–Whitney U-test.
4.1.1.2.3 Effect of E. excelsa whole extract on the liver IGFBP-1 and CaBP9k
mRNA expression
The 50 mg/kg dose of E. excelsa similar to E2 significantly increased the
mRNA expression of IGFBP-1 and CaBP9k, whereas the 100 mg/kg dose led to a
significant down regulation of the two genes (Figure 18).

Results
M.Sc. Sadrine Tchoukouegno Ngueu 74
Figure 18: Relative mRNA expression of (A) IGFBP-1 and (B) CaBP9k (normalized to the
mRNA amount of 1A and compared to the vehicle control group) in the liver of
ovariectomized Wistar rats after 3-day subcutaneous administration of 50 and 100 mg/kg
BW/day E. excelsa ethanolic extract and 4 µg/kg/BW/day E2. *P≤0.05, Kruskal-Wallis H-test
followed by Mann-Withney U-test.
4.1.1.3 MTT assay
The effect of E. excelsa whole extract on the growth of breast (MCF-7) and
colon (HT-29) cancer cell lines was investigated using the MTT cytotoxicity/cell
viability assay. A slight and non significant increase of the viability of MCF-7 cells was
caused by lower concentrations of E. excelsa. At higher concentrations however, cell
death was observed. The extract was cytotoxic on HT-29 cells at all tested
concentrations and was less potent than the chemotherapeutic drug cisplatin (Figure
19).

Results
M.Sc. Sadrine Tchoukouegno Ngueu 75
Figure 19: Effects of E. excelsa whole extract on the growth of (A) MCF-7 and (B) HT-29
cells. Results present the average and standard deviation of n≥3 independent experiments
performed in 6 replicates for each concentration.
4.1.2 Effect of a phytoestrogenic fraction of E. excelsa on the growth of the
human breast cancer line MCF-7.
Here, the above mentioned crude ethanol extract was fractionated by vacuum
liquid chromatography and the fractions monitored for both estrogenic (YES) and
cancer growth inhibitory (XTT) properties.
4.1.2.1 Yeast estrogen screen
From the 5 fractions obtained by VLC, F4 and F5 were able to induce the
transactivation of the hERα as depicted on Figure 20. Based on the calculated EC50
values (Table 14) F4 was almost as potent as the whole extract and 13.47 times
more potent than F5.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 76
Figure 20: Effect of fractions of E. excelsa on the expression of β-galactosidase in yeast
cells. (A) VLC fractions, (B) active fractions compared to the whole extract. The graphs
depict the concentration of test chemicals plotted against the CAUs of the medium after 3
days of incubation.
Table 14: Yeast assay results with VLC fractions.
Substance EC 50(μg/ml) RIE (%) RP
E2 5.08 x 10-5
± 4.64 x10-6
100
F1 - -
F2 - -
F3 - -
F4 1.1 ± 0.0174 70.37 4.62 x 10-3
F5 14.8 ± 6.7 35.06 3.43 x 10-3
4.1.2.2 XTT
To test whether the two estrogenic fractions also conserved the cytotoxic
ability exhibited by the whole extract, their effect on MCF-7 cell growth was assessed
using the XTT assay. To minimize the influence of the steroids presents in FBS, the
assay was performed with a medium supplemented with charcoal stripped FBS. As
shown in Figure 21, both fractions at lower concentrations induced a slight but not
significant increase in cell viability. At higher concentrations, F4 induced a

Results
M.Sc. Sadrine Tchoukouegno Ngueu 77
concentration-dependent decrease of cell number but was less cytotoxic than the
whole extract.
Figure 21: (A) Effect of estrogenic fractions of E. excelsa ethanol extract on MCF-7 cell„s
viability. (B) F4 compared to the whole extract. MCF-7 cells were incubated for 72 h with
increasing concentrations of fractions or whole extract and the cell viability examined by
XTT.* (p≤0.05) and *** (p<0.001) vs. control analyzed by Kruskal Wallis H-test followed by
Mann Whitney U-test.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 78
4.1.2.3 HPLC fingerprints of F4 and E. excelsa whole extract
Figure 22: HPLC chromatograms of E. excelsa whole extract and F4 fraction
Based on the results obtained in YES and XTT, F4 was the most potent
fraction with respect to both estrogenicity and cytotoxicity and was chosen for further
analysis.
4.1.2.4 Effect of F4 on MCF-7 cell cycle distribution
As on cell viability, a biphasic effect of F4 on cell cycle distribution was
observed. Similarly to E2, F4 at lower concentrations stimulated the G1/S transition
leading to an increased percentage of cells in S and G2/M phases. At the same time,
a concentration-dependent decrease of the percentage of subG1 population (death
cells) was observed. With higher concentrations of F4, although not significant, a
concentration-dependent increase of the percentage of cells in G0/G1 was observed
followed by a decrease of the percentage of cells in S but not in G2/M phase. Higher
concentrations of F4 also increased the percentage of the subG1 population (Figure
23).

M. Sc. Sad
rine T
cho
uk
ou
egno
Ngu
eu
Page 7
9
Resu
lts
Figure 23: Effects of F4 on the cycle distribution of MCF-7 cells. MCF-7 cells were incubated for 48 h with increasing concentrations of F4 and
the cell cycle distribution assessed by flow cytometry. (A) Representative Histogramms and (B) mean of n≥ 3 independent experiments. a
(p≤0.05), b (p<0.01), c (p<0.001) vs. control analyzed by Kruskal Wallis H-test followed by Mann Whitney U-test.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 80
4.1.2.4.1 Antagonisation with ZK
To elucidate the role of ER in the estrogen-like effects of lower concentrations
of F4, an antagonization experiment with a pure antiestrogen (ZK) was performed.
Treatment with ZK alone led to G1 arrest (indicated by a significant increase of the
percentage of the cells in G1 with a concomitant significant decrease in the
percentage of S and G2/M populations) and to an increase of cell death. ZK
antagonized the effects of E2 as well as F4 on almost all phases of the cell cycle
(Figure 24).
Figure 24: Role of ER in F4-mediated effects on the cycle distribution of MCF-cells. MCF-7
cells were cotreated for 48 h with F4 and the pure anti-estrogen ZK and the cell cycle
analyzed by flow cytometry. * (p≤0.05), ** (p<0.01), *** (p<0.001) vs. Control; + (p≤0.05), ++
(p<0.01), +++ (p<0.001) vs. single treatment analyzed by Kruskal Wallis H-test followed by
Mann Whitney U-test.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 81
4.1.2.4.2 Cotreatment with E2
To test for ER antagonism activity of F4, MCF-7 cells were simultaneously
treated with increasing concentrations of F4 and 10-10 M E2 (Figure 25). Analysis of
the cell cycle following cotreatment with lower concentrations of F4 and 0.1 nM E2
shows some antagonistic patterns on both SubG1 and S phases. In fact single
treatment with F4 at those concentrations or with E2 decreased the percentage of the
subG1 cell population and cotreatment brought this percentage back to control levels.
The same effect was observed on the percentage of S-phase cells with 1 μg/mL F4.
In contrast to subG1 and S, additive effects were observed on Go/G1 and M phases.
Cotreatment with E2 did not rescue the cells from death induced by higher
concentrations of F4 rather, E2 addition led to even more pronounced cell death. On
the other hand E2 addition induced more cells to transit from G1 to S phase.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 82
Figure 25: Effect of the cotreatment with E2 and lower (A) and higher (B) concentrations of
F4 on the cycle distribution of MCF-7 cells. MCF-7 cells were treated for 48h with various
concentrations of F4 together with 0.1 nM E2 and the cell cycle distribution analyzed by flow
cytometry. * (p≤0.05), ** (p<0.01), *** (p<0.001) vs. Control; + (p≤0.05), ++ (p<0.01), +++
(p<0.001) vs. single treatment analyzed by Kruskal Wallis H-test followed by Mann Whitney
U-test.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 83
4.1.2.5 M30 staining
To assess the cell death process sustaining the increase of subG1 population
induced by higher concentrations of F4, the caspase cleaved cytokeratin 18 that is a
marker of early apoptosis was investigated. Figure 26 shows clear tendency of a
concentration-dependent increase of the percentage of M30 positive cells following
treatment with F4.
Figure 26: Effect of F4 on caspase-cleaved cytokeratin 18 in MCF-7 cells. MCF-cells were
exposed for 6h to either ZK (10-6
M) or increasing concentrations of F4 and the protein
expression of caspase-cleaved cytokeratin 18 measured by indirect immunofluorescence
using the M30 CytoDEATH antibody as described in material and methods. (A)
Representative fluorescence microscopy images (100 X magnitude), (B) quantitative analysis
of M30 positive cells.
4.1.2.6 Effect of F4 on the mRNA expression of CCND1, Bax and Bcl-2 in MCF-7
cells
CCND1 mRNA was upregulated following treatment with E2 and 10 μg/mL F4,
and downregulated by ZK and 50 μg/mL of F4. E2 and F4 at low concentration acted
differently on Bax and Bcl-2 expression but both significantly downregulated the
Bax/Bcl-2 ratio. E2 had no effect on Bax and upregulated Bcl-2 whereas F4 had no
effect on Bcl-2 but downregulated Bax. At high concentration, F4 as well as ZK
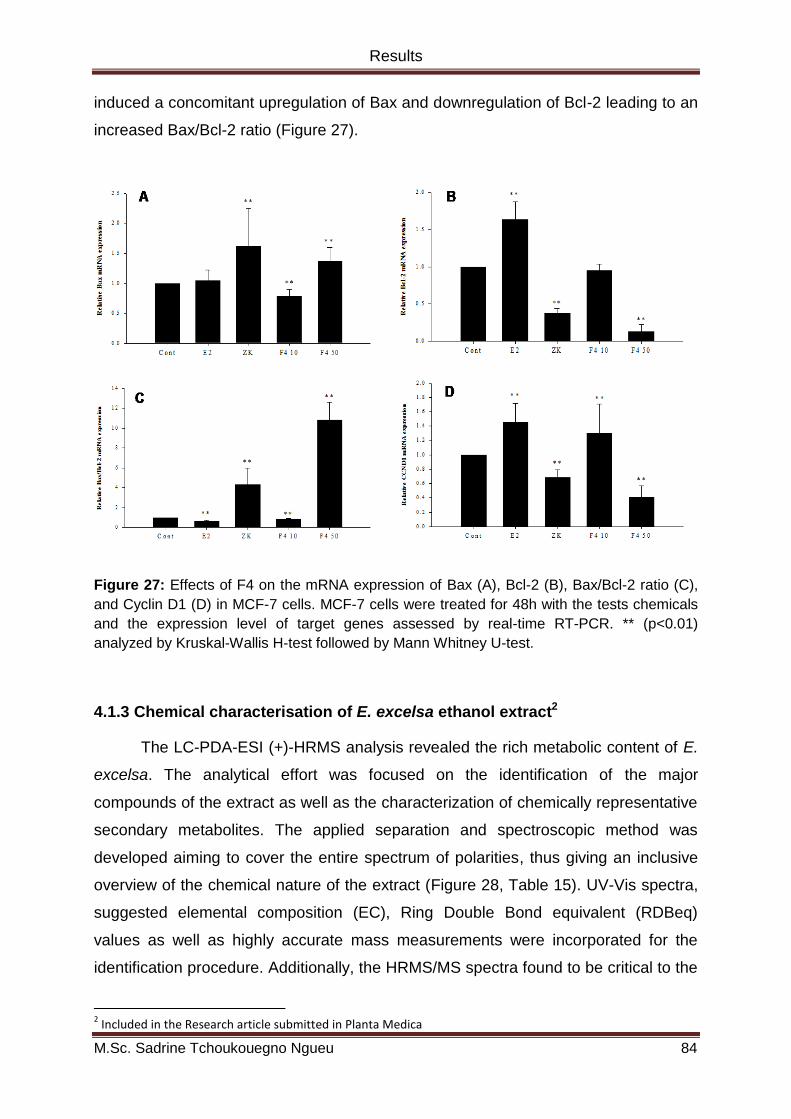
Results
M.Sc. Sadrine Tchoukouegno Ngueu 84
induced a concomitant upregulation of Bax and downregulation of Bcl-2 leading to an
increased Bax/Bcl-2 ratio (Figure 27).
Figure 27: Effects of F4 on the mRNA expression of Bax (A), Bcl-2 (B), Bax/Bcl-2 ratio (C),
and Cyclin D1 (D) in MCF-7 cells. MCF-7 cells were treated for 48h with the tests chemicals
and the expression level of target genes assessed by real-time RT-PCR. ** (p<0.01)
analyzed by Kruskal-Wallis H-test followed by Mann Whitney U-test.
4.1.3 Chemical characterisation of E. excelsa ethanol extract2
The LC-PDA-ESI (+)-HRMS analysis revealed the rich metabolic content of E.
excelsa. The analytical effort was focused on the identification of the major
compounds of the extract as well as the characterization of chemically representative
secondary metabolites. The applied separation and spectroscopic method was
developed aiming to cover the entire spectrum of polarities, thus giving an inclusive
overview of the chemical nature of the extract (Figure 28, Table 15). UV-Vis spectra,
suggested elemental composition (EC), Ring Double Bond equivalent (RDBeq)
values as well as highly accurate mass measurements were incorporated for the
identification procedure. Additionally, the HRMS/MS spectra found to be critical to the
2 Included in the Research article submitted in Planta Medica

Results
M.Sc. Sadrine Tchoukouegno Ngueu 85
identification procedure supporting the structural information were obtained from the
HRMS analysis in full scan (Figures 29-36).
Figure 28: Base Peak chromatogram of Erythrina excelsa ethanol extract and Extracted Ion
Chromatograms (XICs) of representative isoflavonoids detected.

