binding of soybean agglutinin by normal and trypsin-treated red blood cells
TRANSCRIPT

BIOCHIMICA ET BIOPHYSICA ACTA 387
BBA Report
BBA 21332
Binding of soybean agglutinin by normal and trypsin-treated red blood cells
JULIUS A. GORDON*, NATHAN SHARON** and HALINA LIS
Department of Biophysics, Weizmann Institute o f Science, Rehovoth (Israel)
(Received February 21st, 1972)
SUMMARY
Trypsinized human erythrocytes were found to bind 3 times as much
soybean agglutinin as the untreated cells. With rabbit erythrocytes, no effect of trypsinization on the binding of the agglutinin was observed. However, the susceptibility to agglutination increased 100- to 200-fold for all erythrocytes. The
increased erythrocyte agglutinability can therefore not be explained simply by an
increase in the number of agglutinin binding sites.
Prior treatment of erythrocytes with proteolytic enzymes can dramatically
reduce the amount of agglutinin (lectin) required to produce hemagglutination ~- 4. The
increased ease of agglutination following enzymatic treatment has been attributed to the
exposure of additional agglutinin-specific receptor sites which are thought to be in a "cryptic" form on the untreated cell 3' 4 and more recently to rearrangements of pre-
existing sites s . We report here the results of our quantitative binding studies of
12 s I-labelled soybean agglutinin with human and with rabbit erythrocytes; these studies
indicate the absence of a simple relationship between the extent of binding of the agglutinin and the increased agglutinability of the erythrocyte following trypsinization.
Soybean agglutinin was isolated and purified as previously described 6. The
labelling was performed according to the chloramine-T method of Greenwood et al. 7,8
using carrier-free Na 12s I (Radiochemical Centre, Amersham, England, 20 mCi/ml). The
labelled protein was separated from the low molecular weight radioactive material by
Abbreviation: GalNAc, N-acetyl-D-galactosamine.
*Permanent address: Department of Pathology, University of Colorado Medical School, Denver, Colo., U.S.A. **To whom correspondence should be addressed.
Biochim. Biophys. Acta, 264 (1972) 387-391

388 BBA REPORT
gel filtration on a Sephadex G-150 column (1,4 cm × 45 cm) in saline; the fractions containing the labelled protein were combined and dialysed 48 tl at 4 °C against saline to remove the last traces of low molecular weight radioactive material. The preparation obtained behaved upon chromatography on columns of calcium phosphate in a fashion identical to that of unlabelled soybean agglutinin 6, and there was no change m agglutinating activity or specificity of the agglutinin after iodination. The counting
rate of the ~2 s l-labelled soybean agglutinin preparation used in the reported experiments was 4500 cpm//ag protein. Erythrocytes were prepared for use by washing 3-4 times with
saline containing 0.01 M potassium-sodium phosphate, pH 7.4 (phosphate-buffered saline) and suspending m phosphate-buffered saline (4% cell suspension, about 3.108 cells/ ml) to give an absorbance of 2 at 620 nm. The absorbance was measured in a Colernan Junior Spectrophotometer equipped with a special adaptor 2' 9 using 10 mm X 75 mm round cuvettes. Determination of agglutinating activity on untreated and trypsinized erythrocytes was perforined by the quantitative spectrophotometric method of Liener as previously described 2' 9. Trypsinization was carried out with Bacto-trypsin, Difco, I mg/ml, at 37 °C. At specified times samples were withdrawn, immediately cooled, the trypsinized erythrocytes washed 5 times with phosphate-buffered saline and re-suspended in the original volume of phosphate-buffered saline.
