atpy glicólisis (bioenergética) - fabián rodríguez
TRANSCRIPT

Universidad Central de Venezuela
Facultad de Medicina
Escuela de Medicina José María Vargas
Caracas, Agosto 2012
ATP y Glicólisis
(Bioenergética)
Fabián Rodríguez
Médico Cirujano - UCV
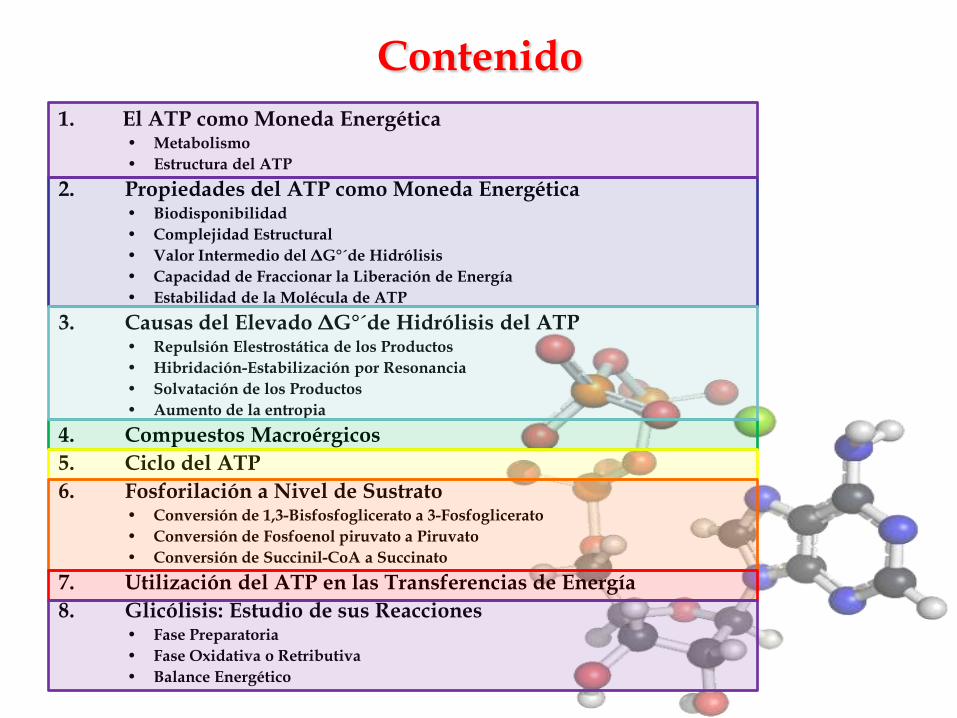
Contenido
1. El ATP como Moneda Energética• Metabolismo
• Estructura del ATP
2. Propiedades del ATP como Moneda Energética• Biodisponibilidad
• Complejidad Estructural
• Valor Intermedio del ΔG°´de Hidrólisis
• Capacidad de Fraccionar la Liberación de Energía
• Estabilidad de la Molécula de ATP
3. Causas del Elevado ΔG°´de Hidrólisis del ATP• Repulsión Elestrostática de los Productos
• Hibridación-Estabilización por Resonancia
• Solvatación de los Productos
• Aumento de la entropia
4. Compuestos Macroérgicos
5. Ciclo del ATP
6. Fosforilación a Nivel de Sustrato• Conversión de 1,3-Bisfosfoglicerato a 3-Fosfoglicerato
• Conversión de Fosfoenol piruvato a Piruvato
• Conversión de Succinil-CoA a Succinato
7. Utilización del ATP en las Transferencias de Energía
8. Glicólisis: Estudio de sus Reacciones• Fase Preparatoria
• Fase Oxidativa o Retributiva
• Balance Energético

El ATP COMO MONEDA ENERGÉTICA
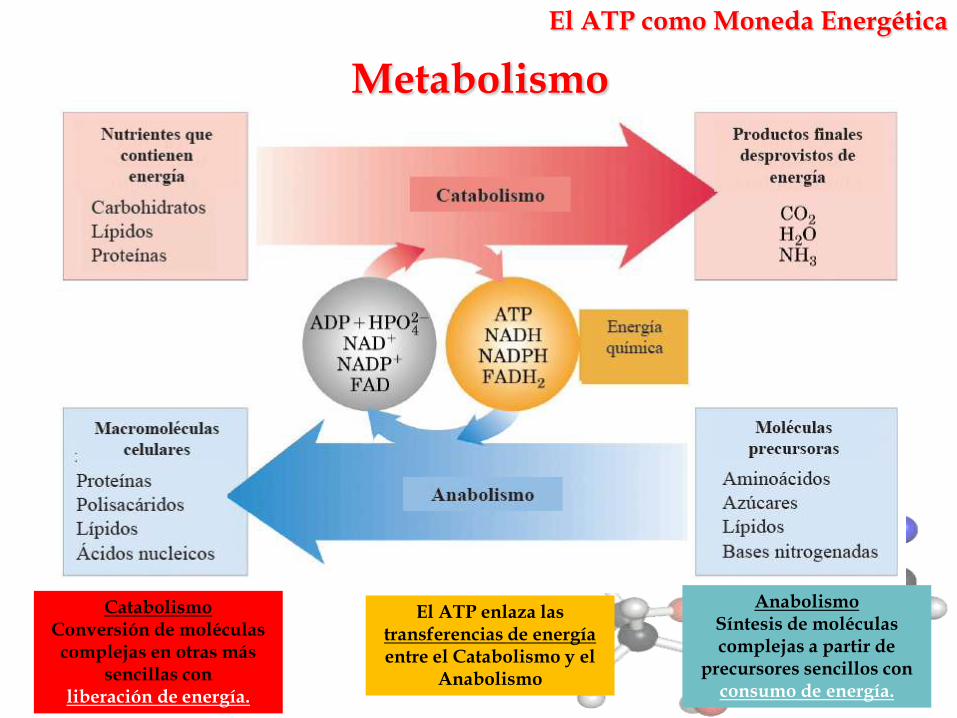
El ATP como Moneda Energética
CatabolismoConversión de moléculas complejas en otras más
sencillas con liberación de energía.
Metabolismo
AnabolismoSíntesis de moléculas complejas a partir de
precursores sencillos conconsumo de energía.
El ATP enlaza las transferencias de energía entre el Catabolismo y el
Anabolismo
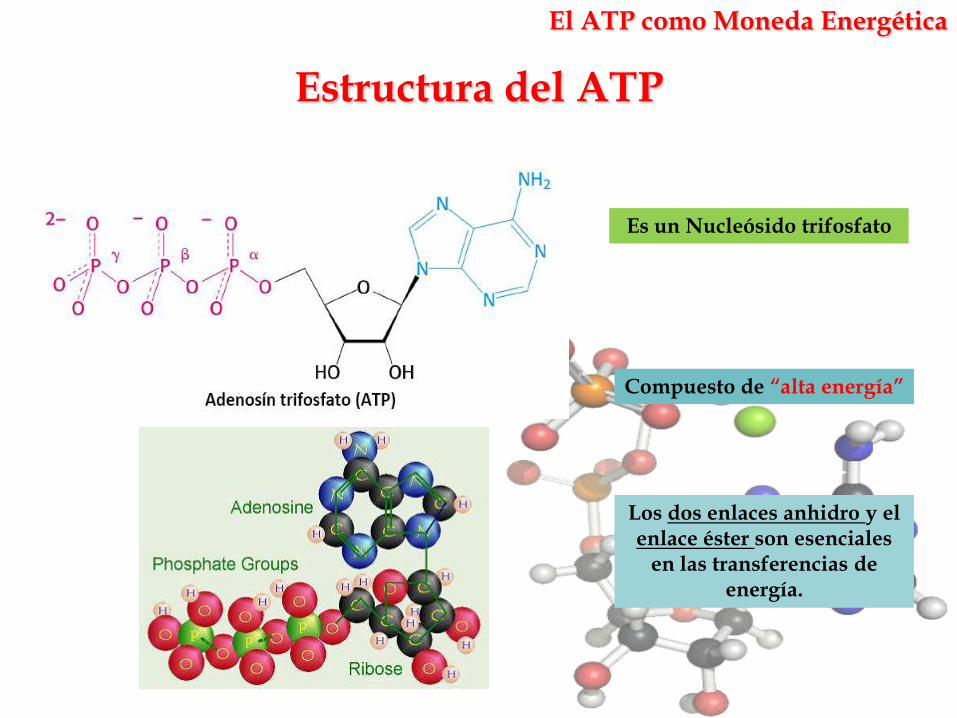
Es un Nucleósido trifosfato
Compuesto de “alta energía”
Los dos enlaces anhidro y el enlace éster son esenciales
en las transferencias de energía.
El ATP como Moneda Energética
Estructura del ATP

PROPIEDADES DEL ATP COMO MONEDA ENERGÉTICA

Propiedades como Moneda Energética
1. Biodisponibilidad.
2. Complejidad Estructural.
3. Valor Intermedio del ΔG°´ de Hidrólisis.
4. Capacidad de Fraccionar la liberación de Energía.
5. Estabilidad de la Molécula de ATP
Propiedades del ATP
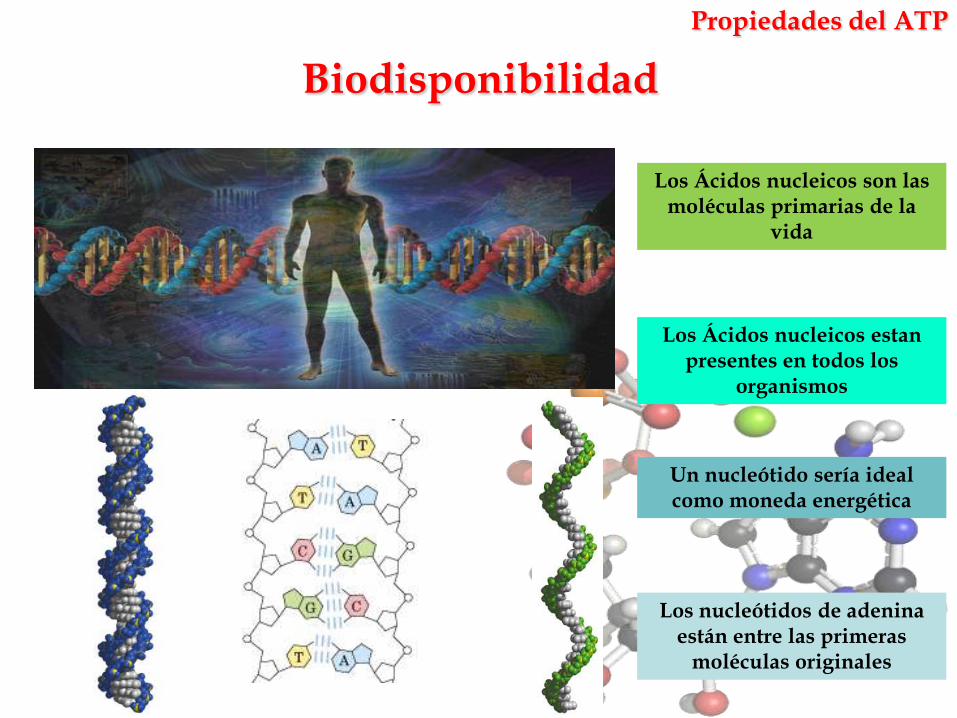
Biodisponibilidad
Propiedades del ATP
Los Ácidos nucleicos son las moléculas primarias de la
vida
Un nucleótido sería ideal como moneda energética
Los Ácidos nucleicos estanpresentes en todos los
organismos
Los nucleótidos de adeninaestán entre las primeras
moléculas originales
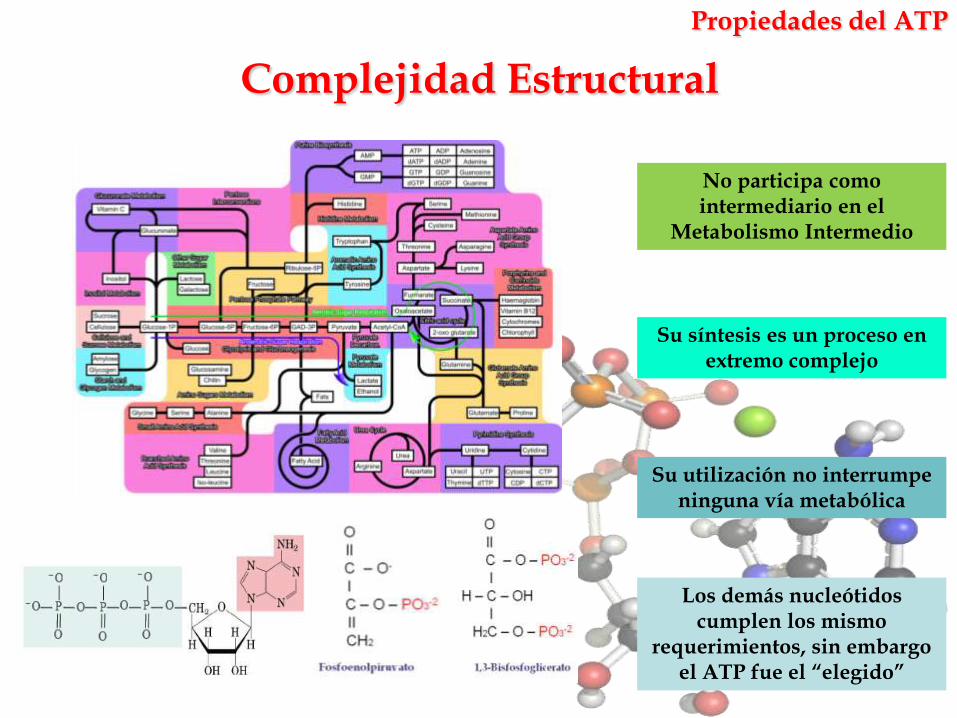
Complejidad Estructural
Propiedades del ATP
No participa como intermediario en el
Metabolismo Intermedio
Su utilización no interrumpe ninguna vía metabólica
Su síntesis es un proceso en extremo complejo
Los demás nucleótidos cumplen los mismo
requerimientos, sin embargo el ATP fue el “elegido”
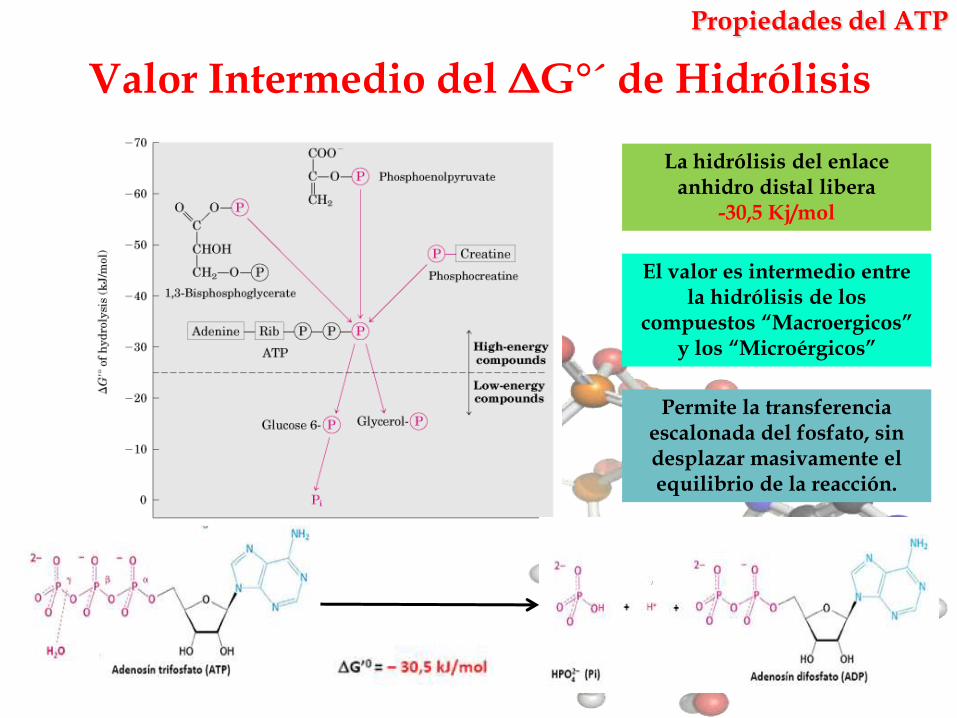
Valor Intermedio del ΔG°´ de Hidrólisis
Propiedades del ATP
La hidrólisis del enlace anhidro distal libera
-30,5 Kj/mol
Permite la transferencia escalonada del fosfato, sin desplazar masivamente el equilibrio de la reacción.
