author's personal copy - mentalhealth.kr · author's personal copy available ......
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Available online at www.sciencedirect.com
European Journal of Integrative Medicine 4 (2012) e392–e399
Original article
Antidepressant-like effects of Bupleuri Radix extract
Mi Kyoung Seo a,c, Jae Chun Song c, Sun Jung Lee c, Kyung Ah Koo d, Yong Ki Park e,Jung Goo Lee a,b,c, Sung Woo Park c,∗, Young Hoon Kim a,b,c,∗
a Department of Psychiatry, School of Medicine, Inje University, Busan, Republic of Koreab Haeundae Paik Hospital, Inje University, Busan, Republic of Korea
c Paik Institute for Clinical Research, Inje University, Busan, Republic of Koread Department of Biological Science, University of the Science in Philadelphia, Philadelphia, USA
e College of Oriental Medicine, Dongguk University, Gyeongju, Republic of Korea
Received 2 February 2012; received in revised form 9 July 2012; accepted 26 July 2012
Abstract
Study aims: Some oriental herbal medicines have been used to treat neuropsychiatric disorders. We found previously that Kami-shoyo-san (KSS)possesses antidepressant-like effects at both the behavioral and molecular levels. Bupleuri Radix (BR), a major component of KSS, has affinityfor both dopamine D2 and 5-HT1A receptors, which may be responsible for its antidepressant-like effects. In the present study, we investigated theantidepressant-like effects of BR in rats using the forced swimming test (FST) and the potential mechanisms of BR action in SH-SY5Y cells.Materials and methods: The immobility time of rats treated with BR extract was measured in the FST. Western blotting was performed to examinethe expression of proteins involved in the cAMP response element binding protein (CREB)/brain-derived neurotrophic factor (BDNF)-mediatedsignaling pathway, thought to be a major target of antidepressant drug action.Results: BR extract (900 mg/kg) significantly reduced immobility time in the FST. Treatment with BR extract (10, 100 and 1000 �g/mL) signifi-cantly increased CREB phosphorylation and elevated BDNF levels in SH-SY5Y cells. BR extract increased phosphorylation of Akt and glycogensynthase kinase-3� (GSK-3�). Moreover, BDNF small interfering RNA (siRNA) reduced elevated Akt and GSK-3� phosphorylation induced byBR extract. A phosphatidylinositol 3-kinase (PI3K) inhibitor blocked the BR-induced increase in phosphorylated Akt levels.Conclusions: These findings suggest that an aqueous extract of BR can exert antidepressant-like effects in behavioral and molecular assays. Theseresults demonstrate that BR may exert its effects through actions on CREB and BDNF activation, leading to stimulation of the PI3K/Akt/GSK-3�signaling pathway.© 2012 Elsevier GmbH. All rights reserved.
Keywords: Antidepressant effect; Brain-derived neurotrophic factor; Bupleuri Radix; Forced swimming test; Glycogen synthase kinase-3�
Introduction
Depression is one of the most common psychiatric disorders,with a lifetime incidence of up to 20% in the general population,and with high associated morbidity and mortality [1]. Despitethe development of various classes of antidepressant drugs, theyare not completely effective and have a high propensity to cause
∗ Corresponding authors at: Paik Institute for Clinical Research, Inje Univer-sity, 633-165 Gaegum-dong, Jin-gu, Busan 614-735, Republic of Korea.Tel.: +82 51 890 6749; fax: +82 51 894 6709.
E-mail addresses: [email protected] (S.W. Park), [email protected](Y.H. Kim).
serious side effects [2]. Thus, there is a continuing need forresearch into alternative antidepressant drug therapies.
Herbal medicines have been used for centuries in a numberof Asian countries to treat various neuropsychiatric disor-ders. Kami-shoyo-san (KSS) is a traditional medicine used inKorea, Japan, and China to treat stress-related neuropsychiatricdisorders, such as depression or anxiety [3,4]. We previ-ously reported that KSS, a combined preparation of PaeoniaeRadix, Bupleuri Radix, Atractylodis Macrocephalae Rhizoma,Liriopis Tuber, Angelicae Gigantis Radix, Hoelen, MenthaeFolium, Glycyrrhizae Radix, and Zingiberis Rhizoma, producedantidepressant-like effects at both the behavioral and molecu-lar levels [5]. Among KSS components, we sought to identifythe active compound that exerts antidepressant effects. Bupleuri
1876-3820/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.http://dx.doi.org/10.1016/j.eujim.2012.07.979

Author's personal copy
M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399 e393
Radix (BR), a major component of KSS, is frequently includedin herbal formulas designed to treat stress-related mental illness[6]. BR, one of the most important Chaihu-Shugan-San com-ponents, reportedly has significant antidepressant-like effectsin animal models of depression [7]. However, no studies haveexamined the BR mechanisms involved in producing the antide-pressant effects. The major chemical constituents of BR aresaikosaponin, adonitol, �-spinasterol, and other volatile oils,and aqueous extracts of this herb have affinity for 5-HT1A anddopamine D2 receptors [8,9].
In vitro and in vivo data provide evidence that the transcrip-tion factor, cAMP response element binding protein (CREB),and one of its target genes, brain-derived neurotrophic factor(BDNF), are major mediators in the action of antidepressantdrugs [10]. CREB and BDNF levels are elevated followingtreatment with different classes of antidepressant drugs, sug-gesting that CREB and BDNF play important roles in thetherapeutic response to antidepressants drugs [11,12]. Phospho-rylation of CREB at serine133 can occur via activation of thecAMP/protein kinase A (PKA) pathway, which can be regulatedby various antidepressant drugs via serotonin, norepinephrine,and dopamine receptors [10]. CREB activation causes increasedsecretion of BDNF, which acts via tyrosine kinase B (TrkB)receptors. Furthermore, transgenic over-expression of TrkB inthe brain results in antidepressant-like behavioral response inanimal models [13].
However, it is unclear how increased BDNF influencesantidepressant action. We suggest that it may be associated withthe activity of glycogen synthase kinase-3� (GSK-3�). GSK-3�, an ubiquitous cellular serine/threonine protein kinase, wasoriginally shown to regulate glycogen synthesis in responseto insulin, but is now recognized as a downstream regulatorthat mediates multiple signaling pathways coupled to receptors(e.g., neurotransmitter and neurotrophic factor receptors) [14].BDNF activation of TrkB stimulates several signaling pathways,including the phosphatidylinositol 3-kinase (PI3K)/Akt andRas/mitogen-activated protein kinase pathway [15]. In partic-ular, activation of PI3K/Akt signaling inhibits GSK-3� activityby phosphorylation on serine9 [16]. GSK-3� is implicated inthe pathophysiology of bipolar disorder because of the effectsof mood stabilizers (e.g., lithium or valproate) on GSK-3� activ-ity [17]. Moreover, GSK-3� dysfunction might also be causativein depression [18]. Similarly, inhibition of GSK-3 activity pro-duces antidepressant-like effects in the forced swimming test(FST) in rodents [19–21]. This suggests, therefore, that GSK-3� inhibition may be involved in antidepressant action. Despitegrowing interest in the possible involvement of GSK-3�, thereare few studies regarding signaling pathways associated withGSK-3� in antidepressant action.
