basement membrane integrity in oral cancer repair
TRANSCRIPT

Basement Membrane Integrity
In Oral Cancer
Presented by
Dr. Thein Tun

The basement membrane (BM) is a major functional
component of the extracellular matrix that underlies all
epithelia.
provides structural support to epithelia and forms a
mechanical connection between epithelia and underlying
connective tissue.
The integrity of the basement membrane is essential for
cellular growth, and is often altered in disease.

In general, basement membranes appear very similar
to each other by extracellular matrix (EM).
But all are not alike!
There is a wealth of molecular and functional
heterogeneity among basement membranes, due
primarily to isoform variations of basement
membrane components.

Remodeling of the BM is a normal process that occurs
throughout development.
The BM is structurally remodeled by cells during
movement and proliferation.
Breach of the BM is a critical step for malignant cells to
invade locally and metastasize.

• The role of the BM in carcinoma biology is not well
understood.
• Based mostly on morphologic data, the loss of BM
continuity has been associated with increasing
malignancy. (Bosman, 1994)

Four layers of the basement
membrane zone
(1) The basal keratinocyte layer,
(2) the lamina lucida,
(3) the lamina densa, and
(4) the sublamina densa (lamina reticularis).
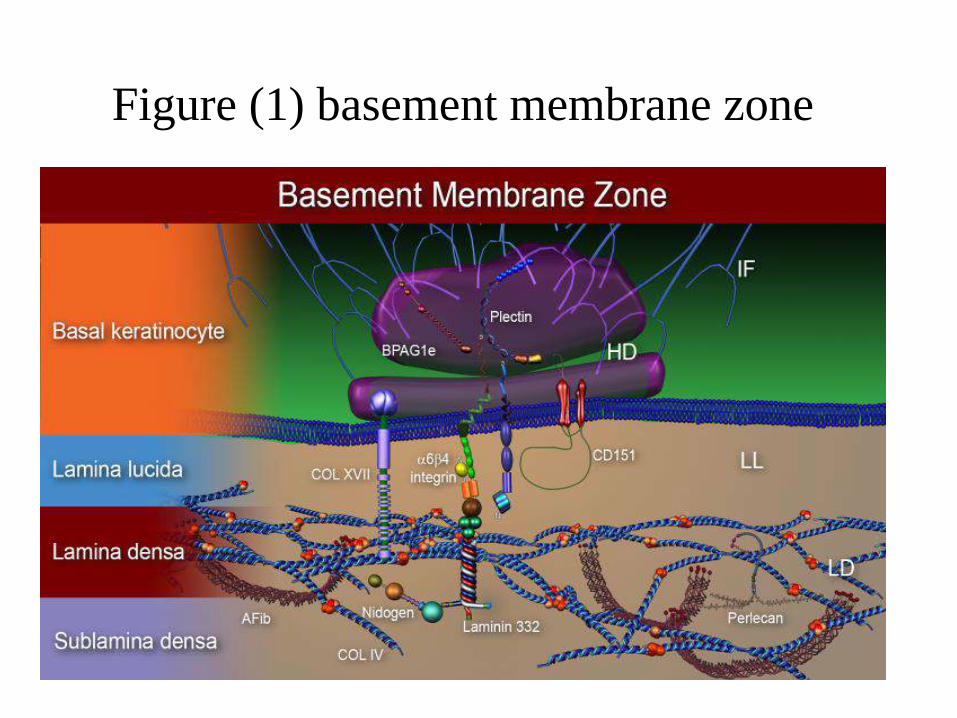
Figure (1) basement membrane zone

Within the basal keratinocyte,
• keratin intermediate filaments (IF),
• hemidesmosomes (HD),
• composed of bullous pemphigoid antigen 1 (BPAG1),
• plectin,

• Within the lamina lucida (LL),
• anchoring filaments (AF), composed predominantly
of Laminin 332.
• the extracellular portions of collagen XVII
• Integrin and CD151.

