antigenotoxic properties of selenium compounds on potassium dichromate and hydrogen peroxide
TRANSCRIPT

Antigenotoxic Properties of SeleniumCompounds on Potassium Dichromateand Hydrogen Peroxide
Eduardo Cemeli,1 Joanna Carder,1 Diana Anderson,1n Emma Guillamet,2
Marıa Jose Morillas,2 Amadeu Creus,2 and Ricardo Marcos2
1Department of Biomedical Sciences, University of Bradford, Richmond Road,Bradford, United Kingdom2Grup de Mutagenesi, Department de Genetica i de Microbiologia,Universitat Autonoma de Barcelona, Bellaterra, Spain
Selenium is an environmental metal that occurs ubiquitously and is producedthroughout the world for various industrial activities. Selenium has been reportedto have anticarcinogenic and preventive effects in clinical and epidemiologicalstudies. Selenium supplements can inhibit chemically-induced tumours. From theviewpoint of genotoxicity, selenium has not been adequately studied and anIARC review concluded that there were not sufficient data to consider it acarcinogen for man. In contrast, hexavalent chromium is classified as a knownrespiratory carcinogen producing DNA damage through free oxygen radicals. Inthe present study, a collaborative study has been carried out to evaluate thegenotoxicity of selenium compounds and their possible interactions withpotassium dichromate and hydrogen peroxide. Thus, in laboratory 1 (U.K.),the genotoxic effects of three selenium compounds were examined. Sodiumselenate, sodium selenite, and selenous acid were investigated in the Ames testusing strain TA102 and in the Comet assay using human lymphocytes, and alsoinvestigated for their interaction with potassium dichromate. In the Ames test, itwas shown that potassium dichromate produced a highly mutagenic response,
E.C. and E.G. contributed equally to this work.
Contract grant sponsor: CAEB of the Balearic Islands; Contract grant sponsor: Generalitat de Catalunya;
Contract grant number: SGR2002-00197; Contract grant sponsor: Spanish Ministry of Education and
Science (CAICYT); Contract grant number: PM98-0179.
nCorrespondence to: Professor Diana Anderson, Department of Biomedical Sciences, University of
Bradford, Richmond Road, Bradford, West Yorkshire BD7 1DP, UK.
E-mail: [email protected]
Accepted 21 June 2003
Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/tcm.10080
Teratogenesis, Carcinogenesis, and Mutagenesis Supplement 2:53–67 (2003)
r 2003 Wiley-Liss, Inc.

whilst the three selenium compounds did not. In combination, sodium selenatereduced the genotoxicity of potassium dichromate, but sodium selenite andselenous acid had no effect. In the Comet assay, potassium dichromate inducedDNA damage, but so did the selenium compounds. In combination withpotassium dichromate, however, only sodium selenate reduced its effect, whereassodium selenite and selenous acid exacerbated DNA damage. In laboratory 2(Spain), in the TK6 lymphoblastoid cell line, the Comet assay showed thatsodium selenite was non-genotoxic, while potassium dichromate and hydrogenperoxide induced DNA damage. It was also shown that sodium selenite did notdecrease the genotoxicity of potassium dichromate or hydrogen peroxide whenadministered as a pre-treatment or at the same time, or when potassiumdichromate and sodium selenite treatments were for different time periods. Thus,only sodium selenate has shown antigenotoxic properties against potassiumdichromate in the Ames test and in human lymphocytes in the Comet assay.Teratogenesis Carcinog. Mutagen. Suppl. 2:53–67, 2003. �c 2003 Wiley-Liss, Inc.
Key words: sodium selenate; sodium selenite; selenous acid; potassium dichromate; hydrogen peroxide;
Comet assay
INTRODUCTION
In practically all living organisms, heavy metals are present and are essential fornutrition and a wide range of biological functions. Non-essential metals are found ascontaminants of food-stuff and are ingested daily. Thus, the human diet contains bothessential and toxic metals [1]. Selenium is an environmental metal that occursnaturally in air, water, soil, plants, and food. It is currently produced throughout theworld for various industrial activities. At a nutritional level, selenium has been shownto have anticarcinogenic/preventive effects [1–4] supported by clinical and epidemio-logical studies [5]. Selenium supplements can inhibit chemically-induced tumours [6,7]. Selenium interacts with other metals protecting against the toxic effects ofcadmium and is antagonistic toward arsenic, copper, mercury and silver [8–10].
From the viewpoint of genotoxicity, selenium has been inadequately studied,and IARC (1975) [11] concluded that there were not sufficient data to considerselenium as a carcinogen for man.
In contrast, hexavalent chromium is classified as a known respiratorycarcinogen and is extensively absorbed by all routes in humans. After entering thecell, it is reduced, producing free oxygen radicals that cause lipid peroxidation andDNA damage [12]. The genotoxic effects of chromium compounds have beenreviewed by De Flora et al. [13]. Amrani et al. [14] have evaluated the genotoxicactivity of three chromium compounds in the somatic cells of Drosophila showingthat potassium chromate and dichromate are highly genotoxic and that sodiumselenite showed some antigenotoxic effects towards the chromium compound.
In the present study, in laboratory 1 (U.K.), the genotoxic effects of threeselenium compounds have been investigated, alone or in combination withpotassium dichromate, in the Ames test and the Comet assay using humanlymphocytes. The experiments in laboratory 2 (Spain) used a human lymphoblastoidcell line, TK6, in the Comet assay, to evaluate sodium selenite genotoxicity alone, orin combination with hydrogen peroxide and potassium dichromate. Hydrogenperoxide is a very well-known inducer of genotoxicity acting through oxygen radicalmechanisms. In the Comet assay [15, 16] its effects are abolished by catalase [15]. It issuggested that selenium acts by strengthening the antioxidative enzymes, namely
54 Cemeli et al.

catalase and peroxidase [17]. The aim of the present study was to confirm anyantigenotoxic properties of selenium in bacterial and mammalian cells.
METHODS
In laboratory 1, the Ames test and the Comet assay in human lymphocytes wereconducted according to standard methods. In laboratory 2, the Comet assay wasconducted in the human lymphoblastoid TK-6 cell line, that is a standard cell linewith a stable karyotype.