The aim of the present study was to explore possibleantidepressant-like effects of Bupleuri Radix extract. First, weinvestigated the potential antidepressant effects of BR usingthe FST in rats, a standard procedure used to screen com-pounds for antidepressant activity. Second, we investigated theBR mechanism of action associated with BDNF: specifically,PI3K/Akt/GSK-3� pathways in SH-SY5Y human neuroblas-toma cells.
Materials and methods
Materials
Imipramine was purchased from Sigma (St. Louis, MO,USA). Dulbecco’s modified Eagle’s medium (DMEM), fetalbovine serum (FBS) and antibiotic–antimycotic were purchasedfrom Gibco BRL (Grand Island, NY, USA). Antibodies used forWestern blotting were obtained from the following sources: anti-phosphorylated GSK-3� (Ser 9), anti-GSK-3 and anti-BDNFwere obtained from Santa Cruz Biotechnology (Santa Cruz,CA, USA); anti-phosphorylated CREB (Ser 133) was fromUpstate Biotech (Upstate, NY, USA); anti-phosphorylated-Akt(Ser 473), anti-Akt and anti-CREB were from Cell SignalingTechnology (Cell Signaling, Beverly, MA, USA); anti-�-tubulinwas purchased from Sigma; ECL anti-mouse IgG, horseradishperoxidase linked species-specific whole antibody was fromAmersham Pharmacia (Amersham, Little Chalfont, Bucking-hamshire, UK) and anti-goat and anti-rabbit IgG-horseradishperoxide conjugates from Santa Cruz Biotechnology. The PI3Kinhibitor wortmannin was purchased from Invitrogen (Carlsbad,CA, USA). All other chemicals were purchased from commer-cial sources.
Plant material
Bupleuri Radix root was authenticated by Dr. Y.-K. Park, oneof the authors. A voucher specimen (BR-W-0303) was depositedin the College of Oriental Medicine, Dongguk University herbar-ium.
Preparation of Bupleuri Radix extract
BR was air dried and extracts were prepared by boiling thesample in 200 mL of water at 100 ◦C for 2 h. The procedure wasrepeated twice. The extracts were then filtered, concentratedusing a vacuum evaporator, and lyophilized into powder. Thetotal extract yield was 20% (w/w). In herbal formulas designedto treat mental illness, the human adult recommended daily doseis 4–8 g [5,7]. Previous animal studies reported antidepressantaction at 10 times above the clinical human dose [5,7,22]. Thus,rats were administered doses at 10 times and 1.5 times the recom-mended daily dose for humans (i.e., approximately 600 mg/kgand 900 mg/kg, respectively).
Animals
All animal experiments were performed in accordance withanimal care and use guidelines of the US National Insti-tutes of Health (NIH publication no. 23-85, revised 1996) andthe Korean Academy of Medical Science. All experimentsinvolving animals were approved by the Committee for Ani-mal Experimentation and the Institutional Animal LaboratoryReview Board of Inje Medical College (approval no. 2009-037).Male Sprague–Dawley rats (Orient Bio, GyeongGi-Do, Korea)weighing 200–250 g were housed (three per cage) with freely

Author's personal copy
e394 M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399
accessible food and water, in a room maintained at 21 ◦C with a12 h:12 h light:dark cycle.
Effects of Bupleuri Radix on immobility time in the FST
The FST, developed by Porsolt [23,24], was performed withminor modifications as described in our previous report [5].Briefly, rats forced to swim in a restricted space adopt a typi-cal immobile posture after an initial period of vigorous attemptsto escape. The observed immobility reflects a state of despair,and studies have shown that immobility is selectively reducedby a variety of treatments that are therapeutically effectivein depression. We used transparent plastic containers (40 cmheight × 18 cm diameter) filled with water (23–25 ◦C), deepenough that rats could not touch the bottom or escape. The swimtest consisted of three stages. First, each rat was given an ini-tial 15-min pretest. Twenty-four hours later, four groups of rats(6–8 animals) were administered saline, 20 mg/kg of imipramine(i.p.), BR 600 mg/kg (p.o.), or BR 900 mg/kg (p.o). All drugswere administered at 14:00–15:00 h for 7 days. Finally, a 5-minswimming session was performed 1 h after the last treatment.This session was videotaped for later scoring. The amount oftime the animal was active (e.g., swimming, exploring, or try-ing to escape) and immobile (not struggling and making onlythose movements necessary to keep its head above water) weremeasured.
Cell culture
SH-SY5Y human neuroblastoma cells, obtained from theAmerican Type Tissue Culture Collection (ATCC; Rockville,MD, USA), were cultured in medium with an equal amount ofDMEM supplemented with 10% heat-inactivated FST and 1%antibiotic–antimycotic. The cells were maintained in a humid-ified atmosphere of 5% CO2 at 37 ◦C. The culture mediumwas changed every 2–3 days. BR extract was dissolved in dis-tilled water (DW) and diluted with DMEM to a concentration of1000 mg/mL. Cells were treated with various BR extract con-centrations (final concentrations; 10, 100, and 1000 �g/mL).
Small interfering RNA-mediated BDNF knockdown
BDNF knockdown using small interfering RNAs (siRNAs)was performed as follows. BDNF-specific (101263) and con-trol scrambled (SN-1013) siRNA were purchased from Bioneer(Daejon, Korea). For Western blotting, SH-SY5Y cells (50–60%confluent; 6-well plates) were transfected with 50 nM of siRNAusing the Lipofectamine RNAiMAX transfection reagent (Invit-rogen, CA, USA) according to the manufacturer’s protocol. Thegrowth medium was changed 6 h later, and cells were treatedwith BR extract during the exchange. The Western blot anal-yses to detect Akt and GSK-3� phosphorylation and BDNFlevels were performed a minimum of 2 days after siRNAtransfection.