• Within the lamina densa (LD) are
• collagen IV,
• nidogen and perlecan.
• Within the sublamina densa (lamina reticularis)
• anchoring fibrils (AFib), composed of collagen VII.
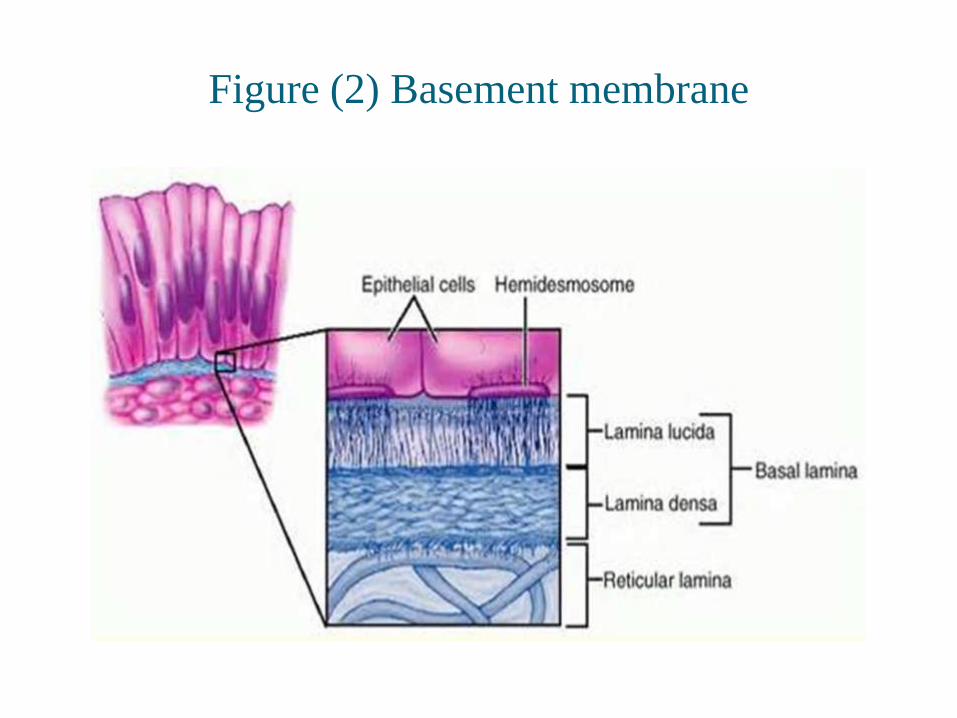
Figure (2) Basement membrane

• Lamina lucida and lamina densa together make up
basal lamina
• Within the lamina lucida, we can see thin strand-like
structures known as anchoring filaments.
• Beneath the lamina lucida is known as lamina densa.
• Beneath the lamina lucida and lamina densa is the
sublamina densa (lamina reticularis).

• The basal keratinocyte is anchored to the basal lamina
via the intermediate filaments and hemidesmosomes.
• Two types of hemidemsmosome have been identified:
• mature type I and
• immature or developing type II hemidesmosomes

• The main components of basal lamina are:
• Type IV collagen, Glycoproteins, Proteoglycans
• The Lamina Lucida is made up of laminin, integrins,
entactins, and dystroglycans.
• The Lamina Densa is made up of type IV collagen
fibers; perlecan (a heparan sulfate proteoglycan)

• Glycoproteins are variously located in the lamina lucida
and lamina densa areas. (Lee, 1988)
• Collagen VII is predominantly localized to the epidermal
basement membrane zone within the lamina densa and
sublamina densa.
• It provides structural support and stability to the
epidermal basement zone.

Many of the components of the epidermal basement
membrane zone have been linked to cancer, with
several basement membrane zone components having
a significant role in tumor invasiveness and or
prognosis. (Ko & Marinkovich, 2010)

Although cancers come in a variety of different forms,
one of the basic tenants of cancer is the fact that it has
the ability to invade through basement membranes.
• Early stages of malignancy that are thus limited to the
epithelial layer by the basement membrane are called
carcinoma in situ.

Three-Step Theory of Invasion
1. The first step is tumor cell attachment via cell surface receptors
which specifically bind to components of the matrix such as
laminin (for the basement membrane) and fibronectin (for the
stroma).(Kramer et al., 1980)
2. The anchored tumor cell next secretes hydrolytic enzymes (or
induces host cells to secrete enzymes) which can locally
degrade the matrix (including degradation of the attachment
components).

• Matrix lysis most probably takes place in a highly localized
region close to the tumor cell surface.
3. The third step is tumor cell locomotion into the region of the
matrix modified by proteolysis.
• Continued invasion of the matrix may take place by cyclic
repetition of these three steps.