Chemicals
Laboratory 1
RPMI 1640 medium from Gibco (Paisley, UK); low and normal melting pointagarose, ampicillin, biotin, dimethyl sulphoxide (DMSO), ethidium bromide,histidine, sodium selenite (CAS No. 10102-18-8, purity 98%), sodium ammoniumphosphate, tetracycline, Triton X-100, and trypan blue from Sigma ChemicalCompany (St. Louis, MO); citric acid, magnesium sulphate, sodium chloride,sodium hydroxide, potassium chloride, potassium dichromate (Cas No.7778-50-9),potassium phosphate-dibasic, Tris-base, and EDTA from BDH (Poole, England);agar no. 1, nutrient broth no. 2, and phosphate buffered saline from Oxoid Ltd.(Basingstoke, Kent, UK); sodium selenate (CAS No.13910-01-0, purity 98%), andselenous acid (CAS No.7783-00-8, purity 98%), from Aldrich Chemical Co.(Mil-waukee, WI); anhydrous D-glucose from Fisher Chemicals (Loughborough, UK);Lymphoprep from Axis shield (Oslo, Norway); and foetal calf serum (FCS) fromNalgene (Rochester, NY).
Laboratory 2
Sodium selenite (CAS No. 21-448-5, purity 99%) was obtained from AldrichChemical Co. Potassium dichromate (CAS No. 7689-03-4, purity 99.9%), low andnormal melting temperature agarose, Triton X-100, Na2EDTA, Tris-HCl, PBS, andethidium bromide were obtained from Sigma (St. Louis, MO).
Prior to use, all the compounds were dissolved in distilled water.
Ames test
Procedures employed in the current study were standard [18–20]. Salmonella
typhymurium strain TA102 kindly provided by Safepharm (Derby, UK) was grownin nutrient broth with 8 mg/ml ampicillin and 0.8 mg/ml tetracycline. An aliquotfrom the broth was checked for optical density, (at l¼ 600 nm) before performingeach experiment. Bacteria were grown to a density of approximately 1.2� 109 cells/ml (O.D. about 1.2) and diluted 1:5 in sterile PBS ready for the Ames procedure.Sterile 15-ml screw top tubes were prepared with 2 ml of molten top agar (0.6% agar,5% sodium chloride, excess biotin, and trace histidine allowing just initial growth)kept at 451C. The appropriate chemical dilutions in water were added in 100 mlaliquots to triplicate tubes. Six-hundred microliters of the bacterial suspension werealiquoted in each tube. The mixture was immediately poured onto warm minimalglucose agar plates (2% anhydrous glucose, 1.5% agar with Vogel-Bonner salts [1� ]
Antigenotoxic Properties of Selenium Compounds 55

comprising magnesium sulphate, citric acid, potassium phosphate, and sodiumammonium phosphate). The plates were gently swirled to spread the top agar evenlybefore allowing it to solidify on a horizontal surface. The plates were then invertedand placed in an incubator at 371C for 72 hours. The number of colonies was scoredusing a manual colony counter (Anderman, Colony Counter, Surrey, UK). Onlythose experiments where the number of spontaneous mutants on the control platesfell within the expected mutation rate for TA102 were considered. Three experimentswere carried out, with triplicate plates for each experimental dose. Means of ninevalues per dose and their standard error were calculated and plotted in the graphs.
Comet Assay
In laboratory 1
Blood samples containing 50 ml/ml sodium citrate as an anticoagulant wereobtained from one healthy donor. Lymphocytes were isolated using Lymphoprep,which has a density of 1.077 g/ml and is composed of an aqueous solution of Ficoll(57 g/l) and sodium diotrizeate (90 g/l). Lymphocytes were stored in liquid nitrogencryovials in a mixture of 90% FCS and 10% DMSO at 108 cells/ml. Several cryovialsper experiment were thawed at room temperature prior to each experiment. Onehundred microliters of the cell suspension was added to each microtube containing100 ml of a defined concentration of the selenium compound (either sodium selenate,sodium selenite, or selenous acid to be tested on its own or in combination with 300mM potassium dichromate). Finally, RPMI was added up to a final volume of 1 ml.Microtubes were incubated for 30 min at 31C. Viability of the cells was checked bytrypan blue exclusion straight after incubation and only those tubes in whichviability was over 80% were included in the study.
The Comet assay was performed according to standard methods [15, 21].Conventional microscope slides from BDH (Poole, UK) were dipped into a solutionof 1% normal melting agarose (NMA) in distilled water at about 501C and allowedto dry at room temperature. This layer allowed the attachment of the second andthird layers. Immediately after incubation, the cell suspension was spun down and900 ml of the supernatant was removed. The pellet was resuspended in 75 ml of 1%low melting point agarose (LMA) in PBS at 371C. The cell suspension in agarose wasrapidly pipetted onto the first agarose layer and spread using a coverslip (22� 40mm) from BDH. The slides were placed on an ice-cold flat tray for 5 min to solidify.The coverslips were removed and 100 ml of 0.5% LMA in PBS at about 501C wereadded on the second layer, spread using a coverslip, and again placed on ice for 5min to solidify. Coverslips were removed and the slides were immersed in freshlyprepared cold lysing solution (2.5 M NaCl, 100 mM Na2 EDTA, and 10 mM Tris,pH 13), with 1% (v/v) Triton X-100 and 10% DMSO. The slides were kept in thelysing solution overnight at 41C.
The slides were removed from the lysing solution, drained, and placed in ahorizontal gel electrophoresis tank (model HU20, Scie-Plas, Renfrewshire, UK) sideby side, avoiding spaces and with the agarose ends facing towards the anode. Thetank was filled with fresh electrophoresis solution (1 mM Na2 EDTA and 300 mMNaOH pH 13.5) to a level approximately 0.25 cm above the slides. The slides wereleft in the solution for 30 min to allow the unwinding of the DNA and expression ofalkali labile damage before electrophoresis. Electrophoresis was conducted for
56 Cemeli et al.

30 min, using 25 V and adjusting the current to 300 mA by raising or lowering thebuffer level and using a compact power supply (2300 Microdrive 1, LKB, Sweden).Electrophoresis was conducted at 41C in darkness to prevent the occurrence ofadditional damage. After electrophoresis, the slides were placed horizontally on atray, and they were soaked with Tris buffer (0.4 M Tris, pH 7.5) for 1 min toneutralise the excess alkali. Slides were tipped sideways to drain off the buffer. Thisstep was repeated three times. Sixty microliters of ethidium bromide of a stocksolution (20 mg/ml) were pipetted onto each slide which was then covered with acoverslip (22� 40 mm). The slides were analysed immediately after.
