author's personal copy - ecd group · author's personal copy ... including environmental...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Review Article
Care for Child Development: Basic Science
Rationale and Effects of InterventionsRebecca L. Holt, BA and Mohamad A. Mikati, MD
The past few years have witnessed increasing interest indevising programs to enhance early childhood develop-ment. We review current understandings of brain devel-opment, recent advances in this field, and theirimplications for clinical interventions. An expandingbody of basic science laboratory data demonstratesthat several interventions, including environmentalenrichment, level of parental interaction, erythropoietin,antidepressants, transcranial magnetic stimulation,transcranial direct current stimulation, hypothermia,nutritional supplements, and stem cells, can enhance ce-rebral plasticity. Emerging clinical data, using functionalmagnetic resonance imaging and clinical evaluations,also support the hypothesis that clinical interventionscan increase the developmental potential of children,rather than merely allowing the child to achieve an al-ready predetermined potential. Such interventions in-clude early developmental enrichment programs, whichhave improved cognitive function; high-energy andhigh-protein diets, which have increased brain growthin infants with perinatal brain damage; constraint-induced movement therapy, which has improved motorfunction in patients with stroke, cerebral palsy, and cere-bral hemispherectomy; and transcranial magnetic stim-ulation, which has improved motor function in strokepatients. � 2011 Elsevier Inc. All rights reserved.
Holt RL, Mikati MA. Care for child development: Basicscience rationale and effects of interventions. PediatrNeurol 2011;44:239-253.
Introduction
In 2007, over 200 million children under age 5 yearswere estimated not to be fulfilling their developmental
potential [1]. Child development is largely dependent onthe quality of early experiences. Early intervention canenhance the brain’s potential through cerebral plasticity.The hypothesis underlying intervention programs claimsthat early intervention does not merely help the brain toachieve a predetermined potential, but that it enhancesthe underlying brain’s potential altogether. A convincingbody of evidence from neurobiology supports this hypoth-esis. The challenge will be to identify those interventionsthat are effective, safe, and practical, based not only onfield experience but also on basic science data. Althoughsimilar in action, interventions that enhance developmentalpotential are distinct from factors that provide neuroprotec-tion during or after a specific insult. Fig 1 depicts thepotential relationship between neuroprotection and neuralrecovery and enhancement. We will present basic scienceevidence for both kinds of interventions. The implementa-tion of early interventions will require the support andcommitment of developmental psychologists, primarycare providers, parents, policymakers, and local and globalcommunities. Basic scientists can also contribute to thisprocess. Through a cooperative effort, we should be ableto increase the effectiveness of programs aimed at enhanc-ing childhood development and the awareness of thebenefits of early intervention.
Cerebral Plasticity and Its Role in CerebralDevelopment and Recovery
Cerebral plasticity refers to the brain’s ability to learn, re-member, forget, reorganize, and recover from injury. Theconceptual framework for plasticity was formulated in1949 by Hebb, who postulated that when one cell excitesanother repeatedly, a change occurs in one or both cellsthat contributes to their stability [2]. In other words,
From the Department of Pediatric Neurology, Children’s Health Center,Duke University Medical Center, Durham, North Carolina.
Communications should be addressed to:Dr. Mikati; Department of Pediatric Neurology; Children’s Health Center;Duke University Medical Center; 2301 Erwin Road; Box 3936;Durham, NC 27710.E-mail: [email protected] June 30, 2010; accepted November 16, 2010.
� 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.pediatrneurol.2010.11.009 � 0887-8994/$ - see front matter
Holt and Mikati: Care for Child Development 239

Author's personal copy
‘‘neurons that fire together, wire together.’’ Neurogenesisand synaptogenesis comprise the activity-dependent mech-anisms underlying cerebral plasticity. Plasticity is greatestin the developing brain, partly because of the overproduc-tion of neurons in the fetus and the overproduction of syn-apses during postnatal development [3]. Excess synapsesare pruned during childhood and early adolescence, inresponse to external experience [3]. Heightened plasticityin the developing brain results in critical periods duringinfancy and childhood when environmental stimuli can cre-ate adaptive or maladaptive changes in the brain’s structure.A threshold level of stimulation may be required for thebrain to develop normally during these critical periods.Plasticity accounts for a child’s superior ability to learna second language or to recover the ability towalk after rad-ical surgeries, such as a hemispherectomy to treat epilepsy[4]. The idea that a child’s brain has more plasticity than anadult brain and can recover more easily from injury isknown as the Kennard principle. Studies of traumatic braininjury indicate that this principle does not always hold true.In fact, adults and older children with diffuse traumaticbrain injury demonstrated better recovery than young chil-dren whomanifested equivalent injuries [5]. Although plas-ticity allows a child’s brain to learn and recover, it can alsobe responsible for maladaptive structural changes that per-sist into adulthood, after environmental insult and abuse [4].Exposure to early stress in humans is associated with neuro-biologic consequences, including the development of epi-leptiform electroencephalogram abnormalities, alterationsin the corpus callosum, and reduced volume and synapticdensity of the hippocampus [6]. As we will review, experi-mental data strongly support the clinical impression that thephysical, cognitive, and psychologic development of chil-dren depends on the quality of early relationships and expe-riences, and that early intervention can enhance early
childhood development and the brain’s ability to recoverfrom injury (Tables 1 and 2).
Evidence Demonstrating Enhancement of CerebralDevelopment and Stages of Plasticity in CerebralDevelopment
The processes of neurulation and neuronal proliferation,migration, and differentiation begin at conception. At birth,most neurons have migrated to their final locations withinthe brain. Subcortical structures can be clearly delineated,and resemble their adult forms (Figs 2, 3). Organizationof the human cortex begins during gestation andcontinues postnatally and into early adulthood (Fig 4). Cor-tical organization is characterized by dendritic and axonalgrowth, the production of synapses, and neuronal and syn-aptic pruning.Differential growth is evident between differ-ent cortical regions [7]. Measures of synaptic densityindicate that synaptogenesis varies by age and brain region[8]. Synaptic production in the occipital cortex begins earlyin the postnatal period and reaches peak density at age 8months, declining to adult levels by early adolescence.The increase in synaptic density in the parietotemporal re-gion occurs after its increase in the occipital regions andpeaks later, at age 3-4 years, and also declines to adult levelsby early adolescence. The frontal lobes lag behind the pari-etotemporal regions by several months throughout infancy.Synaptic density in the frontal regions peaks at age 3-4years, but pruning continues into early adulthood. Datafrom positron emission tomography studies demonstratedthat the maturation of local metabolic rates also varies byage and region, and closely parallels the time course of syn-aptogenesis [9]. Again, the frontal cortex exhibits pro-longed maturation relative to the visual cortex. Theprocess of myelination also varies by age and brain region
Figure 1. Potential depiction of the relationship between neural protection and neural recovery and enhancement. This review emphasizes neural recoveryand enhancement, because this area does not receive as much attention in the literature as neuroprotection.
240 PEDIATRIC NEUROLOGY Vol. 44 No. 4

Author's personal copy
[10,11]. Myelination begins in the peripheral nervoussystem just before birth. Shortly thereafter, myelinappears in the central nervous system in the brainstemand cerebellum and in components of some major sensoryand motor systems. By age 3 years, all of the main fibertracts are identifiable [12]. Myelination within the cerebralhemispheres, particularly those regions involved in higher-level associative functions and sensory discrimination, oc-curs well after birth and progresses over decades (Figs 2, 3).
Neurogenesis and Factors That Can Enhance It
Neurogenesis and programmed cell death are carefullybalanced during fetal development to assure that the correctnumber of neurons populate each region of the brain.Neuro-trophins, such as brain-derived growth factor, promote neu-ronal proliferation and survival in response to environmentalcues. A surplus of neurons is produced in the fetus that al-lows neurons to rewire and recover functionality that islost because of injury. Although neurogenesis peaks duringdevelopment, evidence indicates that neurogenesis persistsinto adulthood in certain regions of the brain, includingthe olfactory system and the hippocampus [13]. In the olfac-tory system, precursor cells reside in the anterior portion ofthe subventricular zone, in thewalls of the lateral ventricles.In the hippocampus, precursor cells are evident in the sub-granular zone of the dentate gyrus [13]. Furthermore, neuro-genesis increases in response to neuronal injury. In neonatalrats with experimental hypoxic injury, neurogenesis is en-hanced and continues in the subventricular zone for monthsafter injury [4]. Sustained neurogenesis in this brain regionwas also documented in adult rodents after stroke [4]. A re-cent study of human patients who died after acute ischemicstroke presented unequivocal evidence of active cell prolif-eration in the ipsilateral subventricular zone [14].