Western blotting
Cells were seeded at a density of 2 × 106 cells/100-mmdish. After 24 h incubation, cells were treated with various BRextract concentrations for 72 h. Whole cell lysates were preparedas follows. Cells were washed twice with ice-cold phosphatebuffered saline (PBS). Lysis buffer [20 mM Tris–HCl, 137 mMNaCl, 10% glycerol, 1% Nonidet p-40, 0.1% sodium dode-cyl sulfate (SDS), 0.5% sodium deoxycholate, 2 mM EDTA,1 tablet complete protease inhibitor (Roche, Laval, Quebec,Canada), 20 mM NaF and 1 mM Na3VO4] was then added.Cells were then centrifuged (1000 × g, 10 min, 4 ◦C). Equalamounts of protein (20 �g) from the cell extracts of each treat-ment condition were separated using SDS polyacrylamide gelelectrophoresis (SDS-PAGE) and then electrophoretically trans-ferred onto polyvinylidene fluoride (PVDF) membranes. PVDFmembranes were blocked by incubation in 5% (w/v) nonfat milkin Tris buffered saline (TBS) with 0.15% Tween 20 (TBS-T)for 1 h. After primary antibody (anti-phosphor-ser9-GSK-3�,1:1000; anti-phosphor-ser473-Akt, 1:1000; anti-BDNF, 1:1000;anti-phosphor-ser133-CREB, 1:1000; anti-GSK-3, 1:1000; anti-Akt, 1:1000, anti-CREB, 1:1000; or anti-�-tubulin, 1:2000)in TBS-T at 4 ◦C overnight, the membranes were washedthree times in TBS-T for 10 min. The membranes werethen incubated for 1 h in TBS-T containing a horseradishperoxidase-conjugated secondary antibody (donkey anti-goatIgG for anti-phospho-ser9-GSK-3�, 1:10,000; goat-anti-rabbitIgG for anti-phospho-ser473-Akt, 1:1000, anti-Akt, 1:2000,anti-phospho-ser133-CREB, 1:1000, anti-CREB, 1:2000, anti-BDNF, 1:10,000; ECL anti-mouse IgG for anti-GSK-3, 1:2000,or anti-�-tubulin, 1:10,000). Immunoreactive bands were visual-ized and quantified using enhanced chemiluminescence (ECL+).Western blotting reagents, with chemifluorescence detected byLas-3000 Image Reader (Fuji Film, Tokyo, Japan) software.Protein levels were normalized to the housekeeping protein �-tubulin to adjust for variability in protein loading, and expressedas a percentage of vehicle control (deemed to be 100%).
Statistical analysis
For the FST, the Kruskal–Wallis was used to detect signif-icant differences in immobility time among groups, followedby Mann–Whitney U test. To analyze cell experiments, weperformed repeated-measures one-way analysis of variance(ANOVA) followed by Scheffe’s post hoc tests. p values ≤ 0.05were considered statistically significant.
Results
Effects of Bupleuri Radix on immobility time in the forcedswimming test
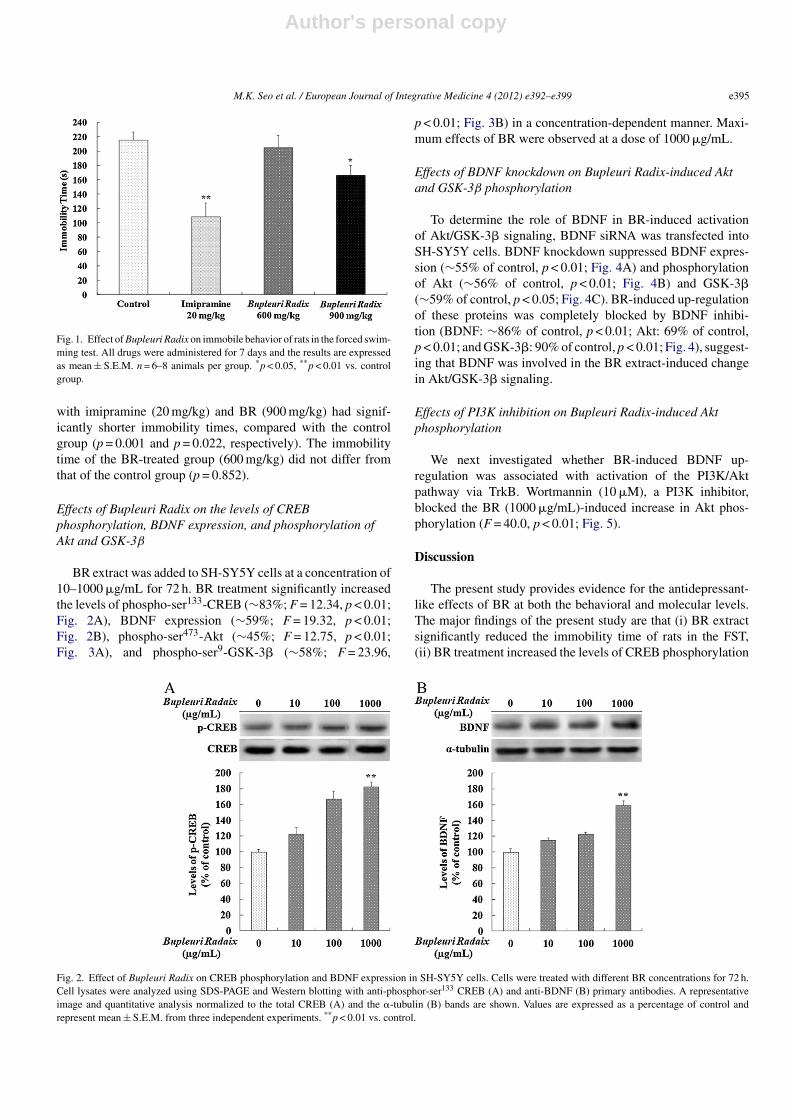
Immobility times in the forced swimming test of the fourgroups are shown in Fig. 1. The immobility times of thecontrol, imipramine (20 mg/kg)-, BR (600 mg/kg)-, and BR(900 mg/kg)-treated groups were 215.8 ± 11.1, 108.7 ± 19.0,205.3 ± 16.7, and 166.1 ± 14.0 s, respectively. Groups treated
Author's personal copy
M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399 e395
Fig. 1. Effect of Bupleuri Radix on immobile behavior of rats in the forced swim-ming test. All drugs were administered for 7 days and the results are expressedas mean ± S.E.M. n = 6–8 animals per group. *p < 0.05, **p < 0.01 vs. controlgroup.
with imipramine (20 mg/kg) and BR (900 mg/kg) had signif-icantly shorter immobility times, compared with the controlgroup (p = 0.001 and p = 0.022, respectively). The immobilitytime of the BR-treated group (600 mg/kg) did not differ fromthat of the control group (p = 0.852).