M. Sc. Sad
rine T
cho
uk
ou
egno
Ngu
eu
Page 8
6
Resu
lts
Table 15: Representative Isoflavonoids of Erythrina excelsa ethanol extract using LC-PDA-ESI (+)-LTQ-Orbitrap platform.
Rt
(min)
[M+H]+ Experimental
m/z
Theoritical
m/z
Δm(mmu/ppm)/
ppm)
RDB λmax
(nm)
Main MS / MS fragment ions (Relative abundance, %) Chemicalclass
(Identification) 1 7.91 C15H11O4 255.0663 255.0652 1.125 / 4.409 10.5 261 255 (100), 237 (18), 227 (43), 211 (4), 199 (85), 137 (43)
Simple isoflavone
(Daidzein)
2 9.53 C15H9O5 269.0435 269.0444 -0.900 / -3.345 11.5 262, 342 Not found Coumestan (coumestrol)
3 11.43 C20H19O6 355.1187 355.1176 1.095 / 3.084 11.5 267 337 (100), 319 (5), 309 (8), 295 (37), 283 (77), 271 (2) Prenyl isoflavonoid
4 12.11 C25H27O7 439.1761 439.1751 0.960 / 2.255 12.5 234, 270 421 (11), 403 (49), 385 (9), 368 (24), 349 (100), 295 (8),
213 (6) Dipyran isoflavonoid
5 14.83 C20H9O5 339.1234 339.1227 0.710 / 2.093 11.5 228, 267 283 (100) Prenyl isoflavonoid
6 16.12 C25H25O6 421.1651 421.1646 0.535 / 1.270 13.5 232, 280 403 (100), 349 (10) Dipyran isoflavonoid
7 16.69 C20H17O5 337.1076 337.1071 0.520 / 1.542 12.5 228, 282 337 (8), 322 (19), 319 (69), 309 (15), 295 (31), 283 (100),
267 (15) Pyran isoflavonoid
8 17.22 C25H27O6 423.1803 423.1802 0.085 / 0.201 12.5 237, 271 405 (42), 367 (100), 351 (3), 349 (39), 295 (2) Pyran prenyl
isoflavonoid
9 18.56 C25H27O5 407.1851 407.1853 -0.180 / -0.443 12 .5 239, 271 351 (100), 295 (10) Diprenyl isoflavonoid

Results
M.Sc. Sadrine Tchoukouegno Ngueu 87
The great majority of the detected secondary metabolites were isoflavonoids.
For instance compound 1 (Rt = 7.91 min) was identified as daidzein based on its
accurately measured pseudomolecular ion [M+H]+ at m/z 255.0663 with an RDBeq.
value of 10.5 (positive mode). HRMS/MS spectra verified its structure through the
characteristic [M+H-CO]+, [M+H-CO-CO]+ and 1,3A+ ions (Figure 29) (Lee et al.,
2005).
Figure 29: HRMS/MS spectrum of compound 1.
Coumestans were also detected. For example, the well-known phytoestrogen
coumestrol (compound 2, Rt = 9.23 min) was identified by its pseudomolecular ion at
m/z 269.0435 and its characteristic UV spectra as well as the RDBeq. value of 12.5
(positive mode) (Antignac et al., 2003).
Except isoflavones and coumestans, several mono- and di-pyran and prenyl
derivatives of isoflavonoids seem to dominate the extract and thus comprising its
most characteristic chemical group. Some representative prenylated and pyran
isoflavonoid derivatives are listed in Table 15. Similarly to the above mentioned
compounds, they were identified from their accurately measured spectrometric data
and additional spectrometric features while fragmentation patterns mostly related to
specific mass losses at their HRMS/MS spectra supported drastically their

Results
M.Sc. Sadrine Tchoukouegno Ngueu 88
identification. In particular, the nominal mass losses of 56 Da (C4H8) characteristic of
prenyl group and 54 Da (C4H6) for pyran ring were widely observed at the HRMS/MS
spectra of compounds 3-9 (Simons et al., 2009). Specifically, at the HRMS/MS
spectra of compounds 3 (Figure 30) (Rt = 11.43 min), 5 (Figure 31) (Rt = 14.83 min)
and 9 (Figure 32) (Rt = 19.56 min), the typical loss of 56 Da (C4H8) is observed
indicating the occurrence of at least one prenyl side chain at the basic skeleton of
these isoflavonoids (Zhang et al., 2008; Simons et al., 2009). Particularly for
compound 9, two successive losses of 56 Da were observed from its
pseudomolecular ion. Based on this observation as well as the suggested molecular
formula and the RDBeq value, diprenylation could be proposed.
Figure 30: HRMS/MS spectrum of compound 3.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 89
Figure 31: HRMS/MS spectrum of compound 5.
Figure 32: HRMS/MS spectrum of compound 9.
In contrast, at the HRMS/MS spectra of compounds 4 (Figure 33) (Rt = 12.11
min), 6 (Figure 34) (Rt = 16.12 min) and 7 (Figure 35) (Rt = 16.69 min), the
characteristic loss of 54 Da is observed indicating the presence of a pyran ring
(Takayama and Nomura, 1995). Moreover, similarly to compound 9, a successive

Results
M.Sc. Sadrine Tchoukouegno Ngueu 90
loss of 54 Da was observed at the HRMS/MS spectra of compound 4 indicating
likewise the presence of two pyran rings on the basic isoflavonoid skeleton.
Figure 33: HRMS/MS spectrum of compound 4.
Figure 34: HRMS/MS spectrum of compound 6.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 91
Figure 35: HRMS/MS spectrum of compound 7.
Finally, compound 8 (Figure 36) exhibited both type of mass losses indicating
probably the concurrent presence of prenyl substitution and pyran ring.
Figure 36: HRMS/MS spectrum of compound 8.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 92
4.2 Effects of estrogens and chemicals of plant origin on skeletal
muscle growth
4.2.1 E2, genistein, and Ecdysone induce hypertrophy of C2C12 murine skeletal
muscle cells
Differentiated C2C12 cells were used as a model to evaluate the effect of
estrogens, PEs and the phytoecdysteroid ecdysterone on skeletal muscle growth. As
shown in Figure 37 all the chemicals except the whole extract and fraction F4 were
able to significantly increase the diameter of C2C12 myotubes.
Figure 37: Effect of E2, PEs, and the phytoecdysteroid ecdysone on C2C12 myotubes
hypertrophy. 4-6 days old myotubes incubated for 48 h with test chemicals were fixed and
photographied by glutaraldehyde induced autofluorescence. Depicted are mean of n≥2
independent experiments. * p≤0.05 vs. Control by Kruskal Wallis H-test and Mann Withney
U-test.
4.2.2 Role of ER and AR on ecdysone induced myotube hypertrophy
To get more insights on the mode of action of ecdysterone, further
experiments were performed with both AR (Flutamide) and ER (ZK) antagonists. ZK
but not flutamide was able to block to hypertrophic response of C2C12 myotubes to
ecdysterone treatment (Figure 38).

Results
M.Sc. Sadrine Tchoukouegno Ngueu 93
Figure 38: Role of AR (A) and ER (B) on ecdysterone induced C2C12 myotubes
hypertrophy. 4-6 days old myotubes incubated for 48 h with test chemicals were fixed and
photographied by glutaraldehyde induced autofluorescence. Represented are mean± SD of
n=2 independent experiments. * p≤0.05 vs. Control, + p≤0.05 vs. Single treated Kruskal
Wallis H-test and Mann Withney U-test.
4.2.3 Role of ERα and ERβ in skeletal muscle growth
ERs subtype selective agonists (ALPHA and BETA) were used to specify the
implication of each ER isoform on the hypertrophic response of C2C12 to estrogens
and estrogen-like chemicals. The dose-response effect of both agonists is
represented in Figure 39. The increase of mytobute diameter was positively
correlated with higher concentrations of ALPHA and lower concentrations of BETA.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 94
Figure 39: Effect of ER subtype selective agonists on C2C12 myotubes hypertrophy. 4-6
days old myotubes incubated for 48 h with test chemicals were fixed and photographied by
glutaraldehyde induced autofluorescence. Depicted are mean of n=3 independent
experiments.
At a concentration where both agonists selectively bind to their specific
receptors, BETA but not ALPHA was able to induce myotube hypertrophy (Figure
40).

M. Sc. Sad
rine T
cho
uk
ou
egno
Ngu
eu
Page 9
5
Resu
lts
Figure 40: Effect of ER subtype selective agonists on C2C12 myotubes hypertrophy. 4-6 days old myotubes incubated for 48 h with test
chemicals were fixed and photographied by glutaraldehyde induced autofluorescence. Depicted are mean of n=3 independent experiments. *
p≤0.05 vs. Control by Kruskal Wallis H- and Mann Withney U-tests.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 96
Moreover, in the presence of an ERβ selective antagonist the hypertrophic response
to E2 and Ecdy was reduced (Figure 41) confirming therefore an ERβ-mediated
effect.
Figure 41: Ecdysterone induced myotube hypertrophy is ERβ-mediated. 4-6 days old
myotubes incubated for 48 h with test chemicals were fixed and photographied by
glutaraldehyde induced autofluorescence. * p≤0.05 vs. Control, + p≤0.05 vs. Single treated
Kruskal Wallis H-test and Mann Withney U-test.
4.2.4 ERα and ERβ expression in C2C12 myotubes
Both ERs are expressed in differentiated C2C12 cells as shown below (Figure
42). Expression of ERα was shown at the mRNA level and ERβ at the protein level.

Results
M.Sc. Sadrine Tchoukouegno Ngueu 97
Figure 42: ERα mRNA expression (A) and ERβ protein expression (B) in C2C12 myotubes.

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 98
CHAPTER 5: DISCUSSION

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 99
The aims of this thesis were to evaluate the biological activities that may
explain the use of the extract of E. excelsa to alleviate menopausal symptoms and to
treat female gynecological tumors and to characterize its main chemical constituents;
and to elucidate the mechanisms underlying the anabolic effect of ecdysterone on
mammalian skeletal muscle. The major findings that emerged are:
1. E. excelsa exhibits both estrogenic and antiestrogenic effects on several
estrogens responsive endpoints.
2. A phytoestrogenic fraction of E. excelsa at low and maybe
physiologically relevant concentrations stimulates the growth of MCF-7
breast cancer cell line in an E2-free environment but exerts antagonistic
effects in the presence of E2.
3. Prenyl- and pyran-substituted isoflavonoids are the main constituents
of E. excelsa extract.
4. ERβ is involved in Ecdysterone-mediated skeletal muscle growth.
5.1 Biological activities of Erythrina excelsa
5.1.1 Estrogenic/anti-estrogenic and cytotoxic properties of E. excelsa
Based on the ethnomedical use of E. excelsa to alleviate menopausal
complaints and to treat tumors in the female genital tract, we hypothetized that the
extract may possess estrogenic and cytotoxic properties. The yeast estrogen screen
(YES) and the uterotrophic assay were used as tests for estrogenicity and the MTT
assay as test for cytototoxic effects.
Yeast cells transfected with ERs function as steroid-dependent transcriptional
activators and are therefore a suitable tool used to measure the ability of a substance
to activate the transactivation of the receptor. To assess the functionality of our
assay, the EC50 of E2 was determined and was 86 pM, a value that is comparable to
the median EC50 value of 100 pM in mammalian cells based transcriptional
activation assays (The National Toxicology Program (NTP), 2002). Genistein was

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 100
used as reference phytoestrogen and its calculated EC50, RP and RIE were in line
with published values (Dhooge et al., 2006). E. excelsa was able to induce a
concentration-dependent transactivation of ER with a RP of 1.3 x 10-3 which is in the
range of other RPs reported for estrogenic plants extracts in the YES (Lee et al.,
2005; Zhang et al., 2005). The dose-dependent transactivation of ER by E. excelsa
was antagonized by a pure anti-estrogen (Figure 14B) demonstrating an ER-
mediated effect. It is worth noting that the calculated potency of the extract was only
12.3 times lower than that of the pure phytoestrogen genistein suggesting the
presence of potent estrogenic secondary metabolites in the extract. This was later
confirmed by LC-HRMS and LC-HRMS/MS analysis which showed that E. excelsa
whole ethanol extract contains isoflavonoids (Figure 28) known to exhibit estrogenic
activity in several test systems.
Because in vitro test systems are usually specific for one mechanism of action
and cannot totally predict what may happen in an organism, we also used the 3-day
uterotrophic assay with adult OVX rats according to the OECD guidelines (Owens
and Ashby, 2002; OECD, 2007) to test the estrogenic activity of E. excelsa. This
assay is an established model for studying the effect of estrogen and estrogen-like
substances on estrogen-responsive tissues. Uterine weight is the major endpoint
measured in the uterotrophic assay as recommended by OECD. However, because
several PEs and endocrine disruptor substances have been shown to exhibit
estrogenic effects not visible on the uterine weight, other endpoints (uterine and
vaginal epithelial heights and uterine and liver gene expression analysis) were also
included in the investigation.
Treatment of OVX rats with E.excelsa led to mixed estrogenic and anti-
estrogenic effects on uterine morphometric parameters (Figure 15).The same profile
was observed at the molecular level on uterine (Figure 16) and liver (Figure 18) gene
expression. Up-regulation of C3 and down-regulation of clusterin are highly sensitive
molecular markers to detect estrogenicity in the uterus (Diel et al., 1995). Analysis of
the uterine expression of these genes in E. excelsa-treated animals indicates a
strong agonistic activity after treatment with the 50 mg/kg dose, whereas treatment
with the 100 mg/kg dose resulted in an expression pattern indicative of anti-
estrogenic activity. The liver that is the central organ in metabolic pathways is also a
non-classical target of estrogen and estrogen-like substances (Geis et al., 2005). The

