bilirubin uptake in vitro by the rat intestinal mucosa
TRANSCRIPT

Biochimica et BiophyMca Actl, 291 (1973) 308-314
© Elsevier Scientific Publishing Company, Amsterdam - Printed in The Netherlands
BBA 7 6144
BILIRUBIN UPTAKE I N V I T R O BY THfi RAT INTESTINAL MUCOSA
JUAN LELIO CORCHS, RAQUEL ELENA SER~ ANI AND EMILIO RODRIGUEZ GARAY
Depavtamento de Ciencias Fisioldgica, s, Facultad de Ciencias Mddicas, Universidad Nacional de Rosario, Santa Fd 3too, Rosario (Arg,;ntina)
(Received June 6th, 1972)
SUMMARY
Flat sheets of rat jejunum incubated in the presence of unconjugated bilirubin solutions were shown to incorporate bilirubin iato the tissue.
Bilirubin mucosal uptake, expressed ~.s a function of the incubation time showed a tendency to reach a constant level within 12o min.
Solutions of bilirubin in sodium tauroc'~olate gave an incorporation of signifi- cantly greater amounts of bilirubin than those, prepared with albumin.
A structurally similar substance (biliver 5fin) inhibited bilirubin uptake in a way that suggested competitive inhibition. The res,.dts support the view that the mechan- ism of bilirubin uptake by the rat intestinal rqucosa cannot be entirely explained by simple passive diffusion.
INTRODUCTION
It has been demonstrated in vivo that L:aconjugated bilirubin ~.an be absorbed from the intestinal lumen into the blood and that it can also pass from the plasma into the intestinal lumen x-4. Recent experiments hz vitro have shown that lq2-1abelled unconjugated bilirubin can cross the intestinal wall when placed inside normal or everted intestinal segments. The unidirectional fluxes were linear with concentration within the range from o.oi to o.I mM and were approximately equal in both direc- tions 5.
In order to obtain further insight into -he mechanism by which uneonjugated bilirubin is transported across the gut wall, the uptake of pigment by isolated sheets of rat jejunum was investigated using an in v, 'to tissue accumulation technique ~.
MATERIALS AND METHODS
Chemicals
Bilirubin, biliverdin, sodium taurocholate and bovine serum albumin were obtained from Sigma (St. Louis, Mo., U.S.A). All the other reagents used were of analytical grade.
Preparation of tissue
Adult Wistar rats of both sexes were killed by cervical dislocation. The proximal

BILIRUBIN UPTAKE BY I N T E S T I N E in vitro 309
jejunum was removed and kept in Ringer-Krebs solution ~. Segments about I -2 cm in length were cut open to expose the mucosa. The segments were then incubated at 37 °C in a chamber provided with a reflux condenser.
Incubation conditions Unconjugated bilirubin was dissolved in O.lO-O.15 ml o.i M NaOH and then
added to Ringer's solution (equilibrated with O:-CO 2 (95.5, v/v)) containing (I) bovine serum albumin to give a molar ratio of bilirubin to albumin of 2: I (bilirubin- albumin solution), (2) 5 mM sodium taurochola~e to give a molar ratio of I : I6 .~ (bilirubin-taurocholate solution). Unless otherwise stated, the final concentration of bilirubin in the incubation medium was 0.3 mM. The amount of wet tissue added ranged from IOO to 15o mg for 5 m! medium. During incubation stirring was achieved using the gas mixture.
Biliverdin was dissolved in o.I nil of sodium carbonate solution (47.2 mM Na~CO a, 88. 9 mM NaCI) and then mixed with the incubation medium.
At the end of the incubation period, the segments were removed, rinsed twice with cold Ringer's solution and gently blotted on filter paper.
Mucosal and serosal sheets were obtained as described by Dickens and Weil- Malherbe 9, weighed on a Mettler balance and homogenized in citrate-phosphate buffer pH 2.2 (ref. xo) to give IOO mg wet tissue per 2.5 ml buffcr.
After 12o min of incubation the bilirubin concentration in the medium had decreased by IO % of its initial value in bilirubin- albumin experiments and 2o % in the bilirubin-taurocholate experiments.