Growing evidence fromanimal studies indicates that envi-ronmental enrichment stimulates neurogenesis in the brain[4,15]. A recent study in mice demonstrated that enrichedstimuli increased progenitor cell numbers and suppressedcell death in the amygdale [16]. In contrast, environmentalstress is known to decrease neurogenesis independent of spe-cies, stressor, or life stage [17]. A study of chronicallystressed rats indicated that environmental enrichment ame-liorated stress-induced decreases in the survival and differ-entiation of progenitor cells, and reversed signs ofbehavioral depression [18]. The increased neurogenesisobserved with enrichment may be mediated by mechanismsinvolvingvascular endothelial growth factor, the recruitmentof T cells, and the activation of microglia [19,20].Enrichment is also thought to promote neurogenesis byincreasing levels of neurotrophins such as brain-derived neu-rotrophic factor andnervegrowth factor [19]. Environmentalenrichment was demonstrated to promote neurogenesis byincreasing neurotrophin levels in animal models of Hunting-ton’s disease, Alzheimer’s disease, Parkinson’s disease, epi-lepsy, traumatic brain injury, and stroke [19].
Synaptogenesis and Factors That Can Enhance It
Neurons transmit information to other neurons throughsynapses. Synaptogenesis refers to the formation of newsynapses. Both spontaneous and environmentally inducedactivity results in the formation and stabilization of synap-ses. An early postnatal burst in synaptogenesis is followedby an activity-dependent pruning of synapses throughoutchildhood. Critical periods for synaptogenesis and pruningvary by brain region. The failure to receive exposure to ap-propriate stimuli during a critical period is difficult orimpossible to remedy subsequently. For example, theproduction of synapses in the visual cortex begins early,plateaus early, and declines to adult levels by early adoles-cence. Consequently, the reversal of unilateral amblyopiamay not be possible after age 12 years, when numbers ofsynapses in the occipital lobes are rapidly declining [21].However, recent animal studies suggest that unilateralamblyopia may be at least partly reversible via anti-gamma-aminobutyric acidergic drugs (whereas gamma-aminobutyric acidergic drugs exert the opposite effect)[22]. In addition, the primary somatosensory cortex is capa-ble of cortical reorganization after injury or of differentialuse into adulthood in response to glutamatergic andgamma-aminobutyric acidergic input [4]. These pharmaco-logic manipulations of cerebral plasticity may hold futureclinical implications.
Synaptogenesis and pruning are mediated by eventsoccurring at the molecular level. Excitatory transmissioncan increase or decrease the strength of a synapse throughlong-term potentiation or long-term depression. Long-term potentiation is produced by the rapid stimulation ofsynapses that use glutamate, whereas long-term depressionis produced by the slow, repetitive stimulation of the samesynapses. Rapid stimulation results in the opening of N-methyl-D-aspartate-type glutamate receptors in the postsyn-aptic membrane. This event leads to an influx of calcium,the activation of signaling cascades (including calcium cal-modulin II), and the insertion of alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionic acid-type glutamate re-ceptors in the postsynaptic membrane. A threshold levelof calcium seems to exist for long-term depression and forthe transition from long-term depression to long-term po-tentiation [23]. The detection of glutamate by astrocytesleads to the activity-dependent release of neurotrophins,including brain-derived neurotrophic factor and tumornecrosis factor-a, which enhance long-term potentiationand promote neuronal survival [24]. In contrast, the slow,repetitive stimulation of the synapse results in long-termdepression through a reduction of alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionic acid-type glutamate re-ceptors in the postsynaptic membrane.
Growing evidence from animal studies suggests that en-vironmental enrichment enhances synaptogenesis in vari-ous regions of the brain. Enrichment was associated withincreased dendritic branching and an increase in the num-ber of dendritic spines in the hippocampus [19].
Holt and Mikati: Care for Child Development 241

Author's personal copy
Enrichment also increases the expression of synaptic pro-teins such as synaptophysin and postsynaptic density-95protein [19]. A study of immature mice indicated that envi-ronmental stimulation resulted in improvements in learn-
ing and memory that were associated with elevatedhippocampal levels of synaptophysin and nerve growthfactor [19]. A study of rats exposed to chronic prenatalstress determined that environmental enrichment during
Table 1. In vivo interventions proven to enhance cerebral plasticity and development in animals
Intervention Model
Environmental enrichment Healthy mouse and rat pups and adults
Murine models:Pups with epilepsyAdults with epilepsy, Huntington’s disease, Alzheimer’s disease, Parkinson’s disease, fragile X, Rett syndrome, amyotrophic lateralsclerosis
Rat models:Pups with stress, epilepsyAdults with stress, amblyopia, hypoxia-ischemia, traumatic brain injury
Licking and grooming Rat models:Healthy rat pupsPups exposed to high levels of corticosterone
Erythropoietin Murine models:Pups with oxidative injury
Rat models:Pups with hypoxia-ischemia, hyperoxiaAdults with epilepsy, traumatic brain injury, intracerebral hemorrhage
Rabbit models:Subarachnoid hemorrhage
Antidepressants Healthy rat adults
Murine models:Pups with epilepsyAdults with diabetes
Rat models:Adults with amblyopia, hypoxia-ischemia
Prenatal choline supplementation Adult rats in status epilepticus
Polyphenol Murine models:Neonates with hypoxia-ischemia
Rat models:Adults with stroke
Omega-3 fatty acids Murine models:Adults with Alzheimer’s disease, Huntington’s disease
Rat models:Healthy neonatesPups with ischemia, epilepsyAdults with Alzheimer’s disease
Hypothermia Rat models:Pups with hypoxia-ischemia
Sheep models:Fetus with hypoxia-ischemia
Gerbil models:Pups with hypoxia-ischemia
Pig models:Piglets with hypoxia-ischemia
Stem cells Murine models:Pups with hypoxia-ischemia
Rat models:Pups with hypoxia-ischemiaAdults with 6-hydroxydopamine lesioned model of Parkinson’s disease
Calcium signaling Murine models:Adults with overexpression of neuronal calcium sensor-1 in dentate gyrus, genetic inhibition of calcineurin in forebrain
Interference with cell-deathpathways
Murine models:Adults with nitric oxide synthase inhibitor in MPTP model of Parkinson’s disease
Rat models:Pups with neuronal nitric oxide synthase inhibitor in hypoxia-ischemia
Growth factors Vascular endothelial growth factor in healthy adult miceNerve growth factor in healthy in adult ratsBasic fibroblast growth factor in healthy rat pups and adults
Murine models:Adults with vascular endothelial growth factor in amyotrophic lateral sclerosis and hypoxia-ischemia
Rat models:Pups with insulin-like growth factor-1 in hypoxia-ischemiaAdults with nerve growth factor in 6-hydroxy-dopamine lesioned model of Parkinson’s disease
Abbreviation:
MPTP = 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine
242 PEDIATRIC NEUROLOGY Vol. 44 No. 4

Author's personal copy
the early postnatal period enhanced hippocampal long-term potentiation and ameliorated stress-induced deficitsin spatial learning and memory [25]. In a recent study,ras-grf knockout mice with defective long-term potentia-
tion and contextual fear-conditioning memory were ex-posed to an enriched environment. This exposureenhanced long-term potentiation not only in the enrichedmice, but in their future offspring through early
Behavioral Effects Cellular and Molecular Effects References
Improved learning, memory, auditory function Increased neurogenesis, decreased apoptosis, increased dendritic branching,increased number of dendritic spines, increased levels of synaptic proteins,increased neurotrophin levels, facilitated long-term potentiation, increasedangiogenesis
15,16,19,26,78
Improved learning, memory, exploratory behavior,restored visual acuity and ocular dominance,reduced reactivity to stress, motor deficits, seizures
Increased neurogenesis, decreased apoptosis, increased number of astrocytes andoligodendrocytes, increased dendritic branching, increased number of dendriticspines, increased levels of synaptic proteins, increased neurotrophin levels,facilitated long-term potentiation
18,19,25,79-87
Enhanced learning under basal conditions,impaired learning under high stress
Longer dendritic branch length, increased number of dendritic spines 27
Improved neurobehavioral outcomes,reduced mortality, motor deficits, seizures
Increased neurogenesis, decreased apoptosis, activated neuroprotective signalingpathways, increased neurotrophin levels, increased neuropeptide Y
28-34,88
Increased neurogenesis, increased number of dendritic spines, increased levels ofsynaptic proteins
40,41,89
Improved motor deficits, restored ocular dominance,visual function
Increased neurogenesis, reduced apoptosis, increased neurotrophin levels,decreased inflammation
4,44,90,91
Improved memory and learning Enhanced preseizure hippocampal levels of growth factors, decreasedseizure-induced hippocampal degeneration, enhanced dentate cellproliferation, attenuated glial fibrillary acidic protein levels, decreased loss ofglutamic acid decarboxylase, altered growth factor expression
54,55
Decreased brain tissue loss, decreased apoptosis 56