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 101
gene expression of IGFBP-1 and CaBP9k in the liver has been shown to be
upregulated following estradiol administration (Diel et al., 1995; Geis et al., 2005; Lee
et al., 2005). Here, like in the uterus, E. excelsa exhibited a pattern of activity
indicative of mixed estrogenic and anti-estrogenic effects. E. excelsa extract contains
ISO that are known to exhibit hormetic dose-responses on many physiological
endpoints. Hormesis is defined as the phenomenon by which chemicals are able to
induce biologically opposite effects at different doses (Siow and Mann, 2010; Vargas
and Burd, 2010). It is defined in toxicology as a dose-response phenomenon that is
characterized by low dose stimulation, high dose inhibition, resulting in either a J-
shaped or an inverted U-shaped dose-response. Both estrogenic and anti-estrogenic
effects of ISO depending on the dose have been reported (Patisaul and Jefferson,
2010; Pilsakova et al., 2010; Zhao and Mu, 2010). Moreover, according to the MS-
based investigation, pyrano as well as prenylated isoflavonoids comprise the main
chemical classes of secondary metabolites of the extract. There are several
published data investigating the role of prenyl chains and pyran rings in the
estrogenicity of isoflavonoids (Halabalaki et al., 2006; Okamoto, 2006; Djiogue et al.,
2010; Simons et al., 2012). Depending on their position on the isoflavonoids skeleton,
these substitutions have been shown to modify the biological activity of the parent
molecule. Djiogue et al (2010) showed that prenylation of genistein at various
positions was associated with either increase or decrease of its estrogenic
activity/potency, and with a modification of its ER selectivity. Okamoto et al (2006)
and Simons et al (2012) on the other hand showed that prenylation at certain
positions of the isoflavone skeleton was associated with anti-estrogenic activity.
Okamoto et al (2006) showed that four structural features among which the presence
of prenyl groups at the 6- and 8-positions of the A-ring of the isoflavone structure are
essential for the anti-estrogenicity of isoflavones. Tanee Fomum et al. (1986) isolated
a 5, 7, 4'-trihydroxy-6, 8-diprenylisoflavone from the stem bark of E. excelsagrown in
Cameroon. We therefore suggest that the diprenyl isoflavonoid identified in the
ethanol extract by LC-MS analysis may correspond to the same compound and may
also account for the anti-estrogenic effect observed.
Although the estrogenic response is predominantly mediated by the same ER
subtype in uterus and vagina, E. excelsa elicited significant estrogenic effects on the
uterus and not on the vagina. This selectivity of action may be explained by different

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 102
ratio of coactivators to corepressors proteins or by the fact that the extract may
induce a different conformation of the ER in both organs. Some authors have
proposed that phytoestrogens mainly isoflavones due to their mixed agonist
/antagonistic effects on ER targets may be considered as natural selective estrogen
receptor modulators (SERMS). For instance, Mersereau et al (2008) showed that
liquiritigenin, a plant derived phytoestrogen exhibits similar binding affinities for ERα
and ERβ but induces selective transcriptional change of the ERβ only. To explain this
difference in transcriptional regulation, they demonstrated that liquiritigenin only
recruits coactivators to ERβ. Moreover, Shanle and Xu (2011) stated that given the
importance of coactivators in stimulating transcription and their potential role in
stabilizing ligand-bound ER in an active conformation, the estrogenic effects of
potential endocrine disruptors‟ compounds will be highly dependent on the availability
of coactivators which can vary among cell types, thereby contributing to tissue
specific effects.
The effect of E. excelsa on both estrogen-dependent and estrogen-
independent cancer cell lines was investigated using the MTT assay. E. excelsa
induced a slight increase of the viability of the ER positive MCF-7 cells. This effect
that may be correlated to an ERα- mediated cell proliferation since we showed in the
YES that the extract is able to induce the transactivation of ERα. At higher
concentrations, E. excelsa was cytotoxic on MCF-7 cells, a result that is in agreement
with the general knowledge on PEs action. It has been shown that this cytotoxic
effect may be ER-independent as demonstrated here when considering that the
extract was also cytotoxic on HT-29 cells which are ERα-negative (Hartman et al.,
2009; Edvardsson et al., 2011). There are some evidences suggesting that ERα-
negative cells may have different responses to phytoestrogen exposure. Van der
Woude et al (2005) demonstrated that the phytoestrogen quercetin at low
concentrations dose-dependently promoted the growth of the ER-positive MCF-7
cells and inhibited proliferation at high concentrations. They also showed that
quercetin in the same concentration range caused a significant inhibition of the
proliferation of MDA-MB-231 ERα-negative cells. Similarly, genistein was shown to
increase the growth of estrogen-sensitive cells at low concentrations but decreased
cell growth at high concentrations (Seo et al., 2006). On HT-29 cells, genistein has

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 103
been shown to have no effect on the proliferation at low concentration and to inhibit
cell growth at high concentration (Arai et al., 2000).
In summary our data demonstrated that E. excelsa exhibits several biological
activities (estrogenic, antiestrogenic, and cytotoxic) that may explain its
aforementioned ethnomedical use for women‟s health problems. We also showed
that prenyl- and pyran-substituted isoflavonoids were the main constituents of the
extract and may explain the variety of biological activities observed since prenyl
substitution for example, has been shown to improve interaction with membranes and
proteins enlarging therefore the range of biological activities.
5.1.2 Effect of a phytoestrogenic faction of E. excelsa on the growth of the
human breast cancer line MCF-7.
Breast cancer is the most common cancer in women with about 60-70% of
cases classified as ER positive and dependent on estrogens for growth (Mohibi et al.,
2011). Initial results of this work showed that the whole extract of E. excelsa a part
from its estrogenic effect also inhibit the growth of MCF-7 cells, a wel established in
vitro model for ER (+) breast cancer. With this backgroung, we then aimed this step
of our study to get more insights into the “active principle” of the extract accounting
for these two biological activities. To do this, the ethanol crude extract was
fractionated over silica gel into five fractions that were further screened for biological
activity. We concentrated our field of investigations on fractions exhibiting both
estrogenic and cytotoxic activities. The YES was used as first step screen to select
estrogenic fractions and the XTT assay used to access their effect on the viability of
MCF-7 cells. F4 was almost as potent as the whole extract in respect to the
estrogenic effect but was less cytotoxic suggesting that other non-estrogenic
compounds of the whole extract may also account for its effects. The HPLC profiling
method showed that some major compounds present in the whole extract are either
absent or in low amount in F4 (Figure 22).
To gain more understanding of the mechanisms underlying F4-effect on the
growth of MCF-7 cells, cell cycle analysis was performed following 48h-treatment with
increasing concentrations (1-100 µg/ml). At Lower concentrations, F4 similar to E2
promoted cell cycle progression and inhibited cell death (Figure 23) and this effect
was ER-dependent (Figure 24). Gene expression analysis showed that F4 at low

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 104
concentration like E2 increased CCND1 level and modulated the Bax/Bcl-2 ratio in
favor of cell survival (Figure 27). Cell cycle progression is a tightly controlled process
controlled by cyclins and cyclin-dependent kinases (CDKs) complexes and the
deregulation of their expression frequently impacts on breast cancer outcome
including response to therapy (Nigg, 1995; Caldon et al., 2006). CyclinD1 along with
its binding CDKs is an essential sensor and activator of cell cycle initiation and
progression (particularly the transition from G1 to S-phase) and is overexpressed in
up to 50% of human invasive breast cancers (Sun et al., 2012). Bcl-2 family members
play an important role in the mediation and regulation of apoptotic signaling pathways
acting to either inhibit (e.g. Bcl-2) or promote cell death (e.g. Bax). The Bax/Bcl-2
ratio has been recognized as a key factor and determines whether a cell will undergo
apoptosis (Leung and Wang, 1999; Gewies, 2003). As CCND1, Bcl-2 overexpression
is common in breast cancer (Oakes et al., 2011). It is well known that estrogens
through several mechanisms among which their stimulating effects on cell cycle
progression and pro-survival Bcl-2 are an important risk factor for breast cancer
development and progression (Sutherland et al., 1995). F4 at low and maybe
physiologically relevant concentrations behaved in the same way. However, the
relevance of this result for the in vivo situation where the mammary gland is
surrounded by other tissues and is under the influence of growth factors and other
hormones is still to be determined. In fact the local estrogen level in the tissue, the
expressed cofactors, and the expression level of ERα and ERβ are important factors
to take into consideration when considering the in vivo situation (Bondesson and
Gustafsson, 2010). For instance PEs have been shown to exhibit antagonistic
properties in the presence of E2 and this ability was also confirmed here as we
showed that F4 at the same concentration range where growth stimulation was
observed, acted as an antagonist in the presence of E2. This antagonistic ability may
be of interest because it is generally believed that treatments that trigger ER
antagonist effects are protective against breast cancer and favorably affect the
course of breast cancer once diagnosed (Bolca et al., 2010).
At higher concentrations, F4 modulated the G1/S transition as confirmed by
decreased CCND1 mRNA level and induced apoptosis sustained by M30 staining
results and increased Bax/ Bcl-2 ratio (Figures 23, 26, and 27). Impairment of
apoptosis is implicated in not only cancer development but also in the therapy

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 105
(Adams and Cory, 2007). Cancer cells are resistant to apoptosis and it is proposed
that drugs or treatment strategies that can restore the apoptotic signaling pathways
towards normality have the potential to eliminate cancer cells (Wong, 2011). It is
however worth to note that this effect was achieved with relatively high and probably
supraphysiologic concentrations of the extract fraction.
Cotreatment with E2 and higher concentrations of F4 induced more cells to
transit from G1 to S as compared to single F4 treatment. But at the same time an
increase of the percentage of cells in SubG1 was observed. At high concentrations
F4 acted almost the same way as the pure antiestrogen ZK with the difference that
ZK caused cell cycle arrest whereas F4 blocked G1/S transition without preventing
the cells to transit from S to G2/M. (Dalvai and Bystricky, 2010) showed that cells in
the S-phase are more sensitive to antiestrogens-induced apoptosis. Therefore,
cotreatment with E2 by inducing more cells transit from G1 to S also increases their
susceptibility to undergo F4 induced cell death.
Overall these results show that F4 has a biphasic profile on the growth MCF-7
breast cancer cells: low dose stimulation and high dose inhibition through induction of
apoptosis. However, whether this is relevant for breast cancer outcomes is still to be
determined because a direct translation of in vitro results to the in vivo situation would
be oversimplifying and unrealistic. One interesting aspect is that the effects of low
and physiologically relevant concentrations of F4 depended on the presence or
absence of E2. In the presence of E2, F4 could counteract E2-induced stimulation of
cancer cells growth and may therefore be of significance and of interest for
premenopausal women.
5.2 Effect of estrogens and chemicals of plant origin on C2C12
murine skeletal muscle cells growth.
Increasing evidences have emerged over the past decades indicating that
estrogens can regulate skeletal muscle mass (established by a net difference
between muscle protein synthesis rate and muscle protein breakdown). Galluzzo et al
(2009) proposed that estrogens (E2) should be included in the list of skeletal muscle
trophic factors. Skeletal muscle hypertrophy is a dynamic process involving both an
increase in the number of nuclei (due to the stimulation of the proliferation,

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 106
differentiation and fusion of quiescent satellite cells) as well as an increase in
cytoplasmic volume (due to an increase in protein synthesis or remodeling). Here, we
were interested in the effects of our test chemicals on the increase in cytoplamic
volume. Therefore, to differentiate between effects on myoblast differentiation and
those on the trophic state of existing myofibers, fusion of C2C12 myoblasts towards
myotubes was induced prior to the treatment. E2, genistein and Ecdy induced a
significant increase of myotube diameter compared to untreated myotubes probably
due to a shift of the balance in favor of protein synthesis. The Akt signaling pathway
plays an important role in skeletal muscle hypertrophy. E2 (Stefanetti et al, 2008),
and Ecdy (Gorelick-Feldman et al, 2010) have been shown to increase Akt activity
and its downstream anabolic signaling proteins. Conversely to Ecdy that increase the
protein synthesis in the muscle, E2 and genistein have been shown to rather
decrease protein breakdown in porcine skeletal muscle cells (Mau, 2007).
As previously shown by others (Kahlert et al., 1997; Galluzzo et al., 2009;
Milanesi et al., 2009), the presence of both ER isoforms in C2C12 cells was
confirmed in this study. We also showed for the first time that the Ecdy-induced
hypertrophic response is ER-mediated (Figure 38). ERα and ERβ selective agonists
were used to further elucidate the contribution of each ER subtype on E2 and Ecdy-
induced myotube hypertrophy. 16α-LE2 (ALPHA) and 8β-VE2 (BETA) are subtype-
selective steroidal ER agonists designed on the basis of the available protein
structure information (Hillisch et al., 2004). In a transactivation assay with APHA and
BETA, Hillisch et al (2004) showed that the selectivity of both agonists was lost when
used at higher concentrations (Figure 43). Based on this knowledge, dose-response
effects of each agonist were investigated and the results demonstrated that the
hypertrophic response was ERβ-mediated. Moreover, the use of an ERβ antagonist
allowed us to unequivocally show that E2 and Ecdy-induced hypertrophy is mediated
through ERβ (Figure 41). With respect to E2, these results are in line with previous
findings of our lab which indicate that the anabolic effect of E2 is primordially ERβ-
mediated (Velders et al., 2012; Weigt et al., 2012) but are in contradiction with the
findings of Galluzzo et al (2009) who found that ERα was the principal mediator of E2
effect in skeletal muscle. However, it is worth to note that their experiments were
performed on differentiating myotubes, while we used already differentiated
myotubes. With respect to Ecdy, published data on its ability to bind ERs are

Discussion
M.Sc. Sadrine Tchoukouegno Ngueu 107
conflicting. Gao et al (2008) showed that Ecdy was able to transactivate ERβ in
embryonic kidney stem cells (293T) transfected with ERβ expression vector. Others
(Bathori et al., 2008; Seidlova-Wuttke et al., 2010) however demonstrated that Ecdy
was not able to bind ER.
Figure 43: Transactivation studies with ERα agonist (16α-LE2) and ERβ agonist (8β-VE2) in
comparison to E2 (Hillisch et al., 2004)
In summary, we successfully used a system with differentiated C2C12
myoblasts to demonstrate the skeletal muscle growth promoting effects of E2, the
phytoestrogen genistein, and the phytoecdysteroid ecdysterone. Moreover, we
confirmed the results of previous studies showing that ERβ was the ER isoform
mediating E2 anabolic effects, and showed for the first time that the anabolic effect of
Ecdy is ERβ-mediated.