• Liotta and Stetler-Stevenson have described the
following 3-step theory for the invasion of carcinoma
cells through the BM:
• (i) the tumour cells bind to the BM and extracellular
matrix;
• (ii) proteolytic enzymes degrade the BM; and
• (iii) the carcinoma cells are able to migrate and spread.
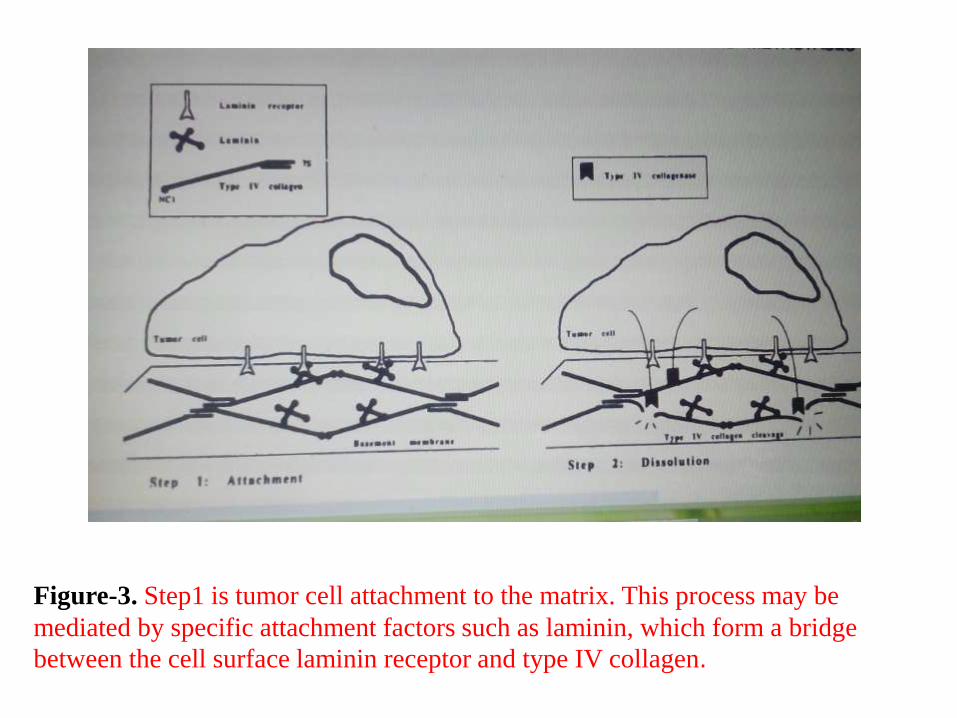
Figure-3. Step1 is tumor cell attachment to the matrix. This process may be
mediated by specific attachment factors such as laminin, which form a bridge
between the cell surface laminin receptor and type IV collagen.
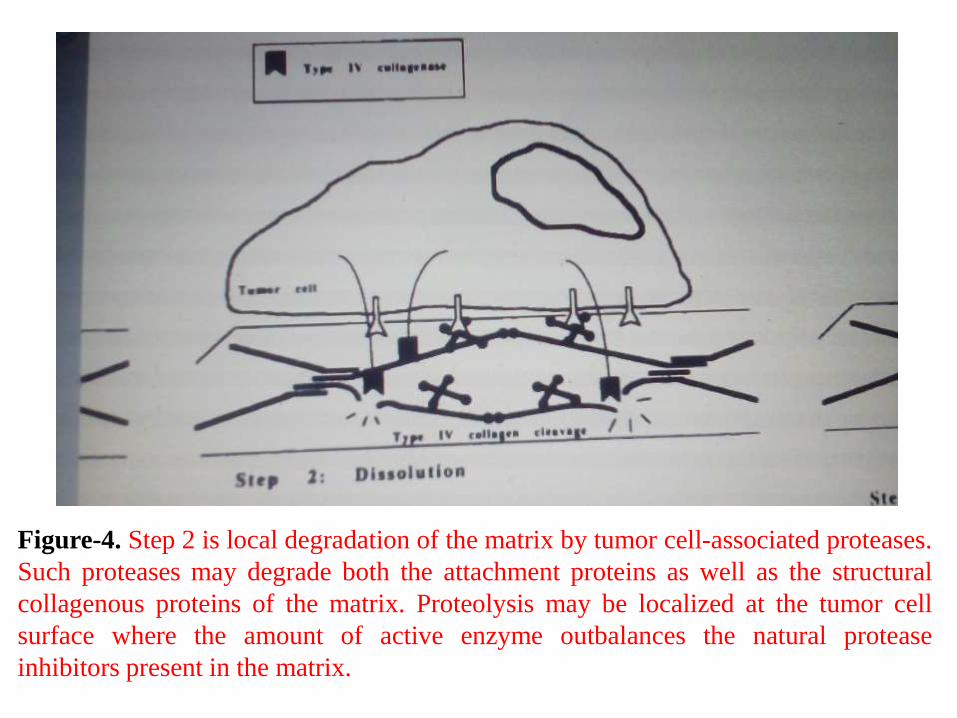