For binding experiments, 5 ml of cell suspension was treated with the specified amount of ~2Sl.labelled soybean agglutinin for 30 min at room temperature. The supernatant was removed from the cells which were then washed 2 times with 5 ml
phosphate-buffered saline; the cell-bound radioactivity was determined in a Packard series 5000 Auto-Gamma spectrometer. Another washing did not remove any significant amount of radioactivity. The extent of binding was found to be dependent
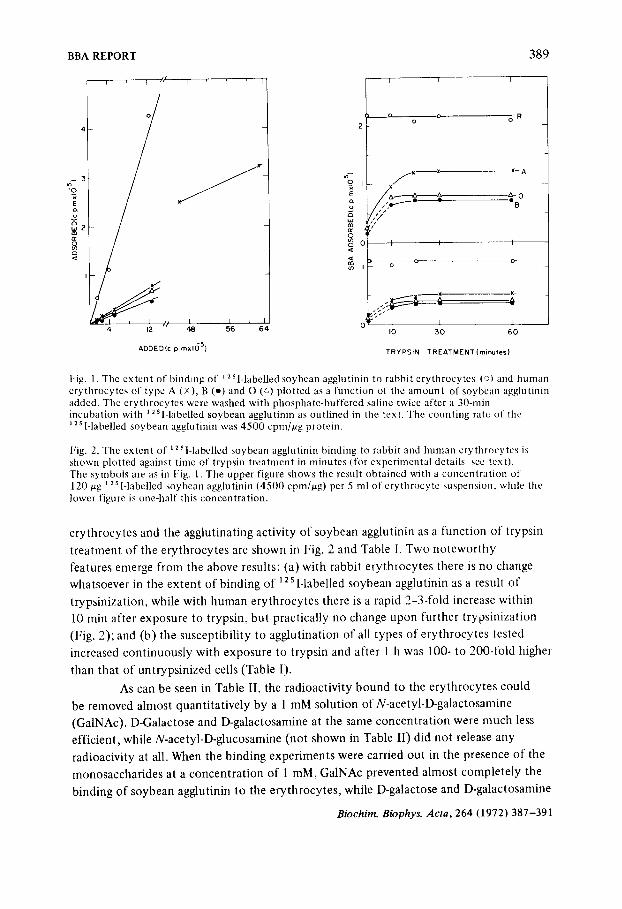
only on the ratio agglutinin:cells and to be invariant after 5 min of incubation. It can be seen from Fig. 1 that at a given soybean agglutinin concentration,
rabbit erythrocytes bound 5-6 times more soybean agglutinin per cell than did human erythrocytes. With human erythrocytes, the amount of soybean agglutinin bound was highest with type A, somewhat lower with type O and lowest with type B erythrocytes. When increasing amounts of ~2 s l-labelled soybean agglutinin were added to a constant
number of erythrocytes (30-300 IJg/5 ml erythrocyte suspension and in one
experiment with type A erythrocytes as much as 1500/ag/5 ml), the amount of radioactivity bound to the cells increased linearly with tire amount of soybean agglutinin added (Fig. 1 ). Our inability to saturate the erythrocyte with soybean agglutinin is in accord with the observations by Boyd e l al. J o on tire binding of lima bean agglutinin to human erythrocytes. At concentrations of soybean agglutinin greater than those reported here, hemolysis of the erythrocytes occurred before the experiment could be completed. The binding experiments to be described were therefore carried out with two amounts of soybean agglutinin (60/ag and 120 ,ug per 5 ml erythrocyte suspension) high enough to give marked agglutination under our conditions but without leading to significant hemolysis.
The results of experiments on the binding of 12s l-labelled soybean agglutinin to
Biochim. Biophys. Acta, 264 (1972) 387-391
BBA REPORT 389
~ 3
x E cL
o
4 12
'*'I
I J I L 48 56 64
ADDED(c p mxl6 5)
' 0 x E
u
I I I
0 o R
S x ~ - A
x
~V
- I I I
D o o o-
I0 30
I
6O
TRYPSIN TREATMENT (minules)
Fig. 1. The extent of binding of ] 2Sl-labelled soybean agglutinin to rabbit erythrocytes (o) and human erythrocytes of type A (x), B (-) and O (P~) plotted as a function of the amount of soybean agglutinin added. The erythrocytes were washed with phosphate-buffered saline twice after a 30-rain incubation with ~ 2SI-labelled soybean agglutinin as outlined in the text. The counting rate of the ;2 s I-labelled soybean agglu tinin was 4500 cpm/~g protein.