Slides were examined at 625� magnification resulting from a 40� objective, a12.5� projection lens, and a 12.5� eyepiece on a fluorescence microscope (Leica,Germany) equipped with an excitation filter of BP 546/10 nm and a barrier filter of590 nm. The microscope was connected to the computer through a charge coupledevice (CCD) camera to transport images to Komet 4.0 software for analysis. Theimages appeared on the computer monitor screen at a magnification of 1,000.Twenty-five cells were scored from each replicate slide (i.e., 50 cells in total).
Values of DNA damage are automatically given as Tail moment. Tail momentis equivalent to the integrated value of density multiplied by migration distance,being considered the most sensitive measurement. The system set-up for thisequipment was as follows: head threshold 2%; tail threshold 0%; smoothing value 1;background height 20; and tail break lengths 5.
In laboratory 2
In laboratory 2, 106 cells were resuspended in 1 ml of RPMI 1640 medium. Thesingle cell gel electrophoresis (SCGE) or Comet assay was performed basicallyaccording to the lymphocyte protocol described above [15]. Following isolation, thecells were mixed with FDA and ethidium bromide and after 15 min cells werecounted and checked for viability (pre- and post-treatments).
Twenty microliters of TK-6 cells (about 40,000 cells) resuspended in RPMI 1640were incubated with 10 ml of a defined dose of the compound tested for 30 min at 371C.
One day prior to the study, 150 ml of normal melting agarose (NMA) was spreadon conventional slides and left to dry. The following day, a 3� 105 cells in 0.5% lowmelting agarose (LMA) (10 ml cell suspension in 75 ml LMA) were added to the slidesurface. Finally, the slide was covered with a third layer of 75 ml LMA.
Slides were immersed in ice-cold freshly prepared lysing solution (2.5 M NaCl,100 mM Na2EDTA, 10 mM tris-HCl, 1% L-laurosylsarcosinate, 1% Triton X-100,and 10% DMSO, pH¼ 0) to lyse the cells and allow DNA unfolding. After 2 h at41C in the dark, slides were placed on a horizontal electrophoresis unit. The unit wasfilled with fresh buffer (1 mM Na2EDTA, 300 mM NaOH, pH¼ 13.5) to cover theslides. The slides were left to set in the high-pH buffer for 40 min to allow DNAunwinding and expression of alkali-labile sites. Electrophoresis was conducted for20 min (0.73 V/cm and 300 mA). Slides were then gently washed to remove alkali in aneutralisation buffer (0.4 M Tris-HCl, pH¼ 7.5) and fixed with 3 ml of absoluteethanol for 3 min.
The slides were stained with 50 ml ethidium bromide (2 ml/ml). All stepsdescribed above were conducted under yellow light to prevent additional DNAdamage.
Antigenotoxic Properties of Selenium Compounds 57

The scoring was carried out by using a Komet 3.1 Image Analysis system fittedwith a Olympus 3� 50 fluorescence microscope equipped with a wide bandexcitation filter of 480–550 nm and barrier filter of 590 nm. One hundred randomlyselected cells were analysed and the Olive tail moment, % of DNA in tail, and taillength, were measured.
In the pre-treatment experiments, cells were incubated for 30 min with sodiumselenite and washed twice with RPMI before treatment with hydrogen peroxide orpotassium dichromate.
In the co-treatment experiments, the two compounds were added simulta-neously to the cells.
Statistical Analysis
In laboratory 1, statistics were performed on the means of five experiments.Thus, experimental data were considered to be parametric. Consequently, data werestatistically treated by the parametric analysis of variance (ANOVA).
In laboratory 2, statistics were performed using SAS/PC v8.0 (SAS Institute,Cary, NC). Statistical significance was determined by a general linear model with0.05 alpha level.
RESULTS
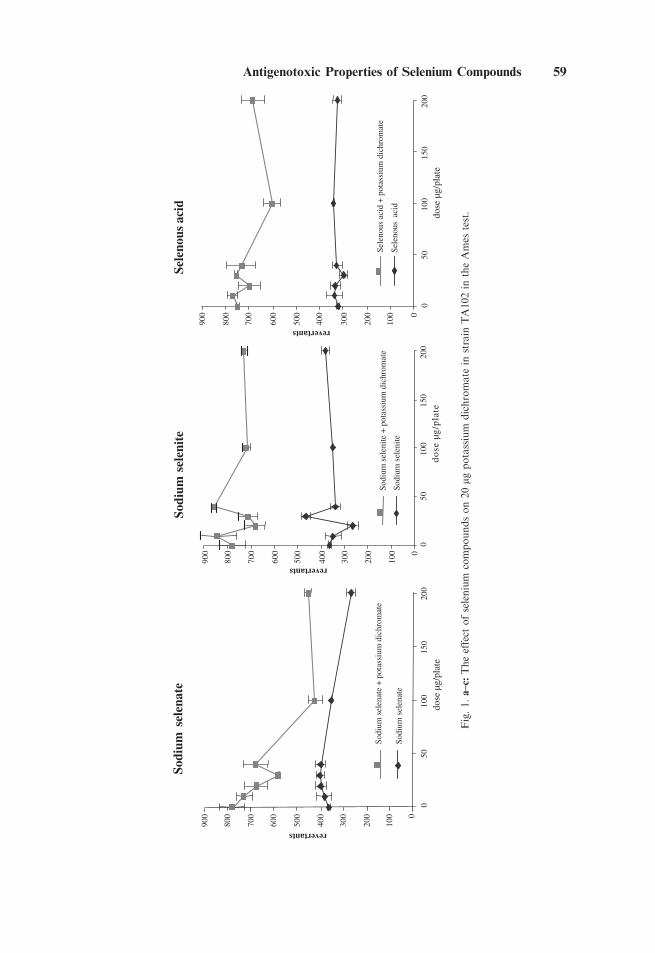
In laboratory 1, the results of the Ames test are shown in Figure 1a–c.Potassium dichromate was highly genotoxic with the greatest non-lethal effect at aconcentration around 20 mg/plate (data not shown). This dose was selected for use incombination with the various doses of sodium selenate, selenite, and selenous acid.The three selenium compounds on their own had little or no effect. In combinationwith sodium selenate, the genotoxic effect of potassium dichromate was significantlyreduced. In contrast, sodium selenite and selenous acid had little effect.