Figure-4. Step 2 is local degradation of the matrix by tumor cell-associated proteases.
Such proteases may degrade both the attachment proteins as well as the structural
collagenous proteins of the matrix. Proteolysis may be localized at the tumor cell
surface where the amount of active enzyme outbalances the natural protease
inhibitors present in the matrix.
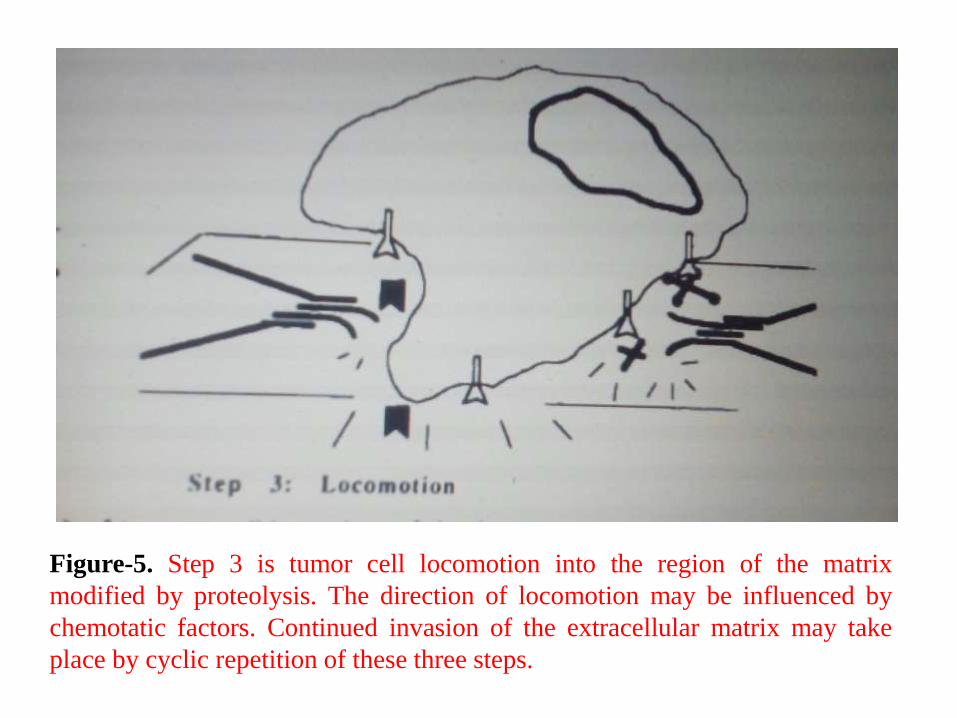
Figure-5. Step 3 is tumor cell locomotion into the region of the matrix
modified by proteolysis. The direction of locomotion may be influenced by
chemotatic factors. Continued invasion of the extracellular matrix may take
place by cyclic repetition of these three steps.