Effects of Bupleuri Radix on the levels of CREBphosphorylation, BDNF expression, and phosphorylation ofAkt and GSK-3β
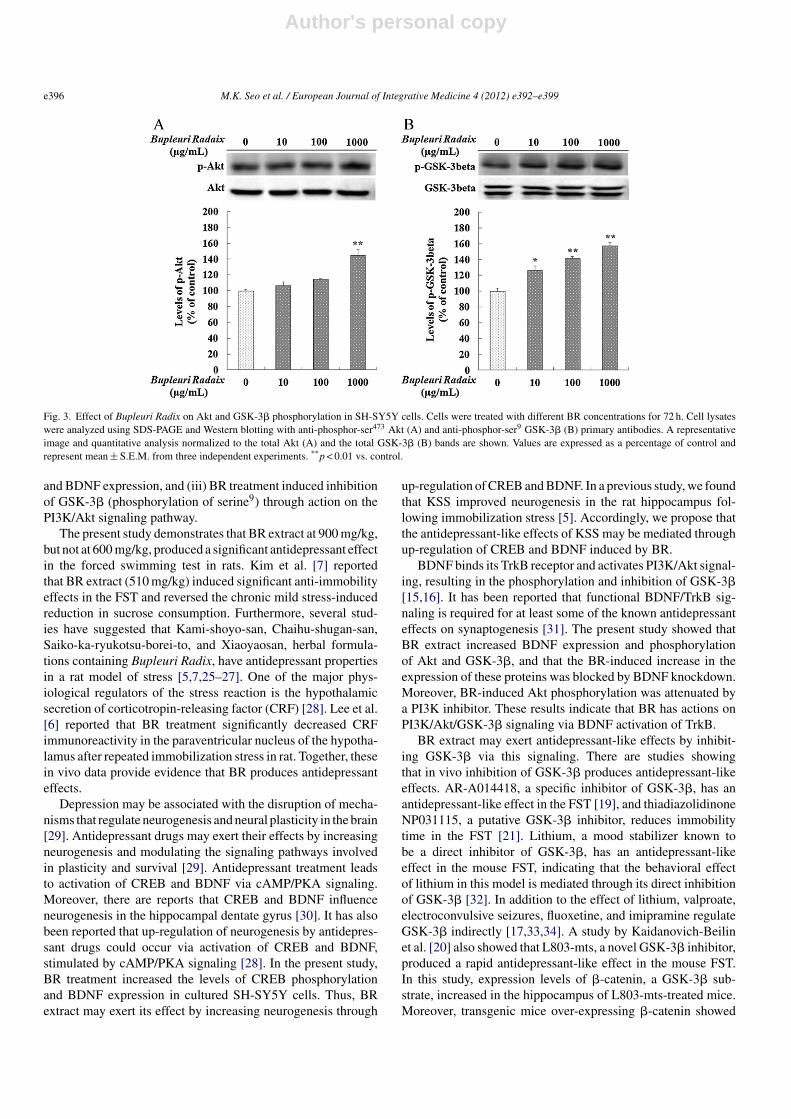
BR extract was added to SH-SY5Y cells at a concentration of10–1000 �g/mL for 72 h. BR treatment significantly increasedthe levels of phospho-ser133-CREB (∼83%; F = 12.34, p < 0.01;Fig. 2A), BDNF expression (∼59%; F = 19.32, p < 0.01;Fig. 2B), phospho-ser473-Akt (∼45%; F = 12.75, p < 0.01;Fig. 3A), and phospho-ser9-GSK-3� (∼58%; F = 23.96,
p < 0.01; Fig. 3B) in a concentration-dependent manner. Maxi-mum effects of BR were observed at a dose of 1000 �g/mL.
Effects of BDNF knockdown on Bupleuri Radix-induced Aktand GSK-3β phosphorylation
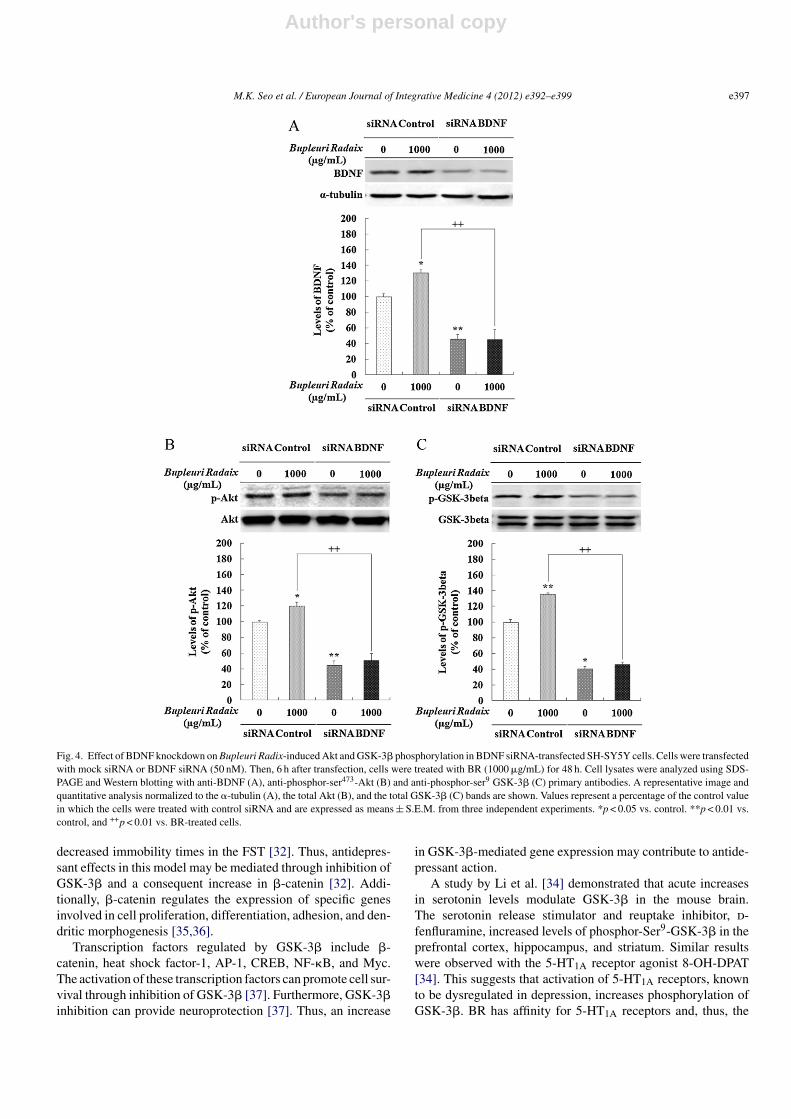
To determine the role of BDNF in BR-induced activationof Akt/GSK-3� signaling, BDNF siRNA was transfected intoSH-SY5Y cells. BDNF knockdown suppressed BDNF expres-sion (∼55% of control, p < 0.01; Fig. 4A) and phosphorylationof Akt (∼56% of control, p < 0.01; Fig. 4B) and GSK-3�(∼59% of control, p < 0.05; Fig. 4C). BR-induced up-regulationof these proteins was completely blocked by BDNF inhibi-tion (BDNF: ∼86% of control, p < 0.01; Akt: 69% of control,p < 0.01; and GSK-3�: 90% of control, p < 0.01; Fig. 4), suggest-ing that BDNF was involved in the BR extract-induced changein Akt/GSK-3� signaling.