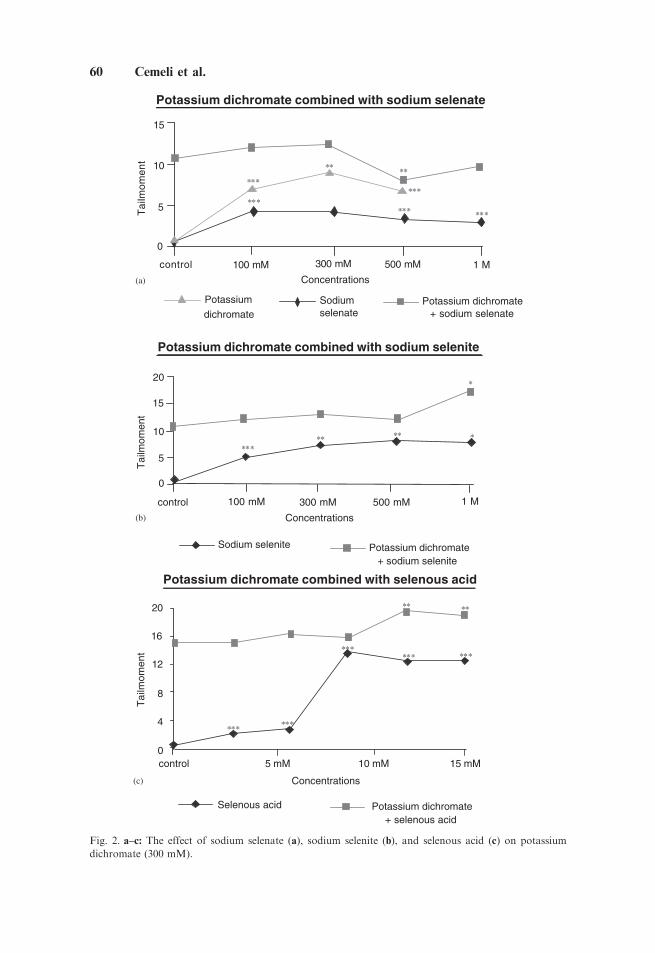
The results of the Comet assay are shown in Figure 2a–c. All selenium compoundsinduced some DNA damage, as did potassium dichromate. In combination withpotassium dichromate, however, sodium selenate reduced the DNA damage (Fig. 2a),but sodium selenite and selenous acid (Fig. 2b,c) exacerbated it.
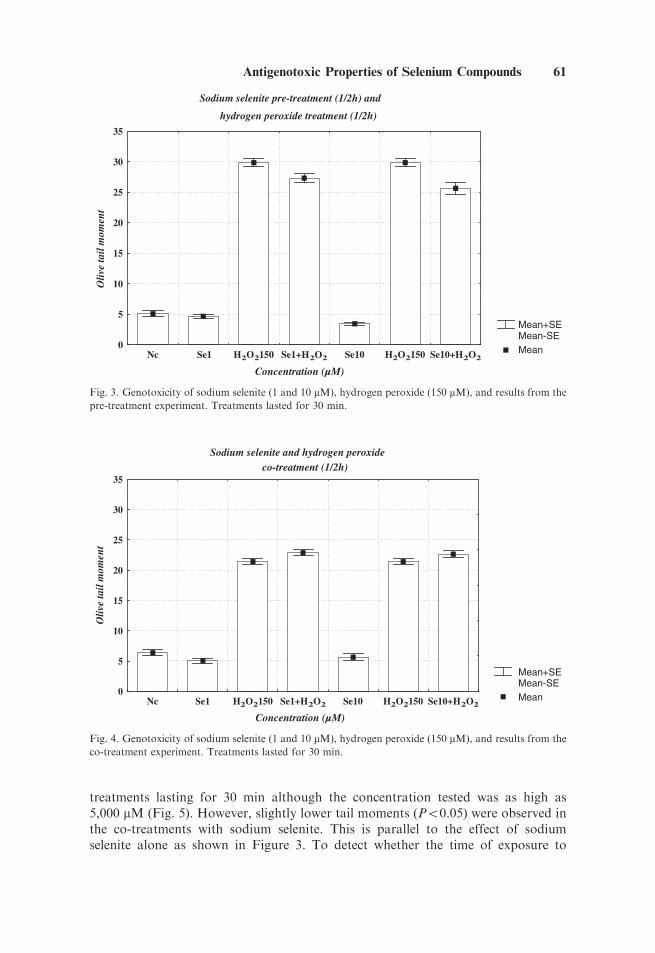
In laboratory 2, hydrogen peroxide was used as the positive control in the Cometassay. Figure 3 shows the effects with two concentrations of sodium selenite (1 and10 mM), and one concentration of hydrogen peroxide (150 mM), in treatments lastingfor 30 min. On its own, sodium selenite did not induce any genetic damage and in factshowed a slight reduction, compared with the negative control, whereas 150 mM ofhydrogen peroxide induced a significant increase in tail moments. Pretreatment of cellswith sodium selenite for 30 min followed by a treatment for 30 min with hydrogenperoxide resulted in a significant decrease in genetic damage induced by hydrogenperoxide alone. Different results were obtained in a second experiment where a co-treatment protocol was followed (Fig. 4). In this case, cells were treated simultaneouslywith sodium selenite and hydrogen peroxide for 30 min. No decrease was observedwith respect to the damage induced by hydrogen peroxide alone. In fact, there was anincrease (Po0.05) in damage with the low concentration (1 mM) of sodium selenite.