• The laminins are important integrin-binding structures
within the lamina lucida and lamina densa.
• Laminins are large glycoproteins composed of three
chains (α,β and ϒ ) bound by disulfide bonds.
• There are at least 15 different laminins, comprised of a
combination of 5 different α-chains, three β-chains, and
three γ-chains.

• laminin-332, when up regulated may serve as a
promoter of tumor invasiveness, and when down
regulated, the basement membrane zone's integrity is
altered such that tumor cells are allowed to cross the
basement membrane.
• laminin-5 (Ln-5) plays a vital role in tumour
migration and shows an increased expression in areas
of direct tumour–stroma interactions in OSCCs.
(Berndt et al., 2001)

• Laminins are important autocrine factors produced by
cancers to promote tumorigenesis.
• Invasion of OSCCs is associated with Ln-5 synthesis,
focal loss from the basement membrane, and
deposition in the stroma beneath invading carcinoma
cell complexes. (Franz et al., 2007)

• Laminin-5 is a component of the epithelial cell adhesion
complex containing hemidesmosomes, anchoring fibrils
and anchoring fibers. (Gerecke et al., 1994)
• Laminin-5 connects the hemidesmosomes with the
underlying connective tissue through type VII collagen
(Fig. 7). (Rousselle et al., 1997)
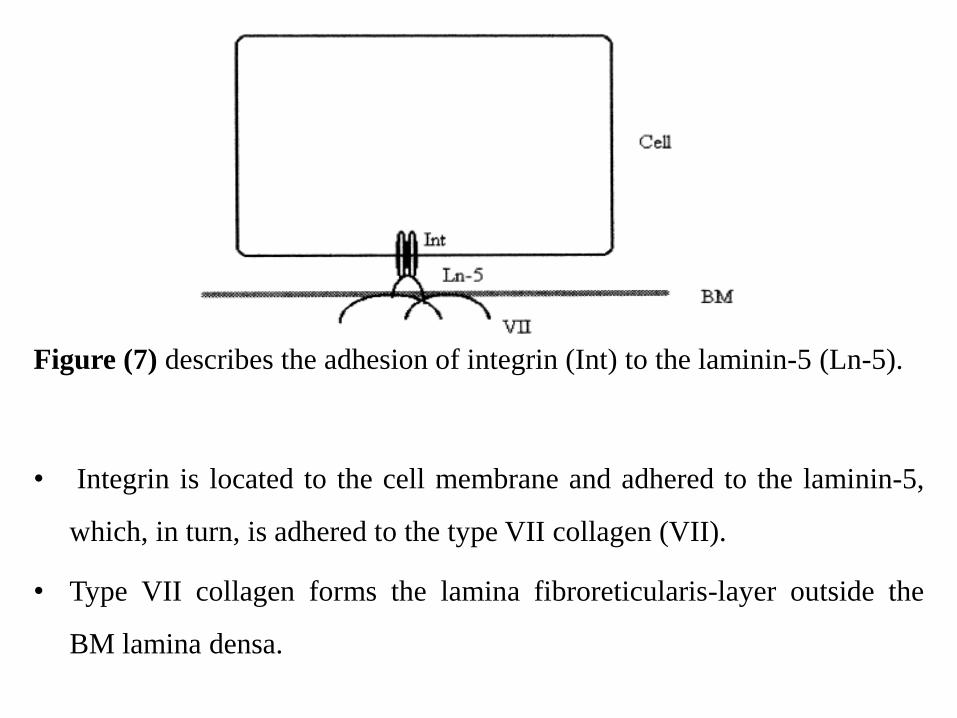
Figure (7) describes the adhesion of integrin (Int) to the laminin-5 (Ln-5).
• Integrin is located to the cell membrane and adhered to the laminin-5,
which, in turn, is adhered to the type VII collagen (VII).
• Type VII collagen forms the lamina fibroreticularis-layer outside the
BM lamina densa.

• Integrin is a two-way signaling molecule that links
the keratinocyte cytoskeleton to the extracellular
matrix.
• Integrin expression by cancer cells at the invasive
front has been related to the mode of invasion and
prognosis in OSCC.

• OSCC patients with higher expression levels of α3,
α6 and β1 integrins have significantly better
prognosis than those with lower expression levels.
• In addition, β1 integrin expression showed the highest
correlation with clinical and pathological
characteristics.(Ohara et al., 2009)

• They are necessary for maintaining epidermal
homeostatic functions such as adhesion, proliferation, and
differentiation.
• Integrin-laminin interactions maintain stem cell
populations and help to ensure their stability.
• Epithelial cells interact with the basement membrane via
integrins along their basal surface.

• Other molecules found within the lamina densa layer
of the basement membrane zone include nidogens
and perlecan.
• These molecules play a supportive role in the
stabilization of the basement membrane.(Kruegel and
Miosge, 2010)

• Nidogens play a supportive role.
• Their primary function appears to be stabilizing
interactions between laminins and collagen IV with the
lamina densa.
• Nidogens are not required for epidermal basement
membrane zone formation.

