bacteriophages against biothreat bacteria: diagnostic ... · biothreat bacteria: diagnostic,...
TRANSCRIPT

Open AccessReview Article
Filippov et al., J Bioterr Biodef 2013, S3 DOI: 10.4172/2157-2526.S3-010
ISSN:2157-2526 JBTBD, an open access journal Advances in Biosciences: BioterrorismJ Bioterr Biodef
therapy of these infections. Many bacterial viruses (bacteriophages or phages) active against Y. pestis, B. anthracis and Brucella species have been described. Characteristics and practical applications of such phages are the subject of this review. Due to the lack of data on lytic phages of F. tularensis and very limited information on practical importance of phages capable of lysing B. pseudomallei and B. mallei, this review does not cover the literature on these bacteria.
General Characteristics of Phages Active against Biothreat Bacteria
Keywords: Yersinia pestis; Bacillus anthracis; Brucella; Bacteriophages;Phage diagnostics; Phage typing; Phage decontamination; Phage therapy
Abbreviations: ATP: Adenosine Triphosphate; CDC: Centers forDisease Control and Prevention; CFU: Colony-forming Unit; EOP: Efficiency of Plating; FDA: Food and Drug Administration; FSU: Former Soviet Union; h: hour; LD50: 50% Lethal Dose; LPS: Lipopolysaccharide; PCR: Polymerase Chain Reaction; qPCR: Quantitative Real-time Polymerase Chain Reaction; PFU: Plaque-forming Unit; RTD: Routine Test Dilution; sbsp.: subspecies; UA: Unavailable
IntroductionBioterrorism or biothreat agents are highly infectious and pathogenic
microorganisms (bacteria, viruses, and fungi) and their toxins that can be used by individuals or groups of terrorists or as biowarfare agents in military operations. The Centers for Disease Control and Prevention (CDC) has established two groups of the most dangerous biothreatagents, categories A and B [1,2]. The highest-priority category A, inaddition to botulinum toxin and several especially dangerous viruses,includes three biothreat bacteria, the causative agents of bubonicand pneumonic plague (Yersinia pestis), anthrax (Bacillus anthracis)and tularemia (Francisella tularensis). Category B comprises severaltoxins, viruses and bacterial pathogens including the causative agentsof brucellosis (Brucella species), glanders (Burkholderia mallei), andmelioidosis (Burkholderia pseudomallei). In this review, we focus onplague, anthrax and brucellosis. Plague [3] and anthrax [4] are severefulminant primarily zoonotic infections that can be transmitted tohumans and easily disseminated in human populations, mainly vianatural or artificial aerosolization of the bacteria, resulting in epidemicswith high mortality rates. Brucellosis is a globally spread zoonosis whichis also prevalent in humans in the form of severe systemic disease. Thereare four species of Brucella pathogenic for humans, Brucella melitensis,Brucella suis, Brucella abortus, and Brucella canis [5]. The serious publichealth concern about these biothreat infections is aggravated by theemergence of multidrug-resistant strains of Y. pestis [6,7] and isolatesof Brucella resistant to some clinically relevant antibiotics [8,9], as wellas by easy induction of antibiotic resistance in B. anthracis [10], anda potential for the application of genetically engineered multidrug-resistant strains of biothreat bacteria in biological attacks [11-13].The severity of these biothreat bacterial infections requires efficientbiosurveillance and biodefense, including availability of a rich arsenalof up-to-date methods of rapid detection and identification of thebacteria, strain characterization, diagnostics, efficient prophylaxis, and
*Corresponding author: Andrey A Filippov, Department of Emerging Bacterial Infections, Bacterial Diseases Branch, Walter Reed Army Institute of Research, Silver Spring, Maryland, USA, E-mail: [email protected]
Received December 27, 2012; Accepted January 18, 2013; Published January 21, 2013
Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Copyright: © 2013 Filippov AA, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits un-restricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic ApplicationsAndrey A Filippov*, Kirill V Sergueev and Mikeljon P NikolichDepartment of Emerging Bacterial Infections, Bacterial Diseases Branch, Walter Reed Army Institute of Research, Silver Spring, Maryland, USA
AbstractPlague, anthrax and brucellosis are severe bacterial infections presenting a serious threat to public health. Their
causative agents can be weaponized and a number of drug-resistant strains have been described. This requires improvement of existing and development of new methods of diagnostics, strain characterization, prophylaxis and therapy of these infections. This review article focuses on lytic bacteriophages (phages) active against Yersinia pestis, Bacillus anthracis and Brucella including the uses of phages for diagnostics, strain typing, specific decontamination, and antibacterial therapy.
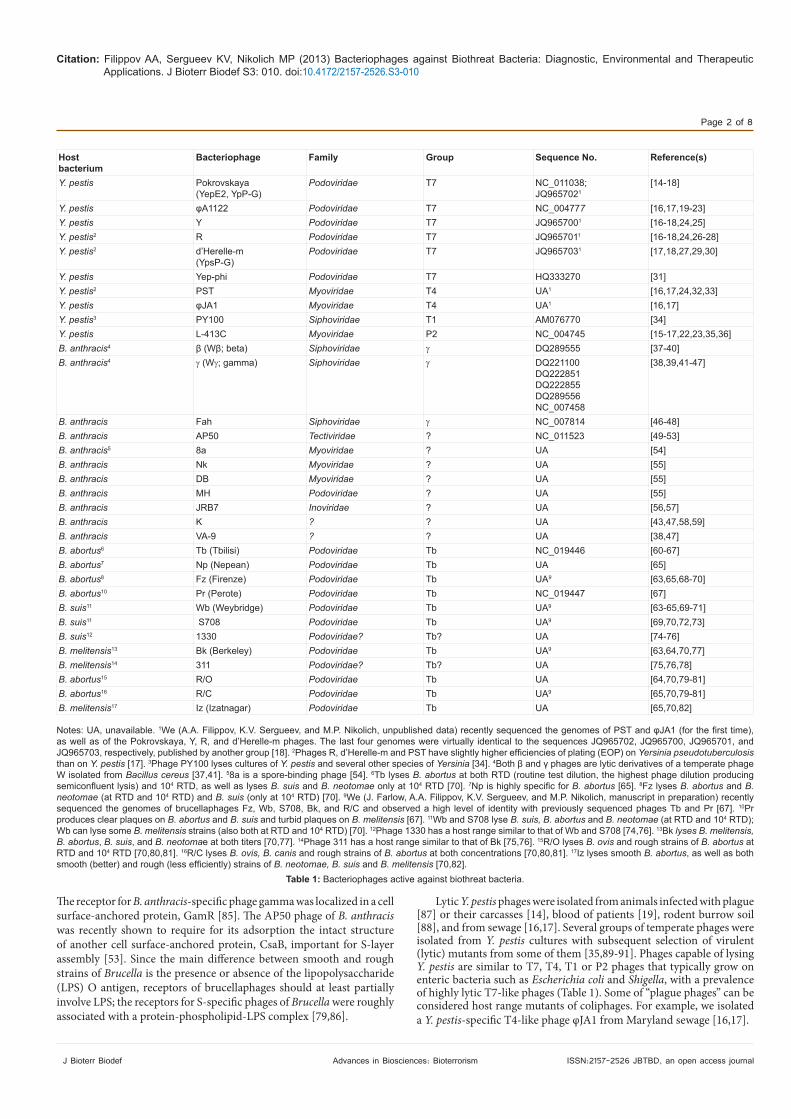
The Most important bacteriophages lytic for Y. pestis, B. anthracis and Brucella species are listed in table 1. The listed bacteriophages are members of the same order, Caudovirales (tailed viruses) but belong to five different families (Myoviridae, Podoviridae, Siphoviridae, Tectiviridae, and Inoviridae) and different groups. The first three families have different tail structures: long contractile, short noncontractile, or long noncontractile tails, respectively. The features of the Tectiviridae phages are a double coat and a tail equivalent, and the Inoviridae family representatives have a filamentous structure. Phages of these families contain linear double-stranded DNA, except the Inoviridae members which carry circular single-stranded DNA [83]. The genomes of many phages active against biothreat bacteria were sequenced (Table 1). The bacterial cell surface receptors were identified for Y. pestis-specific phages φA1122 [16,84], Pokrovskaya, L-413C, Y, φJA1, R, and PST [16], mainly in different sugar residues of the LPS outer and inner core. It was found that φA1122 is specific for Y. pestis when grown at 26-28°C but can efficiently lyse both Y. pestis and Yersinia pseudotuberculosis (the closest phylogenetic relative of Y. pestis) at 37°C [17,20-22,84], due to repressed production of O-antigen which blocks the Y. pseudotuberculosis phage receptor in the LPS inner core when grown at lower temperatures [84].