El valor es intermedio entre la hidrólisis de los
compuestos “Macroergicos” y los “Microérgicos”
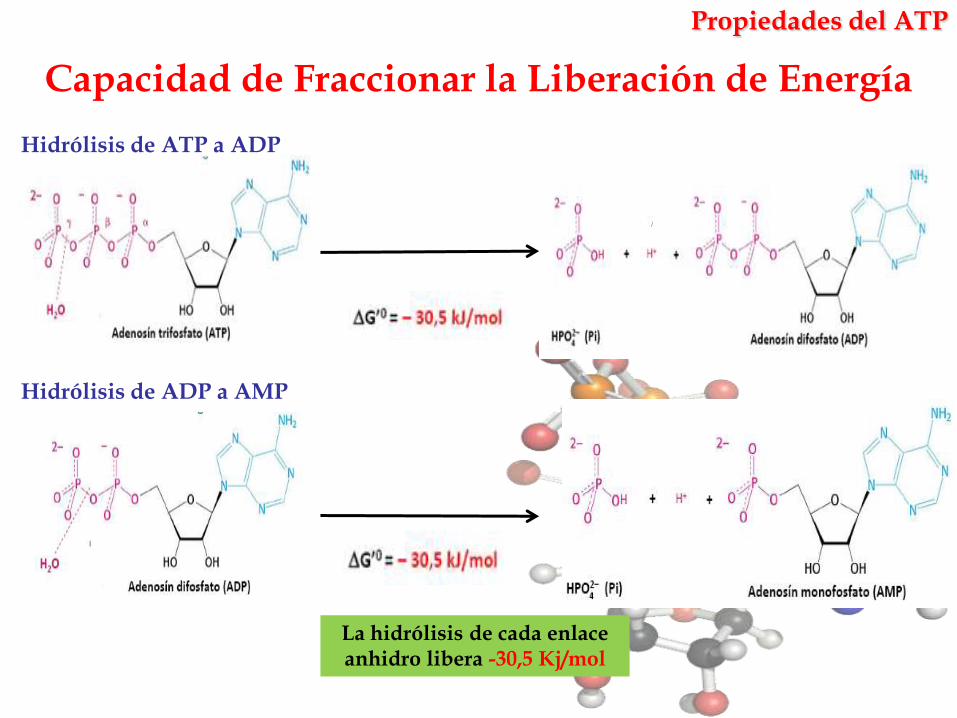
Capacidad de Fraccionar la Liberación de Energía
Propiedades del ATP
La hidrólisis de cada enlace anhidro libera -30,5 Kj/mol
Hidrólisis de ATP a ADP
Hidrólisis de ADP a AMP
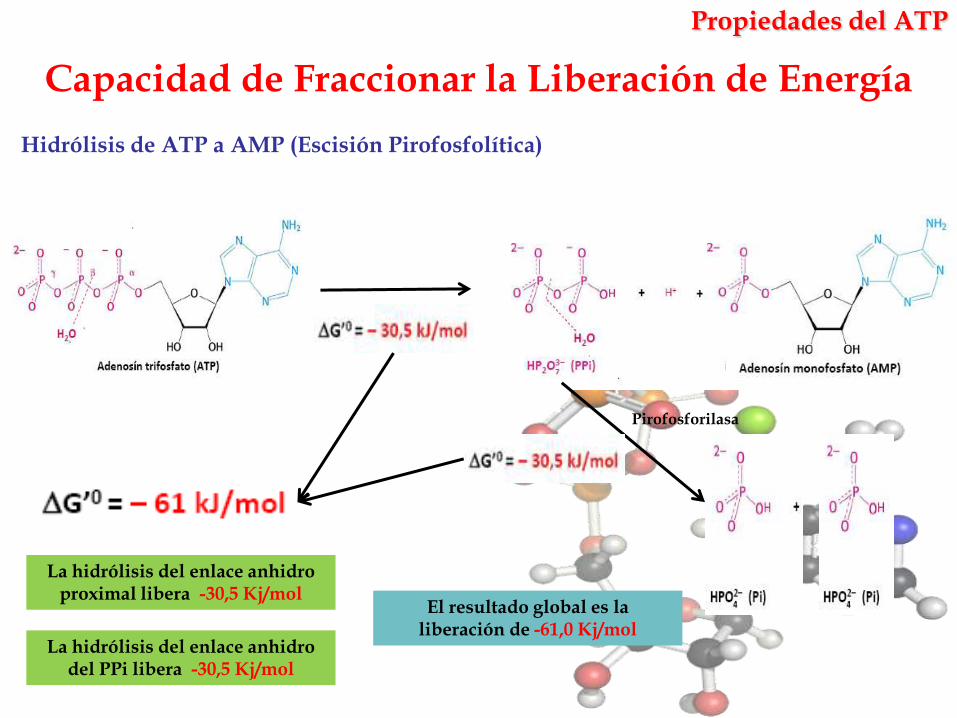
Capacidad de Fraccionar la Liberación de Energía
Propiedades del ATP
La hidrólisis del enlace anhidro proximal libera -30,5 Kj/mol
Hidrólisis de ATP a AMP (Escisión Pirofosfolítica)
Pirofosforilasa
La hidrólisis del enlace anhidro del PPi libera -30,5 Kj/mol
El resultado global es la liberación de -61,0 Kj/mol
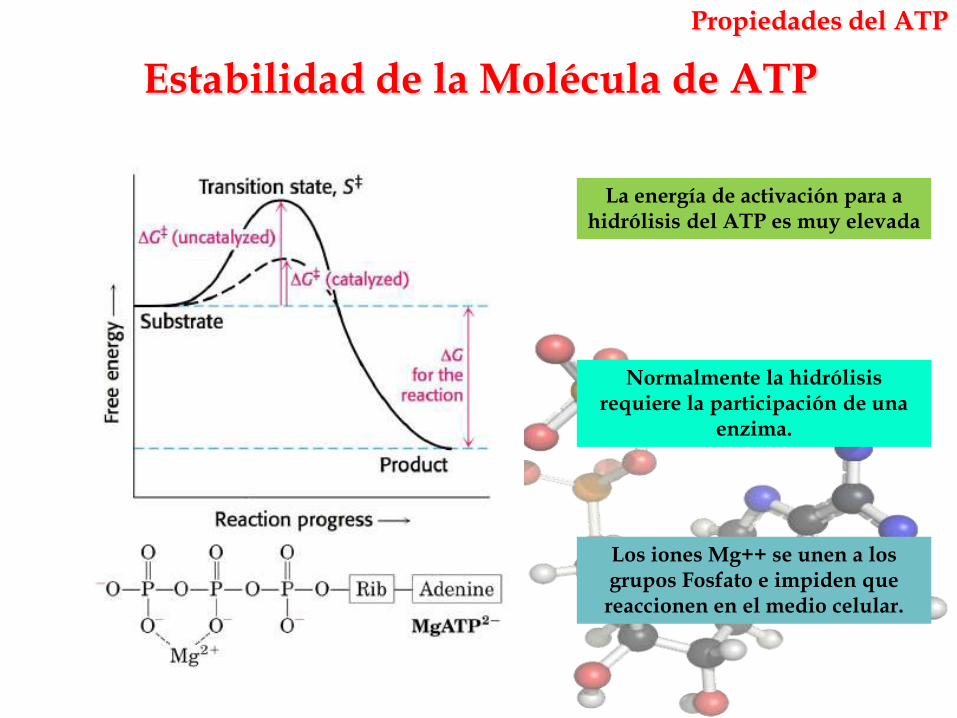
Estabilidad de la Molécula de ATP
Propiedades del ATP
La energía de activación para a hidrólisis del ATP es muy elevada
Los iones Mg++ se unen a los grupos Fosfato e impiden que reaccionen en el medio celular.
Normalmente la hidrólisis requiere la participación de una
enzima.

CAUSAS DEL ELEVADO ΔG°´ DE HIDRÓLISIS DEL ATP
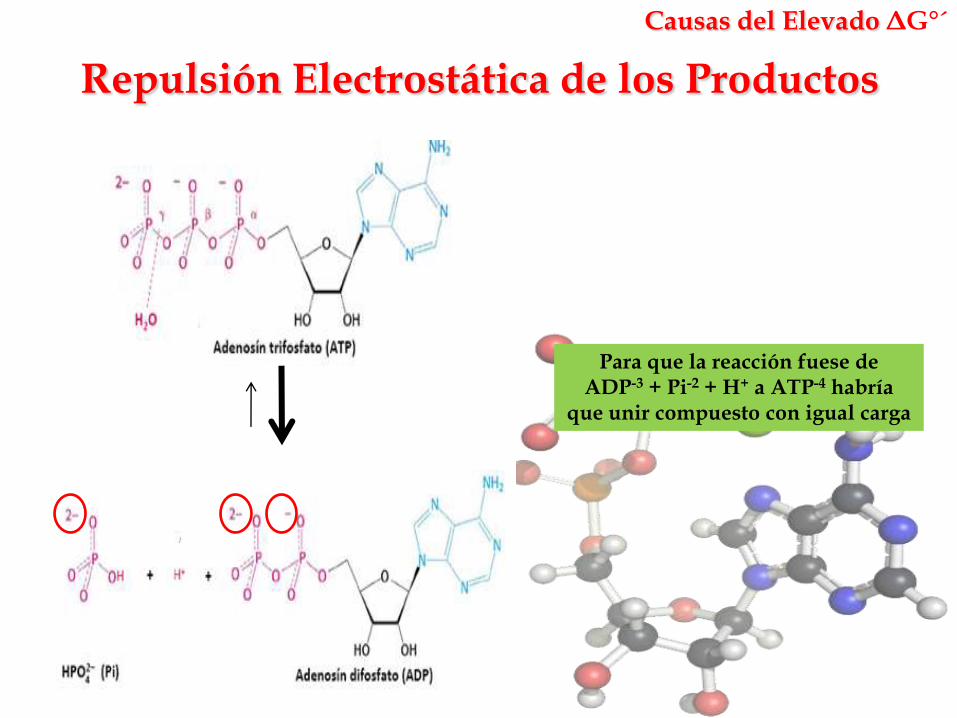
Repulsión Electrostática de los Productos
Causas del Elevado ΔG°´
Para que la reacción fuese de ADP-3 + Pi-2 + H+ a ATP-4 habría
que unir compuesto con igual carga
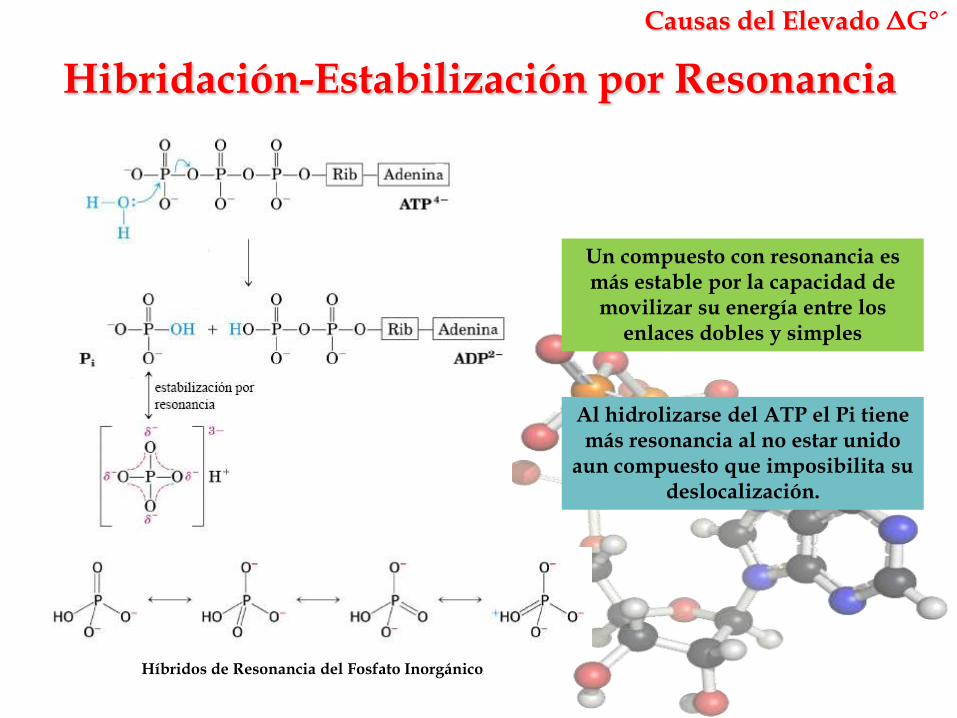
Hibridación-Estabilización por Resonancia
Causas del Elevado ΔG°´
Un compuesto con resonancia es más estable por la capacidad de movilizar su energía entre los
enlaces dobles y simples
Híbridos de Resonancia del Fosfato Inorgánico
Al hidrolizarse del ATP el Pi tiene más resonancia al no estar unido
aun compuesto que imposibilita su deslocalización.
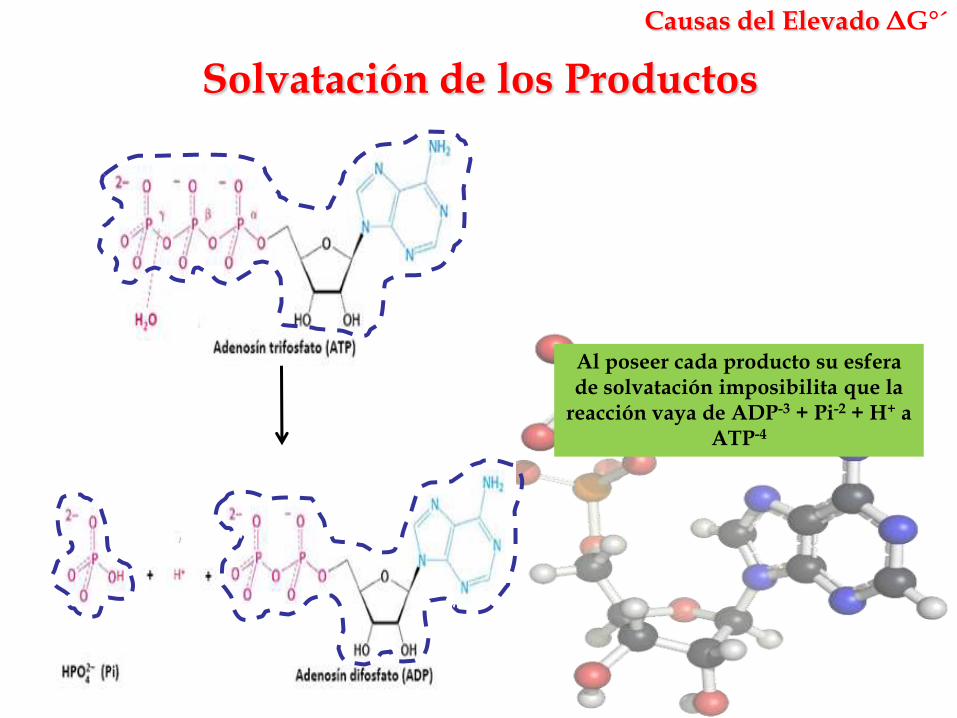
Solvatación de los Productos
Causas del Elevado ΔG°´
Al poseer cada producto su esfera de solvatación imposibilita que la
reacción vaya de ADP-3 + Pi-2 + H+ a ATP-4
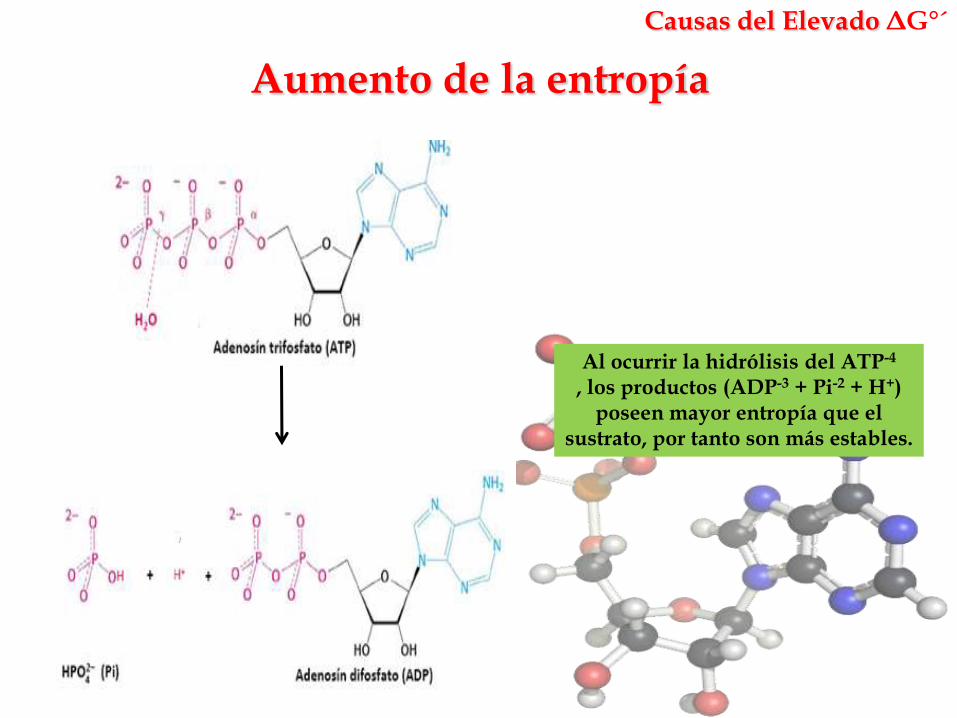
Aumento de la entropía
Causas del Elevado ΔG°´
Al ocurrir la hidrólisis del ATP-4
, los productos (ADP-3 + Pi-2 + H+) poseen mayor entropía que el
sustrato, por tanto son más estables.