Analytical proccdures Determination of bilirubin concentration in the homogenates was carried out
by a modification of Hargreaves' method n using a dilute diazo reagenO g. Tile absorbance of the stlperna:ant was determined at 54 ° n m in a DU Be~-km:,n
spectrophotometer. A 91 + 3 % (n = 6) recovery of the bilirubin added to normal homogenates was obtained.
In some experiments the diazotized supernatant was concentrated ~3 and chromatographed on pape# 4.
Total tissue water in the unsuited specimens was calculated by the differen between wet and dry weight (24 h at IOO °C). Water content was found t(, be con3tant, with a value of 0.8o ml/g wet tissue duriltg the entire period of incubatior:.
The mucosal extracellular space was determined with inulin t~ and gave a value of 0.25 m l ' g -1 wet tissue (S.E. -4- o.o2i (~ = 18).
Oxygen consumption (Qo,) was determined by Warburg's method I~ and expressed as pl O~/mg protein per h. Protein wa_~ determined by Lowrv':~ technique l~.
RESULTS
Bilirubin upgake by nmcosal a;~:d seros,zl '.;hoots I t was found that the uptake of bilirubin by the mucosal fraction was greater
than tha t of the serosa (Fig. I). A difference was detectable after 15 rain and was significantly greater after 12o minute~; incubation (P < o.oi).
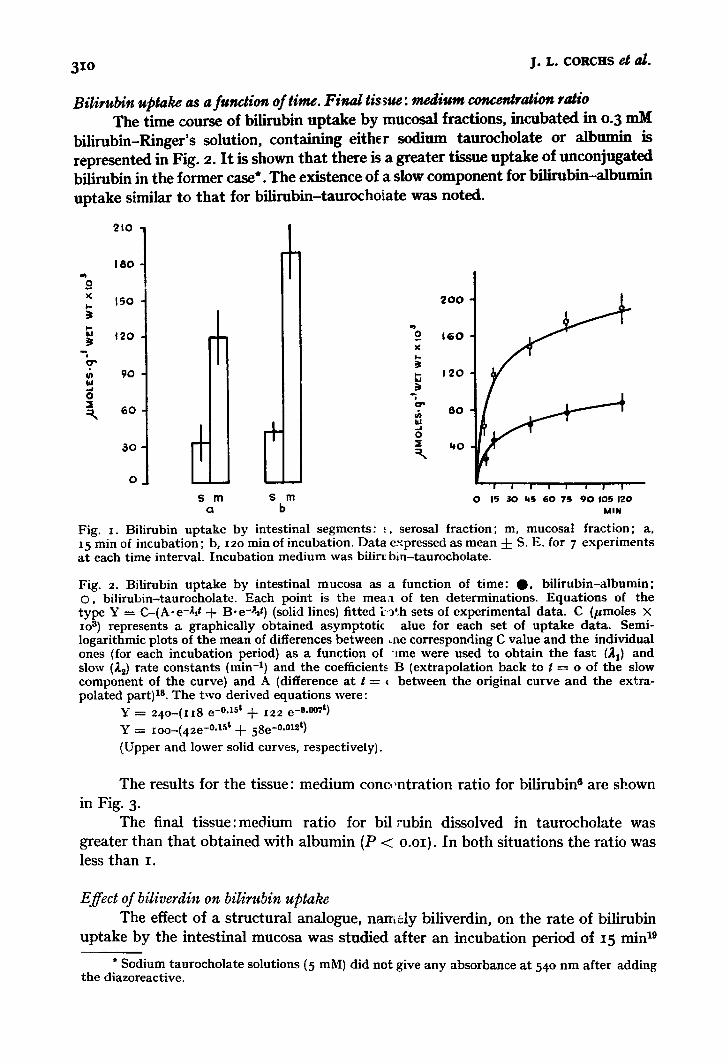
3xo j .L. CORCHS et al.