Decreased motor deficits, improved learning Enhanced myelinogenesis, reduced oxidative stress, decreased apoptosis,decreased levels of Ab and tau protein, reduced infarct volume, prevention ofepileptiform activity on electroencephalogram
57
Delayed expression of brain damage, reduced neuronal loss 52,92,93
Reduced motor deficits Increased neurogenesis, decreased apoptosis 4,58,59,94
Improved learning, memory, exploratory behavior Facilitated long-term potentiation 95,96
Improved neurologic function, reduced motor deficits Prevented neuronal damage, reduced apoptosis 97,98
Increased neurogenesis 99-101
Prolonged survival, reduced motor deficits Increased neurogenesis, prevented neuronal injury, reduced neuronal loss 102-105
Holt and Mikati: Care for Child Development 243
Author's personal copy
Table 2. Clinical interventions proven to enhance cerebral plasticity and development
Intervention Condition Finding References
Erythropoietin Term infants with moderate hypoxic-
ischemic encephalopathy
Adults with acute stroke
Adults with chronic progressive
multiple sclerosis
Reduced infarct size, improved motor
function, improved cognition
37-39
Antidepressants Adults with depression
Adults with stroke
Reduced hippocampal volume
loss, increased neurogenesis,
increased serum brain-derived
neurotrophic factor,
improved motor
recovery
45-47,106
Adults with posttraumatic stress
disorder
Improved declarative memory,
increased hippocampal volume
107
Omega-3 fatty acids Adults with Alzheimer’s disease
Adults with Huntington’s disease
Adults with multiple sclerosis
Improved mood, cooperativity,
short-term memory, appetite,
sleep, spatial orientation,
decreased cerebral atrophy,
improved motor function,
reduced disability
57
Hypothermia Neonates with hypoxic-ischemic
encephalopathy
Reduced rate of cerebral palsy,
improved development and motor
function
52,53
Peripheral nerve stimulation Adults with stroke Enhanced training effects of
functional hand tasks
4
Repetitive transcranial magnetic
stimulation
Adults and children with stroke
Adults with Parkinson’s disease,
dystonia
Improved motor behavior and
corticospinal output to paretic hand,
improved motor function, reduced
dystonia
49-51,108
Human fetal neural stem cells
Human autologous bone marrow
mesenchymal stem cells
Human fetal neural stem cells
Two-month-old infant with hypoxic-
ischemic encephalopathy
Adults with amyotrophic lateral
sclerosis
Adults with Huntington’s disease
Improved muscle tone, movement,
intelligence, slowed linear decline
of forced vital capacity and of
amyotrophic lateral sclerosis
functional rating score,
improved motor and
cognitive function
60,109,110
Constraint-induced movement
therapy
Adults with stroke Improved motor function, bilaterally
increased gray matter in sensory
and motor areas and both
hippocampi on magnetic
resonance imaging, increased
activation of motor cortices
of undamaged hemisphere
during affected hand movement
on functional magnetic
resonance imaging, increased
size of transcranial magnetic
stimulation motor maps in
affected hemisphere motor
cortex
4,111
Patients aged 12-22 years with
cerebral hemispherectomy
Improved motor function,
reorganization of sensorimotor
cortical representations of both
hands in one hemisphere on
functional magnetic resonance
imaging
68
Children (aged 18 months to 4 years)
with hemiplegic cerebral palsy
Improved motor function 4
High-energy and high-protein diet Infants with perinatal brain injury Increased brain and corticospinal
tract growth
69
244 PEDIATRIC NEUROLOGY Vol. 44 No. 4

Author's personal copy
adolescence, although the offspring never experienced en-vironmental enrichment [26].
Studies in rat pups demonstrate that the frequency of ma-ternal licking and grooming alters synaptic structure andfunction, and results in a differential responsiveness to glu-
cocorticoids and stress. Pups that received a low frequencyof licking and grooming demonstrated shorter dendriticbranch lengths and lower spinal density in CA1 cells [27].In addition, these pups also manifested impaired long-term potentiation under basal conditions, but surprisingly,
Figure 2. In utero development. CNS = central nervous system.
Table 2. Continued
Intervention Condition Finding References
Early developmental intervention
program
Occupational therapy, physical
therapy, psychology,
neurodevelopmental therapy in
preterm infants
Improved cognitive function during
infancy and preschool age, no
improvement in cognitive function
at school age, little effect on motor
outcome during infancy and
preschool age
70,71
Psychosocial stimulation in severely
malnourished infants
Increased growth, improved
development at age 6 months
72
Psychosocial stimulation in low birth
weight infants
Improved cognitive function and
behavior at age 6 years
73
Cognitive and psychosocial therapy
in low-income pregnant mothers
and families with infants and
toddlers
Improved cognitive function and
behavior at age 3 years
75
Holt and Mikati: Care for Child Development 245

Author's personal copy
significantly enhanced long-term potentiation in responseto high levels of corticosterone and stressful conditions.
Factors Enhancing Recovery After Discrete Types ofCerebral Injury
Erythropoeitin
The heightened plasticity of the developing brain in-creases its susceptibility to injury, and also permits its re-
covery from injury. Interventions that modulate themolecules and processes involved in neurogenesis and syn-aptogenesis improve the brain’s ability to recover. For ex-ample, the neuroprotective actions of erythropoietin arewell established in animal models of ischemic brain injury.The immature nervous system is especially sensitive to theeffects of hypoxia and hypoxia-induced seizures. A recentstudy in a kainic acid-induced rat model of seizure revealedthat erythropoietin attenuated the severity of seizures andthe mortality rate. These effects were accompanied by an
Figure 3. Postnatal development.
Figure 4. Synaptogenesis according to brain region. Peak synaptic density in visual, auditory, and prefrontal cortex is indicated in black [8].
246 PEDIATRIC NEUROLOGY Vol. 44 No. 4

Author's personal copy
upregulation of neuropeptide Y and a decrease in both ap-optosis and abnormal cell proliferation in the hippocampus[28]. Erythropoietin was also demonstrated to promoterecovery in animal models of traumatic brain injury, sub-arachnoid hemorrhage, intracerebral hemorrhage, hypoxia,and oxidative injury [29-34]. The mechanisms underlyingerythropoietin-mediated neuroprotection include directneurotrophic effects, decreased susceptibility to glutamatetoxicity, an induction of antiapoptotic factors, decreasedinflammation, decreased nitric oxide-mediated injury,direct antioxidant effects, and protective effects on theglia [35].
Basic science provides extensive evidence for the neuro-protective effects of erythropoietin, and supports the ther-apeutic use of erythropoietin in humans [35]. High-doseerythropoietin treatment is a promising option for neonateswith hypoxic brain injury. Two phase I/II trials of high-dose erythropoietin in preterm infants established its phar-macokinetic and safety profiles [36]. A recent prospective,randomized trial of erythropoietin treatment for term in-fants with moderate hypoxic-ischemic encephalopathydemonstrated reduced disability, without apparent side ef-fects [37]. Erythropoietin also exhibited neuroprotectiveactions in clinical studies of adults with central nervoussystem disease. In adults with a middle cerebral arterystroke, treatment with high-dose erythropoietin reducedinfarct size and promoted functional recovery [38]. Eryth-ropoietin also improved motor function and cognition ina study of patients with chronic, progressive multiple scle-rosis [39]. Further clinical trials are needed to investigateand confirm the potential applications of erythropoietinin central nervous system injury and to optimize treatmentregimens.
Antidepressants
Antidepressants were also linked experimentally withenhanced cerebral plasticity. The chronic administrationof antidepressants promoted neurogenesis, synaptogenesis,and neurotrophic signaling in the hippocampus [40-43].Growing evidence suggests that antidepressants induceneurogenesis and synaptogenesis through the activationof brain-derived neurotrophic factor signaling pathways[42]. A recent study of adult amblyopic rats indicatedthat the chronic administration of fluoxetine reinstated oc-ular dominance plasticity in the visual cortex and promotedthe recovery of visual function [4]. These effects were ac-companied by an increased expression of brain-derivedneurotrophic factor in the visual cortex. Fluoxetine wasalso demonstrated to prevent neuronal cell death throughanti-inflammatory effects in a rat model of cerebralischemia [44].
Antidepressants are widely used in humans to treat ma-jor depression because of their safety and efficacy. In pa-tients with major depressive disorder, antidepressantsreduce hippocampal volume loss, increase neural progeni-tor cells, increase serum brain-derived neurotrophic factor
levels, and improve memory [45,46]. Recent animalstudies support the use of antidepressants after discreteforms of central nervous system injury. In humans,antidepressant therapy was associated with improvedmotor recovery after stroke [47]. The observed recovery,however, is likely attributable to the alleviation of depres-sion as well as the direct neurotrophic effects of antidepres-sants and their ability to enhance cerebral plasticity.Further studies using quantitative structural and functionalimaging are needed to investigate these effects and theunderlying mechanisms further.