`
Jour
nal o
f Bioterrorism&Biodefense
ISSN: 2157-2526
Journal of Bioterrorism & Biodefense
Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 2 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
The receptor for B. anthracis-specific phage gamma was localized in a cell surface-anchored protein, GamR [85]. The AP50 phage of B. anthracis was recently shown to require for its adsorption the intact structure of another cell surface-anchored protein, CsaB, important for S-layer assembly [53]. Since the main difference between smooth and rough strains of Brucella is the presence or absence of the lipopolysaccharide (LPS) O antigen, receptors of brucellaphages should at least partially involve LPS; the receptors for S-specific phages of Brucella were roughly associated with a protein-phospholipid-LPS complex [79,86].
Lytic Y. pestis phages were isolated from animals infected with plague [87] or their carcasses [14], blood of patients [19], rodent burrow soil [88], and from sewage [16,17]. Several groups of temperate phages were isolated from Y. pestis cultures with subsequent selection of virulent (lytic) mutants from some of them [35,89-91]. Phages capable of lysing Y. pestis are similar to T7, T4, T1 or P2 phages that typically grow on enteric bacteria such as Escherichia coli and Shigella, with a prevalence of highly lytic T7-like phages (Table 1). Some of “plague phages” can be considered host range mutants of coliphages. For example, we isolated a Y. pestis-specific T4-like phage φJA1 from Maryland sewage [16,17].
Hostbacterium
Bacteriophage Family Group Sequence No. Reference(s)
Y. pestis Pokrovskaya(YepE2, YpP-G)
Podoviridae T7 NC_011038;JQ9657021
[14-18]
Y. pestis φA1122 Podoviridae T7 NC_004777 [16,17,19-23]Y. pestis Y Podoviridae T7 JQ9657001 [16-18,24,25]Y. pestis2 R Podoviridae T7 JQ9657011 [16-18,24,26-28]Y. pestis2 d’Herelle-m
(YpsP-G)Podoviridae T7 JQ9657031 [17,18,27,29,30]
Y. pestis Yep-phi Podoviridae T7 HQ333270 [31]Y. pestis2 PST Myoviridae T4 UA1 [16,17,24,32,33]Y. pestis φJA1 Myoviridae T4 UA1 [16,17]Y. pestis3 PY100 Siphoviridae T1 AM076770 [34]Y. pestis L-413C Myoviridae P2 NC_004745 [15-17,22,23,35,36]B. anthracis4 β (Wβ; beta) Siphoviridae γ DQ289555 [37-40]B. anthracis4 γ (Wγ; gamma) Siphoviridae γ DQ221100
DQ222851DQ222855DQ289556NC_007458
[38,39,41-47]
B. anthracis Fah Siphoviridae γ NC_007814 [46-48]B. anthracis AP50 Tectiviridae ? NC_011523 [49-53]B. anthracis5 8a Myoviridae ? UA [54]B. anthracis Nk Myoviridae ? UA [55]B. anthracis DB Myoviridae ? UA [55]B. anthracis MH Podoviridae ? UA [55]B. anthracis JRB7 Inoviridae ? UA [56,57]B. anthracis K ? ? UA [43,47,58,59]B. anthracis VA-9 ? ? UA [38,47]B. abortus6 Tb (Tbilisi) Podoviridae Tb NC_019446 [60-67]B. abortus7 Np (Nepean) Podoviridae Tb UA [65]B. abortus8 Fz (Firenze) Podoviridae Tb UA9 [63,65,68-70]B. abortus10 Pr (Perote) Podoviridae Tb NC_019447 [67]B. suis11 Wb (Weybridge) Podoviridae Tb UA9 [63-65,69-71]B. suis11 S708 Podoviridae Tb UA9 [69,70,72,73]B. suis12 1330 Podoviridae? Tb? UA [74-76]B. melitensis13 Bk (Berkeley) Podoviridae Tb UA9 [63,64,70,77]B. melitensis14 311 Podoviridae? Tb? UA [75,76,78]B. abortus15 R/O Podoviridae Tb UA [64,70,79-81]B. abortus16 R/C Podoviridae Tb UA9 [65,70,79-81]B. melitensis17 Iz (Izatnagar) Podoviridae Tb UA [65,70,82]
Notes: UA, unavailable. 1We (A.A. Filippov, K.V. Sergueev, and M.P. Nikolich, unpublished data) recently sequenced the genomes of PST and φJA1 (for the first time), as well as of the Pokrovskaya, Y, R, and d’Herelle-m phages. The last four genomes were virtually identical to the sequences JQ965702, JQ965700, JQ965701, and JQ965703, respectively, published by another group [18]. 2Phages R, d’Herelle-m and PST have slightly higher efficiencies of plating (EOP) on Yersinia pseudotuberculosis than on Y. pestis [17]. 3Phage PY100 lyses cultures of Y. pestis and several other species of Yersinia [34]. 4Both β and γ phages are lytic derivatives of a temperate phage W isolated from Bacillus cereus [37,41]. 58a is a spore-binding phage [54]. 6Tb lyses B. abortus at both RTD (routine test dilution, the highest phage dilution producing semiconfluent lysis) and 104 RTD, as well as lyses B. suis and B. neotomae only at 104 RTD [70]. 7Np is highly specific for B. abortus [65]. 8Fz lyses B. abortus and B. neotomae (at RTD and 104 RTD) and B. suis (only at 104 RTD) [70]. 9We (J. Farlow, A.A. Filippov, K.V. Sergueev, and M.P. Nikolich, manuscript in preparation) recently sequenced the genomes of brucellaphages Fz, Wb, S708, Bk, and R/C and observed a high level of identity with previously sequenced phages Tb and Pr [67]. 10Pr produces clear plaques on B. abortus and B. suis and turbid plaques on B. melitensis [67]. 11Wb and S708 lyse B. suis, B. abortus and B. neotomae (at RTD and 104 RTD); Wb can lyse some B. melitensis strains (also both at RTD and 104 RTD) [70]. 12Phage 1330 has a host range similar to that of Wb and S708 [74,76]. 13Bk lyses B. melitensis, B. abortus, B. suis, and B. neotomae at both titers [70,77]. 14Phage 311 has a host range similar to that of Bk [75,76]. 15R/O lyses B. ovis and rough strains of B. abortus at RTD and 104 RTD [70,80,81]. 16R/C lyses B. ovis, B. canis and rough strains of B. abortus at both concentrations [70,80,81]. 17Iz lyses smooth B. abortus, as well as both smooth (better) and rough (less efficiently) strains of B. neotomae, B. suis and B. melitensis [70,82].
Table 1: Bacteriophages active against biothreat bacteria.

Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 3 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
A standardized gamma phage lysis test [45] was approved by the FDA for anthrax diagnostics in 2005 [112]. This phage lysed 96-100% of B. anthracis strains tested including capsulated cultures and 2-8% of strains that belong to relative species: B. cereus, Bacillus megaterium, Bacillus mycoides, and Bacillus subtilis [45,47,113]. Despite its limited specificity, gamma is globally used for the identification of B. anthracis [43,45,47,113-116]. Phages Fah [47,113] and AP50 [117] are strictly B. anthracis-specific but have more narrow lytic spectra: they lyse only 73-89.5% and 33% of strains, respectively. Phages K and VA-9 lyse allB. anthracis strains but have slightly lower specificity than gamma; theK and VA-9 phages are widely used for anthrax diagnosis in the FSUcountries in addition to gamma and Fah [43,47].
The sources of isolation of anthrax phages were lysogenic cultures of B. anthracis [92-94] and Bacillus cereus [37,41], soil [49,54,55,95], sewage [96,97], and the intestinal tract of an earthworm [98]. These phages are very variable and belong to all the five above-mentioned families, however, only genomes of three B. anthracis-specific phages have been sequenced to date (not including prophages; see table 1). Interestingly, phage gamma contains a functional fosfomycin resistance gene, which is evidence of lysogenic conversion and could be the cause of the spread of fosfomycin resistance among B. anthracis strains that is, in turn, important for survival in soil in the presence of Streptomyces producing this antibiotic [39]. Moreover, the infection with temperate phages and lysogenization of B. anthracis can cause multiple phenotypic alterations: blockade or promotion of sporulation, induction of exopolysaccharide production and biofilm formation associated with the ability for long-term colonization of soil and the earthworm gut in a non-sporulating form [99].
The majority of Brucella typing phages were isolated from the bacterial cultures including B. abortus [65,68,72,100], B. suis [71-74], and B. melitensis [72,77,78,101] or host range mutants were derived from known phages following propagation on the bacterial cultures of interest, sometimes with the use of chemical mutagenesis [69,79,80,102]. Phages were isolated from the blood of 16 brucellosis patients out of 23 tested and in one case of severe neurobrucellosis, the phage was isolated both from blood and cerebrospinal fluid [103]. Several brucellaphages were isolated from aborted fetuses of farm animals [104], milk, urine, and feces of bovines [105] and farm sewage [60]. In contrast to the phages of Y. pestis and B. anthracis, brucellaphages have very similar or nearly identical morphology and a high degree of DNA homology despite their differences in the sources of isolation and host ranges; they all seem to belong to the same group of Tb-like phages (Table 1). A very interesting feature was discovered in phages Tb and Pr: the presence of putative DnaA-like protein and oriC-like origin of replication in their genomes suggesting that these phages can replicate by the same mechanism as bacterial chromosomes [67]. We sequenced five more brucellaphages (Fz, Wb, S708, Bk, and R/C) and found the same conserved sequences (J. Farlow, A.A. Filippov, K.V. Sergueev, and M.P. Nikolich, manuscript in preparation) that makes this group unique in the phage world.
Bacteriophage-based Diagnostics and Phage TypingRoutine bacteriophage lysis tests have been used for identification
of biothreat bacteria, discrimination of relative species and differentiation of typical and atypical strains for more than 80 years [96,100,106]. In particular, the phage lysis assay is an essential part of Y. pestis identification and bacteriological diagnosis of plague [107,108]. The specificity of four routinely used plague diagnostic phages was demonstrated on thousands of strains: Pokrovskaya and L-413C [15,107], φA1122 [20,108], and Yep-phi [31]. These phages arecapable of lysing practically all Y. pestis strains and allow differentiationfrom Y. pseudotuberculosis. Phage φA1122 is recommended for plaguediagnosis in the USA by the CDC, whereas Pokrovskaya and L-413C arewidely used in the FSU countries, and Yep-phi is applied in China. Weperformed efficiency of plating (EOP) tests for 11 phages on Y. pestis, Y.pseudotuberculosis and E. coli and showed that the Y and φJA1 phagesalso have high diagnostic value [17]. Phages can be used not only foridentification but also for typing of Y. pestis. For example, LPS-specificphages FP1 and FP3 were successfully used for differentiation of “mainsubspecies”, Y. pestis sbsp. pestis, from sbsp. microti and discriminationof several biovars within the latter subspecies [109].
A standard test of phage lysis requires a pure Y. pestis culture that extends this assay time up to two or three days [107,108]. We recently developed a rapid (4 h) and simple indirect phage-based method of detection and identification of Y. pestis [22]. The approach is based on the detection of φA1122 or L-413C amplification in the presence of Y. pestis by quantitative real-time PCR (qPCR) targeting the phage DNA but omitting the step of DNA isolation. The method is more specific than PCR tests targeting bacterial DNA, because in this case qPCR registers the titer increase of specific phage amplifying if live Y. pestis cells are available. The detection limit was as low as one bacterial cell per 1 µl sample (103 CFU/ml) and can be even lowered by sample concentration. The clinical relevance of this method was demonstrated on artificially spiked samples of human blood [22], sheep serum and mouse spleen suspensions with as high serum assay sensitivity as 104 CFU/ml with sample preparation and DNA isolation steps omitted [110]. We also proved that this method works well with the use of JBAIDS platform (Idaho Technology Inc.), a portable field-deployable thermocycler, with the same sensitivity as when using a full-sized qPCR instrument (Light Cycler 2.0, Roche) [110]. A genetically engineered phage φA1122 expressing luciferase reporter genes was recently exploited for fluorescent labeling and indirect detection of Y. pestis [21]. The method was found to be very rapid (1 h) and allowed detection of 820 or more Y. pestis cells, while the fluorescent signals were also observed with twoY. pseudotuberculosis strains and one Yersinia enterocolitica strain. Inthe latest publication of the same group, a second generation reporterphage was described with a higher level of the bioluminescent signalresponse providing a 10-fold increase in the assay sensitivity and apotential to discriminate drug-susceptible and drug-resistant strains ofY. pestis due to the correlation of bioluminescent signals with antibioticconcentration [111].
The routine phage lysis test is a long procedure that requires isolation of a pure B. anthracis culture (sometimes including spore germination) and its overnight exposure to the phage that overall takes two to four days [114]. Several novel approaches were used to improve phage-based diagnostics. Indirect detection of B. anthracis Sterne, using qPCR targeting the gamma phage DNA took only 5 h. Its sensitivity for pure culture was 2×104 bacterial cells per milliliter, but the specificity of phage propagation and subsequent bacterial detection was not tested on other B. anthracis strains and representatives of other Bacillus species [118].Using a luxAB-tagged reporter phage Wβ, the detection of B. anthraciswas observed within 60 minutes and at low concentration, 103 CFU/ml[40,119]. The lysin of the gamma phage, PlyG, was exploited for rapid(1 h) and sensitive (100 bacteria) detection of B. anthracis based on therelease of ATP from the lysed bacterial cells, which resulted in lightemission in the presence of luciferase and luciferin [44]. C-terminalbinding domain of PlyG was fused to glutathione S-transferase andused with horseradish peroxidase to bind to and detect ca. 103 CFU ofboth non-encapsulated and encapsulated B. anthracis within 3 h [120].

Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 4 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
A short synthetic polypeptide representing the putative PlyG binding motif was coupled with Qdot-nanocrystals and used for the detection of single B. anthracis cells under fluorescent microscope [121].