COMPUESTOS MACROÉRGICOS
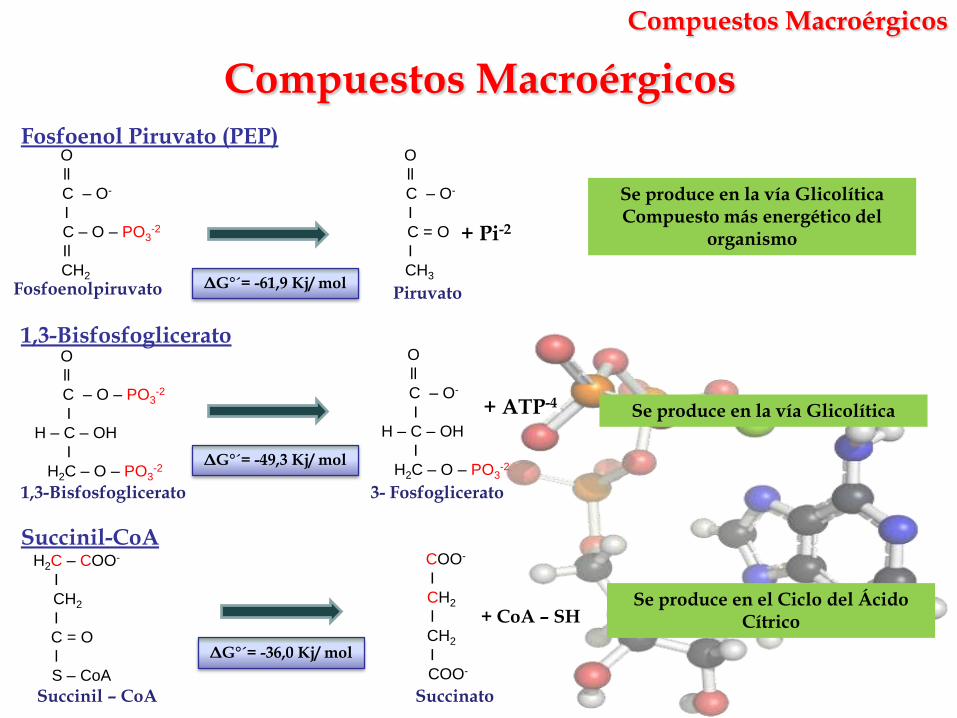
Compuestos Macroérgicos
Compuestos Macroérgicos
Se produce en la vía GlicolíticaCompuesto más energético del
organismo
O
ll
C – O-
I
C – O – PO3-2
Il
CH2
Fosfoenolpiruvato
O
ll
C – O-
I
C = O
I
CH3
Piruvato
+ Pi-2
O
ll
C – O-
I
H – C – OH
I
H2C – O – PO3-2
+ ATP-4
1,3-Bisfosfoglicerato
O
ll
C – O – PO3-2
I
H – C – OH
I
H2C – O – PO3-2
3- Fosfoglicerato
SuccinatoSuccinil – CoA
+ CoA – SH
COO-
I
CH2
l
CH2
I
COO-
H2C – COO-
I
CH2
I
C = O
l
S – CoA
ΔG°´= -61,9 Kj/ mol
ΔG°´= -49,3 Kj/ mol
ΔG°´= -36,0 Kj/ mol
Fosfoenol Piruvato (PEP)
1,3-Bisfosfoglicerato
Succinil-CoA
Se produce en la vía Glicolítica
Se produce en el Ciclo del Ácido Cítrico
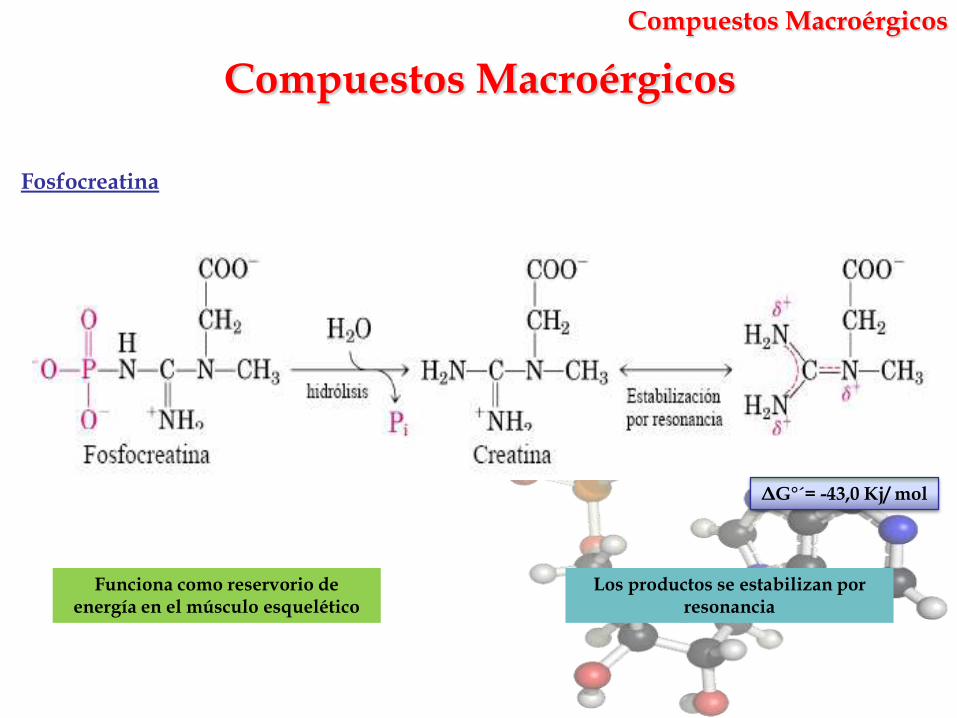
Compuestos Macroérgicos
Compuestos Macroérgicos
Los productos se estabilizan por resonancia
Fosfocreatina
Funciona como reservorio de energía en el músculo esquelético
ΔG°´= -43,0 Kj/ mol
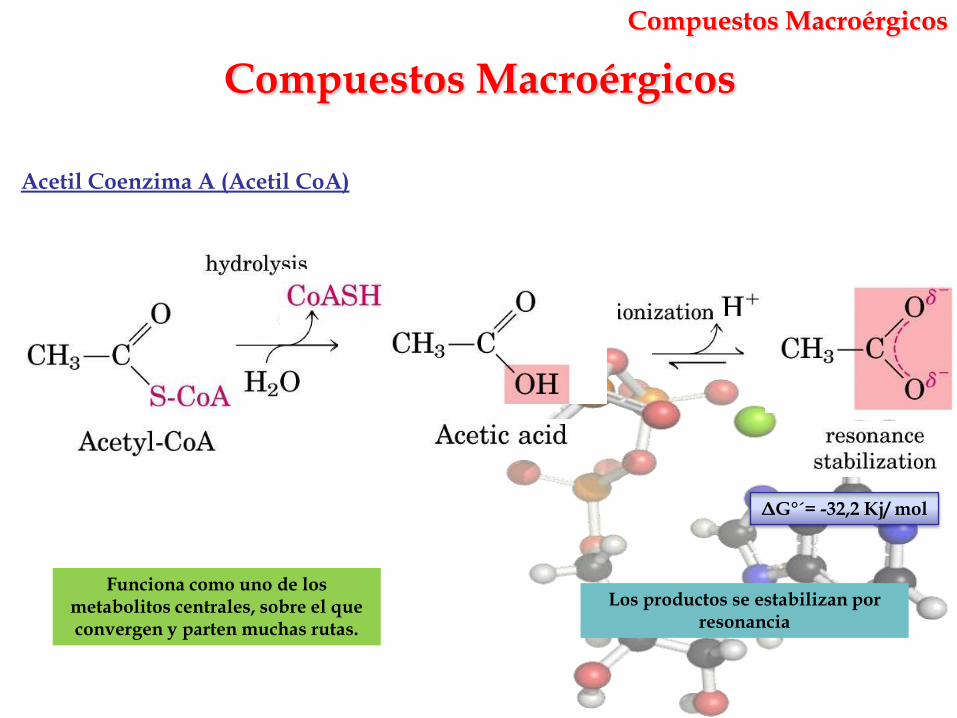
Compuestos Macroérgicos
Compuestos Macroérgicos
Los productos se estabilizan por resonancia
Acetil Coenzima A (Acetil CoA)
Funciona como uno de los metabolitos centrales, sobre el que convergen y parten muchas rutas.
ΔG°´= -32,2 Kj/ mol

CICLO DEL ATP
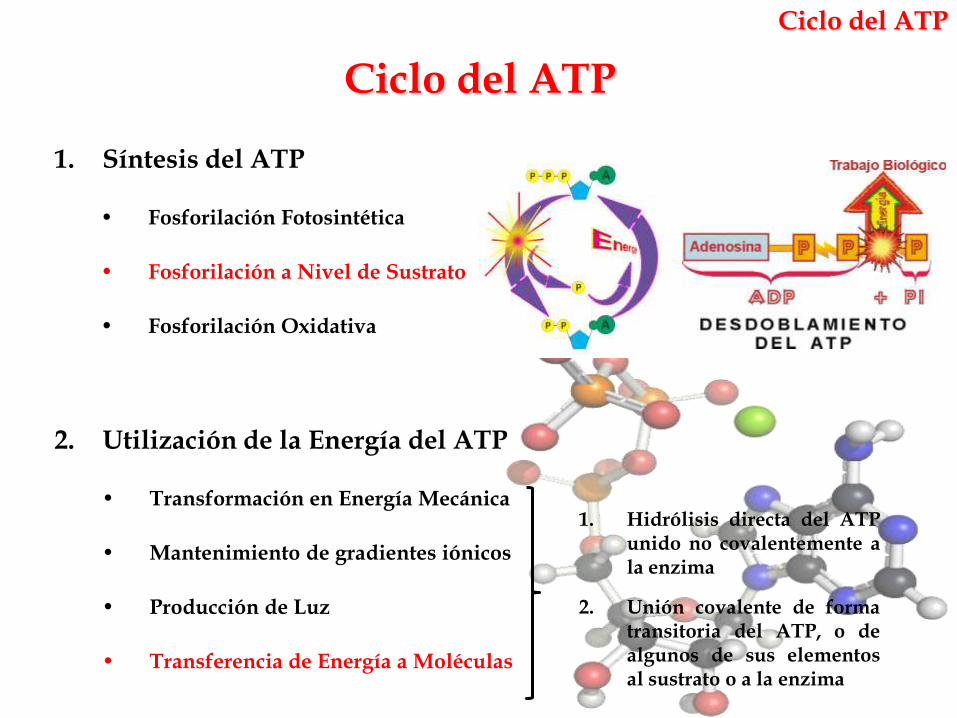
Ciclo del ATP
1. Síntesis del ATP
• Fosforilación Fotosintética
• Fosforilación a Nivel de Sustrato
• Fosforilación Oxidativa
2. Utilización de la Energía del ATP
• Transformación en Energía Mecánica
• Mantenimiento de gradientes iónicos
• Producción de Luz
• Transferencia de Energía a Moléculas
Ciclo del ATP
1. Hidrólisis directa del ATPunido no covalentemente ala enzima
2. Unión covalente de formatransitoria del ATP, o dealgunos de sus elementosal sustrato o a la enzima

FOSFORILACIÓN A NIVEL DE SUSTRATO
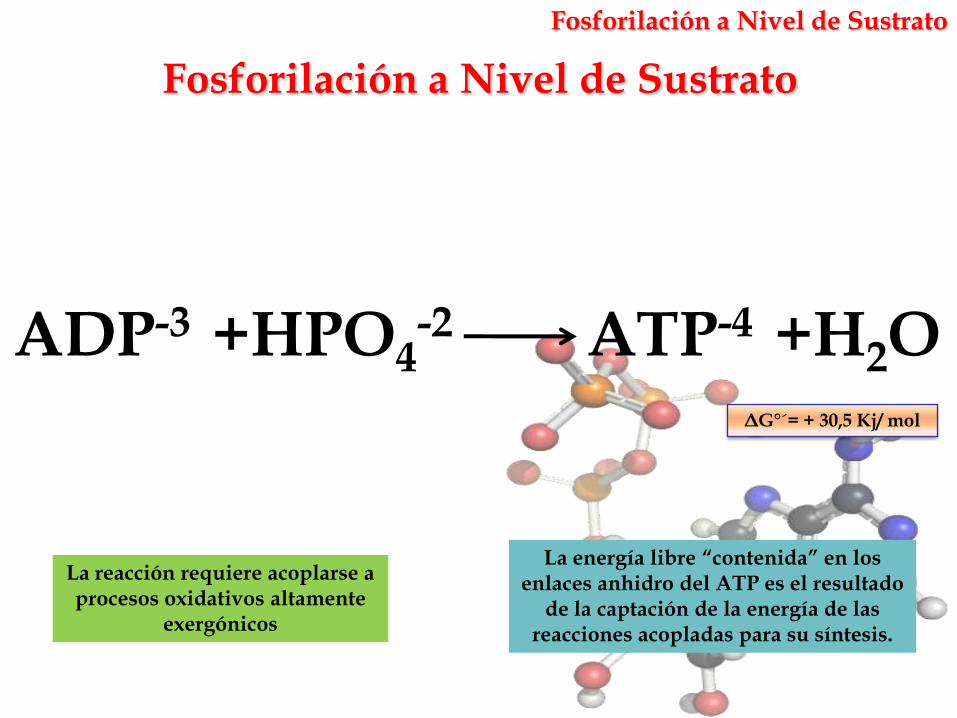
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
La reacción requiere acoplarse a procesos oxidativos altamente
exergónicos
ATP-4 +H2OADP-3 +HPO4-2
ΔG°´= + 30,5 Kj/ mol
La energía libre “contenida” en los enlaces anhidro del ATP es el resultado
de la captación de la energía de las reacciones acopladas para su síntesis.