Effects of PI3K inhibition on Bupleuri Radix-induced Aktphosphorylation
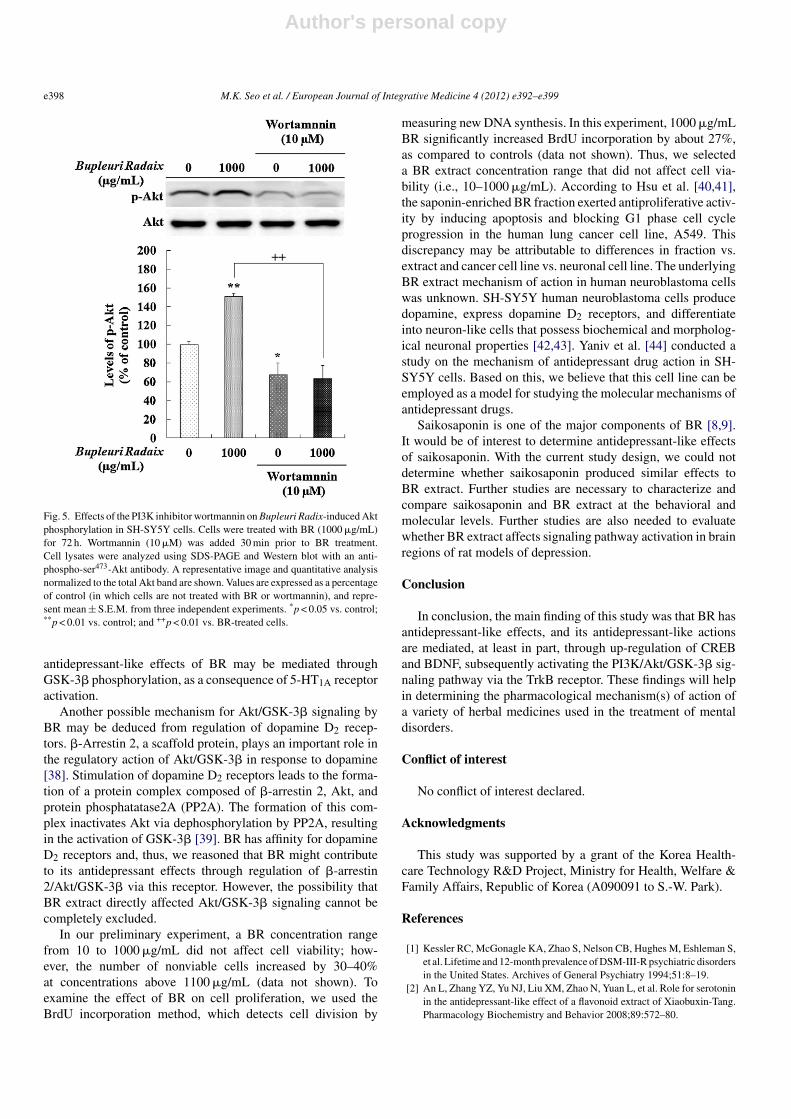
We next investigated whether BR-induced BDNF up-regulation was associated with activation of the PI3K/Aktpathway via TrkB. Wortmannin (10 �M), a PI3K inhibitor,blocked the BR (1000 �g/mL)-induced increase in Akt phos-phorylation (F = 40.0, p < 0.01; Fig. 5).
Discussion
The present study provides evidence for the antidepressant-like effects of BR at both the behavioral and molecular levels.The major findings of the present study are that (i) BR extractsignificantly reduced the immobility time of rats in the FST,(ii) BR treatment increased the levels of CREB phosphorylation
Fig. 2. Effect of Bupleuri Radix on CREB phosphorylation and BDNF expression in SH-SY5Y cells. Cells were treated with different BR concentrations for 72 h.Cell lysates were analyzed using SDS-PAGE and Western blotting with anti-phosphor-ser133 CREB (A) and anti-BDNF (B) primary antibodies. A representativeimage and quantitative analysis normalized to the total CREB (A) and the �-tubulin (B) bands are shown. Values are expressed as a percentage of control andrepresent mean ± S.E.M. from three independent experiments. **p < 0.01 vs. control.
Author's personal copy
e396 M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399
Fig. 3. Effect of Bupleuri Radix on Akt and GSK-3� phosphorylation in SH-SY5Y cells. Cells were treated with different BR concentrations for 72 h. Cell lysateswere analyzed using SDS-PAGE and Western blotting with anti-phosphor-ser473 Akt (A) and anti-phosphor-ser9 GSK-3� (B) primary antibodies. A representativeimage and quantitative analysis normalized to the total Akt (A) and the total GSK-3� (B) bands are shown. Values are expressed as a percentage of control andrepresent mean ± S.E.M. from three independent experiments. **p < 0.01 vs. control.
and BDNF expression, and (iii) BR treatment induced inhibitionof GSK-3� (phosphorylation of serine9) through action on thePI3K/Akt signaling pathway.
The present study demonstrates that BR extract at 900 mg/kg,but not at 600 mg/kg, produced a significant antidepressant effectin the forced swimming test in rats. Kim et al. [7] reportedthat BR extract (510 mg/kg) induced significant anti-immobilityeffects in the FST and reversed the chronic mild stress-inducedreduction in sucrose consumption. Furthermore, several stud-ies have suggested that Kami-shoyo-san, Chaihu-shugan-san,Saiko-ka-ryukotsu-borei-to, and Xiaoyaosan, herbal formula-tions containing Bupleuri Radix, have antidepressant propertiesin a rat model of stress [5,7,25–27]. One of the major phys-iological regulators of the stress reaction is the hypothalamicsecretion of corticotropin-releasing factor (CRF) [28]. Lee et al.[6] reported that BR treatment significantly decreased CRFimmunoreactivity in the paraventricular nucleus of the hypotha-lamus after repeated immobilization stress in rat. Together, thesein vivo data provide evidence that BR produces antidepressanteffects.