Environmental Applications
Bacteriophages were used for specific disinfection of medical catheters [132,133], food products [134,135], and even corals in aquarium experiments [136]. In the cases of bioweapon or bioterrorist
attacks, there is a problem of the “soft” disinfection of military, industrial and medical equipment, especially expensive sensitive equipment (like optical devices) that cannot withstand chemically aggressive disinfectants. The use of antibiotic solutions is not going to be efficient because there is a high probability of the application of natural or engineered multidrug-resistant strains. Recently, a phage cocktail including Pokrovskaya, Y, R, d’Herelle-m, and PST (see table 1) was employed for decontamination of hard surfaces (glass, gypsum board and stainless steel) spiked with Y. pestis, and the complete elimination of the bacteria was observed after a 5-min exposure to the cocktail at concentrations of 109, 108 and 107 PFU/ml [18]. A cocktail of unspecified phages isolated from topsoil retained viability at temperatures from –20°C to 37°C and when applied to B. anthracis spores, prevented their germination [137]. This approach could be expanded for the formulation of a phage cocktail active against multiple biothreat agents.
Phage TherapyGiven the emergence of drug-resistant strains of biothreat agents,
the development of alternative antibacterials has become an important goal. This problem is particularly acute for plague, because at least three multidrug-resistant strains of Y. pestis have been isolated from patients and a wild rodent [6,7]. Easy transmissibility of the resistance genes from these isolates was demonstrated, and one unique isolate was shown to be resistant to all the chemotherapeutic agents recommended for plague treatment and prophylaxis [6]. The widespread dissemination of extensively drug-resistant bacteria has driven recent progress in bacteriophage therapy against experimental, veterinary and human bacterial infections [138,139]. The first therapeutic trial of bubonic plague with bacteriophage was conducted in 1925: four patients received injections of phage suspension into buboes and recovered in a few days [140]. Several more attempts at phage therapy against plague in the 1920’s-1930’s failed or provided limited therapy efficiency which could be explained by a lack of proper laboratory practices with the phages [141]. After the successful treatment of plague with a sulfonamide was first implemented in 1938 [142], studies on bacteriophages as potential therapies against plague were discontinued until recently.
We have addressed the challenges of plague phage therapy from several angles [16,17,22,23,143]. We tested 11 phages lytic for Y. pestis and demonstrated the high lytic activity of T7-like phages and L-413C (see table 1) towards the plague bacterium [17,22], the high specificity of Pokrovskaya, L-413C, φJA1, φA1122, and Y [17,22], and the high stability of purified phage suspensions [23] prepared not only in a phage storage buffer (SM) but also in phosphate-buffered saline supplemented with gelatin suitable for parenteral injection [144]. Phages Pokrovskaya, L-413C, φJA1, φA1122, Y, and d’Herelle-m were shown to have low EOP on E. coli at 37°C [17] and thus, in contrast to antibiotics, they should not affect the normal microflora in animals and humans. We also addressed a potential problem of phage-resistant mutants by: i) demonstrating an extremely low frequency of mutation to φA1122 resistance, ii) identifying seven bacterial cell surface receptors for nine different phages, and iii) finding that most of phage-resistant mutants are attenuated for virulence in mice [16]. These data suggested that: i) potential phage resistance in the case of φA1122 is not a significant problem; ii) Y. pestis mutants resistant to one specific phage should remain susceptible to the other phages, thus the use of these phages in a therapeutic cocktail is likely to prevent cross-resistance [145]; and iii) the majority of phage-resistant mutants are attenuated and will be removed by the immune system and thus should not pose a problem for the efficacy of therapy. Plague diagnostic phages were shown to
There are two important fields in environmental applications of bacteriophages: detection of biothreat bacteria in environmental samples and phage-based decontamination. In contrast to PCR assays targeting bacterial DNA, phage-based techniques that rely on phage propagation detect only live bacterial cells. This is a definite advantage when testing the activity of natural foci of disease and for forensic purposes, such as during the investigation of bioterrorist attacks. For example, natural plague foci cover 6 to 7% of the dry surface of the Earth and require permanent epizootiological monitoring [124,125]. Employing the methods described in the previous section with the use of wild-type [22,110] and engineered [21,111] Y. pestis-specific phages is promising for determining the status of natural plague foci (e.g., when testing soil specimens from rodent burrows) and for the bacterial detection in environmental samples in case of intentional release of Y. pestis. The development of a field-deployable technology of indirect detection of Y. pestis [110] should facilitate the work in plague foci. Similarly, indirect detection methods with the use of intact [118] and genetically modified [40,119] phages or phage lysins [44,120,121] should help to identify B. anthracis in soil and other environmental samples in cases of natural anthrax or bioterrorist attacks. Since B. anthracis persists within soil both as vegetative cells and as spores [99,126,127], and spores can remain infective in soil as long as 70 years [128], the methods of spore detection are particularly important for environmental studies. A filamentous phage JRB7 was shown to specifically bind to B. anthracis spores [129] and was used in phage-coated magnetoelastic micro-biosensors for real-time detection of down to 103 spores per milliliter of water suspension [56,57]. Another spore-binding phage, 8a, was recently isolated and studied in terms of mechanisms of DNA injection [54]. Work is in progress on genome sequencing of the 8a phage and its genetic modification for developing an efficient method of B. anthracis spore detection [130]. Recent identification in the gamma phage lysin PlyG a domain for recognizing B. anthracis spores [131] can also help to improve phage-based spore detection. A successful method to identify live Brucella cells using phage-based qPCR (K.V. Sergueev, A.A. Filippov and M.P. Nikolich, manuscript in preparation) could be used for very sensitive and specific testing of agricultural and environmental samples in brucellosis foci.
There is a set of typing phages routinely and globally used for the identification and speciation of Brucella including Tb, Fz, Wb, S708, Bk, R/C, and Iz (Table 1). The character of lysis of Brucella cultures with the use of these phages at RTD and 104 RTD (see the notes to table 1) allows definitive identification of smooth and rough strains of B. abortus, B. suis, B. melitensis, B. neotomae, B. canis, and B. ovis [63,70,81,122] and even provides the differentiation of some biovars within B. suis[70,123]. Phage typing of Brucella is a multi-day procedure including isolation of a pure bacterial culture of slow-growing Brucella and the phage lysis tests. We recently developed a qPCR assay targeting S708 or Bk phage DNA for indirect identification of B. abortus (K.V. Sergueev, A.A. Filippov and M.P. Nikolich, manuscript in preparation). The assay allowed definitive identification of single cells of B. abortus during 72 hours in pure culture, blood and tissue samples.

Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 5 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
References
1. Atlas RM (2002) Bioterriorism: from threat to reality. Annu Rev Microbiol 56: 167-185.
2. http://www.bt.cdc.gov/agent/agentlist-category.asp
3. Butler T (2009) Plague into the 21st century. Clin Infect Dis 49: 736-742.
4. Sweeney DA, Hicks CW, Cui X, Li Y, Eichacker PQ (2011) Anthrax infection. Am J Respir Crit Care Med 184: 1333-1341.
5. Dean AS, Crump L, Greter H, Schelling E, Zinsstag J (2012) Global burden of human brucellosis: a systematic review of disease frequency. PLoS Negl Trop Dis 6: e1865.
6. Galimand M, Courvalin P (2012) Plague treatment and resistance to antimicrobial agents. In: Carniel E and Hinnebusch BJ, eds. Yersinia: systems biology and control. Caister Academic Press, UK.
7. Kiefer D, Dalantai G, Damdindorj T, Riehm JM, Tomaso H, et al. (2012) Phenotypical characterization of Mongolian Yersinia pestis strains. Vector Borne Zoonotic Dis 12: 183-188.
8. Kinsara A, Al-Mowallad A, Osoba AO (1999) Increasing resistance of Brucellae to co-trimoxazole. Antimicrob Agents Chemother 43: 1531.
9. Memish Z, Mah MW, Al Mahmoud S, Al Shaalan M, Khan MY (2000) Brucella bacteraemia: clinical and laboratory observations in 160 patients. J Infect 40: 59-63.