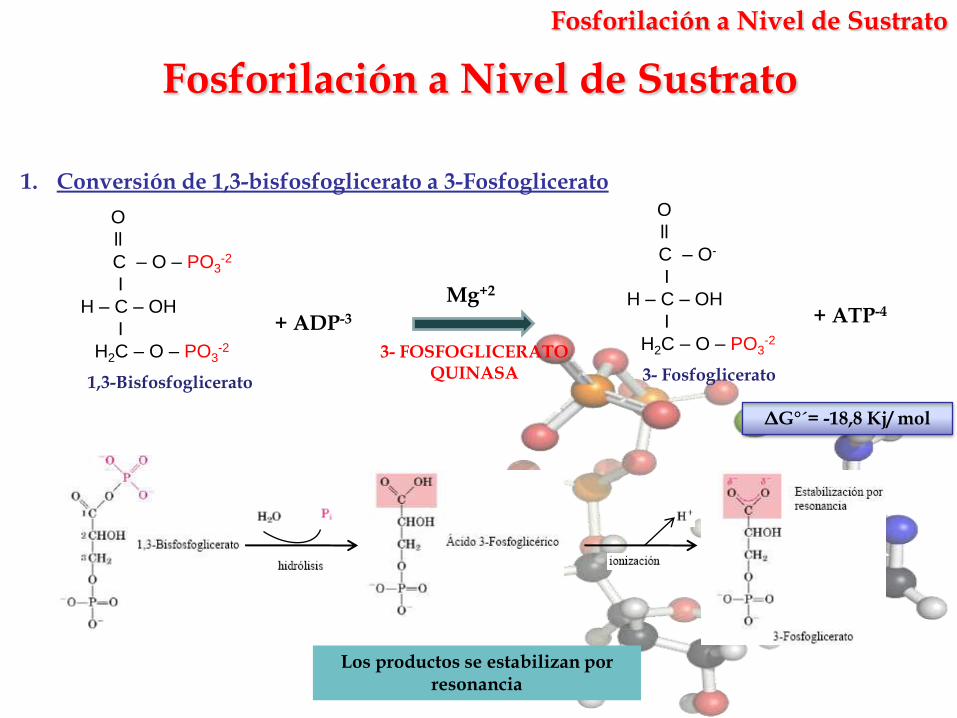
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
Los productos se estabilizan por resonancia
1. Conversión de 1,3-bisfosfoglicerato a 3-Fosfoglicerato
ΔG°´= -18,8 Kj/ mol
O
ll
C – O-
I
H – C – OH
I
H2C – O – PO3-2
+ ADP-3 + ATP-4
1,3-Bisfosfoglicerato
O
ll
C – O – PO3-2
I
H – C – OH
I
H2C – O – PO3-2
3- Fosfoglicerato
Mg+2
3- FOSFOGLICERATO QUINASA
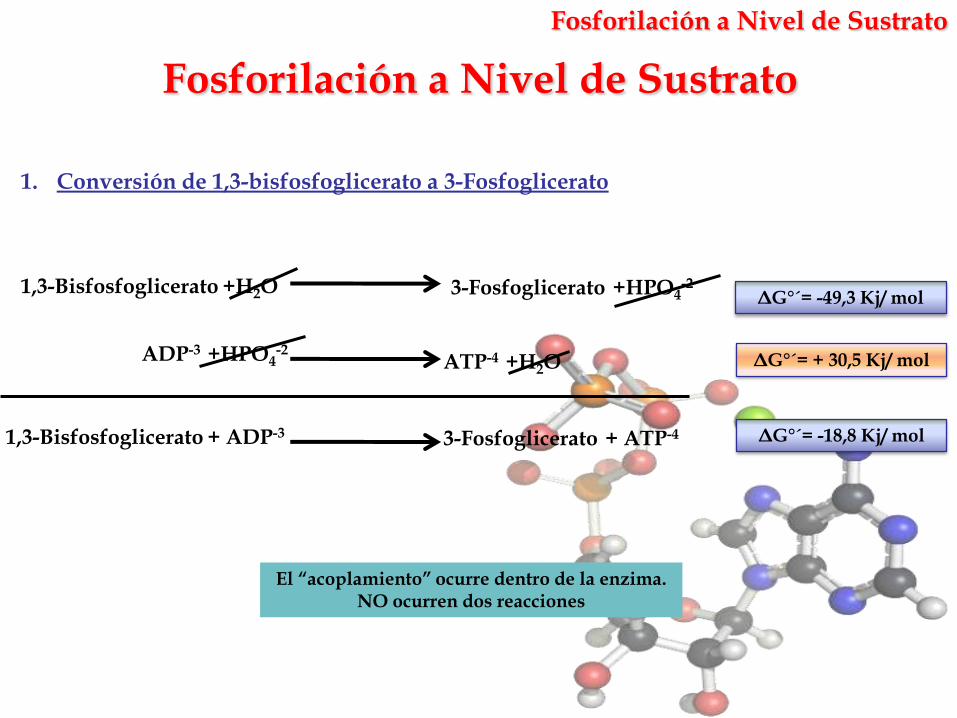
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
El “acoplamiento” ocurre dentro de la enzima. NO ocurren dos reacciones
1. Conversión de 1,3-bisfosfoglicerato a 3-Fosfoglicerato
ΔG°´= -18,8 Kj/ mol
ATP-4 +H2OADP-3 +HPO4
-2
1,3-Bisfosfoglicerato +H2O 3-Fosfoglicerato +HPO4-2
1,3-Bisfosfoglicerato + ADP-3 3-Fosfoglicerato + ATP-4
ΔG°´= + 30,5 Kj/ mol
ΔG°´= -49,3 Kj/ mol
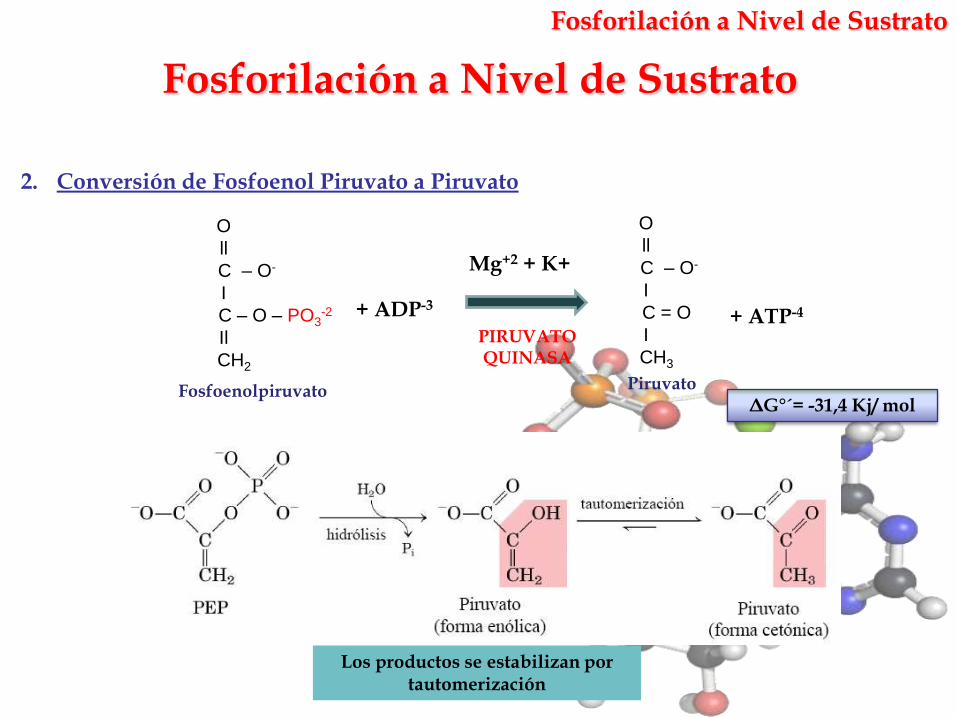
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
Los productos se estabilizan por tautomerización
2. Conversión de Fosfoenol Piruvato a Piruvato
Mg+2 + K+
PIRUVATO QUINASA
O
ll
C – O-
I
C – O – PO3-2
Il
CH2
Fosfoenolpiruvato
+ ADP-3
O
ll
C – O-
I
C = O
I
CH3
Piruvato
+ ATP-4
ΔG°´= -31,4 Kj/ mol
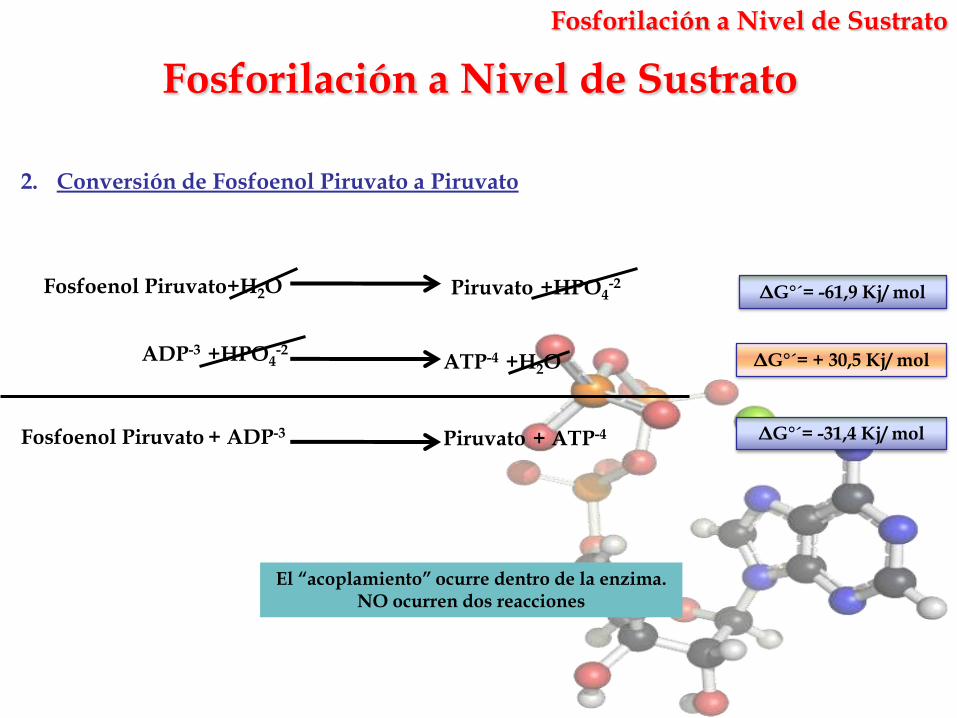
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
El “acoplamiento” ocurre dentro de la enzima. NO ocurren dos reacciones
2. Conversión de Fosfoenol Piruvato a Piruvato
ATP-4 +H2OADP-3 +HPO4
-2
Fosfoenol Piruvato+H2O Piruvato +HPO4-2
Fosfoenol Piruvato + ADP-3 Piruvato + ATP-4
ΔG°´= + 30,5 Kj/ mol
ΔG°´= -31,4 Kj/ mol
ΔG°´= -61,9 Kj/ mol
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
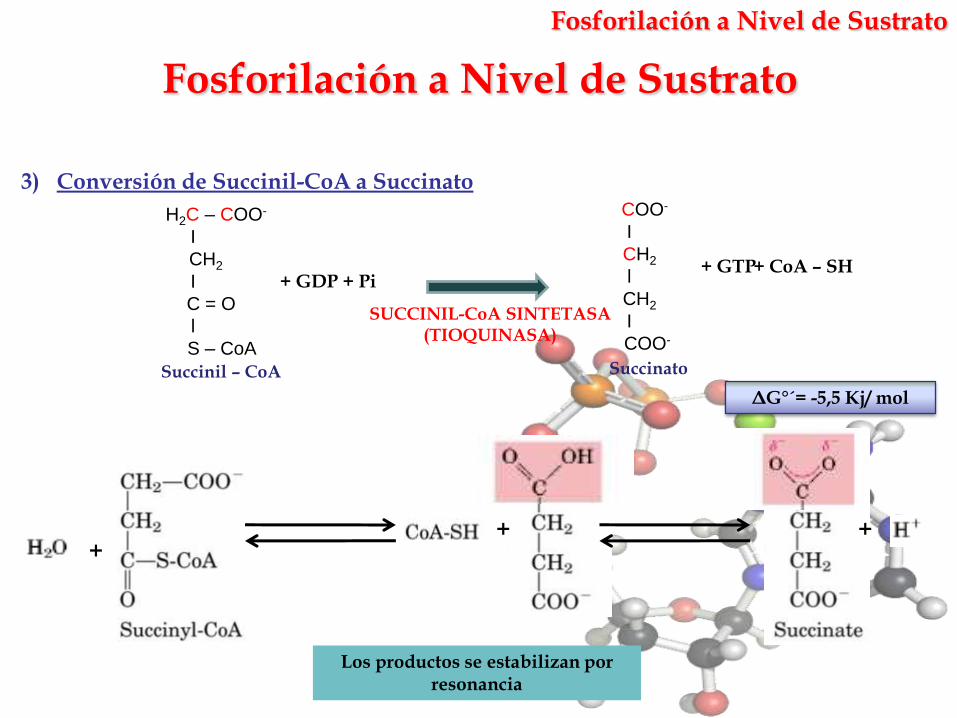
Los productos se estabilizan por resonancia
3) Conversión de Succinil-CoA a Succinato
SUCCINIL-CoA SINTETASA(TIOQUINASA)
Succinato
+ GDP + Pi
Succinil – CoA
+ GTP+ CoA – SH
COO-
I
CH2
l
CH2
I
COO-
H2C – COO-
I
CH2
I
C = O
l
S – CoA
ΔG°´= -5,5 Kj/ mol
++ +
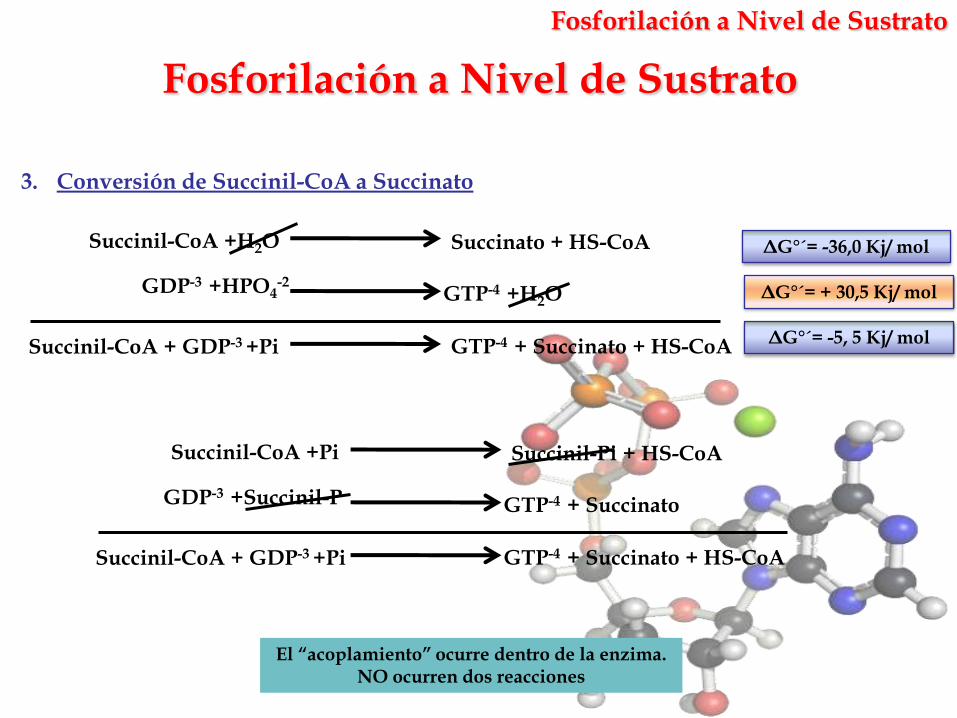
Fosforilación a Nivel de Sustrato
Fosforilación a Nivel de Sustrato
El “acoplamiento” ocurre dentro de la enzima. NO ocurren dos reacciones
3. Conversión de Succinil-CoA a Succinato
GTP-4 +H2OGDP-3 +HPO4
-2
Succinil-CoA +H2O Succinato + HS-CoA
ΔG°´= + 30,5 Kj/ mol
ΔG°´= -36,0 Kj/ mol
GTP-4 + SuccinatoGDP-3 +Succinil-P
Succinil-CoA +Pi Succinil-Pi + HS-CoA
Succinil-CoA + GDP-3 +Pi GTP-4 + Succinato + HS-CoA
Succinil-CoA + GDP-3 +Pi GTP-4 + Succinato + HS-CoA ΔG°´= -5, 5 Kj/ mol

MECANISMOS DE TRANSFERENCIA DE LA ENERGÍA DEL ATP
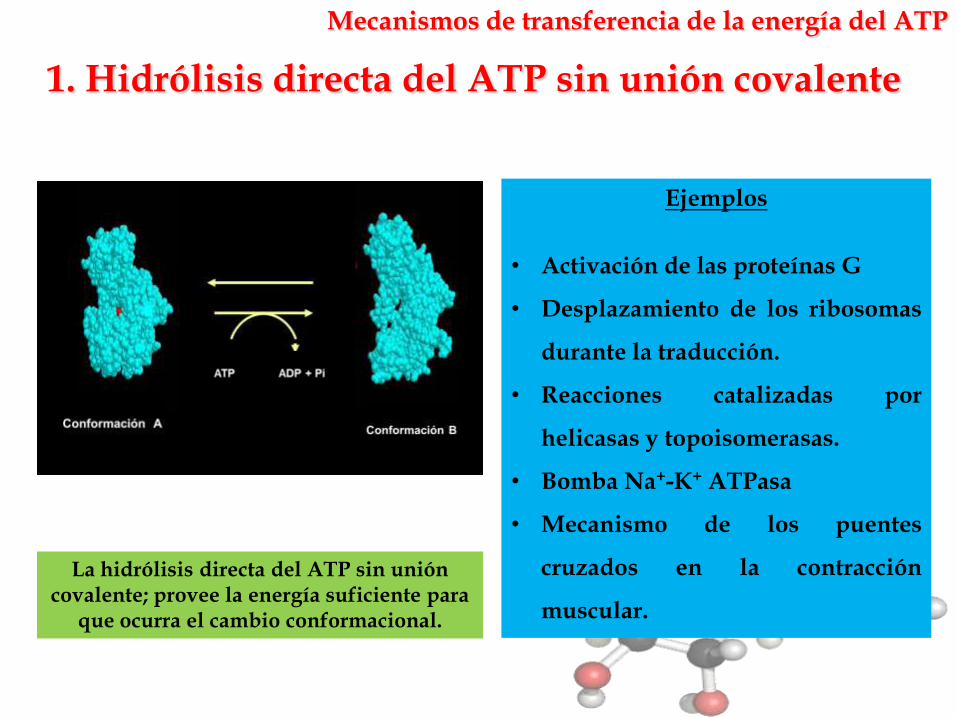
1. Hidrólisis directa del ATP sin unión covalente
Mecanismos de transferencia de la energía del ATP
La hidrólisis directa del ATP sin unión covalente; provee la energía suficiente para
que ocurra el cambio conformacional.