Conclusion and perspectives
M.Sc. Sadrine Tchoukouegno Ngueu 108
CHAPTER 6: CONCLUSION AND PERSPECTIVES

Conclusion and perspectives
M.Sc. Sadrine Tchoukouegno Ngueu 109
The aim of the present thesis was to investigate the biological activities and
mechanisms of action of two plant-related chemicals either as an extract or as a pure
compound. The results showed that an ethanol extract of Erythrina excelsa Baker
exhibits estrogenic and cytotoxic effects that are in close agreement with its
traditionally reported use to alleviate menopausal complaints and to treat female
gynaecological tumors. Chemical characterization revealed a rich metabolic content
of the extract and the isolation of the characterized compounds is ongoing with the
aim to investigate their biological activities as single compounds. We also confirmed
the anabolic efficiency of Ecdysterone and showed for the first time that it is ERβ-
mediated, a result that is of interest and may provide new insights not only for
strategies aiming to treat health conditions related to muscle atrophy (sarcopenia,
Duchenne myopathy), but also for doping. Overall, the findings of this thesis show
that although traditional medicine is often regarded with a lot of skepticism, it may
provide helpful basic knowledge for research orientation. In an era marked by
worldwide increase in life expectancy and its associated chronic diseases, the
maintenance of good health and quality of life is a challenging global issue involving
both developed and developing countries. The potential of traditional medicine is
increasingly recognized but a lot of effort is still needed in other to attain an evidence-
based utilization.
With respect to breast cancer risk, one interesting result that emerged from our
investigations confirming previous recommendations is that local estrogens levels is
an important factor to take into consideration when considering phytoestrogens
consumption and breast cancer outcomes. Intensive research has been made over
the last decade to clarify whether phytoestrogens could be an alternative to hormone
replacement therapy (HRT). Unfortunately most of the performed human clinical trials
failed to show clear evidences. To my opinion, strategies combining HRT and
phytoestrogens maybe of more interest in the future.
Limitations and recommendations for further research
The results of this thesis give an overview of the possible biological activity of
E.excelsa but do not provide sufficient understanding to enable specific
recommandations for use. With respect to Ecdysterone, although we clearly show

Conclusion and perspectives
M.Sc. Sadrine Tchoukouegno Ngueu 110
that its skeletal muscle growth promoting effects are ERβ- mediated, published data
regarding its ER binding ability are conflicting. Future investigations should clarify this
issue.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 111
Bibliography
Adams JM, Cory S (1998). "The Bcl-2 protein family: arbiters of cell survival." Science
281(5381): 1322-6.
Adams JM, Cory S (2007). "The Bcl-2 apoptotic switch in cancer development and
therapy." Oncogene 26(9): 1324-37.
Adlercreutz H, Bannwart C, Wahala K, Makela T, Brunow G, Hase T, Arosemena PJ,
Kellis JT, Jr., Vickery LE (1993). "Inhibition of human aromatase by
mammalian lignans and isoflavonoid phytoestrogens." J Steroid Biochem Mol
Biol 44(2): 147-53.
Adlercreutz H, Hockerstedt K, Bannwart C, Bloigu S, Hamalainen E, Fotsis T, Ollus A
(1987). "Effect of dietary components, including lignans and phytoestrogens,
on enterohepatic circulation and liver metabolism of estrogens and on sex
hormone binding globulin (SHBG)." J Steroid Biochem 27(4-6): 1135-44.
Adlercreutz H, Martin F, Pulkkinen M, Dencker H, Rimer U, Sjoberg NO, Tikkanen MJ
(1976). "Intestinal metabolism of estrogens." J Clin Endocrinol Metab 43(3):
497-505.
Adlercreutz H, Mousavi Y, Hockerstedt K (1992). "Diet and breast cancer." Acta
Oncol 31(2): 175-81.
Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, Itoh N, Shibuya M,
Fukami Y (1987). "Genistein, a specific inhibitor of tyrosine-specific protein
kinases." J Biol Chem 262(12): 5592-5.
Antignac JP, Cariou R, Le Bizec B, Cravedi JP, Andre F (2003). "Identification of
phytoestrogens in bovine milk using liquid chromatography/electrospray
tandem mass spectrometry." Rapid Commun Mass Spectrom 17(12): 1256-64.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 112
Arai N, Strom A, Rafter JJ, Gustafsson JA (2000). "Estrogen receptor beta mRNA in
colon cancer cells: growth effects of estrogen and genistein." Biochem
Biophys Res Commun 270(2): 425-31.
Aronica SM, Katzenellenbogen BS (1993). "Stimulation of estrogen receptor-
mediated transcription and alteration in the phosphorylation state of the rat
uterine estrogen receptor by estrogen, cyclic adenosine monophosphate, and
insulin-like growth factor-I." Mol Endocrinol 7(6): 743-52.
Aronica SM, Kraus WL, Katzenellenbogen BS (1994). "Estrogen action via the cAMP
signaling pathway: stimulation of adenylate cyclase and cAMP-regulated gene
transcription." Proc Natl Acad Sci U S A 91(18): 8517-21.
Ascenzi P, Bocedi A, Marino M (2006). "Structure-function relationship of estrogen
receptor alpha and beta: impact on human health." Mol Aspects Med 27(4):
299-402.
Axelson M, Sjovall J, Gustafsson BE, Setchell KD (1984). "Soya--a dietary source of
the non-steroidal oestrogen equol in man and animals." J Endocrinol 102(1):
49-56.
Balmain N (1991). "Calbindin-D9k. A vitamin-D-dependent, calcium-binding protein in
mineralized tissues." Clin Orthop Relat Res(265): 265-76.
Bannwart C, Adlercreutz H, Fotsis T, Wähälä K, Hase T, Brunow G (1984).
"Identification of O-desmethylangolensin, a metabolite of daidzein, and of
matairesinol, one likely plant precursor of the animal lignan enterolactone, in
human urine. ." Finn Chem Lett 4-5: 120–125.
Barrington R, Zhang M, Fischer M, Carroll MC (2001). "The role of complement in
inflammation and adaptive immunity." Immunol Rev 180: 5-15.
Bateman HL, Patisaul HB (2008). "Disrupted female reproductive physiology
following neonatal exposure to phytoestrogens or estrogen specific ligands is

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 113
associated with decreased GnRH activation and kisspeptin fiber density in the
hypothalamus." Neurotoxicology 29(6): 988-97.
Bathori M, Toth N, Hunyadi A, Marki A, Zador E (2008). "Phytoecdysteroids and
anabolic-androgenic steroids--structure and effects on humans." Curr Med
Chem 15(1): 75-91.
Beato M (1989). "Gene regulation by steroid hormones." Cell 56(3): 335-44.
Bennetts HW, Underwood EJ, Shier FL (1946). "A specific breeding problem of
sheep on subterranean clover pastures in Western Australia." Br Vet J
102(11): 348-52.
Berrino F, Bellati C, Secreto G, Camerini E, Pala V, Panico S, Allegro G, Kaaks R
(2001). "Reducing bioavailable sex hormones through a comprehensive
change in diet: the diet and androgens (DIANA) randomized trial." Cancer
Epidemiol Biomarkers Prev 10(1): 25-33.
Bhat HK, Hacker HJ, Bannasch P, Thompson EA, Liehr JG (1993). "Localization of
estrogen receptors in interstitial cells of hamster kidney and in estradiol-
induced renal tumors as evidence of the mesenchymal origin of this
neoplasm." Cancer Res 53(22): 5447-51.
Bolanos R, Del Castillo A, Francia J (2010). "Soy isoflavones versus placebo in the
treatment of climacteric vasomotor symptoms: systematic review and meta-
analysis." Menopause 17(3): 660-6.
Bolca S, Urpi-Sarda M, Blondeel P, Roche N, Vanhaecke L, Possemiers S, Al-
Maharik N, Botting N, De Keukeleire D, Bracke M, Heyerick A, Manach C,
Depypere H (2010). "Disposition of soy isoflavones in normal human breast
tissue." Am J Clin Nutr 91(4): 976-84.
Bondesson M, Gustafsson JA (2010). "Does consuming isoflavones reduce or
increase breast cancer risk?" Genome Med 2(12): 90.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 114
Bowe J, Li XF, Sugden D, Katzenellenbogen JA, Katzenellenbogen BS, O'Byrne KT
(2003). "The effects of the phytoestrogen, coumestrol, on gonadotropin-
releasing hormone (GnRH) mRNA expression in GT1-7 GnRH neurones." J
Neuroendocrinol 15(2): 105-8.
Brisken C, Rajaram RD (2006). "Alveolar and lactogenic differentiation." J Mammary
Gland Biol Neoplasia 11(3-4): 239-48.
Brooks JD, Thompson LU (2005). "Mammalian lignans and genistein decrease the
activities of aromatase and 17beta-hydroxysteroid dehydrogenase in MCF-7
cells." J Steroid Biochem Mol Biol 94(5): 461-7.
Bruns ME, Overpeck JG, Smith GC, Hirsch GN, Mills SE, Bruns DE (1988). "Vitamin
D-dependent calcium binding protein in rat uterus: differential effects of
estrogen, tamoxifen, progesterone, and pregnancy on accumulation and
cellular localization." Endocrinology 122(6): 2371-8.
Bunone G, Briand PA, Miksicek RJ, Picard D (1996). "Activation of the unliganded
estrogen receptor by EGF involves the MAP kinase pathway and direct
phosphorylation." Embo J 15(9): 2174-83.
Caldon CE, Daly RJ, Sutherland RL, Musgrove EA (2006). "Cell cycle control in
breast cancer cells." J Cell Biochem 97(2): 261-74.
Calmels P, Vico L, Alexandre C, Minaire P (1995). "Cross-sectional study of muscle
strength and bone mineral density in a population of 106 women between the
ages of 44 and 87 years: relationship with age and menopause." Eur J Appl
Physiol Occup Physiol 70(2): 180-6.
Carpinelli MR, Wise AK, Arhatari BD, Bouillet P, Manji SS, Manning MG, Cooray AA,
Burt RA (2012). "Anti-apoptotic gene Bcl2 is required for stapes development
and hearing." Cell Death Dis 3: e362.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 115
Carroll MC, Prodeus AP (1998). "Linkages of innate and adaptive immunity." Curr
Opin Immunol 10(1): 36-40.
Castoria G, Migliaccio A, Bilancio A, Di Domenico M, de Falco A, Lombardi M,
Fiorentino R, Varricchio L, Barone MV, Auricchio F (2001). "PI3-kinase in
concert with Src promotes the S-phase entry of oestradiol-stimulated MCF-7
cells." Embo J 20(21): 6050-9.
Cavalieri E, Chakravarti D, Guttenplan J, Hart E, Ingle J, Jankowiak R, Muti P, Rogan
E, Russo J, Santen R, Sutter T (2006). "Catechol estrogen quinones as
initiators of breast and other human cancers: implications for biomarkers of
susceptibility and cancer prevention." Biochim Biophys Acta 1766(1): 63-78.
Cavalieri E, Frenkel K, Liehr JG, Rogan E, Roy D (2000). "Estrogens as endogenous
genotoxic agents--DNA adducts and mutations." J Natl Cancer Inst
Monogr(27): 75-93.
Cavalieri E, Rogan E, Chakravarti D (2004). "The role of endogenous catechol
quinones in the initiation of cancer and neurodegenerative diseases." Methods
Enzymol 382: 293-319.
Cavalieri EL, Stack DE, Devanesan PD, Todorovic R, Dwivedy I, Higginbotham S,
Johansson SL, Patil KD, Gross ML, Gooden JK, Ramanathan R, Cerny RL,
Rogan EG (1997). "Molecular origin of cancer: catechol estrogen-3,4-quinones
as endogenous tumor initiators." Proc Natl Acad Sci U S A 94(20): 10937-42.
Chakravarti D, Mailander PC, Li KM, Higginbotham S, Zhang HL, Gross ML, Meza
JL, Cavalieri EL, Rogan EG (2001). "Evidence that a burst of DNA
depurination in SENCAR mouse skin induces error-prone repair and forms
mutations in the H-ras gene." Oncogene 20(55): 7945-53.
Chaudhary KD, Lupien PJ, Hinse C (1969). "Effect of ecdysone on glutamic
decarboxylase in rat brain." Experientia 25(3): 250-1.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 116
Chen Z, Yuhanna IS, Galcheva-Gargova Z, Karas RH, Mendelsohn ME, Shaul PW
(1999). "Estrogen receptor alpha mediates the nongenomic activation of
endothelial nitric oxide synthase by estrogen." J Clin Invest 103(3): 401-6.
Chermnykh NS, Shimanovskii NL, Shutko GV, Syrov VN (1988). "[The action of
methandrostenolone and ecdysterone on the physical endurance of animals
and on protein metabolism in the skeletal muscles]." Farmakol Toksikol 51(6):
57-60.
Christoffel J, Rimoldi G, Wuttke W (2006). "Effects of 8-prenylnaringenin on the
hypothalamo-pituitary-uterine axis in rats after 3-month treatment." J
Endocrinol 188(3): 397-405.
Chu S, Nishi Y, Yanase T, Nawata H, Fuller PJ (2004). "Transrepression of estrogen
receptor beta signaling by nuclear factor-kappab in ovarian granulosa cells."
Mol Endocrinol 18(8): 1919-28.
Cowley SM, Parker MG (1999). "A comparison of transcriptional activation by ER
alpha and ER beta." J Steroid Biochem Mol Biol 69(1-6): 165-75.
Cragg GM, Newman DJ (2013). "Natural products: A continuing source of novel drug
leads." Biochim Biophys Acta.
D'Alessandro TL, Boersma-Maland BJ, Peterson TG, Sfakianos J, Prasain JK, Patel
RP, Darley-Usmar VM, Botting NP, Barnes S (2005). "Metabolism of
phytoestrogen conjugates." Methods Enzymol 400: 316-42.
Dalu A, Haskell JF, Coward L, Lamartiniere CA (1998). "Genistein, a component of
soy, inhibits the expression of the EGF and ErbB2/Neu receptors in the rat
dorsolateral prostate." Prostate 37(1): 36-43.
Dalvai M, Bystricky K (2010). "Cell cycle and anti-estrogen effects synergize to
regulate cell proliferation and ER target gene expression." PLoS One 5(6):
e11011.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 117
Danial NN (2007). "BCL-2 family proteins: critical checkpoints of apoptotic cell death."
Clin Cancer Res 13(24): 7254-63.
Danielian PS, White R, Lees JA, Parker MG (1992). "Identification of a conserved
region required for hormone dependent transcriptional activation by steroid
hormone receptors." Embo J 11(3): 1025-33.
Darimont BD, Wagner RL, Apriletti JW, Stallcup MR, Kushner PJ, Baxter JD,
Fletterick RJ, Yamamoto KR (1998). "Structure and specificity of nuclear
receptor-coactivator interactions." Genes Dev 12(21): 3343-56.
Dawson SJ, Makretsov N, Blows FM, Driver KE, Provenzano E, Le Quesne J,
Baglietto L, Severi G, Giles GG, McLean CA, Callagy G, Green AR, Ellis I,
Gelmon K, Turashvili G, Leung S, Aparicio S, Huntsman D, Caldas C, Pharoah
P (2010). "BCL2 in breast cancer: a favourable prognostic marker across
molecular subtypes and independent of adjuvant therapy received." Br J
Cancer 103(5): 668-75.
Dewell A, Hollenbeck PL, Hollenbeck CB (2006). "Clinical review: a critical evaluation
of the role of soy protein and isoflavone supplementation in the control of
plasma cholesterol concentrations." J Clin Endocrinol Metab 91(3): 772-80.
Dhooge W, Arijs K, D'Haese I, Stuyvaert S, Versonnen B, Janssen C, Verstraete W,
Comhaire F (2006). "Experimental parameters affecting sensitivity and
specificity of a yeast assay for estrogenic compounds: results of an
interlaboratory validation exercise." Anal Bioanal Chem 386(5): 1419-28.
Dickson RB, Stancel GM (2000). "Estrogen receptor-mediated processes in normal
and cancer cells." J Natl Cancer Inst Monogr(27): 135-45.
Diel P, Geis RB, Caldarelli A, Schmidt S, Leschowsky UL, Voss A, Vollmer G (2004).
"The differential ability of the phytoestrogen genistein and of estradiol to
induce uterine weight and proliferation in the rat is associated with a substance