Bilirubin uptake as a funaion of time. Final tissue: medium concentration ratio The time course of bilirubin uptake by mucosal fractions, incubated in o.3 mM
bilirubin-Ringer's solution, containing either sodium taurocholate or alb,mln is represented in Fig. 2. It is shown that there is a greater tissue uptake of unconjugated bilirubin in the former case*. The existence of a slow component for bilirubin-albumin uptake similar to that for bilirubin-taurochoiate was noted.
t - ILl
'~10
180
150
120
90 I d
IE
30
0
I
I rl 2OO
~0 IGO
120
? o- 8 o
~, .o
S m s m o t~ so ~5 soT~ 901o~tzo ca b u,~
Fig. x. Bilirubin u p t a k e b y intest inal s egmen t s : .,, serosal f rac t ion ; m, mucosa l f rac t ion; a, 15 min of incuba t ion ; b, i2o min of incubat ion . Da ta expressed as mean 4- S. E. for 7 exper imen t s a t each t ime interval . I n c u b a t i o n m e d i u m was bflir~ b in - t au rocho la t e .
Fig. 2. Bilirubin u p t a k e b y intest inal m u c o s a as a func t ion of t ime : O , b i l i r ub in - a lbumin ; O, b i l i rubin- taurochola tc . Each po in t is the m e a a of t en de te rmina t ions . E q u a t i o n s of the t y p e Y = C- (A-e -&t + B . e -~ t ) (solid lines) f i t ted [-,~th sets of exper imenta l da ta . C (/~moles X xo 3) represents a g raphica l ly ob ta ined a s y m p t o t k alue for each set of u p t a k e da ta . Semi- logar i thmic plots of the mean of differences be tween ,.he co r re spond ing C value and t he individual ones (for each incuba t ion period) as a func t ion of t r ee were used to ob ta in t he fast (~1) and slow (~2) ra te c o n s t a n t s (min -x) and the coefficients B (ex t rapola t ion back to 1 = o of the slow c o m p o n e n t of the curve) and A (difference a t t = ~ be tween t he original cu rve a n d the ex t ra- po la ted part) Is. The two der ived equa t ions were:
Y = 2 4 o - ( z i 8 e -0"lSt + I22 e -0-o07t)
Y = ioo-(42e-0.15t + 58e -°.°t2t)
(Upper and lower solid curves, respect ively) .
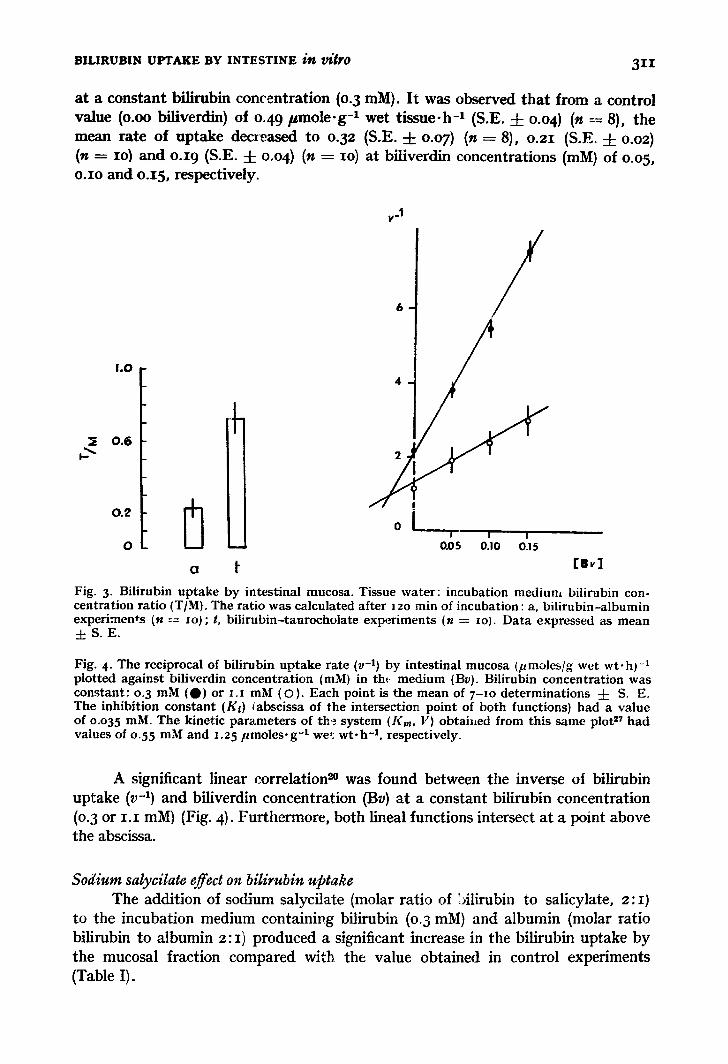
The results for the tissue: medium cono~ntration ratio for bilirubin e are shown in Fig. 3.