Ejemplos
• Activación de las proteínas G
• Desplazamiento de los ribosomas
durante la traducción.
• Reacciones catalizadas por
helicasas y topoisomerasas.
• Bomba Na+-K+ ATPasa
• Mecanismo de los puentes
cruzados en la contracción
muscular.
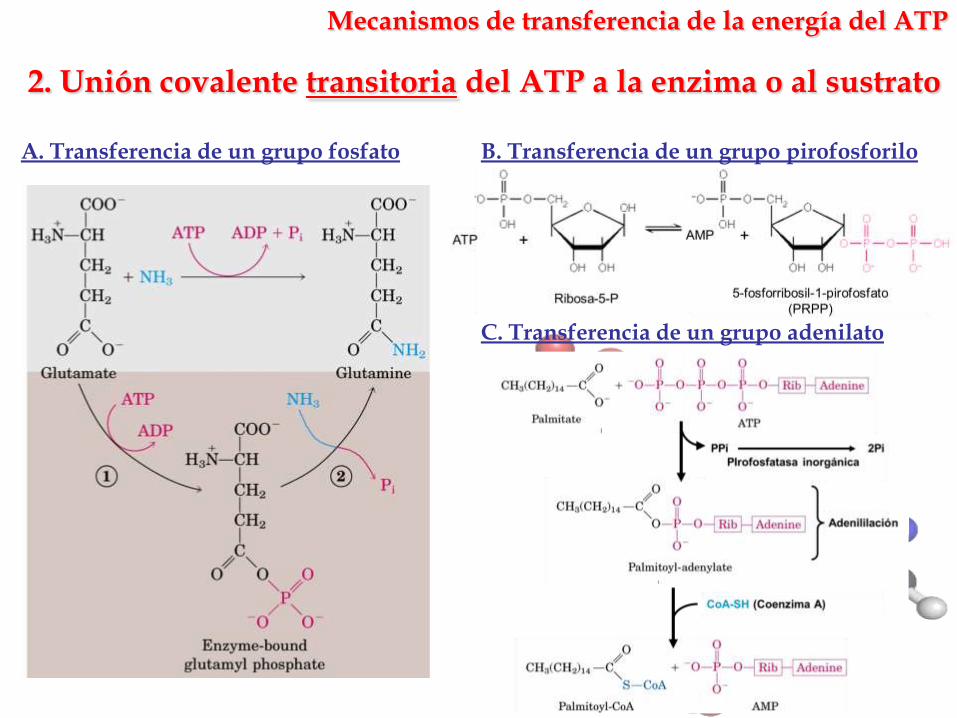
2. Unión covalente transitoria del ATP a la enzima o al sustrato
Mecanismos de transferencia de la energía del ATP
A. Transferencia de un grupo fosfato B. Transferencia de un grupo pirofosforilo
C. Transferencia de un grupo adenilato
UTILIZACIÓN DEL ATP EN LA TRANSFERENCIA DE ENERGÍA A
OTRAS MOLÉCULAS
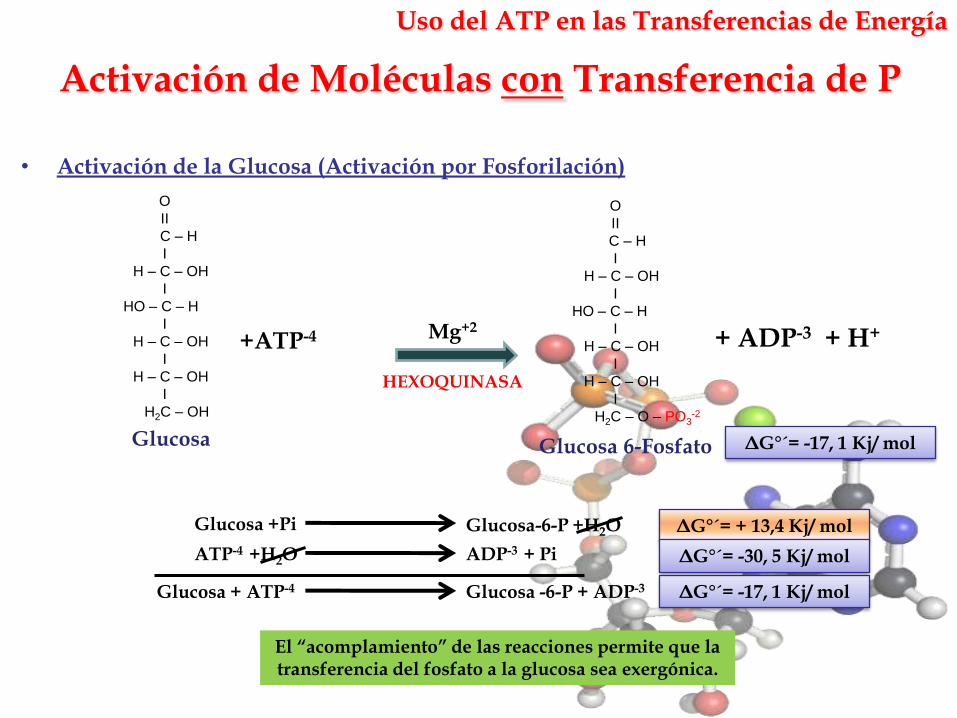
Activación de Moléculas con Transferencia de P
Uso del ATP en las Transferencias de Energía
El “acomplamiento” de las reacciones permite que la transferencia del fosfato a la glucosa sea exergónica.
• Activación de la Glucosa (Activación por Fosforilación)
HEXOQUINASA
O
II
C – H
I
H – C – OH
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – OH
O
II
C – H
I
H – C – OH
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
ΔG°´= -17, 1 Kj/ molGlucosa Glucosa 6-Fosfato
+ATP-4 + ADP-3 + H+Mg+2
ADP-3 + PiATP-4 +H2O
Glucosa +Pi Glucosa-6-P +H2O
Glucosa + ATP-4 Glucosa -6-P + ADP-3 ΔG°´= -17, 1 Kj/ mol
ΔG°´= + 13,4 Kj/ mol
ΔG°´= -30, 5 Kj/ mol
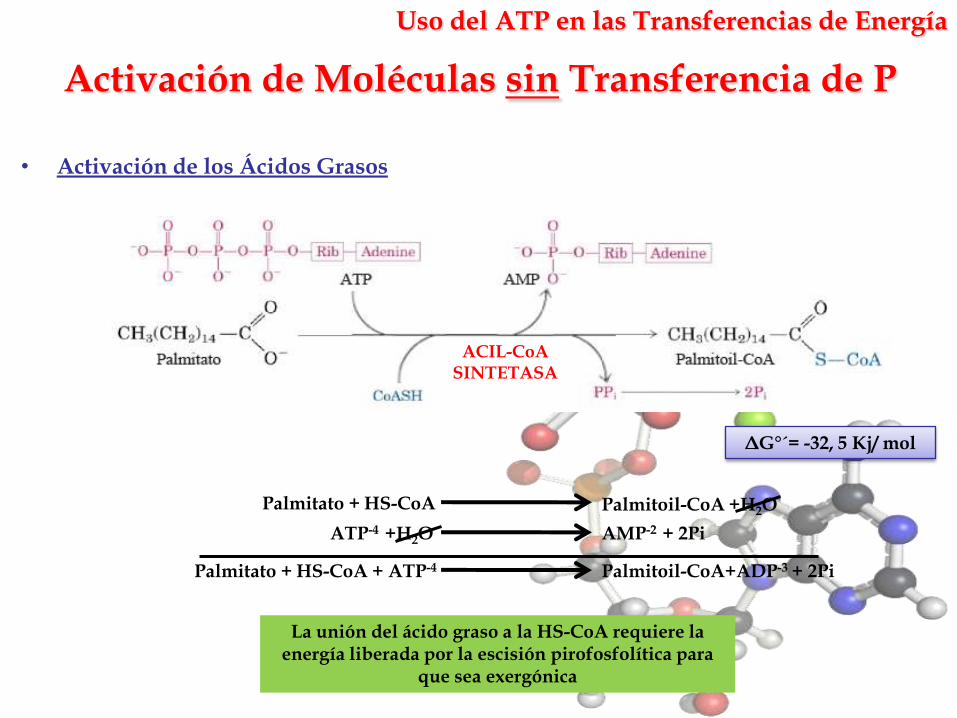
Activación de Moléculas sin Transferencia de P
Uso del ATP en las Transferencias de Energía
La unión del ácido graso a la HS-CoA requiere la energía liberada por la escisión pirofosfolítica para
que sea exergónica
• Activación de los Ácidos Grasos
ΔG°´= -32, 5 Kj/ mol
AMP-2 + 2PiATP-4 +H2O
Palmitato + HS-CoA Palmitoil-CoA +H2O
Palmitato + HS-CoA + ATP-4 Palmitoil-CoA+ADP-3 + 2Pi
ACIL-CoASINTETASA
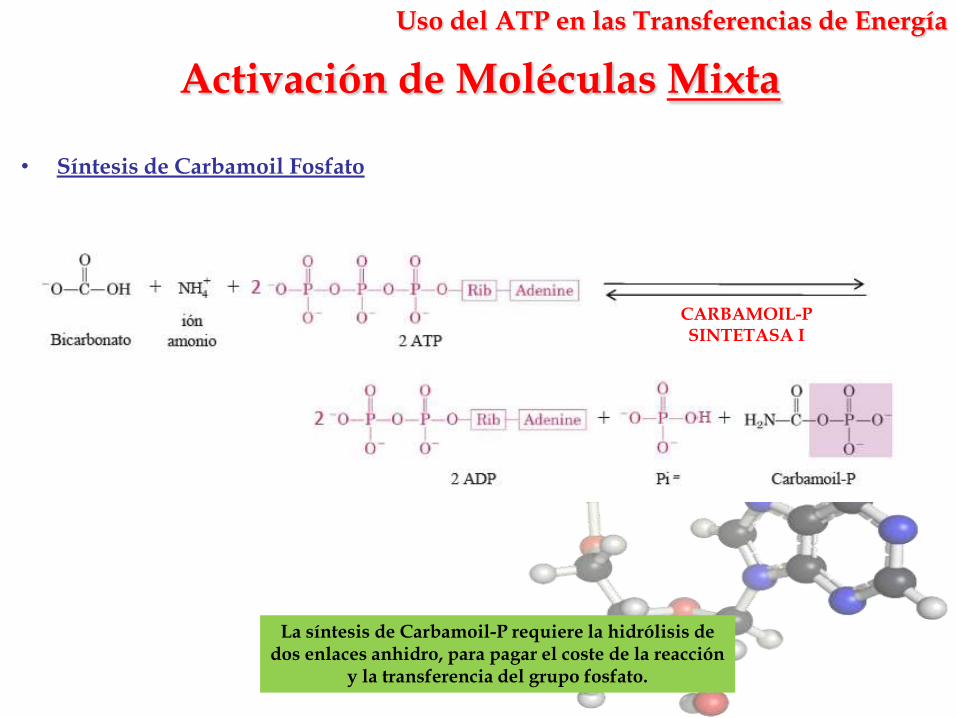
Activación de Moléculas Mixta
Uso del ATP en las Transferencias de Energía
La síntesis de Carbamoil-P requiere la hidrólisis de dos enlaces anhidro, para pagar el coste de la reacción
y la transferencia del grupo fosfato.