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 118
specific modulation of uterine gene expression." Mol Cell Endocrinol 221(1-2):
21-32.
Diel P, Schulz T, Smolnikar K, Strunck E, Vollmer G, Michna H (2000). "Ability of
xeno- and phytoestrogens to modulate expression of estrogen-sensitive genes
in rat uterus: estrogenicity profiles and uterotropic activity." J Steroid Biochem
Mol Biol 73(1-2): 1-10.
Diel P, Walter A, Fritzemeier KH, Hegele-Hartung C, Knauthe R (1995).
"Identification of estrogen regulated genes in Fe33 rat hepatoma cells by
differential display polymerase chain reaction and their hormonal regulation in
rat liver and uterus." J Steroid Biochem Mol Biol 55(3-4): 363-73.
Dionne IJ, Kinaman KA, Poehlman ET (2000). "Sarcopenia and muscle function
during menopause and hormone-replacement therapy." J Nutr Health Aging
4(3): 156-61.
Djiogue S, Njamen D, Halabalaki M, Kretzschmar G, Beyer A, Mbanya JC,
Skaltsounis AL, Vollmer G (2010). "Estrogenic properties of naturally occurring
prenylated isoflavones in U2OS human osteosarcoma cells: Structure-activity
relationships." J Steroid Biochem Mol Biol 120(4-5): 184-91.
Edvardsson K, Strom A, Jonsson P, Gustafsson JA, Williams C (2011). "Estrogen
receptor beta induces antiinflammatory and antitumorigenic networks in colon
cancer cells." Mol Endocrinol 25(6): 969-79.
EICCAM (European Information Center for Complementary and Alternative Medicine)
(2009). http://www.eiccam.eu/portal.html. Accessed April 2013.
Enns DL, Tiidus PM (2010). "The influence of estrogen on skeletal muscle: sex
matters." Sports Med 40(1): 41-58.
Eriksson H, Gustafsson JA (1971). "Excretion of steroid hormones in adults. Steroids
in faeces from adults." Eur J Biochem 18(1): 146-50.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 119
Evans BA, Griffiths K, Morton MS (1995). "Inhibition of 5 alpha-reductase in genital
skin fibroblasts and prostate tissue by dietary lignans and isoflavonoids." J
Endocrinol 147(2): 295-302.
Faulds MH, Zhao C, Dahlman-Wright K, Gustafsson JA (2012). "The diversity of sex
steroid action: regulation of metabolism by estrogen signaling." J Endocrinol
212(1): 3-12.
Feigelson HS, Henderson BE (1996). "Estrogens and breast cancer." Carcinogenesis
17(11): 2279-84.
Feng W, Ribeiro RC, Wagner RL, Nguyen H, Apriletti JW, Fletterick RJ, Baxter JD,
Kushner PJ, West BL (1998). "Hormone-dependent coactivator binding to a
hydrophobic cleft on nuclear receptors." Science 280(5370): 1747-9.
Fishman J, Bradlow HL, Gallagher TF (1960). "Oxidative metabolism of estradiol." J
Biol Chem 235: 3104-7.
Galluzzo P, Rastelli C, Bulzomi P, Acconcia F, Pallottini V, Marino M (2009). "17beta-
Estradiol regulates the first steps of skeletal muscle cell differentiation via ER-
alpha-mediated signals." Am J Physiol Cell Physiol 297(5): C1249-62.
Gao L, Cai G, Shi X (2008). "Beta-ecdysterone induces osteogenic differentiation in
mouse mesenchymal stem cells and relieves osteoporosis." Biol Pharm Bull
31(12): 2245-9.
Geis RB, Diel P, Degen GH, Vollmer G (2005). "Effects of genistein on the
expression of hepatic genes in two rat strains (Sprague-Dawley and Wistar)."
Toxicol Lett 157(1): 21-9.
Gewies M. (2003). "Introduction to Apoptosis." Accessed April 2013 from
http://www.celldeath.de/encyclo/aporev/aporev.htm.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 120
Giguere V, Yang N, Segui P, Evans RM (1988). "Identification of a new class of
steroid hormone receptors." Nature 331(6151): 91-4.
Goping IS, Gross A, Lavoie JN, Nguyen M, Jemmerson R, Roth K, Korsmeyer SJ,
Shore GC (1998). "Regulated targeting of BAX to mitochondria." J Cell Biol
143(1): 207-15.
Gorelick-Feldman J, Cohick W, Raskin I (2010). "Ecdysteroids elicit a rapid Ca2+ flux
leading to Akt activation and increased protein synthesis in skeletal muscle
cells." Steroids 75(10): 632-7.
Gorelick-Feldman J, Maclean D, Ilic N, Poulev A, Lila MA, Cheng D, Raskin I (2008).
"Phytoecdysteroids increase protein synthesis in skeletal muscle cells." J Agric
Food Chem 56(10): 3532-7.
Hahn WC, Weinberg RA (2002). "Rules for making human tumor cells." N Engl J Med
347(20): 1593-603.
Halabalaki M, Alexi X, Aligiannis N, Lambrinidis G, Pratsinis H, Florentin I, Mitakou S,
Mikros E, Skaltsounis AL, Alexis MN (2006). "Estrogenic activity of
isoflavonoids from Onobrychis ebenoides." Planta Med 72(6): 488-93.
Hartman J, Edvardsson K, Lindberg K, Zhao C, Williams C, Strom A, Gustafsson JA
(2009). "Tumor repressive functions of estrogen receptor beta in SW480 colon
cancer cells." Cancer Res 69(15): 6100-6.
Havelock JC, Rainey WE, Carr BR (2004). "Ovarian granulosa cell lines." Mol Cell
Endocrinol 228(1-2): 67-78.
Heldring N, Pike A, Andersson S, Matthews J, Cheng G, Hartman J, Tujague M,
Strom A, Treuter E, Warner M, Gustafsson JA (2007). "Estrogen receptors:
how do they signal and what are their targets." Physiol Rev 87(3): 905-31.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 121
Henttu PM, Kalkhoven E, Parker MG (1997). "AF-2 activity and recruitment of steroid
receptor coactivator 1 to the estrogen receptor depend on a lysine residue
conserved in nuclear receptors." Mol Cell Biol 17(4): 1832-9.
Hertrampf T, Gruca MJ, Seibel J, Laudenbach U, Fritzemeier KH, Diel P (2007). "The
bone-protective effect of the phytoestrogen genistein is mediated via ER
alpha-dependent mechanisms and strongly enhanced by physical activity."
Bone 40(6): 1529-35.
Hikino H, Takemoto T (1972). "Arthropod moulting hormones from Plants,
Achyranthes and Cyathula." Naturwissenschaften 59(3): 91-8.
Hillisch A, Peters O, Kosemund D, Muller G, Walter A, Schneider B, Reddersen G,
Elger W, Fritzemeier KH (2004). "Dissecting physiological roles of estrogen
receptor alpha and beta with potent selective ligands from structure-based
design." Mol Endocrinol 18(7): 1599-609.
Hofer N, Diel P, Wittsiepe J, Wilhelm M, Degen GH (2009). "Dose- and route-
dependent hormonal activity of the metalloestrogen cadmium in the rat
uterus." Toxicol Lett 191(2-3): 123-31.
Howes LG, Howes JB, Knight DC (2006). "Isoflavone therapy for menopausal
flushes: a systematic review and meta-analysis." Maturitas 55(3): 203-11.
Ignar-Trowbridge DM, Nelson KG, Bidwell MC, Curtis SW, Washburn TF, McLachlan
JA, Korach KS (1992). "Coupling of dual signaling pathways: epidermal growth
factor action involves the estrogen receptor." Proc Natl Acad Sci U S A 89(10):
4658-62.
Ignar-Trowbridge DM, Pimentel M, Parker MG, McLachlan JA, Korach KS (1996).
"Peptide growth factor cross-talk with the estrogen receptor requires the A/B
domain and occurs independently of protein kinase C or estradiol."
Endocrinology 137(5): 1735-44.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 122
Improta-Brears T, Whorton AR, Codazzi F, York JD, Meyer T, McDonnell DP (1999).
"Estrogen-induced activation of mitogen-activated protein kinase requires
mobilization of intracellular calcium." Proc Natl Acad Sci U S A 96(8): 4686-91.
Ince BA, Montano MM, Katzenellenbogen BS (1994). "Activation of transcriptionally
inactive human estrogen receptors by cyclic adenosine 3',5'-monophosphate
and ligands including antiestrogens." Mol Endocrinol 8(10): 1397-406.
Isotton AL, Wender MC, Casagrande A, Rollin G, Czepielewski MA (2012). "Effects
of oral and transdermal estrogen on IGF1, IGFBP3, IGFBP1, serum lipids, and
glucose in patients with hypopituitarism during GH treatment: a randomized
study." Eur J Endocrinol 166(2): 207-13.
Jefferson WN, Padilla-Banks E, Newbold RR (2005). "Adverse effects on female
development and reproduction in CD-1 mice following neonatal exposure to
the phytoestrogen genistein at environmentally relevant doses." Biol Reprod
73(4): 798-806.
Jefferson WN, Patisaul HB, Williams CJ (2012). "Reproductive consequences of
developmental phytoestrogen exposure." Reproduction 143(3): 247-60.
Jiofack T, Fokunang C, Guedje N, Kemeuze V, Fongnzossie E, Nkongmeneck BA,
Mapongmetsem PM, Tsabang N (2010). "Ethnobotanical uses of medicinal
plants of two ethnoecological regions of Cameroon " International Journal of
Medicine and Medical Sciences 2(3): 60-79.
Joannou GE, Kelly GE, Reeder AY, Waring M, Nelson C (1995). "A urinary profile
study of dietary phytoestrogens. The identification and mode of metabolism of
new isoflavonoids." J Steroid Biochem Mol Biol 54(3-4): 167-84.
Kadi F, Karlsson C, Larsson B, Eriksson J, Larval M, Billig H, Jonsdottir IH (2002).
"The effects of physical activity and estrogen treatment on rat fast and slow
skeletal muscles following ovariectomy." J Muscle Res Cell Motil 23(4): 335-9.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 123
Kahlert S, Grohe C, Karas RH, Lobbert K, Neyses L, Vetter H (1997). "Effects of
estrogen on skeletal myoblast growth." Biochem Biophys Res Commun
232(2): 373-8.
Kao YC, Zhou C, Sherman M, Laughton CA, Chen S (1998). "Molecular basis of the
inhibition of human aromatase (estrogen synthetase) by flavone and
isoflavone phytoestrogens: A site-directed mutagenesis study." Environ Health
Perspect 106(2): 85-92.
Kelly GE, Nelson C, Waring MA, Joannou GE, Reeder AY (1993). "Metabolites of
dietary (soya) isoflavones in human urine." Clin Chim Acta 223(1-2): 9-22.
Kim SM, Jung EM, An BS, Hwang I, Vo TT, Kim SR, Lee SM, Choi KC, Jeung EB
(2012). "Additional effects of bisphenol A and paraben on the induction of
calbindin-D(9K) and progesterone receptor via an estrogen receptor pathway
in rat pituitary GH3 cells." J Physiol Pharmacol 63(5): 445-55.
Kirkman H (1959). "Estrogen-induced tumors of the kidney. III. Growth characteristics
in the Syrian hamster." Natl Cancer Inst Monogr 1: 1-57.
Kluxen FM, Hofer N, Kretzschmar G, Degen GH, Diel P (2012). "Cadmium modulates
expression of aryl hydrocarbon receptor-associated genes in rat uterus by
interaction with the estrogen receptor." Arch Toxicol 86(4): 591-601.
Knight DC, Eden JA (1996). "A review of the clinical effects of phytoestrogens."
Obstet Gynecol 87(5 Pt 2): 897-904.
Kouki T, Kishitake M, Okamoto M, Oosuka I, Takebe M, Yamanouchi K (2003).
"Effects of neonatal treatment with phytoestrogens, genistein and daidzein, on
sex difference in female rat brain function: estrous cycle and lordosis." Horm
Behav 44(2): 140-5.
Kraus WL, McInerney EM, Katzenellenbogen BS (1995). "Ligand-dependent,
transcriptionally productive association of the amino- and carboxyl-terminal