Depression may be associated with the disruption of mecha-nisms that regulate neurogenesis and neural plasticity in the brain[29]. Antidepressant drugs may exert their effects by increasingneurogenesis and modulating the signaling pathways involvedin plasticity and survival [29]. Antidepressant treatment leadsto activation of CREB and BDNF via cAMP/PKA signaling.Moreover, there are reports that CREB and BDNF influenceneurogenesis in the hippocampal dentate gyrus [30]. It has alsobeen reported that up-regulation of neurogenesis by antidepres-sant drugs could occur via activation of CREB and BDNF,stimulated by cAMP/PKA signaling [28]. In the present study,BR treatment increased the levels of CREB phosphorylationand BDNF expression in cultured SH-SY5Y cells. Thus, BRextract may exert its effect by increasing neurogenesis through
up-regulation of CREB and BDNF. In a previous study, we foundthat KSS improved neurogenesis in the rat hippocampus fol-lowing immobilization stress [5]. Accordingly, we propose thatthe antidepressant-like effects of KSS may be mediated throughup-regulation of CREB and BDNF induced by BR.
BDNF binds its TrkB receptor and activates PI3K/Akt signal-ing, resulting in the phosphorylation and inhibition of GSK-3�[15,16]. It has been reported that functional BDNF/TrkB sig-naling is required for at least some of the known antidepressanteffects on synaptogenesis [31]. The present study showed thatBR extract increased BDNF expression and phosphorylationof Akt and GSK-3�, and that the BR-induced increase in theexpression of these proteins was blocked by BDNF knockdown.Moreover, BR-induced Akt phosphorylation was attenuated bya PI3K inhibitor. These results indicate that BR has actions onPI3K/Akt/GSK-3� signaling via BDNF activation of TrkB.
BR extract may exert antidepressant-like effects by inhibit-ing GSK-3� via this signaling. There are studies showingthat in vivo inhibition of GSK-3� produces antidepressant-likeeffects. AR-A014418, a specific inhibitor of GSK-3�, has anantidepressant-like effect in the FST [19], and thiadiazolidinoneNP031115, a putative GSK-3� inhibitor, reduces immobilitytime in the FST [21]. Lithium, a mood stabilizer known tobe a direct inhibitor of GSK-3�, has an antidepressant-likeeffect in the mouse FST, indicating that the behavioral effectof lithium in this model is mediated through its direct inhibitionof GSK-3� [32]. In addition to the effect of lithium, valproate,electroconvulsive seizures, fluoxetine, and imipramine regulateGSK-3� indirectly [17,33,34]. A study by Kaidanovich-Beilinet al. [20] also showed that L803-mts, a novel GSK-3� inhibitor,produced a rapid antidepressant-like effect in the mouse FST.In this study, expression levels of �-catenin, a GSK-3� sub-strate, increased in the hippocampus of L803-mts-treated mice.Moreover, transgenic mice over-expressing �-catenin showed
Author's personal copy
M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399 e397
Fig. 4. Effect of BDNF knockdown on Bupleuri Radix-induced Akt and GSK-3� phosphorylation in BDNF siRNA-transfected SH-SY5Y cells. Cells were transfectedwith mock siRNA or BDNF siRNA (50 nM). Then, 6 h after transfection, cells were treated with BR (1000 �g/mL) for 48 h. Cell lysates were analyzed using SDS-PAGE and Western blotting with anti-BDNF (A), anti-phosphor-ser473-Akt (B) and anti-phosphor-ser9 GSK-3� (C) primary antibodies. A representative image andquantitative analysis normalized to the �-tubulin (A), the total Akt (B), and the total GSK-3� (C) bands are shown. Values represent a percentage of the control valuein which the cells were treated with control siRNA and are expressed as means ± S.E.M. from three independent experiments. *p < 0.05 vs. control. **p < 0.01 vs.control, and ++p < 0.01 vs. BR-treated cells.
decreased immobility times in the FST [32]. Thus, antidepres-sant effects in this model may be mediated through inhibition ofGSK-3� and a consequent increase in �-catenin [32]. Addi-tionally, �-catenin regulates the expression of specific genesinvolved in cell proliferation, differentiation, adhesion, and den-dritic morphogenesis [35,36].
Transcription factors regulated by GSK-3� include �-catenin, heat shock factor-1, AP-1, CREB, NF-�B, and Myc.The activation of these transcription factors can promote cell sur-vival through inhibition of GSK-3� [37]. Furthermore, GSK-3�inhibition can provide neuroprotection [37]. Thus, an increase
in GSK-3�-mediated gene expression may contribute to antide-pressant action.
A study by Li et al. [34] demonstrated that acute increasesin serotonin levels modulate GSK-3� in the mouse brain.The serotonin release stimulator and reuptake inhibitor, d-fenfluramine, increased levels of phosphor-Ser9-GSK-3� in theprefrontal cortex, hippocampus, and striatum. Similar resultswere observed with the 5-HT1A receptor agonist 8-OH-DPAT[34]. This suggests that activation of 5-HT1A receptors, knownto be dysregulated in depression, increases phosphorylation ofGSK-3�. BR has affinity for 5-HT1A receptors and, thus, the
Author's personal copy
e398 M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399
Fig. 5. Effects of the PI3K inhibitor wortmannin on Bupleuri Radix-induced Aktphosphorylation in SH-SY5Y cells. Cells were treated with BR (1000 �g/mL)for 72 h. Wortmannin (10 �M) was added 30 min prior to BR treatment.Cell lysates were analyzed using SDS-PAGE and Western blot with an anti-phospho-ser473-Akt antibody. A representative image and quantitative analysisnormalized to the total Akt band are shown. Values are expressed as a percentageof control (in which cells are not treated with BR or wortmannin), and repre-sent mean ± S.E.M. from three independent experiments. *p < 0.05 vs. control;**p < 0.01 vs. control; and ++p < 0.01 vs. BR-treated cells.
antidepressant-like effects of BR may be mediated throughGSK-3� phosphorylation, as a consequence of 5-HT1A receptoractivation.