• Síntesis de Carbamoil Fosfato
CARBAMOIL-PSINTETASA I

GLICÓLISIS :
ESTUDIO DE SUS REACCIONES
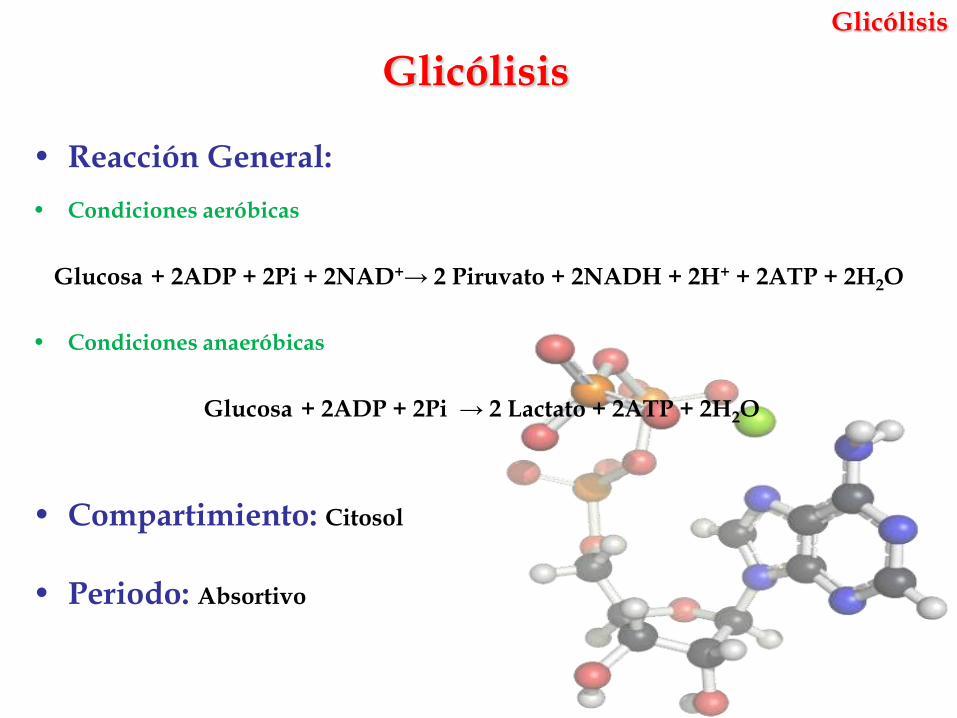
Glicólisis
• Reacción General:
• Condiciones aeróbicas
Glucosa + 2ADP + 2Pi + 2NAD+→ 2 Piruvato + 2NADH + 2H+ + 2ATP + 2H2O
• Condiciones anaeróbicas
Glucosa + 2ADP + 2Pi → 2 Lactato + 2ATP + 2H2O
• Compartimiento: Citosol
• Periodo: Absortivo
Glicólisis
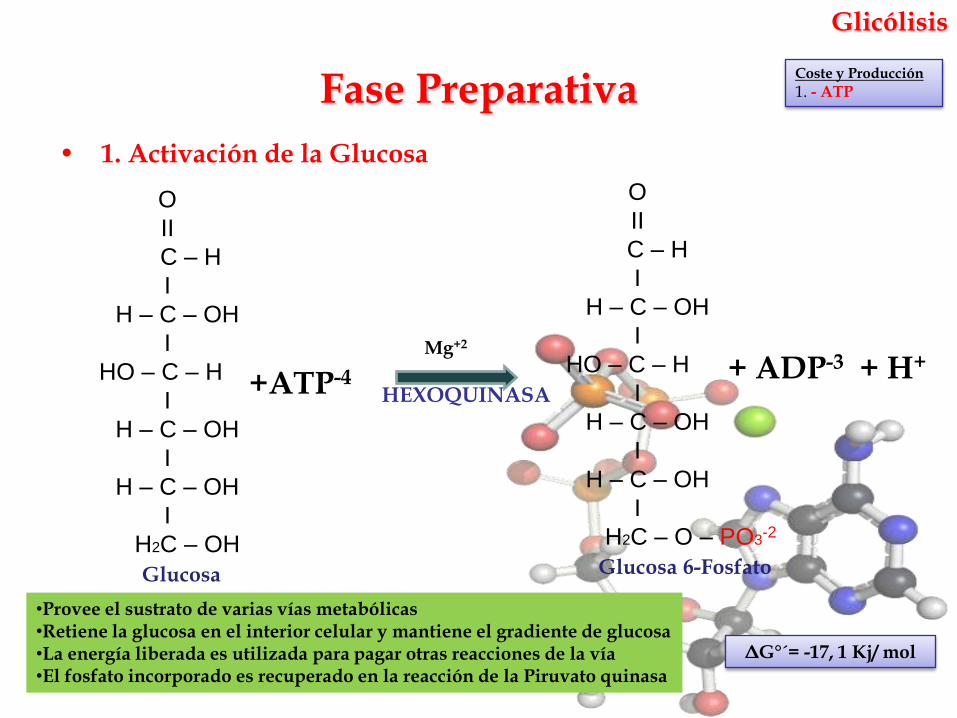
Glicólisis
• 1. Activación de la Glucosa
Mg+2
HEXOQUINASA
Fase Preparativa
O
II
C – H
I
H – C – OH
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – OH
O
II
C – H
I
H – C – OH
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
Coste y Producción
1. - ATP
ΔG°´= -17, 1 Kj/ mol
Glucosa Glucosa 6-Fosfato
+ATP-4 + ADP-3 + H+
•Provee el sustrato de varias vías metabólicas•Retiene la glucosa en el interior celular y mantiene el gradiente de glucosa•La energía liberada es utilizada para pagar otras reacciones de la vía•El fosfato incorporado es recuperado en la reacción de la Piruvato quinasa
Glicólisis
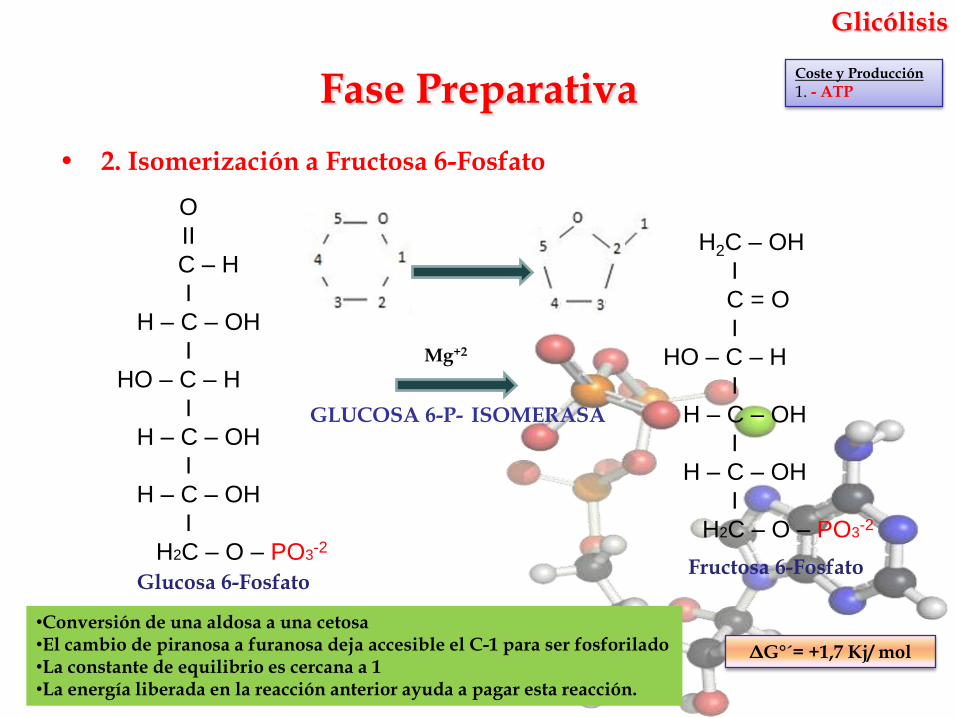
• 2. Isomerización a Fructosa 6-Fosfato
Mg+2
GLUCOSA 6-P- ISOMERASA
Fase Preparativa
O
II
C – H
I
H – C – OH
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
Coste y Producción
1. - ATP
ΔG°´= +1,7 Kj/ mol
Glucosa 6-FosfatoFructosa 6-Fosfato
H2C – OH
I
C = O
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
•Conversión de una aldosa a una cetosa•El cambio de piranosa a furanosa deja accesible el C-1 para ser fosforilado•La constante de equilibrio es cercana a 1•La energía liberada en la reacción anterior ayuda a pagar esta reacción.
Glicólisis
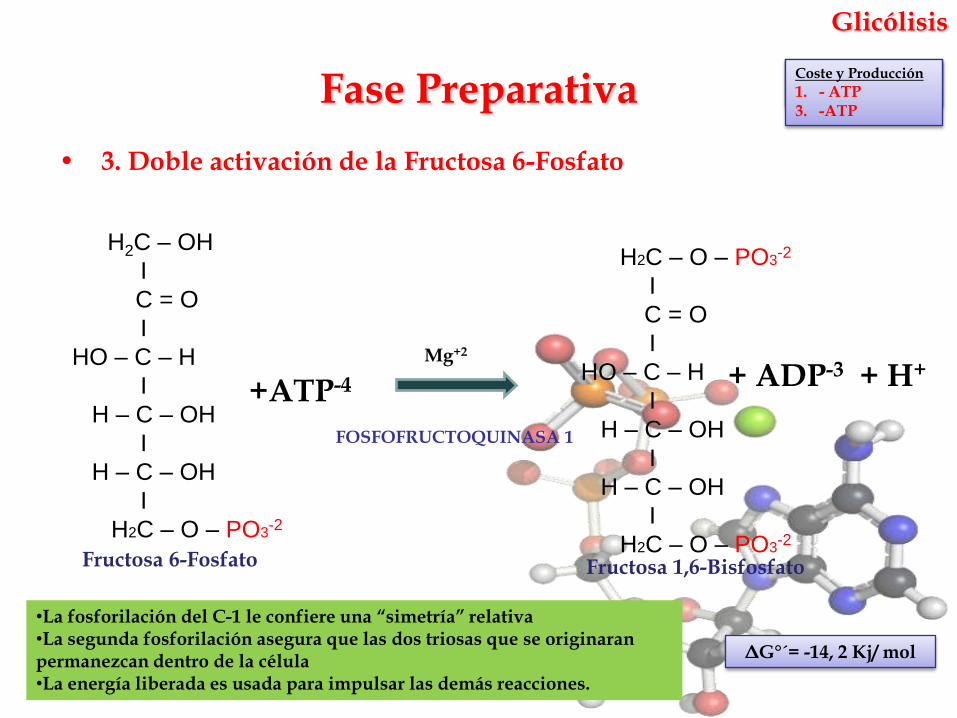
• 3. Doble activación de la Fructosa 6-Fosfato
Mg+2
FOSFOFRUCTOQUINASA 1
Fase PreparativaCoste y Producción
1. - ATP
ΔG°´= -14, 2 Kj/ mol
Fructosa 6-Fosfato Fructosa 1,6-Bisfosfato
+ATP-4 + ADP-3 + H+
H2C – OH
I
C = O
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
H2C – O – PO3-2
I
C = O
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
Coste y Producción
1. - ATP3. -ATP
•La fosforilación del C-1 le confiere una “simetría” relativa•La segunda fosforilación asegura que las dos triosas que se originaran permanezcan dentro de la célula•La energía liberada es usada para impulsar las demás reacciones.
Glicólisis
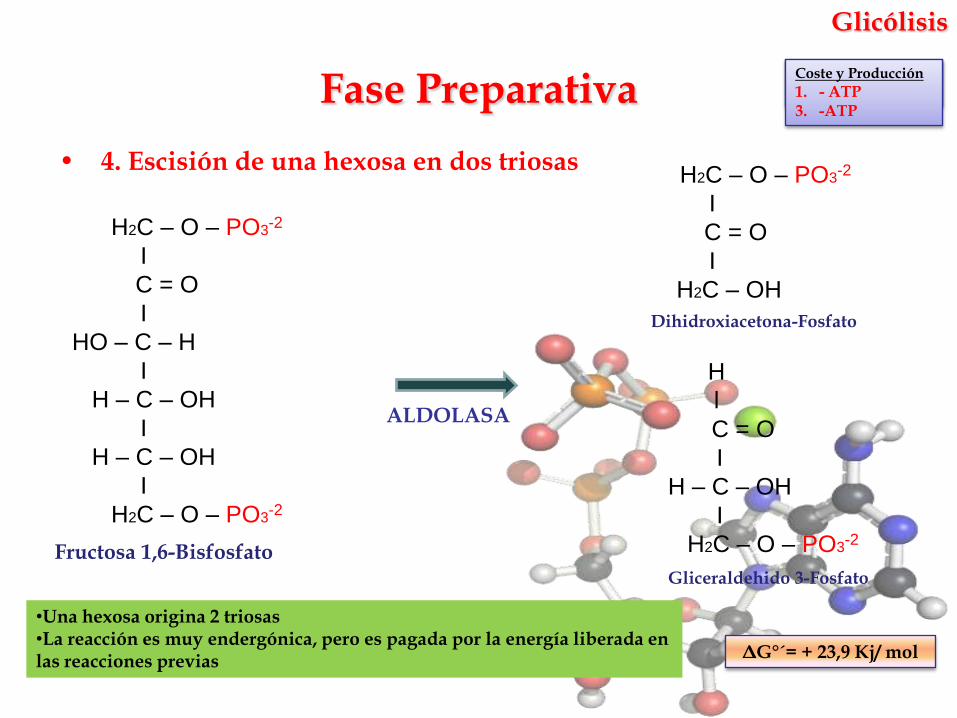
• 4. Escisión de una hexosa en dos triosas
ALDOLASA
Fase PreparativaCoste y Producción
1. - ATP
ΔG°´= + 23,9 Kj/ mol
Fructosa 1,6-Bisfosfato
Dihidroxiacetona-Fosfato
H2C – O – PO3-2
I
C = O
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – O – PO3-2
H2C – O – PO3-2
I
C = O
I
H2C – OH
H
l
C = O
I
H – C – OH
I
H2C – O – PO3-2
Gliceraldehido 3-Fosfato
Coste y Producción
1. - ATP3. -ATP
•Una hexosa origina 2 triosas•La reacción es muy endergónica, pero es pagada por la energía liberada en las reacciones previas
Glicólisis
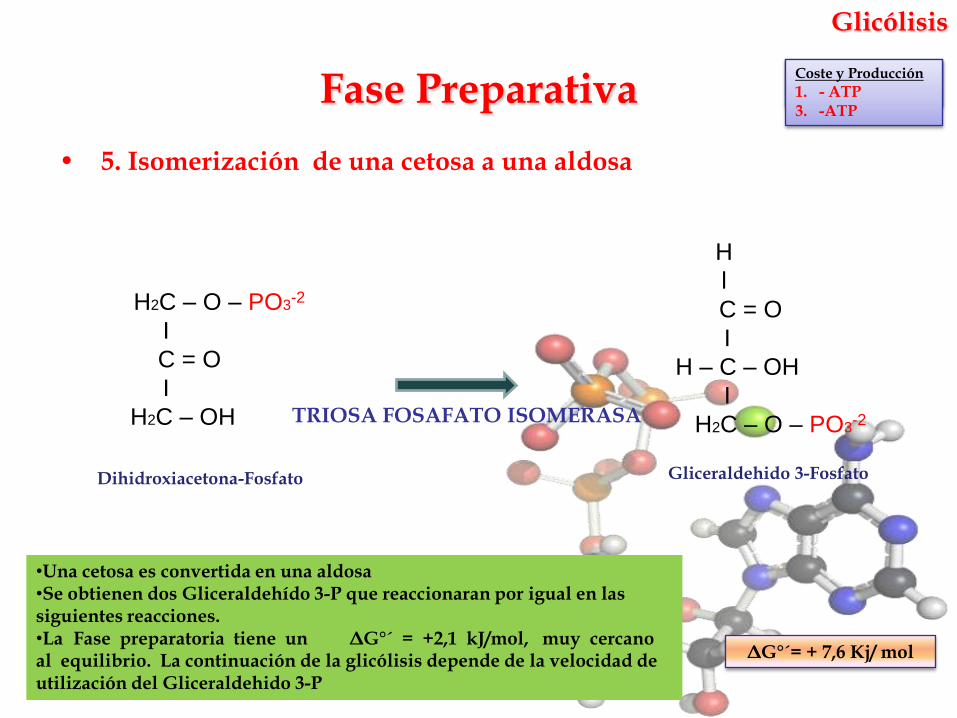
• 5. Isomerización de una cetosa a una aldosa
TRIOSA FOSAFATO ISOMERASA
Fase PreparativaCoste y Producción
1. - ATP
ΔG°´= + 7,6 Kj/ mol
Dihidroxiacetona-Fosfato
H2C – O – PO3-2
I
C = O
I
H2C – OH
H
l
C = O
I
H – C – OH
I
H2C – O – PO3-2
Gliceraldehido 3-Fosfato
Coste y Producción
1. - ATP3. -ATP
•Una cetosa es convertida en una aldosa•Se obtienen dos Gliceraldehído 3-P que reaccionaran por igual en las siguientes reacciones.•La Fase preparatoria tiene un ΔG°´ = +2,1 kJ/mol, muy cercano al equilibrio. La continuación de la glicólisis depende de la velocidad de utilización del Gliceraldehido 3-P
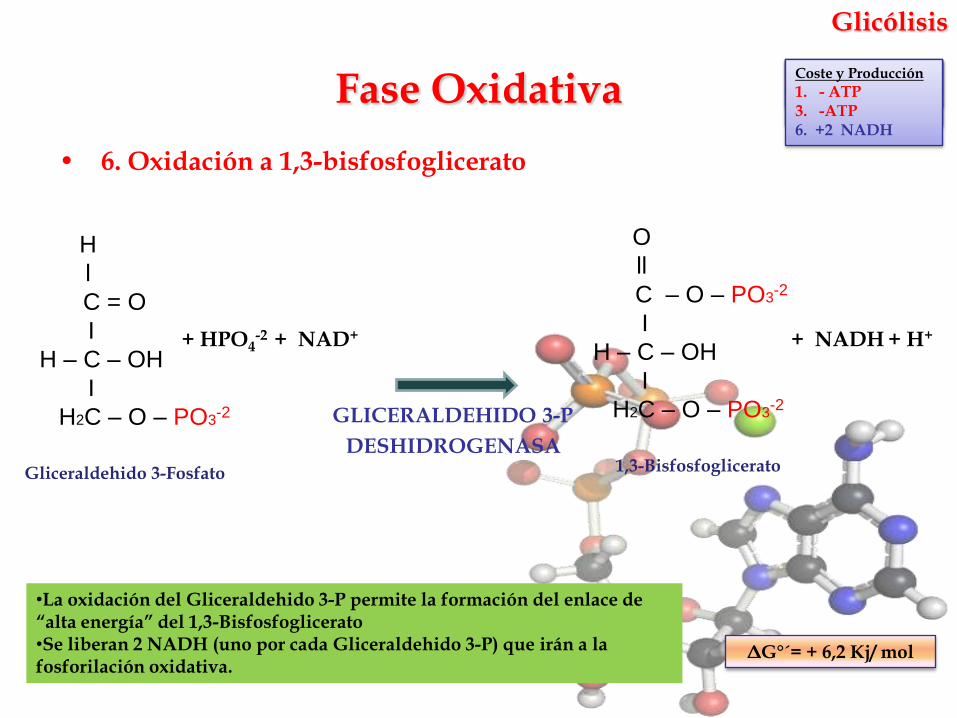
Glicólisis
• 6. Oxidación a 1,3-bisfosfoglicerato
GLICERALDEHIDO 3-P
DESHIDROGENASA
Fase OxidativaCoste y Producción
1. - ATP
ΔG°´= + 6,2 Kj/ mol
H
l
C = O
I
H – C – OH
I
H2C – O – PO3-2
Gliceraldehido 3-Fosfato
Coste y Producción
1. - ATP3. -ATP
O
ll
C – O – PO3-2
I
H – C – OH
I
H2C – O – PO3-2
+ HPO4-2 + NAD+ + NADH + H+
1,3-Bisfosfoglicerato
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
•La oxidación del Gliceraldehido 3-P permite la formación del enlace de “alta energía” del 1,3-Bisfosfoglicerato•Se liberan 2 NADH (uno por cada Gliceraldehido 3-P) que irán a la fosforilación oxidativa.