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 124
regions of a steroid hormone nuclear receptor." Proc Natl Acad Sci U S A
92(26): 12314-8.
Krisinger J, Setoyama T, Leung PC (1994). "Expression of calbindin-D9k in the early
pregnant rat uterus: effects of RU 486 and correlation to estrogen receptor
mRNA." Mol Cell Endocrinol 102(1-2): 15-22.
Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gustafsson
JA (1997). "Comparison of the ligand binding specificity and transcript tissue
distribution of estrogen receptors alpha and beta." Endocrinology 138(3): 863-
70.
Kulling SE, Metzler M (1997). "Induction of micronuclei, DNA strand breaks and
HPRT mutations in cultured Chinese hamster V79 cells by the phytoestrogen
coumoestrol." Food Chem Toxicol 35(6): 605-13.
Kumar R, Zakharov MN, Khan SH, Miki R, Jang H, Toraldo G, Singh R, Bhasin S,
Jasuja R (2011). "The dynamic structure of the estrogen receptor." J Amino
Acids: 812540.
Kurzer MS, Xu X (1997). "Dietary phytoestrogens." Annu Rev Nutr 17: 353-81.
Kushner PJ, Agard DA, Greene GL, Scanlan TS, Shiau AK, Uht RM, Webb P (2000).
"Estrogen receptor pathways to AP-1." J Steroid Biochem Mol Biol 74(5): 311-
7.
Lahooti H, Thorsen T, Aakvaag A (1998). "Modulation of mouse estrogen receptor
transcription activity by protein kinase C delta." J Mol Endocrinol 20(2): 245-
59.
Le Bizec B, Antignac J, Monteau F, Andre F (2002). "Ecdysteroids: one potential new
anabolic family in breeding animals." Analytica Chimica Acta 473: 89-97.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 125
Lee JS, Kim DH, Liu KH, Oh TK, Lee CH (2005). "Identification of flavonoids using
liquid chromatography with electrospray ionization and ion trap tandem mass
spectrometry with an MS/MS library." Rapid Commun Mass Spectrom 19(23):
3539-48.
Lee KY, Oh GT, Kang JH, Shin SM, Heo BE, Yun YW, Paik SG, Krisinger J, Leung
PC, Jeung EB (2003). "Transcriptional regulation of the mouse calbindin-D9k
gene by the ovarian sex hormone." Mol Cells 16(1): 48-53.
Lee PD, Conover CA, Powell DR (1993). "Regulation and function of insulin-like
growth factor-binding protein-1." Proc Soc Exp Biol Med 204(1): 4-29.
Lee SA, Shu XO, Li H, Yang G, Cai H, Wen W, Ji BT, Gao J, Gao YT, Zheng W
(2009). "Adolescent and adult soy food intake and breast cancer risk: results
from the Shanghai Women's Health Study." Am J Clin Nutr 89(6): 1920-6.
Lee SY, Oh SM, Lee SK, Chung KH (2005). "Antiestrogenic effects of marijuana
smoke condensate and cannabinoid compounds." Arch Pharm Res 28(12):
1365-75.
Lee YB, Lee HJ, Kim KS, Lee JY, Nam SY, Cheon SH, Sohn HS (2004). "Evaluation
of the preventive effect of isoflavone extract on bone loss in ovariectomized
rats." Biosci Biotechnol Biochem 68(5): 1040-5.
Lefebvre Y (1993). Fibromyomes utérins. Encyclopédie médicale chirurgicale. Paris,
France. Editions Techniques.
Lemmens R. (2008). "Erythrina excelsa Baker." Accessed August 2012 from
http://database.prota.org/search.htm.
Lemoine S, Granier P, Tiffoche C, Rannou-Bekono F, Thieulant ML, Delamarche P
(2003). "Estrogen receptor alpha mRNA in human skeletal muscles." Med Sci
Sports Exerc 35(3): 439-43.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 126
Leung LK, Wang TT (1999). "Differential effects of chemotherapeutic agents on the
Bcl-2/Bax apoptosis pathway in human breast cancer cell line MCF-7." Breast
Cancer Res Treat 55(1): 73-83.
Li KM, Todorovic R, Devanesan P, Higginbotham S, Kofeler H, Ramanathan R,
Gross ML, Rogan EG, Cavalieri EL (2004). "Metabolism and DNA binding
studies of 4-hydroxyestradiol and estradiol-3,4-quinone in vitro and in female
ACI rat mammary gland in vivo." Carcinogenesis 25(2): 289-97.
Li SH, Liu XX, Bai YY, Wang XJ, Sun K, Chen JZ, Hui RT (2010). "Effect of oral
isoflavone supplementation on vascular endothelial function in
postmenopausal women: a meta-analysis of randomized placebo-controlled
trials." Am J Clin Nutr 91(2): 480-6.
Liu MM, Albanese C, Anderson CM, Hilty K, Webb P, Uht RM, Price RH, Jr., Pestell
RG, Kushner PJ (2002). "Opposing action of estrogen receptors alpha and
beta on cyclin D1 gene expression." J Biol Chem 277(27): 24353-60.
Losa SM, Todd KL, Sullivan AW, Cao J, Mickens JA, Patisaul HB (2011). "Neonatal
exposure to genistein adversely impacts the ontogeny of hypothalamic
kisspeptin signaling pathways and ovarian development in the peripubertal
female rat." Reprod Toxicol 31(3): 280-9.
Lowe DA, Baltgalvis KA, Greising SM (2010). "Mechanisms behind estrogen's
beneficial effect on muscle strength in females." Exerc Sport Sci Rev 38(2):
61-7.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951). "Protein measurement with
the Folin phenol reagent." J Biol Chem 193(1): 265-75.
Mahmoodzadeh S, Fritschka S, Dworatzek E, Pham TH, Becher E, Kuehne A,
Davidson MM, Regitz-Zagrosek V (2009). "Nuclear factor-kappaB regulates
estrogen receptor-alpha transcription in the human heart." J Biol Chem
284(37): 24705-14.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 127
Majinda RRT, Wanjala C, Juma BF (2005). "Boiactive non-alkaloidal constituents
from the genus Erythrina." Studies in natural Products Chemistry 32 (Part 12):
821-853.
Maltais ML, Desroches J, Dionne IJ (2009). "Changes in muscle mass and strength
after menopause." J Musculoskelet Neuronal Interact 9(4): 186-97.
Marino M, Acconcia F, Bresciani F, Weisz A, Trentalance A (2002). "Distinct
nongenomic signal transduction pathways controlled by 17beta-estradiol
regulate DNA synthesis and cyclin D(1) gene transcription in HepG2 cells."
Mol Biol Cell 13(10): 3720-9.
Martin-Millan M, Almeida M, Ambrogini E, Han L, Zhao H, Weinstein RS, Jilka RL,
O'Brien CA, Manolagas SC (2010). "The estrogen receptor-alpha in
osteoclasts mediates the protective effects of estrogens on cancellous but not
cortical bone." Mol Endocrinol 24(2): 323-34.
Mau M, (2007). "In vitro-Wachstum und -Differenzierung prciner Skelettmuskelzellen
unter den Einfluss der Isoflavone Genistein und Daidzein". PhD thesis Faculty
of Mathematics and Natural Sciences, Ernst-Moritz-Arndt University,
Greifswald-Germany.
ub-ed.ub.unigreifswald.de/opus/volltexte/2008/466/pdf/Dissertation_Mau_Marcus.pdf
McInerney EM, Katzenellenbogen BS (1996). "Different regions in activation function-
1 of the human estrogen receptor required for antiestrogen- and estradiol-
dependent transcription activation." J Biol Chem 271(39): 24172-8.
Mense SM, Hei TK, Ganju RK, Bhat HK (2008). "Phytoestrogens and breast cancer
prevention: possible mechanisms of action." Environ Health Perspect 116(4):
426-33.
Mersereau JE, Levy N, Staub RE, Baggett S, Zogovic T, Chow S, Ricke WA,
Tagliaferri M, Cohen I, Bjeldanes LF, Leitman DC (2008). "Liquiritigenin is a