Fig. 2. The extent of ~251-1abelled soybean agglutinin binding to rabbit and human erythrocytes is shown plotted against time of trypsin treatment in minutes (for experimental details see text). 'Fhe symbols are as in Fig. 1. The upper figure shows the result obtained with a concentration of 120 ~g 12 s l-labelled soybean agglutinin (4500 cpm/~g) per 5 ml of erythrocyte suspension, while the lower figure is one-half this concentration.
erythrocytes and the agglutinating activity of soybean agglutinin as a func t ion of trypsin
t rea tment of the erythrocytes are shown in Fig. 2 and Table I. Two no tewor thy
features emerge from the above results: (a) with rabbit erythrocytes there is no change
whatsoever in the ex ten t of b ind ing of 12 s I-labelled soybean agglutinin as a result of
t rypsinizat ion, while with h u man erythrocytes there is a rapid 2-3-fold increase within
10 rain after exposure to t rypsin, bu t practically no change upon further t rypsinizat ion
(Fig. 2); and (b) the susceptibil i ty to agglut inat ion of all types of erythrocytes tested
increased con t inuous ly with exposure to t rypsin and after 1 h was 100- to 200-fold higher
than that of unt ryps in ized cells (Table I).
As can be seen in Table II, the radioactivi ty b o u n d to the erythrocytes could
be removed almost quant i ta t ive ly by a 1 mM solut ion of N-acetyl-D-galactosamine
(GalNAc). D-Galactose and D-galactosamine at the same concen t ra t ion were much less
efficient, while N-acetyl-D-glucosamine (not shown in Table II) did no t release any
radioacivity at all. When the b ind ing exper iments were carried out in the presence of the
monosaccharides at a concen t ra t ion of 1 raM, GalNAc prevented almost complete ly the
b inding of soybean agglutinin to the erythrocytes , while D-galactose and D-galactosamine
Biochim. Biophys. Acta, 264 (1972) 387-391

390 BBA REPORT
TABLE 1
ACTIVITY OF PURIFIED SOYBEAN AGGLUTININ AS A FUNCTION OF TRYPSINIZATION OF ERYTHROCYTES
Agglutinating units/rag soybean agglutinin *
Rabbit ttuman, O,pe
A B
Untreated 40 2 1 2 Trypsinized 10 rain 1200 65 30 60
20 rain 4000 30 rain 6000 120 65 200 60 min 8500 340 100 400
*Agglutinating activity determined by the spectrophotometric method of Liener 2' 9. Results are estimated to be plus or minus 0.5 unit for the untreated human erythrocytes and plus or minus 10% for the remaining values.
TABLE II
RELEASE OF ,2 S I_LABELLED SOYBt';AN AGGLUTIN1N FOLLOWING EXPOSURE OF ERYTHROCYTI".S TO SUGARS
Washed erythrocytes with a known amount of bound radioactive soybean agglutinin were suspended in 5 ml of a 1 mM solution of the indicated sugar in phosphate-buffered saline. After 30 rain incubation at room temperature, the erythrocytes were centrifuged, washed 2 times with 5 ml of phosphate-buffered saline and the residual bound radioactivity determined. The release of radioactivity is expressed as % of radioactivity originally bound. Not shown is the finding that the time of trypsin treatment from 10 rain to 1 h is without effect on the percent release of the 12 S l.labelled soybean agglutinin.