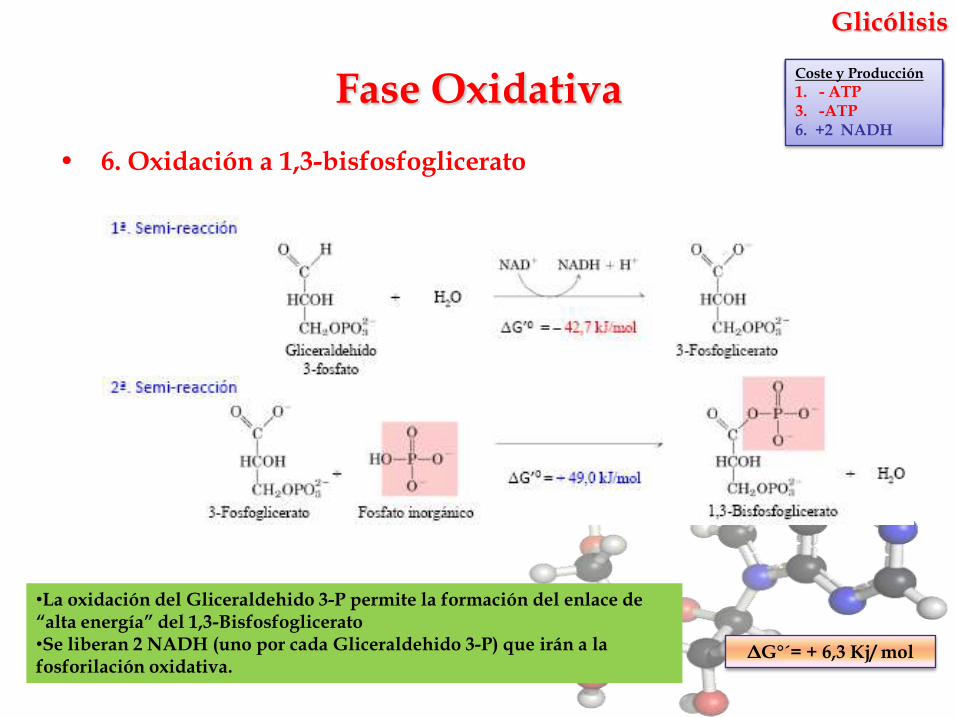
Glicólisis
• 6. Oxidación a 1,3-bisfosfoglicerato
Fase OxidativaCoste y Producción
1. - ATP
ΔG°´= + 6,3 Kj/ mol
Coste y Producción
1. - ATP3. -ATP
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
•La oxidación del Gliceraldehido 3-P permite la formación del enlace de “alta energía” del 1,3-Bisfosfoglicerato•Se liberan 2 NADH (uno por cada Gliceraldehido 3-P) que irán a la fosforilación oxidativa.
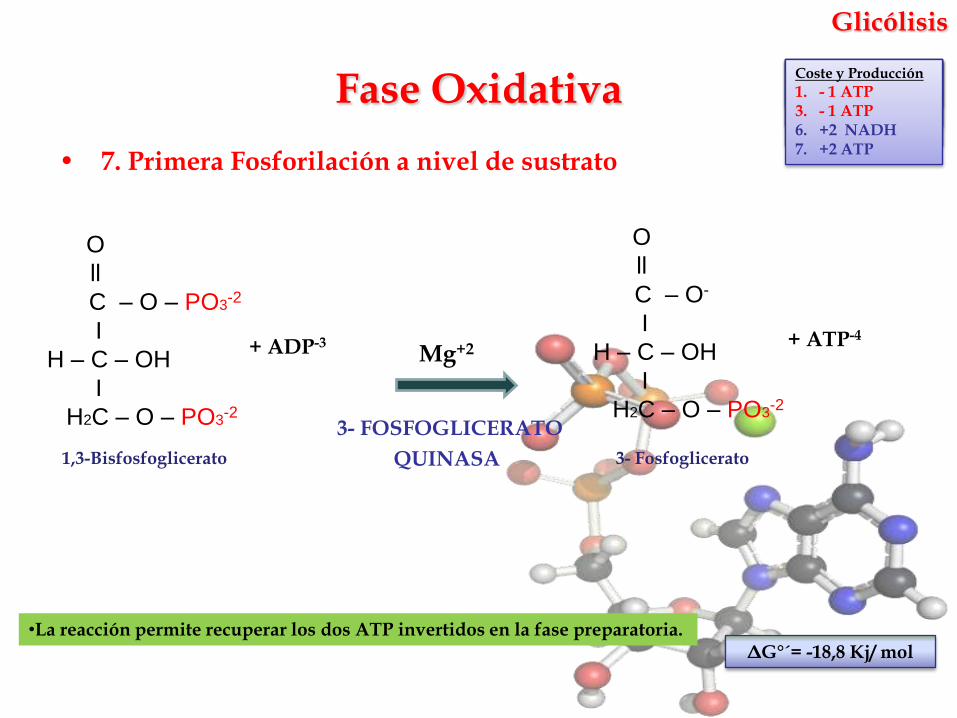
Glicólisis
• 7. Primera Fosforilación a nivel de sustrato
Mg+2
3- FOSFOGLICERATO
QUINASA
Fase OxidativaCoste y Producción
1. - ATP
ΔG°´= -18,8 Kj/ mol
Coste y Producción
1. - ATP3. -ATP
O
ll
C – O-
I
H – C – OH
I
H2C – O – PO3-2
+ ADP-3 + ATP-4
1,3-Bisfosfoglicerato
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
O
ll
C – O – PO3-2
I
H – C – OH
I
H2C – O – PO3-2
3- Fosfoglicerato
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP
•La reacción permite recuperar los dos ATP invertidos en la fase preparatoria.
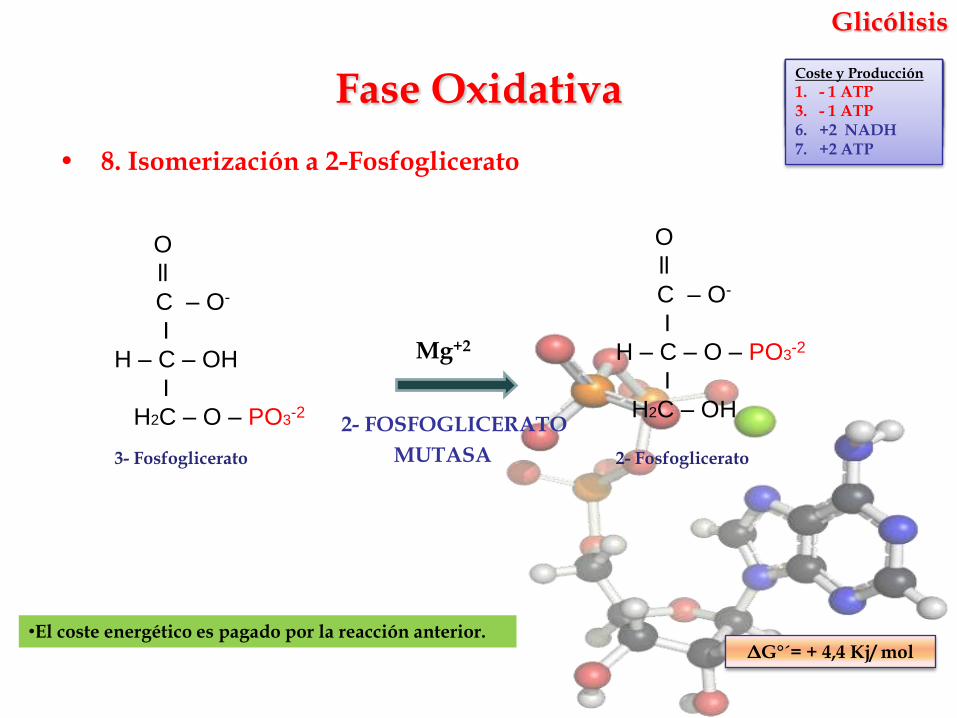
Glicólisis
• 8. Isomerización a 2-Fosfoglicerato
Mg+2
2- FOSFOGLICERATO
MUTASA
Fase OxidativaCoste y Producción
1. - ATP
ΔG°´= + 4,4 Kj/ mol
Coste y Producción
1. - ATP3. -ATP
O
ll
C – O-
I
H – C – O – PO3-2
I
H2C – OH
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
2- Fosfoglicerato
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP
O
ll
C – O-
I
H – C – OH
I
H2C – O – PO3-2
3- Fosfoglicerato
•El coste energético es pagado por la reacción anterior.
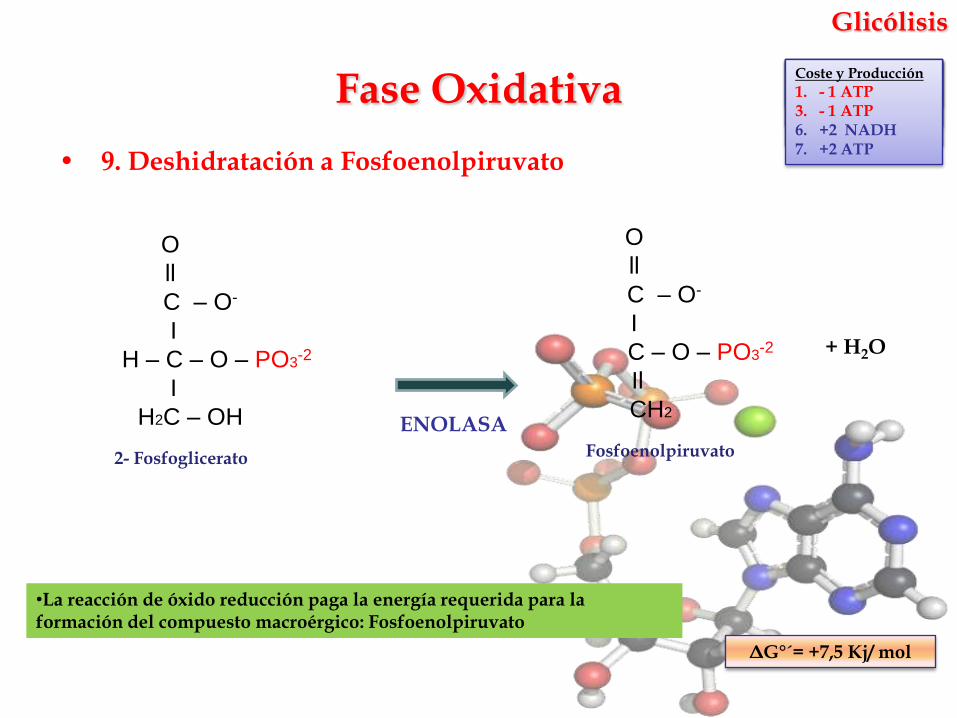
Glicólisis
• 9. Deshidratación a Fosfoenolpiruvato
ENOLASA
Fase OxidativaCoste y Producción
1. - ATP
ΔG°´= +7,5 Kj/ mol
Coste y Producción
1. - ATP3. -ATP
O
ll
C – O-
I
C – O – PO3-2
Il
CH2
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
Fosfoenolpiruvato
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP
2- Fosfoglicerato
O
ll
C – O-
I
H – C – O – PO3-2
I
H2C – OH
+ H2O
•La reacción de óxido reducción paga la energía requerida para la formación del compuesto macroérgico: Fosfoenolpiruvato
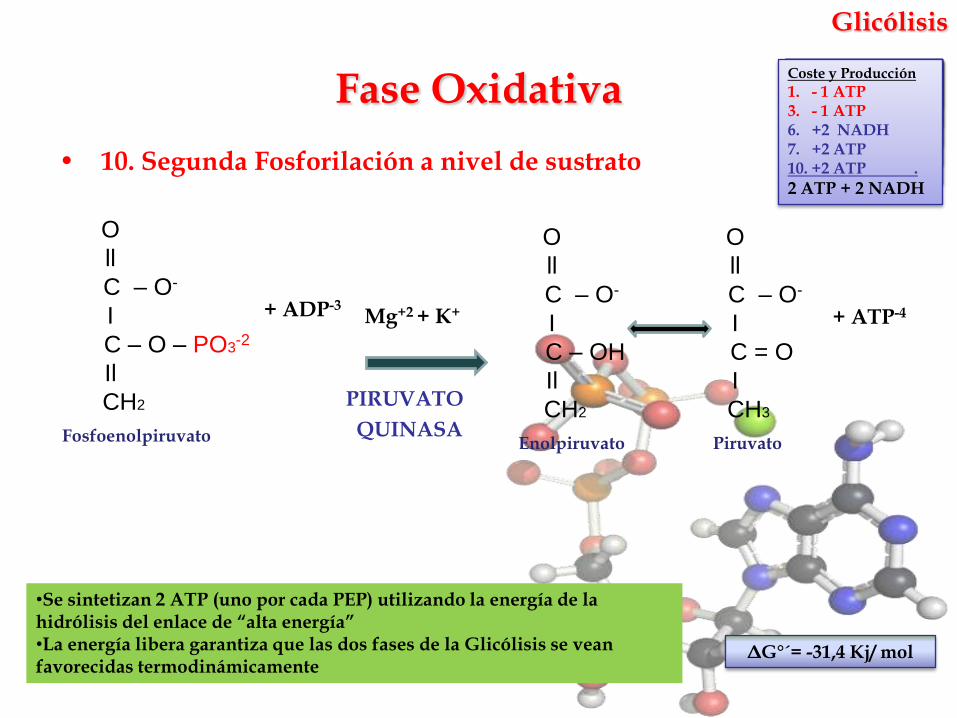
Glicólisis
• 10. Segunda Fosforilación a nivel de sustrato
Mg+2 + K+
PIRUVATO
QUINASA
Fase OxidativaCoste y Producción
1. - ATP
ΔG°´= -31,4 Kj/ mol
Coste y Producción
1. - ATP3. -ATP
O
ll
C – O-
I
C – O – PO3-2
Il
CH2
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
Fosfoenolpiruvato
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP
O
ll
C – O-
I
C – OH
Il
CH2
+ ADP-3
O
ll
C – O-
I
C = O
I
CH3
Enolpiruvato Piruvato
+ ATP-4
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP10. +2 ATP
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP10. +2 ATP .