The final tissue:medium ratio for bil rubin dissolved in taurocholate was greater than that obtained with albumin (P < o.oi}. In both situations the ratio was less than I.
Effect of biliverdin on biIirubin uptake The effect of a structural analogue, namely biliverdin, on the rate of bilirubin
uptake by the intestinal mucosa was studied after an incubation period of x5 min 19
* Sodium t a u roc ho l a t e solutions (5 mM) did no t give a n y absorbance a t 54 ° n m af te r add ing the diazoreact ive.
BILIRUBIN UPTAKE BY INTESTINE i~ /)/tr0 3 I I
at a constant bilirubin con('entration (0. 3 raM). I t was observed tha t from a control value (o.oo biliverdin) of o.49 pmole .g -1 wet tissue-h -1 (S.E. 4- o.o 4) (n := 8), the mean rate of uptake decreased to o.32 (S.E. -4- o.o7) (n = 8), o.21 (S .E 4- o.o~) (n = IO~ and o.i 9 (S.E. 4- o.04) (n = IO) at biliverdin concentrations (mlVl) of o.o5, o.Io and o.15, respectively.
1.0
0 .6
v-1
2
°
0 l I I I
o - - o.os o.lo o.ls
a !" [ m ~ ]
Fig. 3. Bilirubin uptake by intestinal mucosa. Tissue water : incubation medium bilirubin con- centration rat io (T/M). The ratio was calculated after 12o rain of incubation: a, bi l irubin-albumin experiments (n --=- lo); t, bi l i rubin-taurocholate experiments (n = zo). Data expressed as mean d: S. E.
Fig. 4- The reciprocal of bilirubin uptake rate (v -1) by intestinal mucosa (pmoles/g wet wt-h) -1 plotted against biliverdin concentrat ion (raM) in tht medium (By). Bilirubin concentration was constant : 0.3 mM (O) or i . i mM (O). Each point is the mean of 7 - I o determinations 4- S. E. The inhibition constant (Ks) tabscissa of the intersection point of both functions) had a value of o.o35 mM. The kinetic parameters of th ~ . system (Kin, V) obtai~ed from this same plot 2¢ had values of o.55 mM and 1.25 pmoles .g - t we1: w t . h -1, respectively.
A significant linear correlation ~n was found between the inverse oi bilirubin uptake (v -1) and biliverdin concentration (By) at a constant bilirubin concentration (o.3 or I . I mM) (Fig. 4). Furthermore, both lineal functions intersect at a point above the abscissa.
Sodium salycilate effect on bilir.ubin uptake The addition of sodium salycilate (molar ratio of :Alirubin to salicylate, 2:1)
to the incubation medium eontainivg bilirubin (o.3 mM) and albumin (molar ratio bilirubin to albumin z: I ) produced a significant increase in the bilirubin uptake by the mucosal fraction compared with the value obtained in control experiments (Table I).

3 i 2 J.L. CORCltS et d .
T A B L E I
Influence of a lbumin and organic anions on bilirubin u p t a k e by intestinal mucosa. Bilirubin concentrat ion was o.3 raM. The mola r rat io of b i l i rub in-a lbumin and b i l i rubin-sodium salicylate was 2 : i . Concentra t ion of sodium taurochola te was 5 mM. In parentheses are the number of experiments .