• Perlecan has also been implicated in tumorigenesis.
• Perlecan expression is increased in liver tumors, oral
tumors, and serves as a promoter of tumor growth and
angiogenesis. (Iozzo et al., 2009)

• Perlecan can serve as both an enhancer of, or an inhibitor
of angiogenesis.
• Angiogenesis is promoted by the binding of perlecan to
growth factor receptors.(Theocharis et al., 2010)
• Other basement membrane zone molecules such as
CD151, nidogen, and perlecan are associated with
tumorigenesis.

Conclusion
• During the development of invasive cancer, tumour
cells have to penetrate the extracellular matrix including
the basement membrane (BM).
• Tumour cells are hypothesized to traverse basement
membranes by a three-step process: attachment, local
degradation of the basement membrane by type IV
collagenase and other proteases, and locomotion.

• In consequence, the invasion of carcinoma cells
through the BM must be either an active process
effected by the carcinoma cells themselves or is
mediated by structural alterations of the BM occurring
during carcinogenesis and cancer progression.
• It was supposed that invading carcinoma cells are able
to degrade actively the continuous BM by secreting
collagenase and other proteolytic enzymes.

References
• Barsky SH, Siegal GP, Janotta F, et al., (1983). Loss of
basement membrane components by invasive tumors but not
by their benign counterparts. Lab Invest;49:140 –7.
• Franz M, Richter P, Geyer C, Hansen T, Acuna LD, Hyckel P,
et al., (2007). Mesenchymal cells contribute to the synthesis
and deposition of the laminin-5 gamma2 chain in the invasive
front of oral squamous cell carcinoma. J Mol
Histol;38:183-90.

• Gerecke DR, Gordon MK, Wagman DW, et al., (1994).
Hemidesmosomes, anchoring filaments and anchoring fibrils:
components of a unique attachment complex. In: Yurchenko
PD, Bird DE, Mecham RP, eds. Extracellular matrix assembly
and structure. New York: Academic Press,. 417–39.
• Iozzo RV, Zoeller JJ, Nystrom A (2009). Basement membrane
proteoglycans: Modulators Par Excellence of cancer growth
and angiogenesis. Molecules and Cells. May;27(5):503-513.

• Ko MS, Marinkovich MP (2010). Role of Dermal-Epidermal
Basement Membrane Zone in Skin, Cancer, and
Developmental Disorders. Dermatologic Clinics.;28(1):1-16.
• Kramer, R. H., Gonzalez, R., and Nicolson, G. L (1980).
Metastatic tumor cells adhere preferentially to the extracellular
matrix underlying vascular endothelial cells. Int. J. Cancer,
26: 639-642.

• Kruegel J, Miosge N (2010). Basement membrane components
are key players in specialized extracellular matrices. Cell Mol
Life Sci. Sep;67(17):2879-2895.
Lee AK (1988). Basement membrane and endothelial antigens:
Their role in evaluation of tumor invasion and metastasis. In:
Delellis RA, ed. Advances in immunohistochemistry. New
York: Raven Press, :363-393.

• Liotta LA, Stetler-Stevenson WG(1991). Tumor invasion and
metastasis: an imbalance of positive and negative regulation.
Cancer Res;51:5054–9.
• Liotta, L.A., Kleinerman, J., Catanzara, P., and Rynbrandt, D
(1977). Degradation of basement membrane by murine tumor
cells. J. Nati. Cancer Inst., 58: 1427-1439,.

• Ohara T, Kawashiri S, Tanaka A, Noguchi N, Kitahara H,
Okamune A, et al., (2009). Integrin expression levels correlate
with invasion, metastasis and prognosis of oral squamous cell
carcinoma. Pathol Oncol Res;15:429-36.
• Rousselle P, Keene DR, Ruggiero F, et al., (1997). Laminin 5
binds the NC-1 domain of type VII collagen. J Cell Biol; 138:
719 –28.

• Terranova, V. P., Rohrbach, D. H., and Martin, G. R.(1980).
Role of laminin in the attachment of PAM 212 (epithelial) cells
to basement membrane collagen. Cell, 22: 719-726,.

Thank you