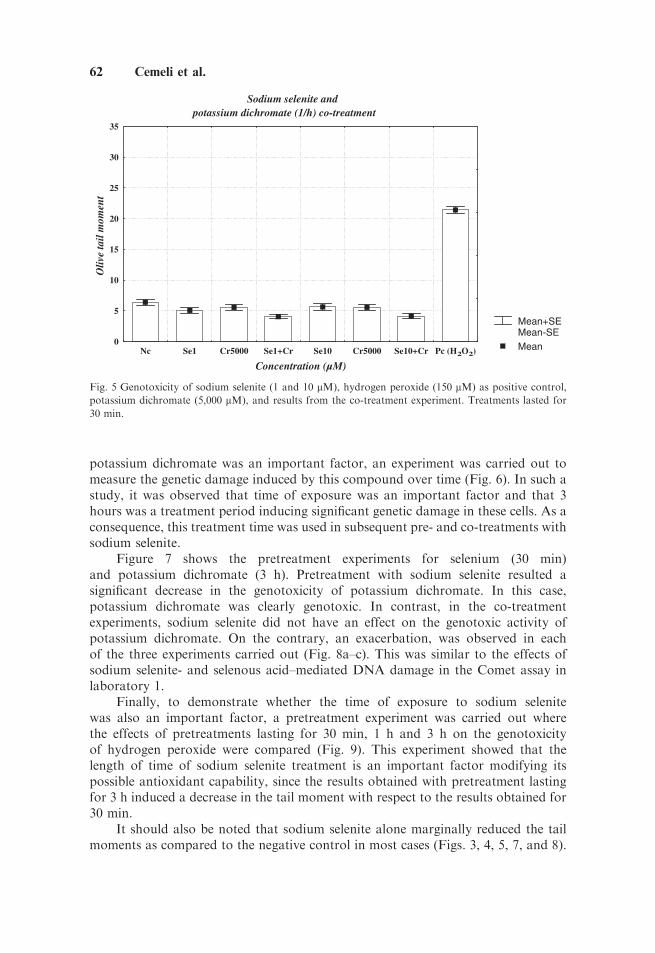
When the co-treatment experiment was carried out with potassium dichromate,it was observed that this compound alone did not induce genotoxicity in TK6 cells in
58 Cemeli et al.
Sodi
um s
elen
ate
0
100
200
300
400
500
600
700
800
900
050
100
150
200
dose
µg/
plat
e
revertants
Sodi
um s
elen
ate
Sodi
um s
elen
ate
+ p
otas
sium
dic
hrom
ate
Sodi
um s
elen
ite
0
100
200
300
400
500
600
700
800
900
050
100
150
200
do
se µ
g/p
late
revertants
Sodi
um s
elen
ite
Sodi
um s
elen
ite +
pot
assi
um d
ichr
omat
e
Sele
nous
aci
d
0
100
200
300
400
500
600
700
800
900
050
100
150
200
dose
µg/
plat
e
revertants
Sele
nous
aci
d
Sele
nous
aci
d +
pot
assi
um d
ichr
omat
e
Fig.1.a–c:
Theeffect
ofselenium
compoundson20mg
potassium
dichromate
instrain
TA102in
theAmes
test.
Antigenotoxic Properties of Selenium Compounds 59
Potassium dichromate combined with sodium selenite
Tai
lmom
ent
Sodium selenite Potassium dichromate+ sodium selenite
0
5
10
15
20
Concentrations
300 mMcontrol 100 mM 500 mM 1 M
Potassium dichromate combined with sodium selenate
Tai
lmom
ent
control
5
0
10
15
Concentrations
100 mM 500 mM 1 M300 mM
Potassium
dichromatePotassium dichromate
+ sodium selenateSodiumselenate
**
Concentrations
0
4
8
12
16
20
control 5 mM 10 mM 15 mM
Tai
lmom
ent ***
*** ***
*** ***
**
Potassium dichromate combined with selenous acid
Selenous acid Potassium dichromate+ selenous acid
***
**
***
**
****** ***
***** **
*
*
(a)
(b)
(c)
Fig. 2. a–c: The effect of sodium selenate (a), sodium selenite (b), and selenous acid (c) on potassium
dichromate (300 mM).
60 Cemeli et al.
treatments lasting for 30 min although the concentration tested was as high as5,000 mM (Fig. 5). However, slightly lower tail moments (Po0.05) were observed inthe co-treatments with sodium selenite. This is parallel to the effect of sodiumselenite alone as shown in Figure 3. To detect whether the time of exposure to
Mean+SEMean-SEMean
Sodium selenite pre-treatment (1/2h) and
hydrogen peroxide treatment (1/2h)
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 H2O2150 Se1+H2O2 Se10 H2O2150 Se10+H2O2
Fig. 3. Genotoxicity of sodium selenite (1 and 10 mM), hydrogen peroxide (150 mM), and results from the
pre-treatment experiment. Treatments lasted for 30 min.
Mean+SEMean-SEMean
Sodium selenite and hydrogen peroxide co-treatment (1/2h)
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 H2O2150 Se1+H2O2 Se10 H2O2150 Se10+H2O2
Fig. 4. Genotoxicity of sodium selenite (1 and 10 mM), hydrogen peroxide (150 mM), and results from the
co-treatment experiment. Treatments lasted for 30 min.