Erythrocytes Release o f ~ 2 Si.labelled soybean agglutinin (%)
D-GalNAc D-Galactosambte D-Galaetose added added added
Rabbit Untreated 90 10 6 Trypsinized 74 13 7
Hi,matt Type A: Untreated 84 34 37
Trypsinized 93 Type B: Untreated 85 32 39
Trypsinized 94 20 30 Type O: Untreated 90 32 40
Trypsinized 97 20 25
caused only 30 -40% inhibi t ion . These results are in comple t e agreement wi th the
previous findings on the saccharide-specif ic i ty o f soybean agglutinin 2.
I1 seems clear f rom these studies that the increased agglutinabil i ty o f rabbit
e ry th rocy t e s fol lowing t ryps in t r ea tmen t is no t the direct result o f an increase in the
n u m b e r o f b inding sites for soybean agglutinin on the cell. With h u m a n e ry th rocy t e s we do
Bioehim. Biophys. Aeta, 264 (1972) 387-391

BBA REPORT 391
observe an increase in the number of sites with binding affinities apparently similar to
those of the sites present on the untreated erythrocytes, yet even here the increase in
the number of sites for soybean agglutinin does not parallel the progressive and dramatic
increase in agglutinability upon continuing exposure of the cells to trypsin. We conclude,
therefore, that factors other than an increase in the number of soybean agglutinin binding
sites are generally controlling the increased agglutinability of trypsin-treated erythrocytes.
Similar conclusions have recently been reached from studies on the binding of
radioactively labelled agglutinins to normal and transformed somatic cells s ' 11,12
Soybean agglutmin interacts with human blood cells of type A, O and B and
with rabbit erythrocytes; these interactions are specifically inhibited by GalNAc and
related sugars. This observation can be explained along the usual lines 13 by suggesting
that soybean agglutinin interacts with erythrocyte surface receptors containing GalNAc
which need not necessarily be part of the blood group substance of type A. Moreover,
the lack of b lood type specificity may be the result of the inabili ty of soybean agglutinin
to distinguish between a and/3 linked GalNAc. In type A blood group substance,
GalNAc is c~ linked, whereas it is possible that on the erythrocyte surface it occurs both
a and/3 linked. Yet one should not overlook the possibility that soybean agglutinin
contains functionally linked but topographically independent sites for binding GalNAc
and for at tachment to the erytbrocyte surface.
This study was supported by Grant FG-Is-247 from the U.S. Department of
Agriculture. One of us (J.A.G.) would like to thank Professor E. Katchalski for support
and facilities and also to thank the American Cancer Society for fellowship support
graciously given.
REFERENCES
1 0 . M~kel~,Ann. Med. Exp. Biol. Fenn., 35 (1957) suppl. 11. 2 H. Lis, B. Sela, L. Sachs and N. Sharon, Biochim. Biophyx. Aeta, 211 (1970) 582. 3 G.I. Pardoe and G. Uhlenbruck, J. Med. Lab Teehnol., 27 (1970) 249. 4 0 . Prokop, G. Uhlenbruck and W. Kohler, Vox Sang., 24 (1968) 321. 5 G.L. Nicolson,NatureNew Biol., 233 (1971) 244. 6 H. Lis, N. Sharon and E. Katchalski, J. Biol. Chem., 241 (1966) 684. 7 F.C. Greenwood, H.H. Hunter and J.S. Glover, Bioehem. J., 89 (1965) 114. 8 B. Sela, H. Lis, L. Sachs and N. Sharon, Biochim. Biophys. Acta, 249 (1971) 564. 9 I.E. Liener, Arch. Biochem. Biophys., 54 (1954) 223.
10 W.C. Boyd, H.M. Bhata, M.A. Diamond and S. Matsubara, J. lmmunol., 89 (1962) 463. 11 M.J. Cline and D.C. Livingstone, Nature, 232 (1971) 155. 12 B. Ozanne and J. Sambrook, Nature, 232 (1971) 156. 13 G.I. Pardoe and G. Uhlenbruck, Med. Lab. Technol., 28 (1971) 1.
Biochim. Biophys. Acta, 264 (1972) 387-391