Transcranial Magnetic Stimulation
Interventions that increase excitatory neurotransmissionpromote plasticity, whereas interventions that enhance in-hibitory neurotransmission are associated with impairedplasticity. These effects are partly mediated through long-term potentiation and long-term depression, which promoteor inhibit synaptogenesis through different patterns ofexcitatory neurotransmission. A recent study of the somato-sensory cortex in rats revealed that excitatory (glutamate)neurotransmission facilitated somatosensory receptivefield enlargements and somatotopic map expansion,whereas inhibitory (gamma-aminobutyric acid) neuro-transmission prevented these effects [4]. Transcranial mag-netic stimulation and transcranial direct current stimulationcomprise noninvasive techniques that modulate corticalexcitability and plasticity. High-frequencymagnetic stimu-lation is excitatory, and was demonstrated to induce long-term potentiation in rat hippocampal slices [48].
Transcranial magnetic stimulation is used as a treatmentmodality in humans with various neurologic conditions. Astudyof strokepatientswithhemiparesis indicated that a sin-gle session of high-frequency, excitatory transcranial mag-netic stimulation over the stroke hemisphere transientlyimprovedmotor behavior and corticospinal output in paretichands [49]. Randomized, controlled trials in adults andchildrenwith stroke indicated that contralesional, inhibitoryrepetitive transcranial magnetic stimulation results in im-provements in hand function that persist beyond the activetreatment phase [50]. Initial clinical studies demonstratedthat transcranialmagnetic stimulationmayalsodemonstratetherapeutic potential in the treatment of patients with move-ment disorders, including Parkinson’s disease and dystonia[51]. However, the benefits of transcranial magnetic stimu-lation may be short-lived, and frequent, long-term therapymay be required. Larger clinical trials are needed to estab-lish the efficacy and determine the practical applicationsof transcranial magnetic stimulation.
Hypothermia
Reductions in brain temperature by 2-5�C were demon-strated to be neuroprotective in newborn and adult animalmodels of ischemia. Hypothermia is thought to reducebrain injury after ischemia by modulating many of the
Holt and Mikati: Care for Child Development 247

Author's personal copy
pathways involved in cerebral plasticity and the cerebralenergy state. Specifically, brain cooling was demonstratedto decrease excitotoxicity, apoptosis, and the production ofnitric oxide, and to increase cerebral blood flow and metab-olism after ischemic injury [52].
Several clinical trials demonstrated the neuroprotectiveeffects of hypothermia in neonates with hypoxic-ischemicencephalopathy. The National Institute of Child Healthand Human Development Study, a randomized trial of 72hours ofwhole-body hypothermia,was performed in infantswith moderate or severe encephalopathy [52]. The out-comes of death or moderate to severe disability and therisk of cerebral palsy were significantly reduced in infantswho received hypothermia. The multicenter, randomizedTotal Body Hypothermia for Neonatal EncephalopathyStudy compared intensive care plus total body cooling for72 hours of moderate hypothermia with intensive care with-out cooling among term infants with asphyxial encephalop-athy [53]. The study reported a significant improvement inneurologic outcomes among infantswho received hypother-mia, but no significant reduction in the combined rate ofdeath and severe disability. Based on the strength of clinicalevidence supporting the use of prolonged moderate hypo-thermia after asphyxia, cooling procedures are increasinglybeing provided in routine clinical care settings.
Dietary Supplementation
Several dietary supplements, including choline, polyphe-nol, and polyunsaturated fatty acids, have demonstratedneuroprotective effects in animals. Prenatal cholinesupplementation was demonstrated to protect against thelong-term cognitive and neuropathologic effects of statusepilepticus in adult rats [54,55]. Choline supplementationis thought to exert these effects through enhanced growthfactor signaling. Adult offspring of rat dams who receivedprenatal choline supplementation manifest enhancedpreseizure hippocampal levels of brain-derived neurotro-phic factor, nerve growth factor-1, and insulin-like growthfactor-1 [54]. After the administration of kainic acid, theserats exhibited decreased seizure-induced hippocampal de-generation, enhanced dentate cell proliferation, attenuatedglial fibrillary acidic protein levels, a decreased loss of glu-tamic acid decarboxylase, and altered growth factor expres-sion patterns 16 days later. These findings were found topersist 11 weeks after status epilepticus [55].
Polyphenol exhibited neuroprotective effects in animalstudies of hypoxic-ischemic injury and stroke. Foods richin polyphenol include pomegranate juice, blueberries,and green tea extract. Pomegranate juice protects the neo-natal murine brain against hypoxic-ischemic injury whengiven to mothers in their drinking water [56]. Offspringof mothers given pomegranate juice demonstrate decreasedbrain tissue loss after injury [56]. The mechanism of actionis thought to involve the polyphenol resveratrol, whichreduces the activation of caspase-3 and the inactivationof calpain, decreasing apoptosis.
Polyunsaturated fatty acids were demonstrated to pro-tect against neurodegeneration in animal models of Alz-heimer’s disease and Huntington’s disease [57]. Walnutsand fish are well known for their high levels of polyunsat-urated fatty acids. In Alzheimer’s disease, the administra-tion of omega-3 fatty acids is associated with reducedlevels of amyloid and tau protein [57]. Omega-3 fatty acidsare thought to exert their neuroprotective effects by reduc-ing excitotoxicity, oxidative stress, and inflammation.
Several clinical trials in humans investigated the effectsof omega-3 fatty acids on the progression of neurodegener-ative disease. Alzheimer’s disease is strongly correlatedwith low levels of omega-3 fatty acids [57]. However,the results of dietary intervention studies in Alzheimer’sdisease have been mixed. Some groups reported improve-ments in mood and short-term memory with supplemen-tation, whereas others reported no difference [57].Preliminary clinical trials of omega-3 fatty acid supple-mentation in patients with Huntington’s disease havedemonstrated more promise. Patients with advanced Hun-tington’s disease treated with omega-3 fatty acids over 6months exhibited reduced cerebral atrophy on magneticresonance imaging, and improved motor function [57].In clinical studies of multiple sclerosis, supplementationwith omega-3 fatty acids reduced disability, with a treat-ment effect similar to that reported for standard medicaltherapies [57]. Additional clinical studies are needed toinvestigate the role of polyunsaturated fatty acids andother nutritional supplements in the treatment of neurode-generative disease.
Stem Cells
Stem cells are capable of promoting plasticity throughneurotrophic support and the replacement of damagedcells. Bone marrow mesenchymal cells, cord blood cells,fetal cells, and adult neural stem cells have all been demon-strated to promote plasticity in animal models. Studies inneonatal rats with hypoxic-ischemic injury demonstratedthat both intracerebral and intravenous grafts of humanbone marrow-derived multipotent progenitor cells amelio-rate motor deficits and reduce ischemic cell loss in the hip-pocampus [58]. The intraperitoneal transplantation ofhuman cord blood mononuclear cells was demonstratedto reduce spastic paresis in rats exposed to perinatalhypoxic-ischemic damage [59]. A recent study in neonatalmice indicated that human neural stem cells transplantedinto the striatum after hypoxic-ischemic injury decreasedhemispheric brain atrophy [4].
Stem cell therapies are under investigation for a largevariety of neurologic disorders in humans. At present, hu-man autologous stem cells from cord blood have comeclosest to clinical application. No viable stem cell treat-ments exist yet for ischemic brain injury. A single casereport in 2005 documented the successful transplantationof human neural stem cells into the cerebral ventricle ofa 75-day-old boy with severe hypoxic-ischemic
248 PEDIATRIC NEUROLOGY Vol. 44 No. 4
Author's personal copy
encephalopathy [60]. Positron emission tomography indi-cated the survival of implanted cells. The infant was alsoreported to demonstrate remarkable improvement in mus-cle tone, movement, and intelligence, 28 days after trans-plantation. Controlled trials are needed for objectiveassessments of the potential effects of stem cell transplan-tation on neurologic disorders. Clinical trials by a numberof groups, including ours, of stem cell therapy for suchdisorders as stroke, cerebral palsy, spinal cord injury,amyotrophic lateral sclerosis, Parkinson’s disease, Hun-tington’s disease, Pelizaeus-Merzbacher disease, and hu-man neuronal ceroid lipofuscinosis (Batten disease) areunderway [61-63].
