anatomy of tomato fruit and fruit pedicel during fruit ...formatex.info/microscopy4/851-861.pdf ·...
TRANSCRIPT

Anatomy of tomato fruit and fruit pedicel during fruit development
D. Rančić, S. Pekić Quarrie and I. Pećinar
Faculty of Agriculture University of Belgrade, Nemanjina 6 11080 Zemun, Serbia
Paper reviews on the anatomy of berry fruit development by comparative analysis of fruit and fruit pedicel in tomato combining our results with other authors. We have presented data on developmental analysis of fruit: measurements of different anatomical paramenters in fruit and fruit pedicel together with parameters of the fruit size. Anatomical background of the water and solute transport during fruit development is elaborated by discussing all possible factors affecting fruit development. The genotypes used in our research were tomato deficient mutant and wild type to try to elucidate role of ABA in these proceses. Fruits of the ABA deficient mutant are smaller as consequence of smaller-sized cells in pericarp since functional xylem area in fruit pedicels and diameter of xylem elements in both genotypes are similar. Wild type has more nonfunctional xylem that could serve as a mechanical support for heavier fruits. Lower fruit dry weight and phloem efficiency in pedicels of ABA deficient mutant compared to the wild type suggests important role of ABA in the fruit sink activity.
Keywords xylem; phloem, ABA, tomato
1. Introduction
In tomato as well as in other crops size of the fruit is the key factor determining yield. This is why mechanisms of regulating fruit growth and development were the research topic for many authors [1, 2]. Crop yield is the result of many morfo-physiological and biochemical processes depending on environmental factors and genetical background [3]. As the growth of fleshy fruits is to the great extent result of water accumulation, for the understanding of the fruit developmental processes it is important to understand water transport as well as coordination between long distance transport of water and solutes and short distance transports such as water absorption of the individual fruit cells [4]. Understanding of the plant and fruit anatomy could be, therefore, the key factor in understanding transport of water, assimilates and signalling molecules within the plant [5] and, consequently its effects on yield. There are only few papers reporting tomato fruit anatomy [6-8], while detailed anatomical and histochemical analysis during tomato fruit development was done by Gillaspy et al. [9]. Cytological analysis of tomato fruit has shown that in the early phase of fruit development cell division is the main limiting factor for fruit growth [10], and also that cytological changes at the this phase are strongly dependent on transport of water and assimilates into the fruit [11]. For understanding factors affecting fruit growth and development it is important to analyse comparatively anatomy of both, fruit and fruit pedicel. Here we present detailed anatomical analysis of the tomato fruit and fruit pedicel anatomy during fruit development to be able to show to which extent anatomy of the fruit pedicel determines/restricts water transport into the fruit and, consequently, affects fruit size and fruit anatomy. Material used in this investigation: tomato ABA deficient mutant and wild type represent good objects for examining of the role of ABA on plant morphology and physiology [12]. For example, fruits of the ABA deficient mutant are smaller [13] and it is not known is it due to the smaller number of cells in the pericarp or their smaller size due to reduced turgor. Comparative analysis of fruit anatomy in both genotypes could provide explanation of these differences in fruit anatomy as well as to the possible role of ABA.
2. Materials and methods
Two tomato genotypes (Lycopersicon esculentum Mill.): wild type (Ailsa Craig) and flacca (CM Rick Tomato Genetics Resource Center, USA) were investigated. Leaves of wild type in optimal water conditions have about 2.9 ng ABA per mg dry weight [14], while flacca leaves only 26% amount of ABA compared with wild type [15]. The plants were grown from seeds in commercial substrate (Potground H, Klasmann-Deilmann, Germany) in chamber operating with a 14h photoperiod with light intensity at plant level 300 µmolm-2s-1, temperature 25/18˚C and relative humidity 70%. The root system was irrigated daily to a soil water content represents 80% of maximum soil water capacity. Fruits and fruit pedicels were collected in three developmental phases: 12, 20 and 39 days after anthesis. Fruit diameter, fresh and dry weight, thickness and number of cell layers in fruit pericarp, egzo and mezocarp, as well as phloem area, diameter of xylem elements, xylem area and functional xylem area of the pedicel at the transversal sections 5mm apart from the fruit were measured. Phloem efficiency was calculated by dividing fruit dry weight by phloem pedicel area at the final stage of fruit development.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 851
______________________________________________

2.1 Light microscopy
Slides for light microscopy were made according to standard procedure. Samples were fixed in FAA for 24h, postfixed in 70% ethanol and dehydrated in a graded ethanol series. After tissue impregnation in Histowax (56-58°C) samples were embedded. After cooling the blocks on a cold plate and solidifying paraffin, histological sections of about 5–7 µm were cut using a microtome (Leica SM 2000 R). Before staining, the paraffin is removed from the sections by xylene, followed by rehidratation in graded series of ethanol, and tissue was stained by safranine and alcian blue.