Antigenotoxic Properties of Selenium Compounds 61
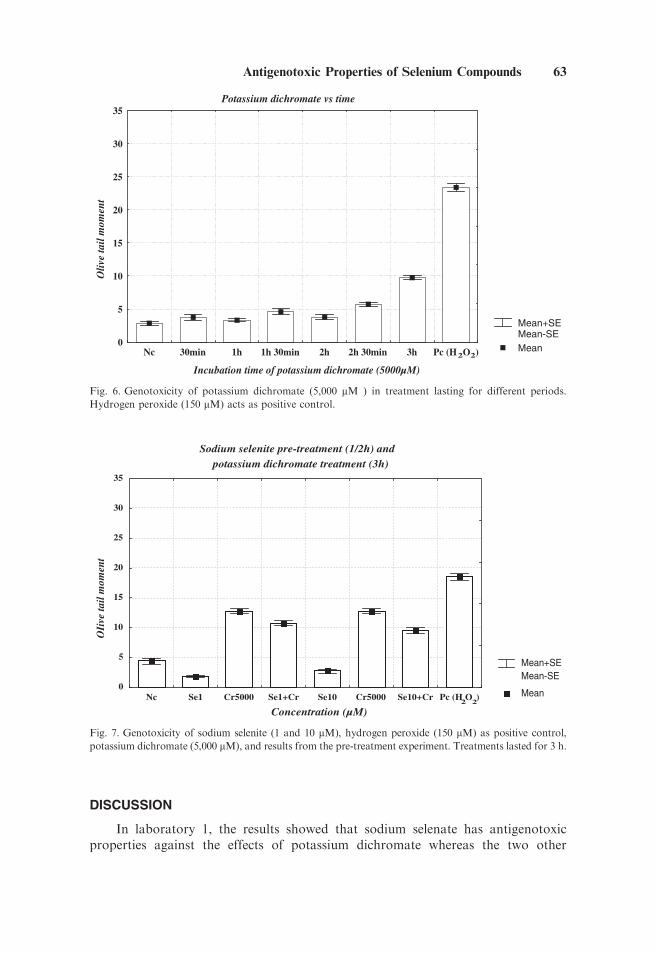
potassium dichromate was an important factor, an experiment was carried out tomeasure the genetic damage induced by this compound over time (Fig. 6). In such astudy, it was observed that time of exposure was an important factor and that 3hours was a treatment period inducing significant genetic damage in these cells. As aconsequence, this treatment time was used in subsequent pre- and co-treatments withsodium selenite.
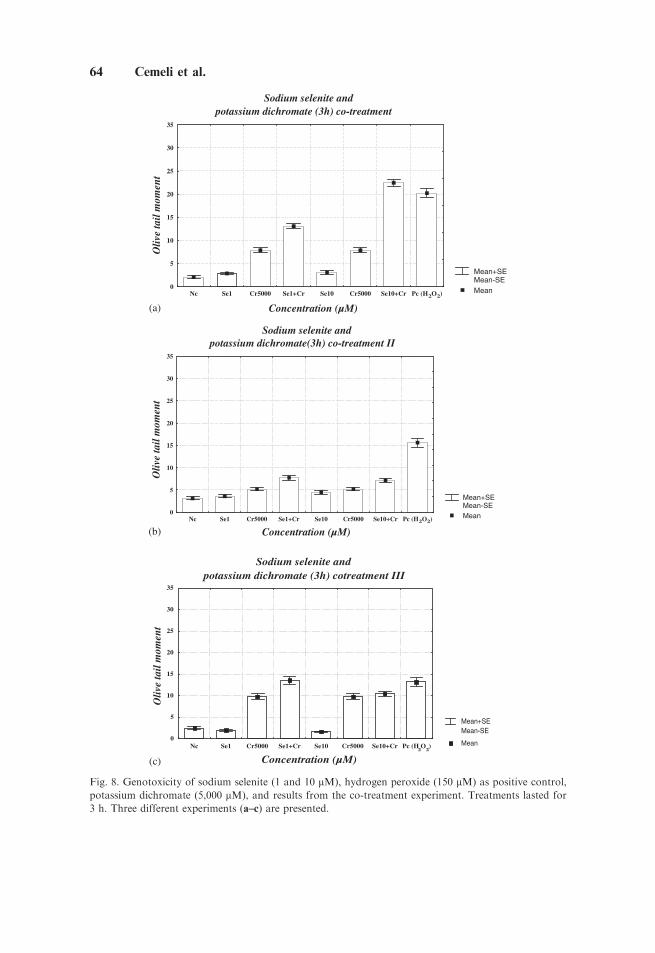
Figure 7 shows the pretreatment experiments for selenium (30 min)and potassium dichromate (3 h). Pretreatment with sodium selenite resulted asignificant decrease in the genotoxicity of potassium dichromate. In this case,potassium dichromate was clearly genotoxic. In contrast, in the co-treatmentexperiments, sodium selenite did not have an effect on the genotoxic activity ofpotassium dichromate. On the contrary, an exacerbation, was observed in eachof the three experiments carried out (Fig. 8a–c). This was similar to the effects ofsodium selenite- and selenous acid–mediated DNA damage in the Comet assay inlaboratory 1.
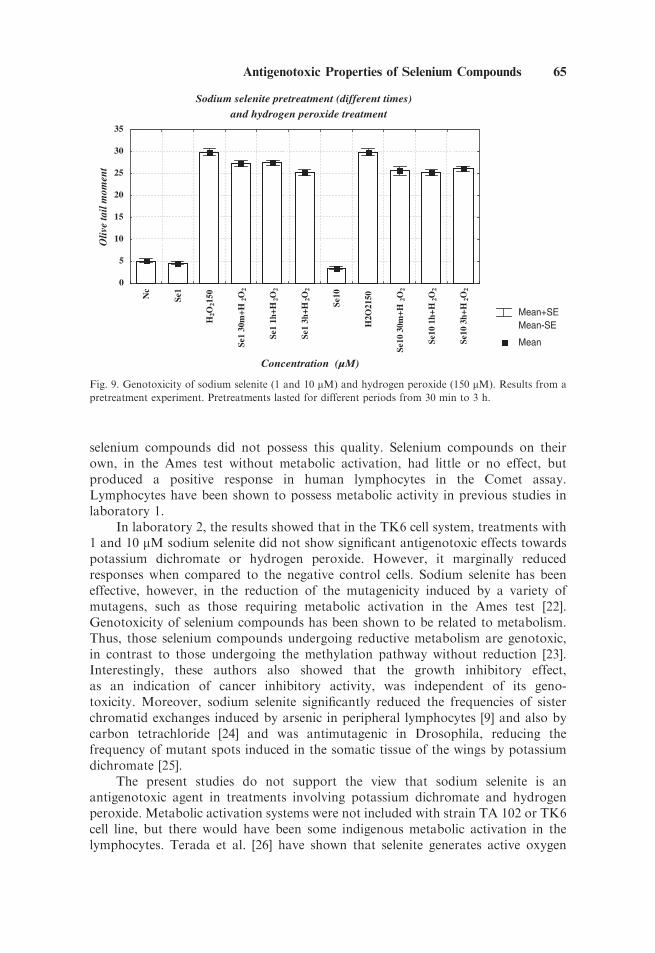
Finally, to demonstrate whether the time of exposure to sodium selenitewas also an important factor, a pretreatment experiment was carried out wherethe effects of pretreatments lasting for 30 min, 1 h and 3 h on the genotoxicityof hydrogen peroxide were compared (Fig. 9). This experiment showed that thelength of time of sodium selenite treatment is an important factor modifying itspossible antioxidant capability, since the results obtained with pretreatment lastingfor 3 h induced a decrease in the tail moment with respect to the results obtained for30 min.