Potential Strategies for Enhancement of CerebralDevelopment and Plasticity
Magnetic Resonance Imaging-Based Studies
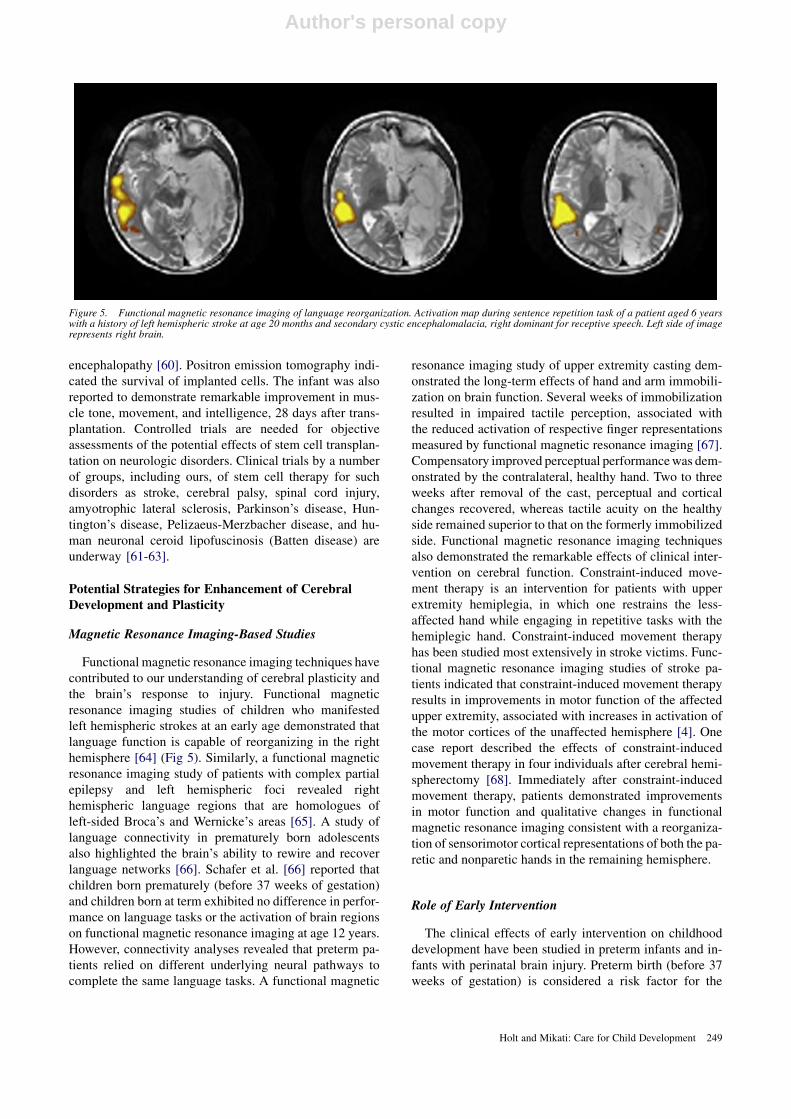
Functional magnetic resonance imaging techniques havecontributed to our understanding of cerebral plasticity andthe brain’s response to injury. Functional magneticresonance imaging studies of children who manifestedleft hemispheric strokes at an early age demonstrated thatlanguage function is capable of reorganizing in the righthemisphere [64] (Fig 5). Similarly, a functional magneticresonance imaging study of patients with complex partialepilepsy and left hemispheric foci revealed righthemispheric language regions that are homologues ofleft-sided Broca’s and Wernicke’s areas [65]. A study oflanguage connectivity in prematurely born adolescentsalso highlighted the brain’s ability to rewire and recoverlanguage networks [66]. Schafer et al. [66] reported thatchildren born prematurely (before 37 weeks of gestation)and children born at term exhibited no difference in perfor-mance on language tasks or the activation of brain regionson functional magnetic resonance imaging at age 12 years.However, connectivity analyses revealed that preterm pa-tients relied on different underlying neural pathways tocomplete the same language tasks. A functional magnetic
resonance imaging study of upper extremity casting dem-onstrated the long-term effects of hand and arm immobili-zation on brain function. Several weeks of immobilizationresulted in impaired tactile perception, associated withthe reduced activation of respective finger representationsmeasured by functional magnetic resonance imaging [67].Compensatory improved perceptual performancewas dem-onstrated by the contralateral, healthy hand. Two to threeweeks after removal of the cast, perceptual and corticalchanges recovered, whereas tactile acuity on the healthyside remained superior to that on the formerly immobilizedside. Functional magnetic resonance imaging techniquesalso demonstrated the remarkable effects of clinical inter-vention on cerebral function. Constraint-induced move-ment therapy is an intervention for patients with upperextremity hemiplegia, in which one restrains the less-affected hand while engaging in repetitive tasks with thehemiplegic hand. Constraint-induced movement therapyhas been studied most extensively in stroke victims. Func-tional magnetic resonance imaging studies of stroke pa-tients indicated that constraint-induced movement therapyresults in improvements in motor function of the affectedupper extremity, associated with increases in activation ofthe motor cortices of the unaffected hemisphere [4]. Onecase report described the effects of constraint-inducedmovement therapy in four individuals after cerebral hemi-spherectomy [68]. Immediately after constraint-inducedmovement therapy, patients demonstrated improvementsin motor function and qualitative changes in functionalmagnetic resonance imaging consistent with a reorganiza-tion of sensorimotor cortical representations of both the pa-retic and nonparetic hands in the remaining hemisphere.
Role of Early Intervention
The clinical effects of early intervention on childhooddevelopment have been studied in preterm infants and in-fants with perinatal brain injury. Preterm birth (before 37weeks of gestation) is considered a risk factor for the
Figure 5. Functional magnetic resonance imaging of language reorganization. Activation map during sentence repetition task of a patient aged 6 yearswith a history of left hemispheric stroke at age 20 months and secondary cystic encephalomalacia, right dominant for receptive speech. Left side of imagerepresents right brain.
Holt and Mikati: Care for Child Development 249

Author's personal copy
development of motor and cognitive disabilities. Dabydeenet al. indicated that a high-energy and high-protein diet in-creases brain and corticospinal tract growth in term andpreterm infants with perinatal brain injury. They did not re-port whether any corresponding improvements in cognitivefunction occurred [69]. Early intervention programs in-volving occupational therapy, physical therapy, psychol-ogy, and neurodevelopmental therapy aim to reducemotor and cognitive impairments in preterm infants. In2007, a largemeta-analysis of 16 studies revealed that earlydevelopmental intervention programs improved cognitiveperformance during infancy and preschool age [70]. Lim-ited evidence suggests that early developmental interven-tions improve outcomes in the long term (during schoolage). Little effect of early intervention on motor outcomeswas evident in the short, medium, or long term, but onlytwo studies have reported outcomes beyond age 2 years.In 2009, the same researchers presented a meta-analysisof 11 randomized studies grouped by intervention, age atoutcome, and study quality [71]. Early developmental in-tervention was observed to improve cognitive outcomesduring infancy and preschool age. The benefit, however,was not sustained at school age. No effect of early interven-tion on motor outcomes was evident during infancy orschool age, and no data were available for motor outcomesat preschool age. Based on these reviews, early develop-mental programs appear to exert a short-term effect on cog-nitive development in preterm infants. However, a long-term effect on cognition and effects on motor developmentcannot be excluded, because of the considerable variabilitybetween studies with regard to primary outcome measure,focus of intervention, and method of delivery. The method-ological quality of studies included in the reviews was vari-able, and sample sizes were quite small in many of thestudies. The length of follow-up and the follow-up rateswere also highly variable. In addition, these studies in-volved premature infants and not other infants manifestingcerebral insult at birth or during infancy, and large-scalesubgroup analysis was not possible.
Research from developing countries and the UnitedStates has demonstrated the benefits of early cognitiveand psychosocial interventions in specific patient popula-tions. A study of severely malnourished infants in Bangla-desh investigated the effects of psychosocial stimulation ongrowth and development [72]. Children who received 2weeks of in-hospital group meetings and play sessionsand 6 months of home visits demonstrated higher scoreson the Bayley Scales of Infant Development and increasedgrowth at age 6 months. In a large, prospective cohort studyof low birth weight Jamaican children, subjects receiveddietary supplementation (1 kg of milk-based formula perweek), psychosocial stimulation, both types of interven-tion, or no intervention. Although the study reported nosignificant effects of dietary supplementation, psychoso-cial stimulation up to age 2 years was demonstrated tobenefit cognition at age 6 years [73,74]. The stimulatedgroup demonstrated higher intelligence quotients and
memory scores and fewer behavioral difficulties than thecontrol group. In the United States, researchers studiedthe effectiveness of Early Head Start, a federal programthat provides cognitive and psychosocial interventions forlow-income pregnant women and families withinfants and toddlers [75]. A randomized trial of 3001families in 17 programs indicated that children in theprogram performed better in cognitive and language devel-opment, and exhibited better behavior and attention at age3 years.