10. Athamna A, Athamna M, Abu-Rashed N, Medlej B, Bast DJ, et al. (2004) Selection of Bacillus anthracis isolates resistant to antibiotics. J Antimicrob Chemother 54: 424-428.
11. Gorelov VN, Gubina EA, Grekova NA, Skavronskaia AG (1991) The possibility of creating a vaccinal strain of Brucella abortus 19-BA with multiple antibiotic resistance. Zh Mikrobiol Epidemiol Immunobiol 2-4.
12. Pomerantsev AP, Staritsyn NA (1996) Behavior of heterologous recombinant plasmid pCET in cells of Bacillus anthracis. Genetika 32: 500-509.
13. Block SM (1999) Living nightmares: biological threats enabled by molecular biology. In: Drell SD, Sofaer AD, Wilson GD, eds. The new terror: facing the threat of biological and chemical weapons. Hoover Institution Press, Stanford 39-75.
14. Pokrovskaya MP (1929) A plague bacteriophage in dead susliks. Gigiena Epidemiol 12: 31-34.
15. Imamaliev OG, Serebryakova VG, Anisimova TI, Plotnikov OP, Sergeeva G. M, et al. (1986) Comparative estimation of activity and specificity of diagnostic plague bacteriophages, L-413C, and Pokrovskaya, In: Bektemirov TA, Zhouravleva YZ, Litvinova MY et al., eds. Standards, strains, and methods of control of bacterial and viral preparations. Mechnikov Institute Press, Moscow 102-106.
17. Filippov AA, Sergueev KV, He Y, Nikolich MP (2012) Bacteriophages capable of lysing Yersinia pestis and Yersinia pseudotuberculosis: efficiency of plating tests and identification of receptors in Escherichia coli K-12. Adv Exp Med Biol 954: 123-134.
18. Rashid MH, Revazishvili T, Dean T, Butani A, Verratti K, et al. (2012) A Yersinia pestis-specific, lytic phage preparation significantly reduces viable Y. pestis on various hard surfaces experimentally contaminated with the bacterium. Bacteriophage 2: 168-177.
19. Advier M (1933) Etude d’un bactériophage antipesteux. Bull Soc Pathol Exotiques 26: 94-99.
20. Garcia E, Elliott JM, Ramanculov E, Chain PSG, Chu MC, et al. (2003) The genome sequence of Yersinia pestis bacteriophage φA1122 reveals an intimate history with the coliphage T3 and T7 genomes. J Bacteriol 185: 5248-5262.
21. Schofield DA, Molineux IJ, Westwater C (2009) Diagnostic bioluminescent phage for detection of Yersinia pestis. J Clin Microbiol 47: 3887-3894.
22. Sergueev KV, He Y, Borschel RH, Nikolich MP, Filippov AA (2010) Rapid and sensitive detection of Yersinia pestis using amplification of plague diagnostic bacteriophages monitored by real-time PCR. PLoS One 5: e11337.
23. Filippov AA, Sergueev KV, He Y, Huang XZ, Gnade BT, et al. (2012)
B.
ConclusionBased on the current body of literature and state of the field, one
can conclude that bacteriophages lytic for Y. pestis, B. anthracis and Brucella species are routinely used for the detection, identification and typing of the host bacteria. New highly efficient indirect approaches with the use of genetically engineered or wild type phages have advanced diagnosis of the biothreat infections but require acceptance to be moved into practical use. Phage cocktails can be successfully used for the specific decontamination of chemically sensitive surfaces and expensive equipment. Highly lytic phages and their lysins are promising antibacterials for the treatment of drug-resistant plague, anthrax and brucellosis.
Acknowledgements
This work was supported in part by the Defense Threat Reduction Agency, Joint Science and Technology Office, Medical S&T Division. The findings and opinions expressed herein belong to the authors and do not necessarily reflect the official views of the WRAIR, the U.S. Army or the Department of Defense.
16. Filippov AA, Sergueev KV, He Y, Huang XZ, Gnade BT, et al. (2011) Bacteriophage-resistant mutants in Yersinia pestis: identification of phage receptors and attenuation for mice. PLoS One 6: e25486.
be safe in cytotoxicity and acute toxicity tests [23]. Intraperitoneal administration of φA1122 provided longer persistence and one log higher concentrations of the phage in mouse spleen, liver and blood vs. intramuscular injections [23]. Pharmacodynamic and therapeutic trials revealed the propagation of φA1122 in mice infected with Y. pestis, a marked reduction of the bacteria in blood and organs, or total clearance of them, and a therapeutic effect of the phage. The most effective result was observed after a challenge with 103 LD50 of Y. pestis and subsequent (1 h later) single intraperitoneal injection of the phage at each of two doses (5×108 and 5×109 PFU): the survival of 20-40% mice and an 84% extension of mean time to death in nonsurvivors [23]. We consider this result very promising, because this is the first attempt of phage therapy against a fulminant plague infection in mice (LD50=2 bacteria). A single injection of the φA1122 phage provided an efficacy of therapy that can be compared with the results of multiple daily injections of a high dose of antibiotic over five days [146,147]. As for possible ways to improve the efficacy of plague therapy, we [143] suggest testing intravenous and intranasal routes of phage administration, as well as injections into or around the buboes [140], the selection of longer (10-14 days) circulating phage mutants [148], the application of different phages and phage cocktails, and the use of alternative animal models (rats and guinea pigs) providing higher lethal doses of Y. pestis and longer infectious processes.
Currently, the concern about drug resistance for B. anthracis and Brucella is not as eminent as in the case of plague. However, the potential introduction of multidrug resistance to current therapeutic options is a serious threat with either pathogen species which could provide dramatic problems in outbreak response. If this problem becomes more serious, there are lytic phages active against both B. anthracis and Brucella as potential alternative therapies. The use of phage lysins is a very promising area because this approach helps to circumvent the problem of phage resistance. The gamma phage PlyG lysin was successfully used in preliminary trials for intravenous therapy of experimental B. cereus and attenuated B. anthracis infection: 70-90% of treated vs. 0-10% of untreated mice survived [44,149]. Interestingly, brucellaphage were isolated from blood of the vast majority of brucellosis patients and a phage titer rise correlated with the improvement of patients’ condition [103]. Administration of phage to guinea pigs and rabbits with experimental brucellosis stimulated the immunity (resulted in higher agglutination titers) and increased the number of animals cleared from Brucella [150].

Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 6 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
Bacteriophage therapy of experimental bubonic plague in mice. Adv Exp Med Biol 954: 337-348.
24. Knapp W (1963) On the varying behavior of Pasteurella phages. Zentralbl Bakteriol 190: 39-46.
25. Hertman I (1964) Bacteriophage common to Pasteurella pestis and Escherichia coli. J Bacteriol 88: 1002-1005.
26. Kotlyarova RI (1956) Pseudotuberculosis bacteriophage and its characteristics. In: Proceedings of Research Anti-Plague Institute of Caucasus and Transcaucasia. Stavropol 234-241.
27. Shashaev MA (1966) Comparative characterization of plague and pseudotuberculosis phages. In: Production of bacterial preparations for prophylaxis and diagnostics of particularly dangerous infections. Saratov 350-356.
28. Arutiunov IuI (1970) The biological characteristics of plague and pseudotuberculosis bacteriophages. Zh Mikrobiol Epidemiol Immunobiol 47: 106-111.
29. Sakal NN, Grischenko RI (1977) On the specificity of pseudotuberculosis phage. Probl Especially Dangerous Infect 53: 48-51.
30. Leshkovich NL (1981) Inhibitory effect of normal serum on phage adsorption. In: Prophylaxis of particularly dangerous infections. Alma-Ata 66-69.
31. Zhao X, Wu W, Qi Z, Cui Y, Yan Y, et al. (2010) The complete genome sequence and proteomics of Yersinia pestis phage Yep-phi. J Gen Virol 92: 216-221.