2.2 Xylem tracing
Water transport through fresh fruit pedicel was investigated using eosin, water soluble xylem mobile dye which easily flows through vessels and pits and could be used as xylem tracer [16]. Pedicels were cut from plants, trimmed under water and the end near cut stem was immediately immersed in 1 ml 1% water solution of eosin for 15 minutes. After short rinsing, pedicels were cut transversally or longitudinally. Sections were investigated using Leica DMLS epifluorescent microscope (filter A 340-380nm). Xylem, thanks to presence of lignin, in UV light shows autofluorescence and emits blue light, but after eosin staining, fluorescence and emitted light is yellow. Since eosin flows only through functional xylem elements, comparing blue areas with yellow ones it is possible to get information about number of functional and non-functional xylem elements.
3. Fruit anatomy-developmental analysis
Tomato fruit consists of pericarp and seeds. Pericarp is composed from: egzocarp, mezocarp and endocarp. Outer layer of cells in the egzocarp is epidermis and below there are two to three layers of hypodermal cells with thick cell walls. Epidermis hasn’t stomata and has relatively thin cuticle, and the thickness of cuticle increases with the fruit growth. Mezocarp is made from large thin wall cells and vascular tissue. Fruit vascular tissue is connected to pedicel vascular tissue [17]. One vascular branch pass trough central and radial mezocarp to the seeds, while other vascular branches radialy pass through outer layer of mezocarp [9] parallel to fruit surface [18], with week branching on proximal side, but more on distal side with simultaneously decreasing ratio of xylem and increased ratio of phloem [19]. Endocarp is unicellular layer boundaring locular cavity. Carpelar septe divide ovarium into two or more loculi. Elongated central placenta, with attached seeds, is made of parenchima tissue and represents primary tissue which later fills the locular cavities. According to Gillaspy et al. [9] growth and development of tomato fruit could be divided into four phases. First phase is development of ovary and fertilisation. Immediately after fertilization, starts the second phase - in ovary starts cell divisions lasting 7-10 days, followed by tissue differentiation, seed development and early growth of embryo. Number of cell divisions and duration of this phase could differ among fruits and both factors determine final fruit size. After cell division phase, during 6-7 weeks fruit growth takes place due to increasing cell volume, until fruit reaches its final size. Although fruit growth depends on cell division as well as on cell growth, in most plants, increasing cell size has major role in determining final fruit size. In tomato, the size of placenta, locular tissue and mezocarp could be increased more than 10-fold [9] due to the cell elongation. At the phase when fruits reach their final size, fruits are green and have most of its final weight. The last phase starts after reaching final fruit size, and fruit development continues with ripening when inflow of carbohydrates in fruits stops [20] and fruit change colour from green to orange and finally red [21]. First change in colour is consequence of transformation of chloroplasts in chromoplasts, and decreasing chlorophyll concentration, followed with increasing in beta-carotene concentration which gives fruit orange colour. Final red colour is due to high concentration of lycopene [19]. Simultaneously with colour changes, metabolic changes occur. At this phase, disintegration of inner and central layer of carpel walls [18] occurs, and placental tissue is softening as a consequence of enzimatous degradation of cell walls and becomes gelatinous. Major biochemical events at this stage are: increase in ethylene synthesis, increase in respiration, fruit acidity rapidly increases and decreases after that, content of starch decreases and sugar content increases. Some other authors, contrary to Gillaspy et al. [9] distinguish three major periods in the growth of tomato fruit: cell division, growth phase and ripening [22]. Illustration of the anatomy of developing tomato fruit in both genotypes is presented at Figs. 1 and 2.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