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 128
plant-derived highly selective estrogen receptor beta agonist." Mol Cell
Endocrinol 283(1-2): 49-57.
Messina M, McCaskill-Stevens W, Lampe JW (2006). "Addressing the soy and breast
cancer relationship: review, commentary, and workshop proceedings." J Natl
Cancer Inst 98(18): 1275-84.
Messina MJ, Wood CE (2008). "Soy isoflavones, estrogen therapy, and breast
cancer risk: analysis and commentary." Nutr J 7: 17.
Migliaccio S, Wetsel WC, Fox WM, Washburn TF, Korach KS (1993). "Endogenous
protein kinase-C activation in osteoblast-like cells modulates responsiveness
to estrogen and estrogen receptor levels." Mol Endocrinol 7(9): 1133-43.
Milanesi L, Vasconsuelo A, de Boland AR, Boland R (2009). "Expression and
subcellular distribution of native estrogen receptor beta in murine C2C12 cells
and skeletal muscle tissue." Steroids 74(6): 489-97.
Misra RR, Hursting SD, Perkins SN, Sathyamoorthy N, Mirsalis JC, Riccio ES,
Crowell JA (2002). "Genotoxicity and carcinogenicity studies of soy
isoflavones." Int J Toxicol 21(4): 277-85.
Mitchell JH, Duthie SJ, Collins AR (2000). "Effects of phytoestrogens on growth and
DNA integrity in human prostate tumor cell lines: PC-3 and LNCaP." Nutr
Cancer 38(2): 223-8.
Mohibi S, Mirza S, Band H, Band V (2011). "Mouse models of estrogen receptor-
positive breast cancer." J Carcinog 10: 35.
Molzberger AF, Vollmer G, Hertrampf T, Moller FJ, Kulling S, Diel P (2012). "In utero
and postnatal exposure to isoflavones results in a reduced responsivity of the
mammary gland towards estradiol." Mol Nutr Food Res 56(3): 399-409.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 129
Moran AL, Nelson SA, Landisch RM, Warren GL, Lowe DA (2007). "Estradiol
replacement reverses ovariectomy-induced muscle contractile and myosin
dysfunction in mature female mice." J Appl Physiol 102(4): 1387-93.
Morley JE, Baumgartner RN, Roubenoff R, Mayer J, Nair KS (2001). "Sarcopenia." J
Lab Clin Med 137(4): 231-43.
Myung SK, Ju W, Choi HJ, Kim SC (2009). "Soy intake and risk of endocrine-related
gynaecological cancer: a meta-analysis." Bjog 116(13): 1697-705.
Nagata C, Takatsuka N, Kurisu Y, Shimizu H (1998). "Decreased serum total
cholesterol concentration is associated with high intake of soy products in
Japanese men and women." J Nutr 128(2): 209-13.
Nakamura T, Imai Y, Matsumoto T, Sato S, Takeuchi K, Igarashi K, Harada Y,
Azuma Y, Krust A, Yamamoto Y, Nishina H, Takeda S, Takayanagi H, Metzger
D, Kanno J, Takaoka K, Martin TJ, Chambon P, Kato S (2007). "Estrogen
prevents bone loss via estrogen receptor alpha and induction of Fas ligand in
osteoclasts." Cell 130(5): 811-23.
Nandi S, Guzman RC, Yang J (1995). "Hormones and mammary carcinogenesis in
mice, rats, and humans: a unifying hypothesis." Proc Natl Acad Sci U S A
92(9): 3650-7.
Neuman E, Ladha MH, Lin N, Upton TM, Miller SJ, DiRenzo J, Pestell RG, Hinds
PW, Dowdy SF, Brown M, Ewen ME (1997). "Cyclin D1 stimulation of estrogen
receptor transcriptional activity independent of cdk4." Mol Cell Biol 17(9):
5338-47.
Newbold RR, Bullock BC, McLachlan JA (1990). "Uterine adenocarcinoma in mice
following developmental treatment with estrogens: a model for hormonal
carcinogenesis." Cancer Res 50(23): 7677-81.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 130
Newman DJ, Cragg GM (2010). "Natural products as sources of new drugs over the
30 years from 1981 to 2010." J Nat Prod 75(3): 311-35.
Nigg EA (1995). "Cyclin-dependent protein kinases: key regulators of the eukaryotic
cell cycle." Bioessays 17(6): 471-80.
Nikaido Y, Yoshizawa K, Danbara N, Tsujita-Kyutoku M, Yuri T, Uehara N, Tsubura A
(2004). "Effects of maternal xenoestrogen exposure on development of the
reproductive tract and mammary gland in female CD-1 mouse offspring."
Reprod Toxicol 18(6): 803-11.
Nilsson S, Gustafsson JA (2010). Estrogen receptors: their actions and functional
roles in health and disease. Nuclear Receptors: Current Concepts and Future
Challenges (C. C. Bunce, M.J, Springer, The Netherlands: 91–141.
Nilsson S, Gustafsson JA (2011). "Estrogen receptors: therapies targeted to receptor
subtypes." Clin Pharmacol Ther 89(1): 44-55.
Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G, Enmark E,
Pettersson K, Warner M, Gustafsson JA (2001). "Mechanisms of estrogen
action." Physiol Rev 81(4): 1535-65.
O'Lone R, Frith MC, Karlsson EK, Hansen U (2004). "Genomic targets of nuclear
estrogen receptors." Mol Endocrinol 18(8): 1859-75.
Oakes SR, Vaillant F, Lim E, Lee L, Breslin K, Feleppa F, Deb S, Ritchie ME, Takano
E, Ward T, Fox SB, Generali D, Smyth GK, Strasser A, Huang DC, Visvader
JE, Lindeman GJ (2011). "Sensitization of BCL-2-expressing breast tumors to
chemotherapy by the BH3 mimetic ABT-737." Proc Natl Acad Sci U S A
109(8): 2766-71.
OECD (2007). "OECD guideline for testing of chemicals. Uterotrophic Bioassay in
Rodents: A short-term screening test for oestrogenic properties." Test
Guidelines 440.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 131
Okamoto Y, Suzuki, A., Ueda, K., Ito C., Itoigawa, M., Furukawa, H., Nishihara, T and
Kojima, N. (2006). "Anti-estrogenic activity of prenylated isoflavone from
Millettia pachycarpa: Implications for pharmacophores and unique
mechanisms." Journal of health Science 52(2): 186-191.
Okura A, Arakawa H, Oka H, Yoshinari T, Monden Y (1988). "Effect of genistein on
topoisomerase activity and on the growth of [Val 12]Ha-ras-transformed NIH
3T3 cells." Biochem Biophys Res Commun 157(1): 183-9.
Ollberding NJ, Lim U, Wilkens LR, Setiawan VW, Shvetsov YB, Henderson BE,
Kolonel LN, Goodman MT (2012). "Legume, soy, tofu, and isoflavone intake
and endometrial cancer risk in postmenopausal women in the multiethnic
cohort study." J Natl Cancer Inst 104(1): 67-76.
Onate SA, Boonyaratanakornkit V, Spencer TE, Tsai SY, Tsai MJ, Edwards DP,
O'Malley BW (1998). "The steroid receptor coactivator-1 contains multiple
receptor interacting and activation domains that cooperatively enhance the
activation function 1 (AF1) and AF2 domains of steroid receptors." J Biol
Chem 273(20): 12101-8.
Otaka T, Okui S, Uchiyama M (1969). "Stimulation of protein synthesis in mouse liver
by ecdysterone." Chem Pharm Bull (Tokyo) 17(1): 75-81.
Otaka T, Uchiyama M, Okui S, Takemoto T, Hikino H (1968). "Stimulatory effect of
insect-metamorphosing steroids from Achyranthes and Cyathula on protein
synthesis in mouse liver." Chem Pharm Bull (Tokyo) 16(12): 2426-9.
Owens JW, Ashby J (2002). "Critical review and evaluation of the uterotrophic
bioassay for the identification of possible estrogen agonists and antagonists: in
support of the validation of the OECD uterotrophic protocols for the laboratory
rodent. Organisation for Economic Co-operation and Development." Crit Rev
Toxicol 32(6): 445-520.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 132
Owuor BO, Kisangau DP (2006). "Kenyan medicinal plants used as antivenin: a
comparison of plant usage." J Ethnobiol Ethnomed 2: 7.
Pfaffl MW (2001). "A new mathematical model for relative quantification in real-time RT-PCR." Nucleic Acids Res 29(9): e45.
Pagliacci MC, Smacchia M, Migliorati G, Grignani F, Riccardi C, Nicoletti I (1994).
"Growth-inhibitory effects of the natural phyto-oestrogen genistein in MCF-7
human breast cancer cells." Eur J Cancer 30A(11): 1675-82.
Palacios S, Pornel B, Vazquez F, Aubert L, Chantre P, Mares P (2010). "Long-term
endometrial and breast safety of a specific, standardized soy extract."
Climacteric 13(4): 368-75.
Patisaul HB, Jefferson W (2010). "The pros and cons of phytoestrogens." Front
Neuroendocrinol 31(4): 400-19.
Petkov V, Roussinov K, Todorov S, Lazarova M, Yonkov D, Draganova S (1984).
"Pharmacological investigations on Rhaponticum carthamoides." Planta Med
50(3): 205-9.
Pilsakova L, Riecansky I, Jagla F (2010). "The physiological actions of isoflavone
phytoestrogens." Physiol Res 59(5): 651-64.
Pollanen E, Sipila S, Alen M, Ronkainen PH, Ankarberg-Lindgren C, Puolakka J,
Suominen H, Hamalainen E, Turpeinen U, Konttinen YT, Kovanen V (2011).
"Differential influence of peripheral and systemic sex steroids on skeletal
muscle quality in pre- and postmenopausal women." Aging Cell 10(4): 650-60.
Porter W, Saville B, Hoivik D, Safe S (1997). "Functional synergy between the
transcription factor Sp1 and the estrogen receptor." Mol Endocrinol 11(11):
1569-80.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 133
Power RF, Mani SK, Codina J, Conneely OM, O'Malley BW (1991). "Dopaminergic
and ligand-independent activation of steroid hormone receptors." Science
254(5038): 1636-9.
Prall OW, Rogan EM, Musgrove EA, Watts CK, Sutherland RL (1998). "c-Myc or
cyclin D1 mimics estrogen effects on cyclin E-Cdk2 activation and cell cycle
reentry." Mol Cell Biol 18(8): 4499-508.
Pritts EA, Parker WH, Olive DL (2009). "Fibroids and infertility: an updated systematic
review of the evidence." Fertil Steril 91(4): 1215-23.
Rosenberg ME, Silkensen J (1995). "Clusterin: physiologic and pathophysiologic
considerations." Int J Biochem Cell Biol 27(7): 633-45.
Rosenfeld MG, Glass CK (2001). "Coregulator codes of transcriptional regulation by
nuclear receptors." J Biol Chem 276(40): 36865-8.
Routledge EJ, Sumpter JP (1996). "Estrogenic activity of surfactants and some of
their degradation products assassed using a recombinant yeast screen "
Environmental Toxicology and Chemistry Vol. 15(No. 3): 241–248.
Samy RP, Thwin MM, Gopalakrishnakone P, Ignacimuthu S (2008). "Ethnobotanical
survey of folk plants for the treatment of snakebites in Southern part of
Tamilnadu, India." J Ethnopharmacol 115(2): 302-12.
Sandberg AA, Slaunwhite WR, Jr. (1957). "Studies on phenolic steroids in human
subjects. II. The metabolic fate and hepato-biliary-enteric circulation of C14-
estrone and C14-estradiol in women." J Clin Invest 36(8): 1266-78.
Saville B, Wormke M, Wang F, Nguyen T, Enmark E, Kuiper G, Gustafsson JA, Safe
S (2000). "Ligand-, cell-, and estrogen receptor subtype (alpha/beta)-
dependent activation at GC-rich (Sp1) promoter elements." J Biol Chem
275(8): 5379-87.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 134
Schoch RD, Willoughby D, Greenwood M (2006). "The regulation and expression of
the creatine transporter: a brief review of creatine supplementation in humans
and animals." J Int Soc Sports Nutr 3: 60-6.
Schwabe JW, Chapman L, Finch JT, Rhodes D (1993). "The crystal structure of the
estrogen receptor DNA-binding domain bound to DNA: how receptors
discriminate between their response elements." Cell 75(3): 567-78.
Schwartz LM, Truman JW (1984). "Hormonal control of muscle atrophy and
degeneration in the moth Antheraea polyphemus." J Exp Biol 111: 13-30.
Seidlova-Wuttke D, Christel D, Kapur P, Nguyen BT, Jarry H, Wuttke W (2010).
"Beta-ecdysone has bone protective but no estrogenic effects in
ovariectomized rats." Phytomedicine 17(11): 884-9.
Seo HS, DeNardo DG, Jacquot Y, Laios I, Vidal DS, Zambrana CR, Leclercq G,
Brown PH (2006). "Stimulatory effect of genistein and apigenin on the growth
of breast cancer cells correlates with their ability to activate ER alpha." Breast
Cancer Res Treat 99(2): 121-34.
Setchell KD, Borriello SP, Hulme P, Kirk DN, Axelson M (1984). "Nonsteroidal
estrogens of dietary origin: possible roles in hormone-dependent disease." Am
J Clin Nutr 40(3): 569-78.
Setchell KD, Cassidy A (1999). "Dietary isoflavones: biological effects and relevance
to human health." J Nutr 129(3): 758S-767S.
Setchell KD, Lydeking-Olsen E (2003). "Dietary phytoestrogens and their effect on
bone: evidence from in vitro and in vivo, human observational, and dietary
intervention studies." Am J Clin Nutr 78(3 Suppl): 593S-609S.
Shanle EK, Xu W (2011). "Endocrine disrupting chemicals targeting estrogen
receptor signaling: identification and mechanisms of action." Chem Res
Toxicol 24(1): 6-19.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 135
Shao W, Brown M (2004). "Advances in estrogen receptor biology: prospects for
improvements in targeted breast cancer therapy." Breast Cancer Res 6(1): 39-
52.
Shiau AK, Barstad D, Loria PM, Cheng L, Kushner PJ, Agard DA, Greene GL (1998).
"The structural basis of estrogen receptor/coactivator recognition and the
antagonism of this interaction by tamoxifen." Cell 95(7): 927-37.
Shivhare MK, Singour PK, Chaurasiya PK, Pawar RS (2012). "Trianthema
portulacastrum Linn. (Bishkhapra)." Pharmacogn Rev 6(12): 132-40.
Short KR, Nair KS (2000). "The effect of age on protein metabolism." Curr Opin Clin
Nutr Metab Care 3(1): 39-44.
Shull JD, Spady TJ, Snyder MC, Johansson SL, Pennington KL (1997). "Ovary-intact,
but not ovariectomized female ACI rats treated with 17beta-estradiol rapidly
develop mammary carcinoma." Carcinogenesis 18(8): 1595-601.
Shwaery GT, Vita JA, Keaney JF, Jr. (1997). "Antioxidant protection of LDL by
physiological concentrations of 17 beta-estradiol. Requirement for estradiol
modification." Circulation 95(6): 1378-85.
Simons R, Gruppen H, Bovee TF, Verbruggen MA, Vincken JP (2012). "Prenylated
isoflavonoids from plants as selective estrogen receptor modulators
(phytoSERMs)." Food Funct 3(8): 810-27.
Simons R, Vincken JP, Bakx EJ, Verbruggen MA, Gruppen H (2009). "A rapid
screening method for prenylated flavonoids with ultra-high-performance liquid
chromatography/electrospray ionisation mass spectrometry in licorice root
extracts." Rapid Commun Mass Spectrom 23(19): 3083-93.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 136
Siow RC, Mann GE (2010). "Dietary isoflavones and vascular protection: activation of
cellular antioxidant defenses by SERMs or hormesis?" Mol Aspects Med
31(6): 468-77.
Smith CL, Conneely OM, O'Malley BW (1993). "Modulation of the ligand-independent
activation of the human estrogen receptor by hormone and antihormone." Proc
Natl Acad Sci U S A 90(13): 6120-4.
Somai S, Chaouat M, Jacob D, Perrot JY, Rostene W, Forgez P, Gompel A (2003).
"Antiestrogens are pro-apoptotic in normal human breast epithelial cells." Int J
Cancer 105(5): 607-12.
Stefanetti AI, Turner MJ, Quick RJ, Russell AP (2008). The effect of estrogen on Akt
signalling and protein synthesis in C2C12 skeletal myotubes. Proceeding of
the Australian Physiological Society. aups.org.au/Proceeding/39/70P/.
Steinberg FM, Murray MJ, Lewis RD, Cramer MA, Amato P, Young RL, Barnes S,
Konzelmann KL, Fischer JG, Ellis KJ, Shypailo RJ, Fraley JK, Smith EO,
Wong WW (2011). "Clinical outcomes of a 2-y soy isoflavone supplementation
in menopausal women." Am J Clin Nutr 93(2): 356-67.
Stitt TN, Drujan D, Clarke BA, Panaro F, Timofeyva Y, Kline WO, Gonzalez M,
Yancopoulos GD, Glass DJ (2004). "The IGF-1/PI3K/Akt pathway prevents
expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO
transcription factors." Mol Cell 14(3): 395-403.
Sun Y, Luo D, Liao DJ (2012). "CyclinD1 protein plays different roles in modulating
chemoresponses in MCF-7 and MDA-MB 231 cells." Journal of carcinogensis
11(12): 18-29.
Sutherland RL, Hamilton JA, Sweeney KJ, Watts CK, Musgrove EA (1995).
"Expression and regulation of cyclin genes in breast cancer." Acta Oncol
34(5): 651-6.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 137
Syrov VN, Kurmukov AG (1976). "[The anabolic properties of turkesterone
phytoecdysone and turkesterone tetraacetate in experiments on male rats]."
Probl Endokrinol (Mosk) 22(3): 107-12.
Takayama M, Nomura T (1995). "Structure of the Diene Originating from a retro-
Diels- Alder Cleavage of the Natural Product Kuwanon G." Rapid Commun
Mass Spectrom 9: 383-386.
Tanee Fomum Z, Ayafor JF, Ifeadike PN, Nkengfack AE, Wandji J (1986). "Erythrina
Studies; Part 3. Isolation of an isoflavone from Erythrina senegalensis and
Erythrina excelsa." Planta Med(4): 341.
Tang M, Abplanalp W, Subbiah MT (1997). "Association of estrogens with human
plasma lipoproteins: studies using estradiol-17beta and its hydrophobic
derivative." J Lab Clin Med 129(4): 447-52.
The National Toxicology Program (NTP) (2002). "Background review document
(BRD) current status of test methods for detecting endocrine disruptors: In
vitro estrogen transcriptional activation assays. National Institute of
Environmental Health Sciences (NIEHS)." NIH Publication. Research Triangle
Park. NC 27709 nº 03-4505.
Tobin NP, Bergh J (2012). "Analysis of Cyclin D1 in Breast Cancer: A Call to Arms."
Curr Breast Cancer Rep 4(3): 171-173.
Toth N, Szabo A, Kacsala P, Heger J, Zador E (2008). "20-Hydroxyecdysone
increases fiber size in a muscle-specific fashion in rat." Phytomedicine 15(9):
691-8.
Trowbridge JM, Rogatsky I, Garabedian MJ (1997). "Regulation of estrogen receptor
transcriptional enhancement by the cyclin A/Cdk2 complex." Proc Natl Acad
Sci U S A 94(19): 10132-7.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 138
Tsai MJ, O'Malley BW (1994). "Molecular mechanisms of action of steroid/thyroid
receptor superfamily members." Annu Rev Biochem 63: 451-86.
Tseng L, Stolee A, Gurpide E (1972). "Quantitative studies on the uptake and
metabolism of estrogens and progesterone by human endometrium."
Endocrinology 90(2): 390-404.
Tsujimoto Y, Finger LR, Yunis J, Nowell PC, Croce CM (1984). "Cloning of the
chromosome breakpoint of neoplastic B cells with the t(14;18) chromosome
translocation." Science 226(4678): 1097-9.
van der Woude H, Ter Veld MG, Jacobs N, van der Saag PT, Murk AJ, Rietjens IM
(2005). "The stimulation of cell proliferation by quercetin is mediated by the
estrogen receptor." Mol Nutr Food Res 49(8): 763-71.
Vargas AJ, Burd R (2010). "Hormesis and synergy: pathways and mechanisms of
quercetin in cancer prevention and management." Nutr Rev 68(7): 418-28.
Velders M, Schleipen B, Fritzemeier KH, Zierau O, Diel P (2012). "Selective estrogen
receptor-beta activation stimulates skeletal muscle growth and regeneration."
Faseb J 26(5): 1909-20.
Wandji J, Nkengfack AE, Fomum ZT, Ubillas R, Killday KB, Tempesta MS (1990). "A
new prenylated isoflavone and long chain esters from two Erythrina species." J
Nat Prod 53(6): 1425-9.
Wasserman RH, Taylor AN (1966). "Vitamin d3-induced calcium-binding protein in
chick intestinal mucosa." Science 152(3723): 791-3.
Webb P, Nguyen P, Shinsako J, Anderson C, Feng W, Nguyen MP, Chen D, Huang
SM, Subramanian S, McKinerney E, Katzenellenbogen BS, Stallcup MR,
Kushner PJ (1998). "Estrogen receptor activation function 1 works by binding
p160 coactivator proteins." Mol Endocrinol 12(10): 1605-18.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 139
Wei H, Wei L, Frenkel K, Bowen R, Barnes S (1993). "Inhibition of tumor promoter-
induced hydrogen peroxide formation in vitro and in vivo by genistein." Nutr
Cancer 20(1): 1-12.
Weigt C, Hertrampf T, Zoth N, Fritzemeier KH, Diel P (2012). "Impact of estradiol, ER
subtype specific agonists and genistein on energy homeostasis in a rat model
of nutrition induced obesity." Mol Cell Endocrinol 351(2): 227-38.
Whitsett TG, Jr., Lamartiniere CA (2006). "Genistein and resveratrol: mammary
cancer chemoprevention and mechanisms of action in the rat." Expert Rev
Anticancer Ther 6(12): 1699-706.
WHO (2008). "Traditional medicine." Fact sheet N°134
Wiik A, Ekman M, Johansson O, Jansson E, Esbjornsson M (2009). "Expression of
both oestrogen receptor alpha and beta in human skeletal muscle tissue."
Histochem Cell Biol 131(2): 181-9.
Wiik A, Glenmark B, Ekman M, Esbjornsson-Liljedahl M, Johansson O, Bodin K,
Enmark E, Jansson E (2003). "Oestrogen receptor beta is expressed in adult
human skeletal muscle both at the mRNA and protein level." Acta Physiol
Scand 179(4): 381-7.
Wong RS (2011). "Apoptosis in cancer: from pathogenesis to treatment." J Exp Clin
Cancer Res 30: 87.
Wunsche W, Tenniswood MP, Schneider MR, Vollmer G (1998). "Estrogenic
regulation of clusterin mRNA in normal and malignant endometrial tissue." Int
J Cancer 76(5): 684-8.
Yamori Y, Moriguchi EH, Teramoto T, Miura A, Fukui Y, Honda KI, Fukui M, Nara Y,
Taira K, Moriguchi Y (2002). "Soybean isoflavones reduce postmenopausal
bone resorption in female Japanese immigrants in Brazil: a ten-week study." J
Am Coll Nutr 21(6): 560-3.