Conclusions and Outlook for the Future
Evidence from neurobiology indicates that early inter-vention is capable of changing the structure and functionof the brain. The biologicmechanisms of neurogenesis, syn-aptogenesis, and rewiring underlie cerebral plasticity andthe brain’s ability to adapt and recover from injury. Environ-mental enrichment alonewas demonstrated to increase neu-rogenesis and synaptogenesis in the brain. Erythropoietin,growth factors, antidepressants, and stem cells comprisesome of the biologic agents capable of augmenting thebrain’s ability to recover from injury. Promising advanceshave occurred in the neurobiology of cerebral plasticity.These advances have implications for our ability to inter-vene with and enhance childhood development. The chal-lenge involves identifying those advances with practical,clinical applications. Currently, theWorldHealthOrganiza-tion and the International Pediatric Association have pro-grams in place, such as Care for Child Development,Child Feeding Modules, and the Early Childhood Develop-ment International Pediatric Association Program, whichprovide health workers with techniques for working withfamilies to improve the growth, health, and developmentof children in the community [76]. These organizationsalso promote research targeted at the development of effec-tive, sustainable early childhood interventions. ThePakistanEarlyChildhoodDevelopmentScale-UpTrial, supported byUNICEF, is in its third year at the Aga Khan UniversityMedical School [77]. This trial aims to evaluate whetherthe integration of early childhood psychosocial stimulation,alone or in combination with nutritional interventions, im-proves development in children up to age 24months in ruralPakistan. Further studies are needed to identify interven-tions that are safe and effective at the clinical level. The im-plementation of early intervention requires the support oflocal and global communities and a commitment to helpingchildren reach their full developmental potential. These ap-proaches have been increasingly supported by convincingclinical as well as basic science data that address not onlyneuroprotection but also the potentiation of cerebral plastic-ity and function.
References
[1] Grantham-McGregor S, Cheung YB, Cueto S, Glewwe P,
Richter L, Strupp B. International Child Development Steering Group.
Lancet 2007;369:60-70.
250 PEDIATRIC NEUROLOGY Vol. 44 No. 4

Author's personal copy
[2] Hebb DO. The organization of behavior. New York: Wiley, 1949.
[3] Johnston MV, Nishimura A, Harum K, Pekar J, Blue ME.
Sculpting the developing brain. Adv Pediatr 2001;48:1-38.
[4] JohnstonMV. Plasticity in the developing brain: Implications for
rehabilitation. Dev Disabil Res Rev 2009;15:94-101.
[5] Laurent-Vannier A, Brugel DG, De Agostini M. Rehabilitation
of brain-injured children. Childs Nerv Syst 2000;16:760-4.
[6] Teicher MH, Tomoda A, Anderson SL. Neurobiological conse-
quences of early stress and childhood maltreatment: Are results from
human and animal studies comparable? Ann NY Acad Sci 2006;1071:
313-23.
[7] Johnson MH. Functional brain development in humans. Nat Rev
Neurosci 2001;2:475-83.
[8] Huttenlocher PR, Dabholkar AS. Regional differences in syn-
aptogenesis in human cerebral cortex. J Comp Neurol 1997;387:
167-78.
[9] ChuganiHT, PhelpsME,Mazziotta JC. Positron emission tomog-
raphy study of human brain functional development. Ann Neurol 1987;22:
487-97.
[10] YakovlevPI,LecoursAR.Themyelogenetic cycles of regionalmat-
uration of the brain. In: Minkowski A, editor. Regional development of the
brain in early life. Oxford: Blackwell Scientific Publications, 1967:3-70.
[11] Gilles FH, Shankle W, Dooling EC. Myelinated tracts: Growth
patterns. In: Wright J, editor. The developing human brain growth and
epidemiologic neuropathology. Boston: John Wright-PSG, Inc., 1983:
117-83.
[12] Matsuzawa J, Matsui M, Konishi T, et al. Age-related volumet-
ric changes of brain gray and white matter in healthy infants and children.
Cereb Cortex 2001;11:335-42.
[13] Balu DT, Lucki I. Adult hippocampal neurogenesis: Regulation,
functional implications, and contribution to disease pathology. Neurosci
Biobehav Rev 2009;33:232-52.
[14] Marti-Fabregas J, Romaguera-Ros M, Gomez-Pinedo U, et al.
Proliferation in the human ipsilateral subventricular zone after ischemic
stroke. Neurology 2010;74:357-65.
[15] Kempermann G, Kuhn HG, Gage FH. More hippocampal
neurons in adult mice living in an enriched environment. Nature 1997;
386:493-5.
[16] OkudaH, Tatsumi K, Makinodan M, Yamauchi T, Kishimoto T,
Wanaka A. Environmental enrichment stimulates progenitor cell prolifer-
ation in the amygdala. J Neurosci Res 2009;87:3546-53.
[17] Mirescu C, Gould E. Stress and adult neurogenesis. Hippocam-
pus 2006;16:233-8.
[18] Veena J, Srikumar BN, Raju TR, Shankaranarayana Rao BS. Ex-
posure to enriched environment restores the survival and differentiation of
new born cells in the hippocampus and ameliorates depressive symptoms
in chronically stressed rats. Neurosci Lett 2009;455:178-82.
[19] Nithianantharajah J, Hannan AJ. Enriched environments,
experience-dependent plasticity and disorders of the nervous system.
Nat Rev Neurosci 2006;7:697-709.
[20] CaoL, Jiao X, Zuzga DS, et al. VEGF links hippocampal activity
with neurogenesis, learning and memory. Nat Genet 2004;36:827-35.
[21] Holmes JM, Clarke MP. Amblyopia. Lancet 2006;367:1343-51.
[22] Sale A, Berardi N, Spolidoro M, Baroncelli L, Maffei L.
GABAergic inhibition in visual cortical plasticity. Front Cell Neurosci
2010;4:10.
[23] Cormier RJ, Greenwood AC, Connor JA. Bidirectional synaptic
plasticity correlated with the magnitude of dendritic calcium transients
above a threshold. J Neurophysiol 2001;85:399-406.
[24] Fellin T. Communication between neurons and astrocytes: Rel-
evance to the modulation of synaptic and network activity. J Neurochem
2009;108:533-44.
[25] Yang J, Hou C, Ma N, et al. Enriched environment treatment re-
stores impaired hippocampal synaptic plasticity and cognitive deficits in-
duced by prenatal chronic stress. Neurobiol Learn Mem 2007;87:257-63.
[26] Arai JA, Li S, HartleyDM, Feig LA. Transgenerational rescue of
a genetic defect in long-term potentiation and memory formation by juve-
nile enrichment. J Neurosci 2009;29:1496-502.
[27] Champagne DL, Bagot RC, van Hasselt F, et al. Maternal
care and hippocampal plasticity: Evidence for experience-dependent
structural plasticity, altered synaptic functioning, and differential re-
sponsiveness to glucocorticoids and stress. J Neurosci 2008;28:
6037-45.
[28] KondoA, Shingo T, Yasuhara T, et al. Erythropoietin exerts anti-
epileptic effects with the suppression of aberrant new cell formation in the
dentate gyrus and upregulation of neuropeptide Y in seizure model of rats.
Brain Res 2009;1296:127-36.
[29] Grasso G, Sfacteria A, Meli F, Fodale V, Buemi M,
Iacopino DG. Neuroprotection by erythropoietin administration after ex-
perimental traumatic brain injury. Brain Res 2007;1182:99-105.
[30] Grasso G, Buemi M, Alafaci C, et al. Beneficial effects of sys-
temic administration of recombinant human erythropoietin in rabbits sub-
jected to subarachnoid hemorrhage. Proc Natl Acad Sci USA 2002;99:
5627-31.
[31] Liao ZB, Zhi XG, Shi QH, He ZH. Recombinant human eryth-
ropoietin administration protects cortical neurons from traumatic brain in-
jury in rats. Eur J Neurol 2008;15:140-9.
[32] Seyfried DM, Han Y, Yang D, Ding J, Chopp M. Erythropoietin
promotes neurological recovery after intracerebral haemorrhage in rats.
Int J Stroke 2009;4:250-6.
[33] Juul SE, McPherson RJ, Bammler TK, Wilkerson J, Beyer RP,
Farin FM. Recombinant erythropoietin is neuroprotective in a novel
mouse oxidative injury model. Dev Neurosci 2008;30:231-42.
[34] MikatiMA, El Hokayem JA, El Sabban ME. Effects of a single
dose of erythropoietin on subsequent seizure susceptibility in rats exposed
to acute hypoxia at P10. Epilepsia 2007;48:175-81.