Another possible mechanism for Akt/GSK-3� signaling byBR may be deduced from regulation of dopamine D2 recep-tors. �-Arrestin 2, a scaffold protein, plays an important role inthe regulatory action of Akt/GSK-3� in response to dopamine[38]. Stimulation of dopamine D2 receptors leads to the forma-tion of a protein complex composed of �-arrestin 2, Akt, andprotein phosphatatase2A (PP2A). The formation of this com-plex inactivates Akt via dephosphorylation by PP2A, resultingin the activation of GSK-3� [39]. BR has affinity for dopamineD2 receptors and, thus, we reasoned that BR might contributeto its antidepressant effects through regulation of �-arrestin2/Akt/GSK-3� via this receptor. However, the possibility thatBR extract directly affected Akt/GSK-3� signaling cannot becompletely excluded.
In our preliminary experiment, a BR concentration rangefrom 10 to 1000 �g/mL did not affect cell viability; how-ever, the number of nonviable cells increased by 30–40%at concentrations above 1100 �g/mL (data not shown). Toexamine the effect of BR on cell proliferation, we used theBrdU incorporation method, which detects cell division by
measuring new DNA synthesis. In this experiment, 1000 �g/mLBR significantly increased BrdU incorporation by about 27%,as compared to controls (data not shown). Thus, we selecteda BR extract concentration range that did not affect cell via-bility (i.e., 10–1000 �g/mL). According to Hsu et al. [40,41],the saponin-enriched BR fraction exerted antiproliferative activ-ity by inducing apoptosis and blocking G1 phase cell cycleprogression in the human lung cancer cell line, A549. Thisdiscrepancy may be attributable to differences in fraction vs.extract and cancer cell line vs. neuronal cell line. The underlyingBR extract mechanism of action in human neuroblastoma cellswas unknown. SH-SY5Y human neuroblastoma cells producedopamine, express dopamine D2 receptors, and differentiateinto neuron-like cells that possess biochemical and morpholog-ical neuronal properties [42,43]. Yaniv et al. [44] conducted astudy on the mechanism of antidepressant drug action in SH-SY5Y cells. Based on this, we believe that this cell line can beemployed as a model for studying the molecular mechanisms ofantidepressant drugs.
Saikosaponin is one of the major components of BR [8,9].It would be of interest to determine antidepressant-like effectsof saikosaponin. With the current study design, we could notdetermine whether saikosaponin produced similar effects toBR extract. Further studies are necessary to characterize andcompare saikosaponin and BR extract at the behavioral andmolecular levels. Further studies are also needed to evaluatewhether BR extract affects signaling pathway activation in brainregions of rat models of depression.
Conclusion
In conclusion, the main finding of this study was that BR hasantidepressant-like effects, and its antidepressant-like actionsare mediated, at least in part, through up-regulation of CREBand BDNF, subsequently activating the PI3K/Akt/GSK-3� sig-naling pathway via the TrkB receptor. These findings will helpin determining the pharmacological mechanism(s) of action ofa variety of herbal medicines used in the treatment of mentaldisorders.
Conflict of interest
No conflict of interest declared.
Acknowledgments
This study was supported by a grant of the Korea Health-care Technology R&D Project, Ministry for Health, Welfare &Family Affairs, Republic of Korea (A090091 to S.-W. Park).
References
[1] Kessler RC, McGonagle KA, Zhao S, Nelson CB, Hughes M, Eshleman S,et al. Lifetime and 12-month prevalence of DSM-III-R psychiatric disordersin the United States. Archives of General Psychiatry 1994;51:8–19.
[2] An L, Zhang YZ, Yu NJ, Liu XM, Zhao N, Yuan L, et al. Role for serotoninin the antidepressant-like effect of a flavonoid extract of Xiaobuxin-Tang.Pharmacology Biochemistry and Behavior 2008;89:572–80.

Author's personal copy
M.K. Seo et al. / European Journal of Integrative Medicine 4 (2012) e392–e399 e399
[3] Mantani N, Hisanaga A, Kogure T, Kita T, Shimada Y, Terasawa K. Fourcases of panic disorder successfully treated with Kampo (Japanese herbal)medicines: Kami-shoyo-san and Hange-koboku-to. Psychiatry and ClinicalNeurosciences 2002;56:617–20.
[4] Yamada K, Kanba S. Herbal medicine (Kami-shoyo-san) in the treatment ofpremenstrual dysphoric disorder. Journal of Clinical Psychopharmacology2002;22:442.
[5] Park SW, Kim YK, Lee JG, Kim SH, Kim JM, Yoon JS, et al.Antidepressant-like effects of the traditional Chinese medicine Kami-shoyo-san in rats. Psychiatry and Clinical Neurosciences 2007;61:401–6.
[6] Lee B, Shim I, Lee H, Hahm DH. Effect of Bupleurum falcatum on thestress-induced impairment of spatial working memory in rats. Biologicaland Pharmaceutical Bulletin 2009;32:1392–8.
[7] Kim SH, Han J, Seog DH, Chung JY, Kim N, Park HY, et al. Antidepressanteffect of Chaihu-Shugan-San extract and its constituents in rat models ofdepression. Life Sciences 2005;76:1297–306.
[8] Liao JF, Jan YM, Huang SY, Wang HH, Yu LL, Chen CF. Evaluation withreceptor binding assay on the water extracts of ten CNS-active Chineseherbal drugs. Proceedings of the National Science Council, Republic ofChina Part B, Life Sciences 1995;19:151–8.
[9] Wing YK. Herbal treatment of insomnia. Hong Kong Medical Journal2001;7:392–402.
[10] Duman RS, Malberg J, Nakagawa S, D’Sa C. Neuronal plasticity andsurvival in mood disorders. Biological Psychiatry 2000;48:732–9.
[11] Nibuya M, Nestler EJ, Duman RS. Chronic antidepressant administra-tion increases the expression of cAMP response element binding protein(CREB) in rat hippocampus. Journal of Neuroscience 1996;16:2365–72.
[12] Castrén E. Neurotrophic effects of antidepressant drugs. Current Opinionin Pharmacology 2004;4:58–64.
[13] Castrén E, Rantamäki T. The role of BDNF and its receptors in depressionand antidepressant drug action: reactivation of developmental plasticity.Developmental Neurobiology 2010;70:289–97.
[14] Grimes CA, Jope RS. The multifaceted roles of glycogen synthase kinase3� in cellular signaling. Progress in Neurobiology 2001;65:391–426.
[15] Nakagawara A, Azar CG, Scavarda NJ, Brodeur GM. Expression andfunction of TRK-B and BDNF in human neuroblastomas. Molecular andCellular Biology 1994;14:759–67.
[16] Li Z, Tan F, Thiele CJ. Inactivation of glycogen synthase kinase-3�
contributes to brain-derived neutrophic factor/TrkB-induced resistance tochemotherapy in neuroblastoma cells. Molecular Cancer Therapeutics2007;6:3113–21.