2 ATP + 2 NADH
•Se sintetizan 2 ATP (uno por cada PEP) utilizando la energía de la hidrólisis del enlace de “alta energía”•La energía libera garantiza que las dos fases de la Glicólisis se vean favorecidas termodinámicamente
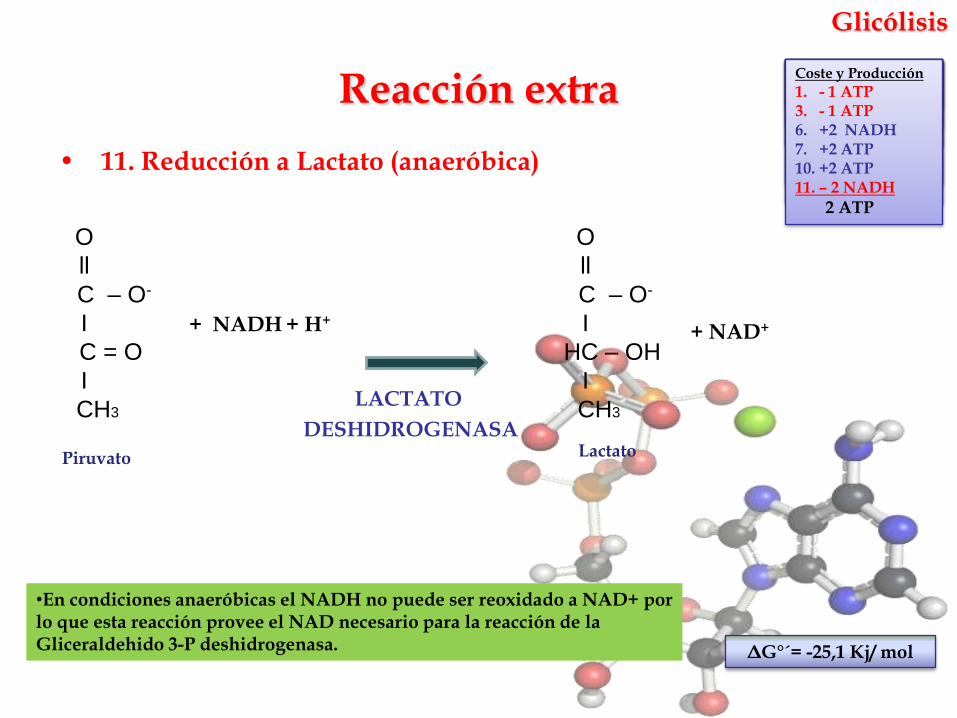
Glicólisis
• 11. Reducción a Lactato (anaeróbica)
LACTATO
DESHIDROGENASA
Reacción extraCoste y Producción
1. - ATP
ΔG°´= -25,1 Kj/ mol
Coste y Producción
1. - ATP3. -ATP
Coste y Producción
1. - ATP3. -ATP6. +2 NADH
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP10. +2 ATP
O
ll
C – O-
I
C = O
I
CH3
Lactato
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP10. +2 ATP11. – 2 NADH
Coste y Producción
1. - 1 ATP3. - 1 ATP6. +2 NADH7. +2 ATP10. +2 ATP 11. – 2 NADH
2 ATP
+ NADH + H+
O
ll
C – O-
I
HC – OH
I
CH3
+ NAD+
Piruvato
•En condiciones anaeróbicas el NADH no puede ser reoxidado a NAD+ por lo que esta reacción provee el NAD necesario para la reacción de la Gliceraldehido 3-P deshidrogenasa.
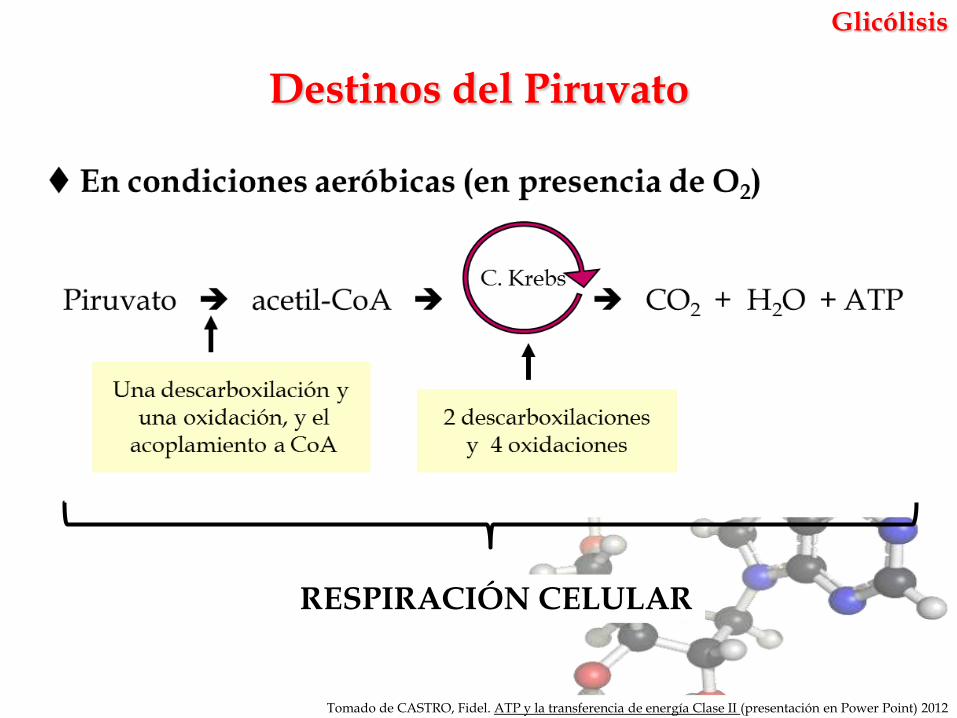
Glicólisis
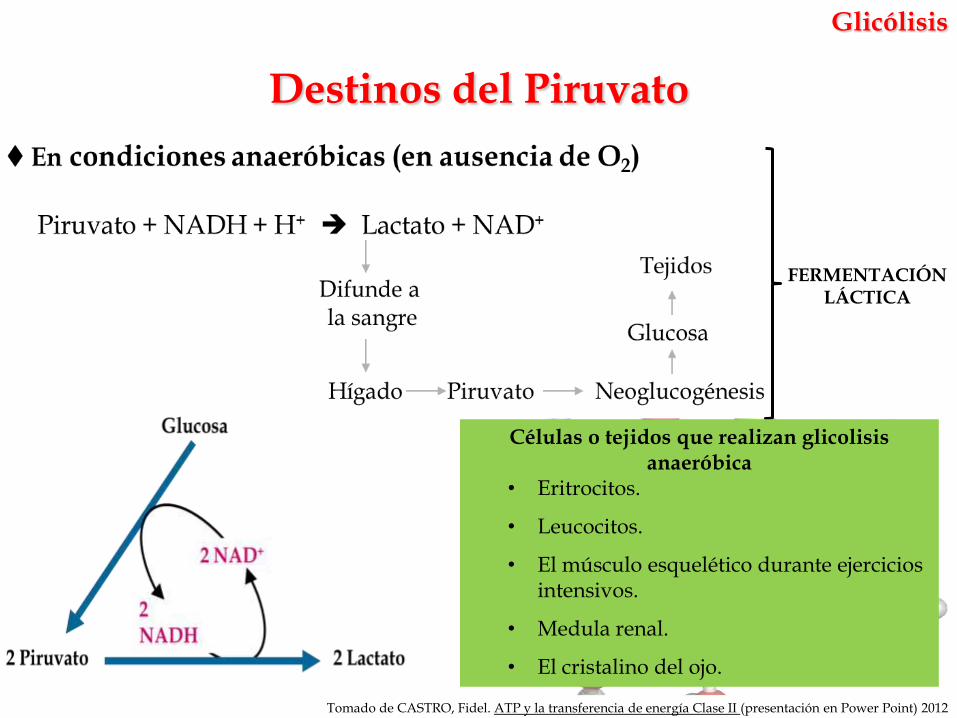
Destinos del Piruvato
RESPIRACIÓN CELULAR
Tomado de CASTRO, Fidel. ATP y la transferencia de energía Clase II (presentación en Power Point) 2012
Glicólisis
Destinos del Piruvato
FERMENTACIÓN LÁCTICA
Células o tejidos que realizan glicolisis anaeróbica
• Eritrocitos.
• Leucocitos.
• El músculo esquelético durante ejercicios intensivos.
• Medula renal.
• El cristalino del ojo.
Tomado de CASTRO, Fidel. ATP y la transferencia de energía Clase II (presentación en Power Point) 2012
Glicólisis
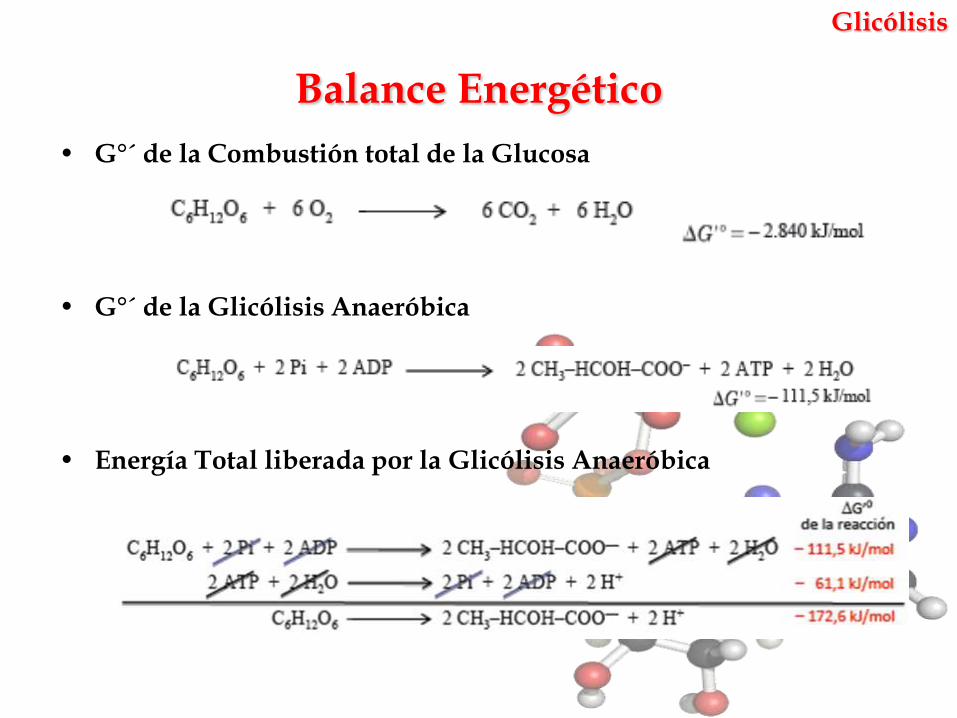
• G°´ de la Combustión total de la Glucosa
• G°´ de la Glicólisis Anaeróbica
• Energía Total liberada por la Glicólisis Anaeróbica
Balance Energético

Glicólisis
• Porcentaje del G°´ de la Combustión total de la Glucosa liberado por la Glicólisis Anaeróbica
• Conservación de los átomos durante la Glicólisis
Balance Energético
O
II
C – H
I
H – C – OH
I
HO – C – H
I
H – C – OH
I
H – C – OH
I
H2C – OH
C6H12O6
O
ll
C – O-
I
HO - C – H
I
CH3
+ 2H+
2 C3H6O3
O
ll
C – O-
I
HO - C – H
I
CH3
+ 2H+
C6H12O6

ESTRUCTURA BIOQUÍMICA DE LA CLASE
ATP y Glicólisis
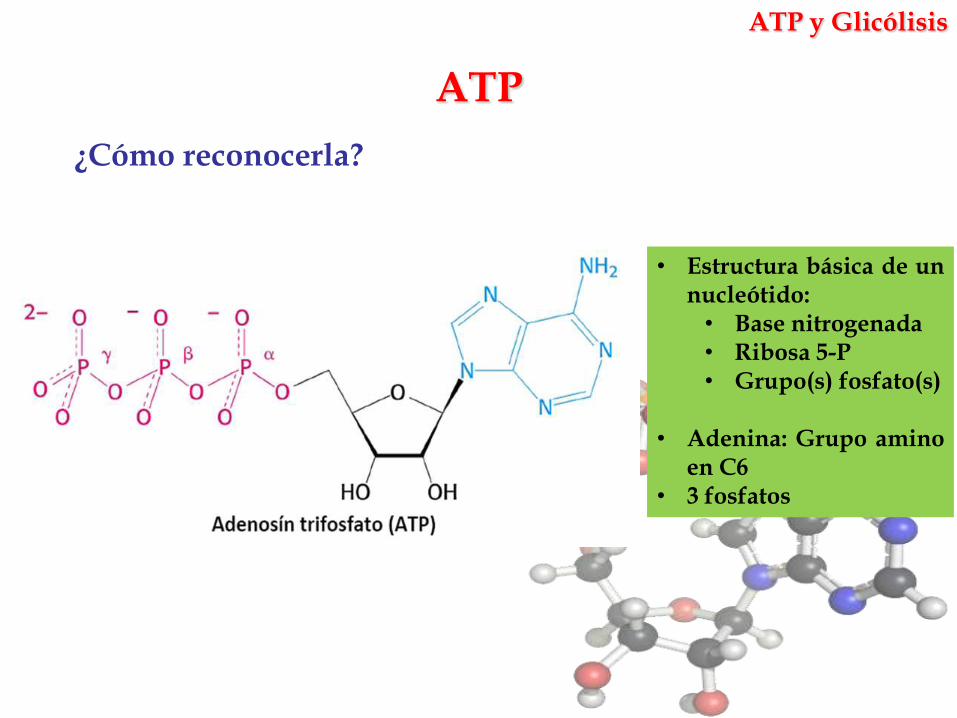
ATP
¿Cómo reconocerla?
• Estructura básica de unnucleótido:• Base nitrogenada• Ribosa 5-P• Grupo(s) fosfato(s)
• Adenina: Grupo aminoen C6
• 3 fosfatos
• Rodríguez, I(2009). El ATP y la Transferencia de la Energía. Presentación enPower Point. Cátedra de Bioquímica, Escuela de Medicina José María Vargas– UCV
• Rodríguez, I. El ATP y la Transferencia de la Energía. Cátedra deBioquímica, Escuela de Medicina José María Vargas – UCV
Bibliografía
“El primer deber de la inteligencia es desconfiar de si misma”
Anónimo
Gracias