[35] McPherson RJ, Juul SE. Recent trends in erythropoietin-
mediated neuroprotection. Int J Dev Neurosci 2008;26:103-11.
[36] McPherson RJ, Juul SE. Erythropoietin for infants with
hypoxic-ischemic encephalopathy. Curr Opin Pediatr 2010;22:139-45.
[37] Zhu C, KangW, Xu F, et al. Erythropoietin improves neurologic
outcomes in newborns with hypoxic-ischemic encephalopathy. Pediatrics
2009;124:e218-26.
[38] Ehrenreich H, Hasselblatt M, Dembowski C, et al. Erythropoi-
etin therapy for acute stroke is both safe and beneficial. Mol Med 2002;8:
495-505.
[39] Ehrenreich H, Fischer B, Norra C, et al. Exploring recombinant
human erythropoietin in chronic progressive multiple sclerosis. Brain
2007;130:2577-88.
[40] Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antide-
pressant treatment increases neurogenesis in adult rat hippocampus.
J Neurosci 2000;20:9104-10.
[41] Hajszan T, MacLusky NJ, Leranth C. Short-term treatment
with the antidepressant fluoxetine triggers pyramidal dendritic spine
synapse formation in rat hippocampus. Eur J Neurosci 2005;21:
1299-303.
[42] Castr�en E. Neurotrophic effects of antidepressant drugs. Curr
Opin Pharmacol 2004;4:58-64.
[43] Santarelli L, Saxe M, Gross C, et al. Requirement of hippocam-
pal neurogenesis for the behavioral effects of antidepressants. Science
2003;301:805-9.
[44] Lim CM, Kim SW, Park JY, Kim C, Yoon SH, Lee JK. Fluoxe-
tine affords robust neuroprotection in the postischemic brain via its anti-
inflammatory effect. J Neurosci Res 2009;87:1037-45.
[45] Boldrini M, Underwood MD, Hen R, et al. Antidepressants in-
crease neural progenitor cells in the human hippocampus. Neuropsycho-
pharmacology 2009;34:2376-89.
[46] Huang TL, Lee CT, Liu YL. Serum brain-derived neurotrophic
factor levels in patients with major depression: Effects of antidepressants.
J Psychiatr Res 2008;42:521-5.
[47] Pariente J, Loubinoux I, Carel C, et al. Fluoxetine modulates
motor performance and cerebral activation of patients recovering from
stroke. Ann Neurol 2001;50:718-29.
[48] Tokay T, Holl N, Kirschstein T, Zschorlich V, K€ohling R. High-frequency magnetic stimulation induces long-term potentiation in rat hip-
pocampal slices. Neurosci Lett 2009;461:150-4.
Holt and Mikati: Care for Child Development 251

Author's personal copy
[49] Talelli P, Greenwood RJ, Rothwell JC. Exploring theta burst
stimulation as an intervention to improve motor recovery in chronic
stroke. Clin Neurophysiol 2007;118:333-42.
[50] Kirton A, Chen R, Friefeld S, Gunraj C, Pontigon AM,
Deveber G. Contralesional repetitive transcranial magnetic stimulation
for chronic hemiparesis in subcortical paediatric stroke: A randomised
trial. Lancet Neurol 2008;7:507-13.
[51] EdwardsM, Talelli P, Rothwell J. Clinical applications of trans-
cranial magnetic stimulation in patients with movement disorders. Lancet
Neurol 2008;7:827-40.
[52] Shankaran S, Laptook AR, Ehrenkranz RA, et al. National In-
stitute of Child Health and Human Development Neonatal Research Net-
work. Whole-body hypothermia for neonates with hypoxic-ischemic
encephalopathy. N Engl J Med 2005;353:1574-84.
[53] Azzopardi DV, Strohm B, Edwards AD, et al.; TOBY Study
Group. Moderate hypothermia to treat perinatal asphyxial encephalopa-
thy. N Engl J Med 2009;361:1349-58.
[54] Wong-Goodrich SJ, Mellott TJ, Glenn MJ, Blusztajn JK,
Williams CL. Prenatal choline supplementation attenuates neuropatholog-
ical response to status epilepticus in the adult rat hippocampus. Neurobiol
Dis 2008;30:255-69.
[55] Wong-Goodrich SJ, Glenn MJ, Mellott TJ, Liu YB,
Blusztajn JK, Williams CL. Water maze experience and prenatal cho-
line supplementation differentially promote long-term hippocampal re-
covery from seizures in adulthood. Hippocampus 2010. Mar 15 (Epub
ahead of print).
[56] WestT, AtzevaM, HoltzmanDM. Pomegranate polyphenols and
resveratrol protect the neonatal brain against hypoxic-ischemic injury.
Dev Neurosci 2007;29:363-72.
[57] Dyall SC, Michael-Titus AT. Neurological benefits of omega-3
fatty acids. Neuromol Med 2008;10:219-35.
[58] Yasuhara T, Hara K, Maki M, et al. Intravenous grafts recapitu-
late the neurorestoration afforded by intracerebrally delivered multipotent
adult progenitor cells in neonatal hypoxic-ischemic rats. J Cereb Blood
Flow Metab 2008;28:1804-10.
[59] MeierC,Middelanis J,Wasielewski B, et al. Spastic paresis after
perinatal brain damage in rats is reduced by human cord blood mononu-
clear cells. Pediatr Res 2006;59:244-9.
[60] Luan Z, Yin GC, Hu XH, et al. Treatment of an infant with se-
vere neonatal hypoxic-ischemic encephalopathy sequelae with transplan-
tation of human neural stem cells into cerebral ventricle. Zhonghua Er Ke
Za Zhi 2005;43:580-3 [in Chinese].
[61] Zietlow R, Lane EL, Dunnett SB, Rosser AE. Human stem cells
for CNS repair. Cell Tissue Res 2008;331:301-22.
[62] Naegele JR, Maisano X, Yang J, Royston S, Ribeiro E. Recent
advancements in stem cell and gene therapies for neurological disorders
and intractable epilepsy. Neuropharmacology 2010;58:855-64.
[63] Schwarz SC, Schwarz J. Translation of stem cell therapies for
neurological diseases. Transl Res 2010;156:155-60.
[64] Li�egeois F, Connelly A, Cross JH, et al. Language reorganization
in children with early-onset lesions of the left hemisphere: An fMRI study.
Brain 2004;127:1229-36.
[65] Rosenberger LR, Zeck J, Berl MM, et al. Interhemispheric and
intrahemispheric language reorganization in complex partial epilepsy.
Neurology 2009;72:1830-6.
[66] Schafer RJ, Lacadie C, Vohr B, et al. Alterations in functional
connectivity for language in prematurely born adolescents. Brain 2009;
132:661-70.
[67] Lissek S,Wilimzig C, Stude P, et al. Immobilization impairs tac-
tile perception and shrinks somatosensory cortical maps. Curr Biol 2009;
19:837-42.
[68] de Bode S, Fritz SL, Weir-Haynes K, Mathern GW. Constraint-
induced movement therapy for individuals after cerebral hemispherec-
tomy: A case series. Phys Ther 2009;89:361-9.
[69] Dabydeen L, Thomas JE, Aston TJ, Hartley H, Sinha SK,
Eyre JA. High-energy and -protein diet increases brain and corticospinal
tract growth in term and preterm infants after perinatal brain injury. Pedi-
atrics 2008;121:148-56.
[70] Spittle AJ, Orton J, Doyle LW, Boyd R. Early developmental in-
tervention programs post hospital discharge to prevent motor and cogni-
tive impairments in preterm infants. Cochrane Database Syst Rev 2007;
2: CD005495.
[71] Orton J, Spittle A, Doyle L, Anderson P, Boyd R. Do early in-
tervention programmes improve cognitive and motor outcomes for pre-
term infants after discharge? A systematic review. Dev Med Child
Neurol 2009;51:851-9.
[72] Nahar B, Hamadani JD, Ahmed T, et al. Effects of psychoso-
cial stimulation on growth and development of severely malnourished
children in a nutrition unit in Bangladesh. Eur J Clin Nutr 2009;63:
725-31.
[73] Walker SP, Chang SM, Younger N, Grantham-McGregor SM.
The effect of psychosocial stimulation on cognition and behaviour at 6
years in a cohort of term, low-birthweight Jamaican children. Dev Med
Child Neurol 2010;52:e148-54.
[74] Walker SP, Chang SM, Powell CA, Simonoff E, Grantham-
McGregor SM. Effects of psychosocial stimulation and dietary
supplementation in early childhood on psychosocial functioning in late
adolescence: Follow-up of randomised controlled trial. Br Med J [Clin
Res] 2006;333:472.