[17] Gould TD, Zarate CA, Manji HK. Glycogen synthase kinase-3: a tar-get for novel bipolar disorder treatments. Journal of Clinical Psychiatry2004;65:10–21.
[18] Jope RS, Roh MS. Glycogen synthase kinase-3 (GSK3) in psychiatric dis-eases and therapeutic interventions. Current Drug Targets 2006;7:1421–34.
[19] Gould TD, Einat H, Bhat R, Manji HK. AR-A014418, a selective GSK-3 inhibitor, produces antidepressant-like effects in the forced swim test.International Journal of Neuropsychopharmacology 2004;7:387–90.
[20] Kaidanovich-Beilin O, Milman A, Weizman A, Pick CG, Eldar-FinkelmanH. Rapid antidepressive-like activity of specific glycogen synthase kinase-3 inhibitor and its effect on �-catenin in mouse hippocampus. BiologicalPsychiatry 2004;55:781–4.
[21] Rosa AO, Kaster MP, Binfaré RW, Morales S, Martín-Aparicio E, Navarro-Rico ML, et al. Antidepressant-like effect of the novel thiadiazolidinoneNP031115 in mice. Progress in Neuro-Psychopharmacology and Biologi-cal Psychiatry 2008;32:1549–56.
[22] Borsini F, Bendotti C, Velkov V, Rech R, Samanin R. Immobility test:effects of 5-hydroxytryptaminergic drugs and role of catecholamines in theactivity of some antidepressants. Journal of Pharmacy and Pharmacology1981;33:33–7.
[23] Porsolt RD, LePichon M, Jalfre M. Depression: new animal model sensitiveto antidepressant treatments. Nature 1977;266:730–2.
[24] Porsolt RD, Anton G, Blavet N, Jalfre M. Behavioral despair in rats:a new model sensitive to antidepressant treatment. European Journal ofPharmacology 1978;47:379–91.
[25] Mizoguchi K, Yuzurihara M, Ishige A, Aburada M, Tabira T. Saiko-ka-ryukotsu-borei-to, a herbal medicine, ameliorates chronic stress-induceddepressive state in rotarod performance. Pharmacology Biochemistry andBehavior 2003;75:419–25.
[26] Gao X, Zheng X, Li Z, Zhou Y, Sun H, Zhang L, et al. Metabonomic studyon chronic unpredictable mild stress and intervention effects of Xiaoyaosanin rats using gas chromatography coupled with mass spectrometry. Journalof Ethnopharmacology 2011;137:690–9.
[27] Zhou Y, Lu L, Li Z, Gao X, Tian J, Zhang L, et al. Antidepressant-like effectsof the fractions of Xiaoyaosan on rat model of chronic unpredictable mildstress. Journal of Ethnopharmacology 2011;137:236–44.
[28] Nemeroff CB, Owens MJ. Treatment of mood disorders. Nature Neuro-science 2002;5(suppl.1):1068–70.
[29] D’Sa C, Duman RS. Antidepressants and neuroplasticity. Bipolar Disorders2002;4:183–94.
[30] Duman RS, Nakagawa S, Malberg J. Regulation of adult neurogenesis byantidepressant treatment. Neuropsychopharmacology 2001;25:836–44.
[31] O’Leary OF, Wu X, Castren E. Chronic fluoxetine treatment increasesexpression of synaptic proteins in the hippocampus of the ovariectomizedrat: role of BDNF signalling. Psychoneuroendocrinology 2009;34:367–81.
[32] Gould TD, Einat H, O’Donnell KC, Picchini AM, Schloesser RJ, Manji HK.�-Catenin overexpression in the mouse brain phenocopies lithium-sensitivebehaviors. Neuropsychopharmacology 2007;32:2173–83.
[33] Roh MS, Kang UG, Shin SY, Lee YH, Jung HY, Juhnn YS, et al.Biphasic changes in the Ser-9 phosphorylation of glycogen synthasekinase-3� after electroconvulsive shock in the rat brain. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2003;27:1–5.
[34] Li X, Zhu W, Roh MS, Friedman AB, Rosborough K, Jope RS. In vivo reg-ulation of glycogen synthase kinase-3� (GSK-3�) by serotonergic activityin mouse brain. Neuropsychopharmacology 2004;29:1426–31.
[35] Yu X, Malenka RC. �-Catenin is critical for dendritic morphogenesis.Nature Neuroscience 2003;6:1169–77.
[36] Logan CY, Nusse R. The Wnt signaling pathway in developmentand disease. Annual Review of Cell and Developmental Biology2004;20:781–810.
[37] Jope RS, Bijur GN. Mood stabilizers, glycogen synthase kinase-3� andcell survival. Molecular Psychiatry 2002;7(suppl.1):S35–45.
[38] Beaulieu JM, Marion S, Rodriguiz RM, Medvedev IO, Sotnikova TD,Ghisi V, et al. A �-arrestin 2 signaling complex mediates lithium action onbehavior. Cell 2008;132:125–36.
[39] Beaulieu JM, Sotnikova TD, Marion S, Lefkowitz RJ, GainetdinovRR, Caron MG. An Akt/�-arrestin 2/PP2A signaling complex mediatesdopaminergic neurotransmission and behavior. Cell 2005;122:261–73.
[40] Hsu YL, Kuo PL, Lin CC. The proliferative inhibition and apoptotic mech-anism of Saikosaponin D in human non-small cell lung cancer A549 cells.Life Sciences 2004;75:1231–42.
[41] Hsu YL, Kuo PL, Weng TC, Yen MH, Chiang LC, Lin CC. The antiprolifer-ative activity of saponin-enriched fraction from Bupleurum Kaoi is throughFas-dependent apoptotic pathway in human non-small cell lung cancerA549 cells. Biological and Pharmaceutical Bulletin 2004;27:1112–5.
[42] Biedler JL, Roffler-Tarlov S, Schachner M, Freedman LS. Multiple neuro-transmitter synthesis by human neuroblastoma cell lines and clones. CancerResearch 1978;38:3751–7.
[43] Farooqui SM. Induction of adenylate cyclase sensitive dopamine D2-receptors in retinoic acid induced differentiated human neuroblastomaSHSY-5Y cells. Life Sciences 1994;55:1887–93.
[44] Yaniv SP, Lucki A, Klein E, Ben-Shachar D. Dexamethasone enhances thenorepinephrine-induced ERK/MAPK intracellular pathway possibly viadysregulation of the alpha2-adrenergic receptor: implications for antide-pressant drug mechanism of action. European Journal of Cell Biology2010;89:712–22.