Bibliography
M.Sc. Sadrine Tchoukouegno Ngueu 140
Zhang CZ, Wang SX, Zhang Y, Chen JP, Liang XM (2005). "In vitro estrogenic
activities of Chinese medicinal plants traditionally used for the management of
menopausal symptoms." J Ethnopharmacol 98(3): 295-300.
Zhang R, Hu SJ, Li C, Zhang F, Gan HQ, Mei QB (2012). "Achyranthes bidentata
root extract prevent OVX-induced osteoporosis in rats." J Ethnopharmacol
139(1): 12-8.
Zhang Y, Zhang P, Cheng Y (2008). "Structural characterization of isoprenylated
flavonoids from Kushen by electrospray ionization multistage tandem mass
spectrometry." J Mass Spectrom 43(10): 1421-31.
Zhao E, Mu Q (2010). "Phytoestrogen Biological Actions on Mammalian
Reproductive System and Cancer Growth." Sci Pharm 79(1): 1-20.
Zhao Z, Kosinska W, Khmelnitsky M, Cavalieri EL, Rogan EG, Chakravarti D, Sacks
PG, Guttenplan JB (2006). "Mutagenic activity of 4-hydroxyestradiol, but not 2-
hydroxyestradiol, in BB rat2 embryonic cells, and the mutational spectrum of 4-
hydroxyestradiol." Chem Res Toxicol 19(3): 475-9.
Zwijsen RM, Wientjens E, Klompmaker R, van der Sman J, Bernards R, Michalides
RJ (1997). "CDK-independent activation of estrogen receptor by cyclin D1."
Cell 88(3): 405-15.

Publications and Presentations
M.Sc. Sadrine Tchoukouegno Ngueu 141
Publications and presentations
Tchoukouegno SN, Job Tchoumtchoua, Dieudonné Njamen, Maria Halabalaki, Ute
Laudenbach-Leschowski, Patrick Diel. Estrogenic and cytotoxic properties of the
ethanolic extract of Erythrina excelsa. Original research article submitted in Planta
Medica.
Tchoukouegno SN et al. A phytoestrogen rich-fraction of Erythrina excelsa
modulates cell cycle progression and induces apoptosis of human breast cancer
MCF-7 cell line. Original research article in preparation.
Parr MK, Haupt O, Hengevoss J, Tchoukouegno Ngueu S, Fritzemeier KH, Marion
Picotta, Muhn P, Diel P. Estrogen receptor beta mediated anabolic effects – insights
from mechanistic studies on the phytoecdysteroid ecdysterone. Original research
article in preparation.
Tchoukouegno SN, Dieudonné Njamen, Ute Laudenbach-Leschowski, Patrick Diel.
Estrogenic effects of the ethanolic extract of Erythrina excelsa Baker
(Leguminoseae). Poster presentation at the Bregenz sommer school of
Endocrinology (Austria, July 2011).
Tchoukouegno SN. Estrogenic and cytotoxic properties of Erythrina excelsa Baker
(Fabaceae). Oral presentation at the symposium on bioactive principles of medicinal
plants and diet. (Dresden, October 2012).
Maria Kristina Parr, PhD, Oliver Haupt, Sadrine Tchoukouegno Ngueu, Karl-
Heinrich Fritzemeier, , Peter Muhn, and Patrick Rene Diel. Estrogen Receptor Beta
mediated anabolic effects – Insights from mechanistic studies on the
Phytoecdysteroid Ecdysterone and Selective Ligands. Poster presentation at the
95thannual Meeting and Expo of the Endocrine Society 14-18 June 2013.

Curriculum Vitae
M.Sc. Sadrine Tchoukouegno Ngueu 142
Curriculum vitae
PERSONAL DETAILS
Name Sadrine Tchoukouegno Ngueu
Family and marital status Married
Date& Place of Birth 20.04.1984 at Bamenyam
Nationality Cameroonian
EDUCATIONAL QUALIFICATIONS
Degree/Diploma Field /
specialty
Institution Marks Yea
r
1. High School Diploma Mathematics
and Natural
Sciences
Lycee de Nkol-
Eton
16.35/20 200
1
2. Bachelor of Science Animal Biology
& Physiology
University of
Yaounde I
200
4
3. Master‟s Degree (without
thesis)
Animal
Physiology
University of
Yaounde I
13.25/20 200
5
4. Master‟s Degree (with
thesis)
Animal
Physiology
University of
Yaounde I
14.37/20 200
7
5. PhD candicate Natural
Sciences
German Sport
University
(Cologne)
from
201
0
AWARDS & SCHOLARSHIP
1. Price of the Best Cameroonian female student at the High School Diploma
exam 2001
2. DAAD Scholarship for PhD Studies in Natural Sciences
INTERSHIPS
Internship at the “Centre Pasteur du Cameroun”
(Biochemistry laboratory)
October-November
2005 (One month)
Internship at the laboratory of Pharmacognosy and
Natural Products Chemistry, Faculty of Pharmacy,
University of Athens, Greece.
February – Mai 2013
(3 months)
TECHNICAL SKILLS
1. Cellular and Molecular Biology: PCR, Western blot, FACS
2. Chromatography: CC, FCPC, HPLC-DAD
3. Scientific softwares: Chromquest, Sigmaplot, SPSS.
LANGUAGES
1. French (Native)
2. English (Very Good)
3. German (Very Good)

Curriculum Vitae
M.Sc. Sadrine Tchoukouegno Ngueu 143
4. Spanish (Basic)
PROFESSIONAL AFFILIATIONS AND MEMBERSHIPS
Member of the Society of Medicinal Plant and Natural Product Research (GA)
Member of ANSA e.v (Alumni Netzwerk Subsahara Afrika)