852 ©FORMATEX 2010
______________________________________________
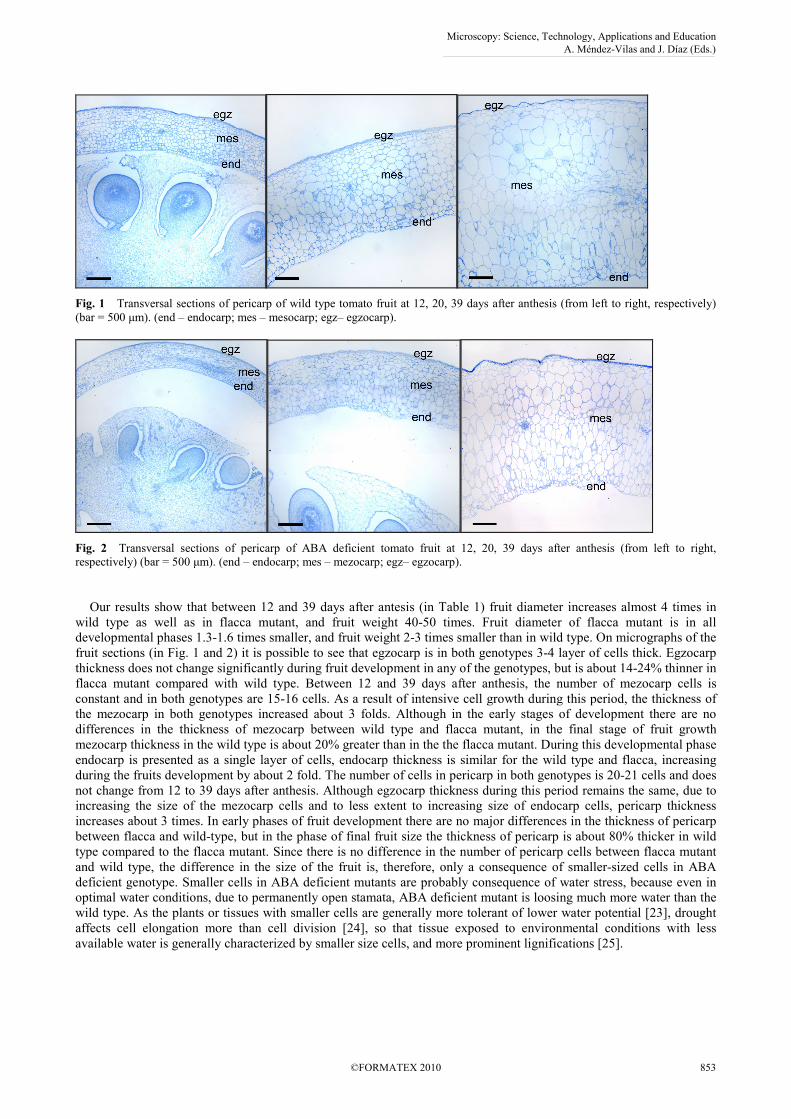
Fig. 1 Transversal sections of pericarp of wild type tomato fruit at 12, 20, 39 days after anthesis (from left to right, respectively) (bar = 500 µm). (end – endocarp; mes – mesocarp; egz– egzocarp).
Fig. 2 Transversal sections of pericarp of ABA deficient tomato fruit at 12, 20, 39 days after anthesis (from left to right, respectively) (bar = 500 µm). (end – endocarp; mes – mezocarp; egz– egzocarp). Our results show that between 12 and 39 days after antesis (in Table 1) fruit diameter increases almost 4 times in wild type as well as in flacca mutant, and fruit weight 40-50 times. Fruit diameter of flacca mutant is in all developmental phases 1.3-1.6 times smaller, and fruit weight 2-3 times smaller than in wild type. On micrographs of the fruit sections (in Fig. 1 and 2) it is possible to see that egzocarp is in both genotypes 3-4 layer of cells thick. Egzocarp thickness does not change significantly during fruit development in any of the genotypes, but is about 14-24% thinner in flacca mutant compared with wild type. Between 12 and 39 days after anthesis, the number of mezocarp cells is constant and in both genotypes are 15-16 cells. As a result of intensive cell growth during this period, the thickness of the mezocarp in both genotypes increased about 3 folds. Although in the early stages of development there are no differences in the thickness of mezocarp between wild type and flacca mutant, in the final stage of fruit growth mezocarp thickness in the wild type is about 20% greater than in the the flacca mutant. During this developmental phase endocarp is presented as a single layer of cells, endocarp thickness is similar for the wild type and flacca, increasing during the fruits development by about 2 fold. The number of cells in pericarp in both genotypes is 20-21 cells and does not change from 12 to 39 days after anthesis. Although egzocarp thickness during this period remains the same, due to increasing the size of the mezocarp cells and to less extent to increasing size of endocarp cells, pericarp thickness increases about 3 times. In early phases of fruit development there are no major differences in the thickness of pericarp between flacca and wild-type, but in the phase of final fruit size the thickness of pericarp is about 80% thicker in wild type compared to the flacca mutant. Since there is no difference in the number of pericarp cells between flacca mutant and wild type, the difference in the size of the fruit is, therefore, only a consequence of smaller-sized cells in ABA deficient genotype. Smaller cells in ABA deficient mutants are probably consequence of water stress, because even in optimal water conditions, due to permanently open stamata, ABA deficient mutant is loosing much more water than the wild type. As the plants or tissues with smaller cells are generally more tolerant of lower water potential [23], drought affects cell elongation more than cell division [24], so that tissue exposed to environmental conditions with less available water is generally characterized by smaller size cells, and more prominent lignifications [25].