Bilirubin uptake {lamoles.g-X wet tissue wt per 45 min~
Bilirubin + a lbumin Bilirubin + a lbumin + sodium salycilate Bilirubin + sodium taurocholate
0.062 4- 0.007 (II) o.Io3 + o.oxo (I2) o.I48 -4- 0.009 (7)
Viability of tissue preparation Oxygen consumption of mucosal sheets was followed in a Warburg respirometer
during a I-h incubation period in t h e , cesence of bilirubin (0.3 mM), sodium tauro- cholate (5 mM) and Ringer's solution after a preincubation time of 3o min in the same medium. The mean value was 13.72 + S.E. 2.1, n = 9 (/~1 O2-mg -t proteirJ.h -l) which did not differ significantly from that obtained in a control group using tissue slices incubated only in Ringer's solution (13.78, S.E. + 1.45, n = IO).
DISCUSSION
In these experiments bilirubin was determined in the _;ntestinal mucosa by a simple and reproducible method. The pigment taken up by the tissue was identified as unconjugated bilirubin because: (a) the unincubated mucosa did not contain any pigment giving a direct diazoreaction, (b) paper chromatography of diazotized homogenates (after incubation) showed only the presence of azopigment A, correspond- ing to unconjugated bilirubin 14.
The distribution of bilirubin (in bilirubin-taurocholate experiments) taken up at 15 and 12o min by the serosal and mucosal fractions is reported in Fig. i. The taurocholate-bilirubin complex is mainly concentrated in the mucosa. Since serosal concentrations attained a constant level after 15 min the possibility that adsorption might be (wholly or in part) the responsible mechanism was not discarded. The relevant point, however, is that while the serosal concentration has reached a cons.tant level during the first 15 min, the mucosal concentration is still increasing after 2 h. This supports the view that the location of the slow component of the uptake process is within the mucosal part of the gut wa!l.
The time course of bilirubin uptake by mucosa (Fig. 2) suggests the existence of at least two different pigment distribution spaces. This is supported by: (a) the fraction taken up by the extracellular space cannot fully account for the total uptake by tissue. If we consider the bilirubin concentration (0.3 mM) in our incubation medium and the value of 0.25 ml ' g -1 wet tissue for extracellular space, it is easily shown that only 0.075/~moles. g-~ wet tissue will reach equilibrium in this space x~dth the incubation medium; (b) unconjugated bilirubin has been detected by histoche- mical methods within epithelial mammalian cells 2x, 2,; (cl data of bilirubin uptake as a function of time (Fig. 2) showed that the curves were composed of two exponential

BILIRUBIN UPTAKE BY INTESTINE i~ Vitt'O 313
terms. This suggests the existence of more than one tissue bilirubin distribution compartment ~a.
In the experiments with bilirubin-taurocholate the tendency to reach a con- stant value with the incubation time (Fig. 2) could be explained as being due to saturation of b'dirubin binding sites in the tissue. }~owever, a similar behaviour was found in bilintbin-albnmin experiments, where the equilibrium takes place with significantly lower levels of bilirubin uptake. Since we can accept that the number of sites which tightly bind b'flirubin would be same in both experimental conditions, this interpretation was discarded. I t seemed more reasonable to interpret the results as indicating that the amount of substrate taken up by the tissue was the expression of a net flux.
The addition of sodium salicylate to the incubation medium produced an increase in pigment uptake (Table I). This was considered to occur as a result of an increase of unbound pigment concentration owing to the displacement of bilirubin from its binding to albumin za. In the experiments where sodium taurocholate was used the increased bilirubin uptake seemed to be due to a similar factor 2a, since in the presence of that salt, bilirubin is completely in the unbound form. However, the possibility of an increase in brush border membrane permeability due to taurocholate cannot be excluded ~.
The rate of bilirubin uptake by tissue was influenced by the presence of a structurally related substance such as biliverdin ~. Although from this plot (Fig. 4) we cannot differentiate between competitive and mixed inhibition, the close structural similarity between substrate (bilirubin) and inhibitor (biliverdin) makes the existence of competitive inhibition the most likely explanation 27.
Uphill transport capacity (final tissue: medium concentration ratio, > I) was not observed in any of the experiments (Fig. 3~- This suggests the presence of only passive mechanisms of bilirubin uptake.
Unidirectional fluxes of unconjugated bilirubin measured g4th the aid of :!4CZ - bilirubin 5 had a value very much greater than the fluxes measured in this work by a dlemical procedure, which could represent a net flux. This discrepancy (and the biliverdin effect on bilirubin uptake) would suggest the existence of a bilirubin transfer mechanism due to facilitated diffusion ~.