32. Ackermann HW, Poty F (1969) Relationship between coliphages T2 and 3, and phage PST of Pasteurella pseudotuberculosis. Rev Can Biol 28: 201-204.
33. Gurleva GG, Arutyunov YI, Khaliapina EE (1981) Serologic relationship and specificity of action of pseudotuberculosis and coli-dysentery phages. Zh Mikrobiol Epidemiol Immunobiol 5: 65-68.
34. Schwudke D, Ergin A, Michael K, Volkmar S, Appel B, et al. (2008) Broad-host-range Yersinia phage PY100: genome sequence, proteome analysis of virions, and DNA packaging strategy. J Bacteriol 190: 332-342.
35. Larina VS, Anisimov PI, Adamov AK (1970) A novel strain of plague bacteriophage for identification of Pasteurella pestis. Probl Especially Dangerous Infect 11: 132-136.
36. Garcia E, Chain P, Elliott JM, Bobrov AG, Motin VL, et al. (2008) Molecular characterization of L-413C, a P2-related plague diagnostic bacteriophage. Virology 372: 85-96.
37. McCloy EW (1951) Unusual behavior of a lysogenic Bacillus strain. J Gen Microbiol 5: xiv–xvi.
38. Levina EN, Arkhipova VR (1967) A comparative study of anthrax bacteriophages. Zh Mikrobiol Epidemiol Immunobiol 44: 24-28.
39. Schuch R, Fischetti VA (2006) Detailed genomic analysis of the Wβ and γ phages infecting Bacillus anthracis: implications for evolution of environmental fitness and antibiotic resistance. J Bacteriol 188: 3037-3051.
40. Schofield DA, Westwater C (2009) Phage-mediated bioluminescent detection of Bacillus anthracis. J Appl Microbiol 107: 1468-1478.
41. Brown ER, Cherry WB (1955) Specific identification of Bacillus anthracis by means of a variant bacteriophage. J Infect Dis 96: 34-39.
42. Watanabe T, Morimoto A, Shiomi T (1975) The fine structure and the protein composition of gamma phage of Bacillus anthracis. Can J Microbiol 21: 1889-1892.
43. Ipatenko NG, Manichev AA, Sedov VA, Guschin VN (1989) Testing anthrax bacteriophages K (VIEV) and gamma (MVA). Veterinariya 10: 58-59.
44. Schuch R, Nelson D, Fischetti VA (2002) A bacteriolytic agent that detects and kills Bacillus anthracis. Nature 418: 884-889.
45. Abshire TG, Brown JE, Ezzell JW (2005) Production and validation of the use of gamma phage for identification of Bacillus anthracis. J Clin Microbiol 43: 4780-4788.
46. Fouts DE, Rasko DA, Cer RZ, Jiang L, Fedorova NB, et al. (2006) Sequencing Bacillus anthracis typing phages gamma and cherry reveals a common ancestry. J Bacteriol 188: 3402-3408.
47. Golovinskaya TM, Buravtseva NP, Tsygankova OI, Eremenko EI (2011) Comparative study of lytic activity and specificity of anthrax bacteriophages
gamma (A-26), K (VIEV), VA-9 and Fah (VNIIV&M). Probl Especially Dangerous Infect 109: 28-30.
48. Minakhin L, Semenova E, Liu J, Vasilov A, Severinova E, et al. (2005) Genome sequence and gene expression of Bacillus anthracis bacteriophage Fah. J Mol Biol 354: 1-15.
49. Nagy E (1974) A highly specific phage attacking Bacillus anthracis strain Sterne. Acta Microbiol Acad Sci Hung 21: 257-263.
50. Ackermann H-W, Roy R, Martin M, Murthy MR, Smirnoff WA (1978) Partial characterization of a cubic Bacillus phage. Can J Microbiol 24: 986-993.
51. Nagy E, Herczegh O, Ivanova N (1982) Lipid-containing anthrax phage AP50: structural proteins and life cycle. J Gen Virol 62: 323-329.
52. Sozhamannan S, McKinstry M, Lentz SM, Jalasvuori M, McAfee F, et al. (2008) Molecular characterization of a variant of Bacillus anthracis-specific phage AP50 with improved bacteriolytic activity. Appl Environ Microbiol 74: 6792-6796.
53. Bishop-Lilly KA, Plaut RD, Chen PE, Akmal A, Willner KM, et al. (2012) Whole genome sequencing of phage resistant Bacillus anthracis mutants reveals an essential role for cell surface anchoring protein CsaB in phage AP50c adsorption. Virol J 9: 246.
54. Fu X, Walter MH, Paredes A, Morais MC, Liu J (2011) The mechanism of DNA ejection in the Bacillus anthracis spore-binding phage 8a revealed by cryo-electron tomography. Virology 421: 141-148.
55. Walter MH, Baker DD (2003) Three Bacillus anthracis bacteriophages from topsoil. Curr Microbiol 47: 55-58.
56. Wan J, Johnson ML, Guntupalli R, Petrenko VA, Chin BA (2007) Detection of Bacillus anthracis spores in liquid using phage-based magnetoelastic micro-resonators. Sens Actuators B Chem 127: 559-566.
57. Shen W, Lakshmanan RJ, Mathison LC, Petrenko VA, Chin BA (2009) Phage coated magnetoelastic micro-biosensors for real-time detection of Bacillus anthracis spores. Sens Actuators B Chem 137: 501-506.
58. Boikov YI (1964) Impact of anthrax bacteriophage on different strains of Bacillus anthracis. Probl Vet Sanitation 24: 120-124.
59. Rusaleev VS (1990) Phage susceptibility of aerobic spore-forming bacteria. Veterinariya 8: 29-31.
60. Popkhadze MZ, Abashidze TG (1957) Characterization of a Brucella phage isolated at Tbilisi Research Institute for Vaccines and Sera. In: Abstracts of Inter-Institute Science Conference on Bacteriophagy, Tbilisi 40.
61. Stinebring WR, Braun W (1959) Brucellaphage. J Bacteriol 78: 736-737.
62. McDuff CR, Jones LM, Wilson JB (1962) Characteristics of brucellaphage. J Bacteriol 83: 324-329.
63. Taran IF, Zanina VM, Liamkin GI, Tsybin BP, Tikhenko NI (1983) Comparative evaluation of the spectrum of lytic effects of bacteriophages Tb, Wb, Fi, Bk2 and R on various Brucella species. Zh Mikrobiol Epidemiol Immunobiol 2: 48-52.
64. Segondy M, Allardet-Servent A, Caravano R, Ramuz M (1988) Common physical map of four Brucella bacteriophage genomes. FEMS Microbiol Lett 56: 177-181.
65. Rigby CE, Cerqueira-Campos ML, Kelly HA, Surujballi OP (1989) Properties and partial genetic characterization of Nepean phage and other lytic phages of Brucella species. Can J Vet Res 53: 319-325.
66. Zhu CZ, Xiong HY, Han J, Cui BY, Piao DR, et al. (2009) Molecular characterization of Tb, a new approach for an ancient brucellaphage. Int J Mol Sci 10: 2999-3011.
67. Flores V, López-Merino A, Mendoza-Hernandez G, Guarneros G (2012) Comparative genomic analysis of two brucellaphages of distant origins. Genomics 99: 233-240.
68. Corbel MJ, Thomas EL (1976) Properties of some new Brucella phage isolates; evidence for lysogeny within the genus. Devs Biol Stand 31: 38-45.
69. Corbel MJ, Thomas EL (1976) Description of a new phage lytic for several Brucella species. J Biol Stand 4: 195-201.
70. Corbel MJ (1987) Brucella phages: advances in the development of a reliable phage typing system for smooth and non-smooth Brucella isolates. Ann Inst Pasteur Microbiol 138: 70-75.

Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 7 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
71. Morris JA, Corbel MJ (1973) Properties of a new phage lytic for Brucella suis. J Gen Virol 21: 539-544.
72. Moreira-Jacob M (1968) New group of virulent bacteriophages showing differential affinity for Brucella species. Nature 219: 752-753.
73. Morris JA, Corbel MJ, Phillip JIH (1973) Characterization of three phages lytic for Brucella species. J Gen Virol 20: 63-73.
74. Kiseleva VI (1968) Isolation of a virulent phage from Brucella suis strain 1330. In: Biochemistry, genetics and immunology (especially dangerous infections). Rostov University Press, Rostov 121-125.
75. Drozhevkina MS, Kiseleva VI (1968) New polyvalent phage for identification of Brucella. In: Diagnostics of especially dangerous infections. Rostov University Press, Rostov 150-154.
76. Drozhevkina MS (1968) Attempting to use bacteriophage for speciation of brucellae isolated from deer. In: Diagnostics of especially dangerous infections. Rostov University Press, Rostov 155-160.
77. Douglas JT, Elberg SS (1976) Isolation of Brucella melitensis phage of broad biotype and species specificity. Infect Immun 14: 306-308.
78. Drozhevkina MS, Uraleva VS, Chernenkova NA (1955) Application of brucellosis bacteriophage for identification of hemocultures isolated from patients. In: Proceedings of Rostov Anti-Plague Institute, Rostov 9: 133-140.
79. Corbel MJ (1979) Isolation and properties of a phage lytic for non-smooth Brucella organisms. J Biol Stand 7: 349-360.
80. Corbel MJ (1984) Properties of brucellaphages lytic for non-smooth Brucella strains. Dev Biol Stand 56: 55-62.
81. Corbel MJ, Thomas EL (1985) Use of phage for the identification of Brucella canis and Brucella ovis cultures. Res Vet Sci 38: 35-40.
82. Corbel MJ, Tolari F, Yadava VK (1988) Characterization of a new phage lytic for both smooth and non-smooth Brucella species. Res Vet Sci 44: 45-49.
83. Ackermann HW (2003) Bacteriophage observations and evolution. Res Microbiol 154: 245-251.
84. Kiljunen S, Datta N, Dentovskaya SV, Anisimov AP, Knirel YA, et al. (2011) Identification of the lipopolysaccharide core of Yersinia pestis and Yersinia pseudotuberculosis as the receptor for bacteriophage φA1122. J Bacteriol 193: 4963-4972.
85. Davison S, Couture-Tosi E, Candela T, Mock M, Fouet A (2005) Identification of the Bacillus anthracis γ phage receptor. J Bacteriol 187: 6742-6749.
86. Corbel MJ (1977) Isolation and partial characterization of a phage receptor from “Brucella neotomae” 5K33. Ann Sclavo 19: 131-142.
87. Shashaev MA (1965) Technique of plague phage isolation from rodents. In: Proceedings of the 4th Conference on natural focality and prophylaxis of plague. Alma-Ata, 298-299.
88. Plotnikov OP, Larina VS, Kondrashin YI, Vorob’yova, TV (1982) Spectrum of lytic activity of bacteriophages isolated from soil of rodent burrows. In: Anisimov PI, Karaseva ZN, Konnov NP, eds. Selection and genetics of causative agents of especially dangerous infections. Institute “Microbe” Press, Saratov 56-61.
89. Leshkovich NL, Arutyunov YI, Tokarev YI, Kirdeev VK (1975) Morphology of plague temperate phages 1701, 1710, and their mutants. Zh Mikrobiol Epidemiol Immunobiol 10: 31-34.
90. Novoseltsev NN, Marchenkov VI (1990) Yersinia pestis phage of a new serovar. Zh Mikrobiol Epidemiol Immunobiol 11: 9-12.
91. Novosel’tsev NN, Marchenkov VI, Arutiunov IuI (1994) Phages of the IV serovar of Yersinia pestis. Zh Mikrobiol Epidemiol Immunobiol 6: 9-10.
92. Buck CA, Anacker RL, Newman FS, Eisenstark A (1963) Phage isolated from lysogenic Bacillus anthracis. J Bacteriol 85: 1423-1430.
93. Larina VS, Petrova LS (1964) Isolation of active bacteriophage from Bacillus anthracis found in soil. In: Microbiology and immunology of especially dangerous infections. Saratov 94-95.
94. Inal JM, Karunakaran KV (1996) phi 20, a temperate bacteriophage isolated from Bacillus anthracis exists as a plasmidial prophage. Curr Microbiol 32: 171-175.
95. Rusaleev VS (1991) Effect of anthrax bacteriophage 3-17 on Bacillus anthracis strains. Veterinariya 5: 26-28.
96. Cowles PB (1931) A bacteriophage for B. anthracis. J Bacteriol 21: 161-166.
97. Terentiev (1937) Bacteriophage of B. anthracis. In: Proceedings of All-Union Institute for Experimental Veterinary. 13: 65-69.
98. Schuch R, Pelzek AJ, Kan S, Fischetti VA (2010) Prevalence of Bacillus anthracis-like organisms and bacteriophages in the intestinal tract of the earthworm Eisenia fetida. Appl Environ Microbiol 76: 2286-2294.
99. Schuch R, Fischetti VA (2009) The secret life of the anthrax agent Bacillus anthracis: bacteriophage-mediated ecological adaptations. PLoS One 4: e6532.
100. Drozhevkina MS (1951) Brucella bacteriophage. The phenomenon of bacteriophagy in Brucella cultures and isolation of the first strains of bacteriophage. Zh Microbiol Epidemiol Immunobiol 11: 34-37.
101. Drozhevkina MS, Kiseleva VI, Kudryakova TA (1967) Virulent phages of Brucella melitensis. In: Genetics, biochemistry and immunochemistry of especially dangerous infections. Rostov University Press, Rostov 54-59.
102. Thomas EL, Corbel MJ (1977) Isolation of a phage lytic for several Brucella species following propagation of Tbilisi phage in the presence of mitomycin C. Arch Virol 54: 259-261.
103. Drozhevkina MS, Mishnaevskii MN, Uraleva VS (1957) Specific bacteriophage in the organism of brucellosis patients. In: Proceedings of Rostov Anti-Plague Institute 12: 403-423.
104. Drozhevkina MS, Tolstokorova VI (1957) Isolation of Brucella bacteriophage from aborted fetuses of farm animals. In: Proceedings of Rostov Anti-Plague Institute 12: 424-427.
105. Mamatsashvili EG (1955) Isolation of phage and determination of some its features. In: Proceedings of Inter-Institute Conference on Bacteriophagy 5: 327-332.
106. Lazarus AS, Gunnison JB (1947) The action of Pasteurella pestis bacteriophage on strains of Pasteurella, Salmonella, and Shigella. J Bacteriol 53: 705-714.
107. Naumov AV, Samoilova LV (1992) Laboratory diagnostics of plague. Institute “Microbe” Press, Saratov.
108. Chu MC (2000) Laboratory manual of plague diagnostic tests. Centers for Disease Control and Prevention, Atlanta, GA.
109. Balakhonov SV, Bel’kova SA, Gremyakova TA, Shaikhutdinova RZ (2001) On the possibility of intraspecific typing of Yersinia pestis with bacteriophages. In: Proceedings of Theoretical and Practical Conference on Important Aspects of Naturally Focal Diseases, Omsk, pp. 170-172.
110. Sergueev KV, Nikolich MP, Filippov AA (2012) Field and clinical applications of advanced bacteriophage-based detection of Yersinia pestis. Adv Exp Med Biol 954: 135-141.
111. Schofield DA, Molineux IJ, Westwater C (2012) Rapid identification and antibiotic susceptibility testing of Yersinia pestis using bioluminescent reporter phage. J Microbiol Methods 90: 80-82.