It should also be noted that sodium selenite alone marginally reduced the tailmoments as compared to the negative control in most cases (Figs. 3, 4, 5, 7, and 8).
Mean+SEMean-SEMean
Sodium selenite and potassium dichromate (1/h) co-treatment
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 Cr5000 Se1+Cr Se10 Cr5000 Se10+Cr Pc (H2O2)
Fig. 5 Genotoxicity of sodium selenite (1 and 10 mM), hydrogen peroxide (150 mM) as positive control,
potassium dichromate (5,000 mM), and results from the co-treatment experiment. Treatments lasted for
30 min.
62 Cemeli et al.
DISCUSSION
In laboratory 1, the results showed that sodium selenate has antigenotoxicproperties against the effects of potassium dichromate whereas the two other
Mean+SEMean-SEMean
Potassium dichromate vs time
Incubation time of potassium dichromate (5000µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc 30min 1h 1h 30min 2h 2h 30min 3h Pc (H2O2)
Fig. 6. Genotoxicity of potassium dichromate (5,000 mM ) in treatment lasting for different periods.
Hydrogen peroxide (150 mM) acts as positive control.
Mean+SEMean-SE
Mean
Sodium selenite pre-treatment (1/2h) and potassium dichromate treatment (3h)
Concentration (µM)
OIi
ve ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 Cr5000 Se1+Cr Se10 Cr5000 Se10+Cr Pc (H2O
2)
Fig. 7. Genotoxicity of sodium selenite (1 and 10 mM), hydrogen peroxide (150 mM) as positive control,
potassium dichromate (5,000 mM), and results from the pre-treatment experiment. Treatments lasted for 3 h.
Antigenotoxic Properties of Selenium Compounds 63
Mean+SEMean-SEMean
Sodium selenite and potassium dichromate(3h) co-treatment II
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 Cr5000 Se1+Cr Se10 Cr5000 Se10+Cr Pc (H2O2)
Mean+SEMean-SE
Mean
Sodium selenite and potassium dichromate (3h) cotreatment III
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 Cr5000 Se1+Cr Se10 Cr5000 Se10+Cr Pc (H2O
2)
Mean+SEMean-SEMean
Sodium selenite and potassium dichromate (3h) co-treatment
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc Se1 Cr5000 Se1+Cr Se10 Cr5000 Se10+Cr Pc (H2O2)
(a)
(b)
(c)
Fig. 8. Genotoxicity of sodium selenite (1 and 10 mM), hydrogen peroxide (150 mM) as positive control,
potassium dichromate (5,000 mM), and results from the co-treatment experiment. Treatments lasted for
3 h. Three different experiments (a–c) are presented.
64 Cemeli et al.
selenium compounds did not possess this quality. Selenium compounds on theirown, in the Ames test without metabolic activation, had little or no effect, butproduced a positive response in human lymphocytes in the Comet assay.Lymphocytes have been shown to possess metabolic activity in previous studies inlaboratory 1.
In laboratory 2, the results showed that in the TK6 cell system, treatments with1 and 10 mM sodium selenite did not show significant antigenotoxic effects towardspotassium dichromate or hydrogen peroxide. However, it marginally reducedresponses when compared to the negative control cells. Sodium selenite has beeneffective, however, in the reduction of the mutagenicity induced by a variety ofmutagens, such as those requiring metabolic activation in the Ames test [22].Genotoxicity of selenium compounds has been shown to be related to metabolism.Thus, those selenium compounds undergoing reductive metabolism are genotoxic,in contrast to those undergoing the methylation pathway without reduction [23].Interestingly, these authors also showed that the growth inhibitory effect,as an indication of cancer inhibitory activity, was independent of its geno-toxicity. Moreover, sodium selenite significantly reduced the frequencies of sisterchromatid exchanges induced by arsenic in peripheral lymphocytes [9] and also bycarbon tetrachloride [24] and was antimutagenic in Drosophila, reducing thefrequency of mutant spots induced in the somatic tissue of the wings by potassiumdichromate [25].
The present studies do not support the view that sodium selenite is anantigenotoxic agent in treatments involving potassium dichromate and hydrogenperoxide. Metabolic activation systems were not included with strain TA 102 or TK6cell line, but there would have been some indigenous metabolic activation in thelymphocytes. Terada et al. [26] have shown that selenite generates active oxygen
Mean+SEMean-SE
Mean
Sodium selenite pretreatment (different times)
and hydrogen peroxide treatment
Concentration (µM)
Oliv
e ta
il m
omen
t
0
5
10
15
20
25
30
35
Nc
Se1
H2O
2150
Se1
30m
+H2O
2
Se1
1h+H
2O2
Se1
3h+H
2O2
Se10
H2O
2150
Se10
30m
+H2O
2
Se10
1h+
H2O
2
Se10
3h+
H2O
2
Fig. 9. Genotoxicity of sodium selenite (1 and 10 mM) and hydrogen peroxide (150 mM). Results from a
pretreatment experiment. Pretreatments lasted for different periods from 30 min to 3 h.
Antigenotoxic Properties of Selenium Compounds 65

species in cultured endothelial cells in the presence of sulphydryl compounds,whereas selenate does not. Bacterial cells do not possess any glutathione peroxidase,whereas mammalian cells do [27]. Glutathione peroxidase catalyses the oxidation ofglutathione. Once glutathione reacts with protein thiols, mixed disulphides areformed that can accumulate with proteins and other thiols causing toxicity as aconsequence of oxidative stress. This could explain the genotoxic effects of sodiumselenite in human lymphocytes. Presumably, selenous acid behaves in a similarmanner to sodium selenite. Also the pH of the medium could affect the capability ofselenium compounds to transfer electrons, and since oxidative systems are involved,differing responses in different systems in the literature might be explained by theswitch between their antioxidant and pro-oxidant properties. The marginalreductions in effects by sodium selenite, by comparison with control TK6 cellsand the antigenotoxic responses with sodium selenate, might well be explained by theantioxidant status pertaining during experimentation.