ACKNOWLEDGEMENT
This work was in part supported by a grant from the Cor.sejo Nacionai de Investigaciones Cientificas y T6cnicas, Repdblica Argentina.
J. L. C. and E. R. G. are investigators from the Consejo Nacional de Investiga- clones Cientificas y T6cnicas, Rep~blica Argentina.
R.E.S. is an investigator from the Universidad Nacional de Rosario, Repfi- blica Argentina.
REFERENCJ'2S
R. Lester and R. J. fichmid, J. Clin. hwest., 42 [i963~ 736. 2 R. Sehmid ant ~ L. Hammaker , J. Clin. l~wesl., 4z (J9o3~ I7zo. 3 R. Lester, Gastroenterology, 47 (x964) 424. 4 E. A. Rodriguez Garay, E. V. Flock and Ch. A. Owen, Am. J. Physiol., 27o (t966} 684

314 j .L . CORCitS etag.
5 J. L. Corcl~ and E. A. Rodriguez Garay, Experientia, z6 (I97 o) 858, 6 R. K. Crane and P. Mandelstam, Biochim. Biophys. Acta, 45 (t96o) 460. 7 I. L Reisin, C. A. Rotunno, L. Corchs0 V. Kowalewsky and M. Cereijido, Physiol. Chem.
Phys., 2 (I97 o) x7I. 8 J. D. Ostrow and R. Schmid, J. Clin. Invest, 42 (I963) I286. 9 F. Dickens and H. Weil-Malherbe, Biochem. J. 35 (I94t) 7-
xo T. C. McIlvaine, J. Biol. Chem., 49 (z92t) x83. Ix T. Hargreaves, Clin. Chim. Acta, II f1965) 278. 12 H. T. Malloy and K. A. Evelyn, J. Biol. Chem., IX9 (I937) 48I. 13 B. A. Noir, E. A. Rodriguez Garay and M. Royer, Biochira. Biophys. Acta. Ioo 11965) 4oJ. t 4 R. Schmid, J . Biol. Chem., 229 (1957) 881. I5 H. H. Roe, J. H. Epstein and N. P. Goldstein, J. Biol. Chem., 178 (x949) 839. z6 W. W. Umbreit, R. H. Burris and J. F. Staufer, Manometric Techniques, Chapter i0 Burgess
Publishing Co., Minneapolis, t957. 17 O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J. Randall, J. Biol. Chem., I93 {I95t) 265. x8 D. S. Riggs, The Mathematical Approach to Physiological Problems, Chapter 6, The Williams
and Wilkins Company, Waverly Press, Baltimore, x963. I9 L. R. Finch and F. J. R. Hird, Biochim. Biophys. Acta, 43 (~96c) 268. 20 G. W. Snedecor, Statistical Methods, Chapter 6, The Iowa State University Press° Ames, Iowa,
I956. 21 S. Raia, in I. A. D. Bouchier and B. H. Billing, Bilirubin Metabolism, Blackweil Scientific
Publications, Oxford and Edinburgh, 1967, p. 285. 22 S. Grzycki and I. Kr61ikowska, Z. Mikrosk-Anat. Forsch., 79 (I968) x5I. 23 G. B. Odell, in A. Sass-Kortsak, Kemictems, University of Toronto Press, Toronto I961
p. 199. 24 M. A. M. Ali, in I. A. D. Bouchier and B. H. Biling, Billirubin Metabolism, Black~ell Scientific
Publications, Oxford and Edingburgh, 1967, p. 245. 25 R. A. Frizzell and S. G. Schultz, Biochim. Biophys. Acta, 211 (I97o) 589. 26 T. K. With, Bile Pigments, Chapters i and 9, Academic Press, NeW York, I958 27 M. Dixon and E. C. Webb, Enzymes, Chapter 8, Longmans Green and Co. Ltd. London, x964 "..8 W. D. Stein, The l~ovement of Molecules across Cell Membranes, Chapter 4, Academic Press
New York, i967 .