112. http://www.sciencedaily.com/releases/2005/08/050830070143.htm
113. Sayapina LV, Abdrashitova AS, Kasina IV, Malakhaeva AN, Rashchepkin LI, et al. (2011) Characterization of new liquid anthrax diagnostic bacteriophage gamma A-26. Biopreparaty 1.
114. Turnbull PCB (1999) Definitive identification of Bacillus anthracis–a review. J Appl Microbiol 87: 237-240.
115. Klee SR, Nattermann H, Becker S, Urban-Schriefer M, Franz T, et al. (2006) Evaluation of different methods to discriminate Bacillus anthracis from other bacteria of the Bacillus cereus group. J Appl Microbiol 100: 673-681.
116. Irenge LM, Gala JL (2012) Rapid detection methods for Bacillus anthracis in environmental samples: a review. Appl Microbiol Biotechnol 93: 1411-1422.
117. Nagy E, Ivanovics G (1977) Association of probable defective phage particles with lysis by bacteriophage AP50 in Bacillus anthracis. J Gen Microbiol 102: 215-219.
118. Reiman RW, Atchley DH, Voorhees KJ (2007) Indirect detection of Bacillus anthracis using real-time PCR to detect amplified gamma phage DNA. J Microbiol Methods 68: 651-653.
119. Schofield DA, Sharp NJ, Westwater C (2012) Phage-based platforms for the clinical detection of human bacterial pathogens. Bacteriophage 2: 105-283.

Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010
Page 8 of 8
ISSN:2157-2526 JBTBD, an open access journal J Bioterr Biodef Advances in Biosciences: Bioterrorism
120. Fujinami Y, Hirai Y, Sakai I, Yoshino M, Yasuda J (2007) Sensitive detection of Bacillus anthracis using a binding protein originating from γ-phage. Microbiol Immunol 51: 163-169.
121. Sainathrao S, Mohan KV, Atreya C (2009) Gamma-phage lysin PlyG sequence-based synthetic peptides coupled with Qdot-nanocrystals are useful for developing detection methods for Bacillus anthracis by using its surrogates, B. anthracis-Sterne and B. cereus-4342. BMC Biotechnol 9: 67.
122. Jones LM, Merz GS, Wilson JB (1968) Phage typing reactions on Brucella species. Appl Microbiol 16: 1179-1190.
123. Thomas EL, Corbel MJ (1982) Differentiation of Brucella suis biotype 4 from other biotypes by Firenze phage. Br Vet J 138: 11-17.
124. Anisimov AP, Lindler LE, Pier GB (2004) Intraspecific diversity of Yersinia pestis. Clin Microbiol Rev 17: 434-464.
125. Gage KL, Kosoy MY (2005) Natural history of plague: perspectives from more than a century of research. Annu Rev Entomol 50: 505-528.
126. Dey R, Hoffman PS, Glomski IJ (2012) Germination and amplification of anthrax spores by soil-dwelling amoebas. Appl Environ Microbiol 78: 8075-8081.
127. http://www.epi.ufl.edu/?q=node/1186
128. Guillemin J (2001) Anthrax: the investigation of a deadly outbreak. University of California Press, Berkeley, CA, USA.
129. Brigati J, Williams DD, Sorokulova IB, Nanduri V, Chen IH, et al. (2004) Diagnostic probes for Bacillus anthracis spores selected from a landscape phage library. Clin Chem 50: 1899-1906.
130. http://www.bioprepwatch.com/news/new-phage-could-fight-anthrax/297236/
131. Yang H, Wang DB, Dong Q, Zhang Z, Cui Z, et al. (2012) Existence of separate domains in lysin PlyG for recognizing Bacillus anthracis spores and vegetative cells. Antimicrob Agents Chemother 56: 5031-5039.
132. Curtin JJ, Donlan RM (2006) Using bacteriophages to reduce formation of catheter-associated biofilms by Staphylococcus epidermidis. Antimicrob Agents Chemother 50: 1268-1275.
133. Fu W, Forster T, Mayer O, Curtin JJ, Lehman SM, et al. (2010) Bacteriophage cocktail for the prevention of biofilm formation by Pseudomonas aeruginosa on catheters in an in vitro model system. Antimicrob Agents Chemother 54: 397-404.
134. Leverentz B, Conway WS, Camp MJ, Janisiewicz WJ, Abuladze T, et al. (2003) Biocontrol of Listeria monocytogenes on fresh-cut produce by treatment with lytic bacteriophages and a bacteriocin. Appl Environ Microbiol 69: 4519-4526.
135. Jassim SA, Abdulamir AS, Abu Bakar F (2012) Novel phage-based bio-processing of pathogenic Escherichia coli and its biofilms. World J Microbiol Biotechnol 28: 47-60.
136. Efrony R, Atad I, Rosenberg E (2009) Phage therapy of coral white plague disease: properties of phage BA3. Curr Microbiol 58: 139-145.
137. Walter MH (2003) Efficacy and durability of Bacillus anthracis bacteriophages used against spores. J Environ Health 66: 9-15.
138. Burrowes B, Harper DR, Anderson J, McConville M, Enright MC (2011) Bacteriophage therapy: potential uses in the control of antibiotic-resistant pathogens. Expert Rev Anti Infect Ther 9: 775-785.
139. Abedon ST, Kuhl SJ, Blasdel BG, Kutter EM (2011) Phage treatment of human infections. Bacteriophage 1: 66-85.
140. d’Hérelle F (1925) Essai de traitement de la peste bubonique par le bactériophage. La Presse Med 33: 1393-1394.
141. Anisimov AP, Amoako KK (2006) Treatment of plague: promising alternatives to antibiotics. J Med Microbiol 55: 1461-1475.
142. Pollitzer R (1954) Plague WHO, Geneva 409-482.
143. Filippov AA, Sergueev KV, Nikolich MP (2012) Can phage effectively treat multidrug-resistant plague? Bacteriophage 2: 186-189.
144. Cerveny KE, DePaola A, Duckworth DH, Gulig PA (2002) Phage therapy of local and systemic disease caused by Vibrio vulnificus in iron-dextran-treated mice. Infect Immun 70: 6251-6262.
145. Levin BR, Bull JJ (2004) Population and evolutionary dynamics of phage therapy. Nat Rev Microbiol 2: 166-173.
146. Byrne WR, Welkos SL, Pitt ML, Davis KJ, Brueckner RP, et al. (1998) Antibiotic treatment of experimental pneumonic plague in mice. Antimicrob Agents Chemother 42: 675-681.
147. Steward J, Lever MS, Russell P, Beedham RJ, Stagg AJ, et al. (2004) Efficacy of the latest fluoroquinolones against experimental Yersinia pestis. Int J Antimicrob Agents 24: 609-612.
148. Merril CR, Biswas B, Carlton R, Jensen NC, Creed GJ, et al. (1996) Long-circulating bacteriophage as antibacterial agents. Proc Natl Acad Sci USA 93: 3188-3192.
149. Fischetti VA (2006) Using phage Lytic Enzymes to Control Pathogenic Bacteria. BMC Oral Health 6: S16.
150. Drozhevkina MD (1963) The present position in Brucella phage research. Bull World Health Organ 29: 43-57.
Thisarticlewasoriginallypublishedinaspecialissue,Advances in Biosci-ences: Bioterrorism handledbyEditor(s).Dr.NicholasEarlBurgis,EasternWashingtonUniversity,USA.
Citation: Filippov AA, Sergueev KV, Nikolich MP (2013) Bacteriophages against Biothreat Bacteria: Diagnostic, Environmental and Therapeutic Applications. J Bioterr Biodef S3: 010. doi:10.4172/2157-2526.S3-010