Thus, in conclusion, sodium selenite did not decrease the genotoxicity ofpotassium dichromate or hydrogen peroxide, when administered as a pre-treatmentor at the same time, or when potassium dichromate and sodium selenite treatmentswere for different time periods. Sodium selenite and selenous acid were shown toexacerbate responses. Only sodium selenate has shown antigenotoxic propertiesagainst potassium dichromate in the Ames test and in human lymphocytes in theComet assay.
ACKNOWLEDGMENTS
Eduardo Cemeli (laboratory 1) received Leonardo da Vinci grants from CAEBof the Balearic Islands. The investigation in laboratory 2 has been supported in partby a grant from the Generalitat de Catalunya (SGR2002-00197) and from theSpanish Ministry of Education and Science (CAICYT, PM98-0179). M.J. Morillasheld a fellowship from the same Ministry.
REFERENCES
1. Rojas E, Herrera LA, Poirier L.A., Ostrosky-Wegman P. Are metals dietary carcinogens? Mutat Res
1999;443:157–181.
2. Griffin AC. Role of selenium in the chemoprevention of cancer. Adv Cancer Res 1979;29:419–442.
3. Shamberger RJ. In: Frieden E, editor. The biochemistry of selenium. New York: Plenum Press. 1983.
p 1–334.
4. Shamberger RJ. The genotoxicity of selenium. Mutat Res 1985;154:29–48.
5. Lange JH. Reanalysis of epidemiological data for selenium anticancer activity. Toxicol Ind Health
1991;7:319 –325.
6. Medina D, Shephard F. Selenium-mediated inhibition of mouse mammary tumorigenesis. Cancer Lett
1980;8:241–245.
7. Whanger PD. Selenium interactions with carcinogens. Fundam Appl Toxicol 1983;3:424–430.
8. Gras G, Mondain J. Mercury and methyl mercury pollution of fishery products: toxicological effects
on human health. Toxicol Eur Res 1981;3:243–259.
9. Hu G, Liu X, Liu J. Protective effects of sodium selenite and selenomethionine on genotoxicity to
human peripheral lymphocytes induced by arsenic Chung hua Yu Fang Ihsueh Tsa Chih 1996;30:
26–29 (in Chinese).
10. Zhang Y, Xiao H. Antagonistic effect of calcium, zinc and selenium against cadmium induced
chromosomal aberrations and micronuclei in root cells of Hordeum vulgare. Mutat Res 1998;420:1–6.
66 Cemeli et al.

11. IARC. Some azaridines, N,-S-, and mustards and selenium. IARC monographs on the evaluation of
carcinogenic risk of chemicals to humans, vol. 9.Lyon, France: International Agency for Research on
Cancer. 1975.
12. Nicolas B, Descotes J. Metals. In: Descotes J, editor. Human toxicology. Amsterdam: Elsevier B.V.
1996. p 515–540.
13. De Flora S, Bagnasco M, Serra D, Zanacchi P. Genotoxicity of selenium compounds: a review. Mutat
Res 1990;238:99–172
14. Amrani S, Rizki M, Creus A, Marcos R. Genotoxic activity of different chromium compounds in
larval cells of Drosophila melanogaster, as measured by the wing spot test. Environ Mol Mutagen
1999;34:47–51.
15. Anderson D, Yu T-W, Phillips BJ and Schmezer P. The effect of various anti-oxidants and other
modifying agents on oxygen radical generated DNA damage in human lymphocytes in the Comet
assay. Mutat Res 1994;307:261–271
16. Morillas MJ, Guillamet E, Surralles J, Creus A, Marcos R. Spontaneous and induced genetic damage
in T-lymphocyte subsets evaluated by the Comet assay. Mutat Res 2002;514:39–48
17. Kaur M, Wadhwa R, Sharma SP. Effect of sodium selenite on antioxidative enzymes of banana
fruitfly. Gerontology 1989;35:188–191
18. Ames BN, McCann J, Yamasaki E. Methods for detecting carcinogens and mutagens with the
Salmonella/mammalian–microsome mutagenicity test. Mutation Res 1975;31:347–364.
19. Maron DM, Ames BN. 1983. Revised methods for the Salmonella mutagenicity test. Mutat Res
1983;113:173–215.
20. Mortelmans K, Zieger E. The Ames Salmonella/microsome mutagenicity. Mutat Res 2000;29–60.
21. Singh NP, Danner DB, Tice RR, Brant L, Schneider EL. A simple technique for quantitation of low
levels of DNA damage in individual cells. Exp Cell Res 1988;175:184–191
22. Martin SE, Adams GH, Schillaci M, Milner JA. Antimutagenic effect of selenium on acridine orange
and 7,12-dimethylbenzo [a]anthracene in the Ames Salmonella/microsomal system. Mutat Res
1981;82:41–46.
23. Lu J, Jiang C, Kaeck M, Ganther H, Vadhanavikit S, Ip C. Thompson H. Dissociation of the
genotoxic and growth inhibitory effects of selenium. Biochem Pharmacol 1995;50:213–219.
24. Sivikova K, Piesova E, Dianovsky J. The protection of Vitamin E and selenium against carbon
tetrachloride-induced genotoxicity in ovine peripheral blood lymphocytes. Mutat Res 2001;494:
134–142
25. Rizki M, Amrani S, Creus A, Xamena N, Marcos R. Antigenotoxic properties of selenium. Studies in
the wing spot test in Drosophila. Environ Mol Mutagen 2001;37:70–75.
26. Terada A, Yoshida M, SekoY, Kobayashi T, Yoshida K, Nakada M, Nakada K, Echizen H, Ogata
H, Rikihisa T. Active oxygen species generation and cellular damage by additives of parenteral
preparations: selenium and sulfhydryl compounds. Nutrition 1999;651–655.
27. Halliwell B, Gutteridge J. Free radicals in Biology and Medicine. Oxford: Oxford University Press.
1999; Vol 15(a).
Antigenotoxic Properties of Selenium Compounds 67