[75] Love JM, Kisker EE, Ross C, et al. The effectiveness of early
head start for 3-year-old children and their parents: Lessons for policy
and programs. Dev Psychol 2005;41:885-901.
[76] WHO-UNICEF. WHO-UNICEF Meeting on Promoting Early
Childhood Development. Colombo, Sri Lanka, July 2009. Geneva:
WHO, 2009.
[77] Aga Khan University. The Pakistan Early Childhood Develop-
ment Scale Up Trial. Bethesda, MD: National Library of Medicine, 2000.
Available at: http://clinicaltrials.gov/ct2/show/study/NCT00715936. Ac-
cessed March 18, 2010.
[78] Ekstrand J, Hellsten J, Tingstr€omA. Environmental enrichment,
exercise and corticosterone affect endothelial cell proliferation in adult rat
hippocampus and prefrontal cortex. Neurosci Lett 2008;442:203-7.
[79] Korbey SM, Heinrichs SC, Leussis MP. Seizure susceptibility
and locus ceruleus activation are reduced following environmental en-
richment in an animal model of epilepsy. Epilepsy Behav 2008;12:
30-8.
[80] Pereira LO, Nabinger PM, Strapasson AC, et al. Long-term ef-
fects of environmental stimulation following hypoxia-ischemia on the ox-
idative state and BDNF levels in rat hippocampus and frontal cortex. Brain
Res 2009;1247:188-95.
[81] Herring A, Ambr�ee O, Tomm M, et al. Environmental enrich-
ment enhances cellular plasticity in transgenic mice with Alzheimer-
like pathology. Exp Neurol 2009;216:184-92.
[82] Hoffman AN, Malena RR, Westergom BP, et al. Environmental
enrichment-mediated functional improvement after experimental trau-
matic brain injury is contingent on task-specific neurobehavioral experi-
ence. Neurosci Lett 2008;431:226-30.
[83] Kondo M, Gray LJ, Pelka GJ, Christodoulou J, Tam PP,
Hannan AJ. Environmental enrichment ameliorates a motor coordination
deficit in a mouse model of Rett syndrome— Mecp2 gene dosage effects
and BDNF expression. Eur J Neurosci 2008;27:3342-50.
[84] G€ortzN, Lewejohann L, TommM, et al. Effects of environmen-
tal enrichment on exploration, anxiety, and memory in female TgCRND8
Alzheimer mice. Behav Brain Res 2008;191:43-8.
[85] Nag N, Moriuchi JM, Peitzman CG, Ward BC, Kolodny NH,
Berger-Sweeney JE. Environmental enrichment alters locomotor behav-
iour and ventricular volume in Mecp2 1lox mice. Behav Brain Res
2009;196:44-8.
[86] Kazl C, Foote LT, Kim MJ, Koh S. Early-life experience alters
response of developing brain to seizures. Brain Res 2009;1285:174-81.
[87] Sale A, Maya Vetencourt JF, Medini P, et al. Environmental en-
richment in adulthood promotes amblyopia recovery through a reduction
of intracortical inhibition. Nat Neurosci 2007;10:679-81.
[88] Kaindl AM, Sifringer M, Koppelstaetter A, et al. Erythropoietin
protects the developing brain from hyperoxia-induced cell death and pro-
teome changes. Ann Neurol 2008;64:523-34.
252 PEDIATRIC NEUROLOGY Vol. 44 No. 4

Author's personal copy
[89] Sairanen M, O’Leary OF, Knuuttila JE, Castr�en E. Chronic an-
tidepressant treatment selectively increases expression of plasticity-
related proteins in the hippocampus and medial prefrontal cortex of the
rat. Neuroscience 2007;144:368-74.
[90] Beauquis J,RoigP,DeNicolaAF,SaraviaF.Neuronal plasticity and
antidepressants in the diabetic brain. Ann NYAcad Sci 2009;1153:203-8.
[91] JinY, LimCM,Kim SW, et al. Fluoxetine attenuates kainic acid-
induced neuronal cell death in the mouse hippocampus. Brain Res 2009;
1281:108-16.
[92] Trescher WH, Ishiwa S, Johnston MV. Brief post-hypoxic-
ischemic hypothermia markedly delays neonatal brain injury. Brain Dev
1997;19:326-38.
[93] George S, Scotter J, Dean JM, et al. Induced cerebral hypother-
mia reduces post-hypoxic loss of phenotypic striatal neurons in preterm
fetal sheep. Exp Neurol 2007;203:137-47.
[94] Yasuhara T, Matsukawa N, Hara K, et al. Transplantation of hu-
man neural stem cells exerts neuroprotection in a rat model of Parkinson’s
disease. J Neurosci 2006;26:12497-511.
[95] Saab BJ, Georgiou J, Nath A, et al. NCS-1 in the dentate gyrus
promotes exploration, synaptic plasticity, and rapid acquisition of spatial
memory. Neuron 2009;63:643-56.
[96] Malleret G, Haditsch U, Genoux D, et al. Inducible and revers-
ible enhancement of learning, memory, and long-term potentiation by ge-
netic inhibition of calcineurin. Cell 2001;104:675-86.
[97] Feng Y, Piletz JE, Leblanc MH. Agmatine suppresses nitric ox-
ide production and attenuates hypoxic-ischemic brain injury in neonatal
rats. Pediatr Res 2002;52:606-11.
[98] Watanabe Y, Kato H, Araki T. Protective action of neuronal ni-
tric oxide synthase inhibitor in theMPTPmousemodel of Parkinson’s dis-
ease. Metab Brain Dis 2008;23:51-69.
[99] Sun Y, Jin K, Childs JT, Xie L, Mao XO, Greenberg DA. Vascu-
lar endothelial growth factor-B (VEGFB) stimulates neurogenesis: Evi-
dence from knockout mice and growth factor administration. Dev Biol
2006;289:329-35.
[100] Frielingsdorf H, Simpson DR, Thal LJ, Pizzo DP. Nerve
growth factor promotes survival of new neurons in the adult hippocampus.
Neurobiol Dis 2007;26:47-55.
[101] Wagner JP, Black IB, DiCicco-BloomE. Stimulation of neona-
tal and adult brain neurogenesis by subcutaneous injection of basic fibro-
blast growth factor. J Neurosci 1999;19:6006-16.
[102] WangY,MaoXO, Xie L, et al. Vascular endothelial growth fac-
tor overexpression delays neurodegeneration and prolongs survival in
amyotrophic lateral sclerosis mice. J Neurosci 2007;27:304-7.
[103] Wang Y, Jin K, Mao XO, et al. VEGF-overexpressing trans-
genic mice show enhanced post-ischemic neurogenesis and neuromigra-
tion. J Neurosci Res 2007;85:740-7.
[104] Chaturvedi RK, Shukla S, Seth K, Agrawal AK. Nerve growth
factor increases survival of dopaminergic graft, rescue nigral dopaminer-
gic neurons and restores functional deficits in rat model of Parkinson’s dis-
ease. Neurosci Lett 2006;398:44-9.
[105] Lin S, Fan LW, Rhodes PG, Cai Z. Intranasal administration of
IGF-1 attenuates hypoxic-ischemic brain injury in neonatal rats. Exp Neu-
rol 2009;217:361-70.
[106] Matrisciano F, Bonaccorso S, Ricciardi A, et al. Changes in
BDNF serum levels in patients with major depression disorder (MDD) af-
ter 6 months treatment with sertraline, escitalopram, or venlafaxine. J Psy-
chiatr Res 2009;43:247-54.
[107] Vermetten E, Vythilingam M, Southwick SM, Charney DS,
Bremner JD. Long-term treatment with paroxetine increases verbal de-
clarative memory and hippocampal volume in posttraumatic stress disor-
der. Biol Psychiatry 2003;54:693-702.
[108] Nowak DA, Grefkes C, Dafotakis M, et al. Effects of low-
frequency repetitive transcranial magnetic stimulation of the contrale-
sional primary motor cortex on movement kinematics and neural activity
in subcortical stroke. Arch Neurol 2008;65:741-7.
[109] Mazzini L, Mareschi K, Ferrero I, et al. Stem cell treatment in
amyotrophic lateral sclerosis. J Neurol Sci 2008;265:78-83.
[110] Bachoud-L�evi AC, Gaura V, Brugi�eres P, et al. Effect of
fetal neural transplants in patients with Huntington’s disease 6 years
after surgery: A long-term follow-up study. Lancet Neurol 2006;5:
303-9.
[111] Wolf SL, Winstein CJ, Miller JP, et al. Retention of upper limb
function in stroke survivorswhohave receivedconstraint-inducedmovement
therapy: The EXCITE randomised trial. Lancet Neurol 2008;7:33-40.
Holt and Mikati: Care for Child Development 253