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 853
______________________________________________
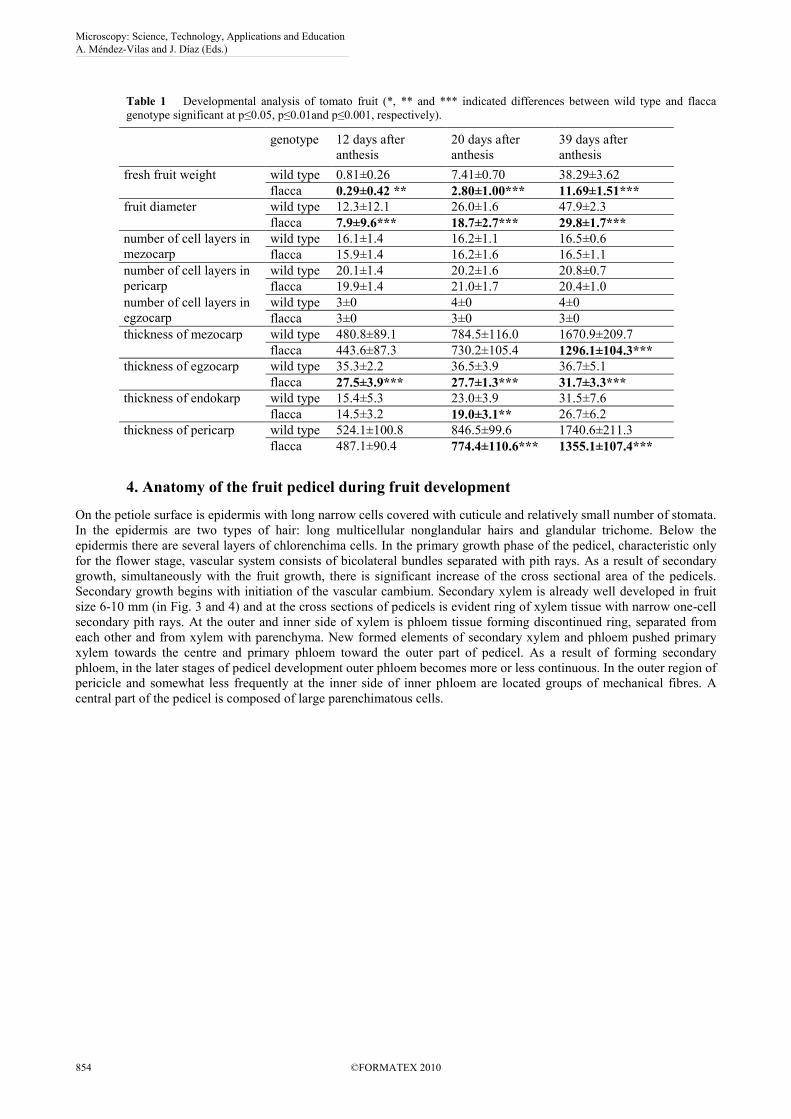
Table 1 Developmental analysis of tomato fruit (*, ** and *** indicated differences between wild type and flacca genotype significant at p≤0.05, p≤0.01and p≤0.001, respectively).
genotype 12 days after anthesis
20 days after anthesis
39 days after anthesis
fresh fruit weight wild type 0.81±0.26 7.41±0.70 38.29±3.62 flacca 0.29±0.42 ** 2.80±1.00*** 11.69±1.51***
fruit diameter wild type 12.3±12.1 26.0±1.6 47.9±2.3 flacca 7.9±9.6*** 18.7±2.7*** 29.8±1.7***
number of cell layers in mezocarp
wild type 16.1±1.4 16.2±1.1 16.5±0.6 flacca 15.9±1.4 16.2±1.6 16.5±1.1
number of cell layers in pericarp
wild type 20.1±1.4 20.2±1.6 20.8±0.7 flacca 19.9±1.4 21.0±1.7 20.4±1.0
number of cell layers in egzocarp
wild type 3±0 4±0 4±0 flacca 3±0 3±0 3±0
thickness of mezocarp wild type 480.8±89.1 784.5±116.0 1670.9±209.7 flacca 443.6±87.3 730.2±105.4 1296.1±104.3***
thickness of egzocarp wild type 35.3±2.2 36.5±3.9 36.7±5.1 flacca 27.5±3.9*** 27.7±1.3*** 31.7±3.3***
thickness of endokarp wild type 15.4±5.3 23.0±3.9 31.5±7.6 flacca 14.5±3.2 19.0±3.1** 26.7±6.2
thickness of pericarp wild type 524.1±100.8 846.5±99.6 1740.6±211.3 flacca 487.1±90.4 774.4±110.6*** 1355.1±107.4***
4. Anatomy of the fruit pedicel during fruit development
On the petiole surface is epidermis with long narrow cells covered with cuticule and relatively small number of stomata. In the epidermis are two types of hair: long multicellular nonglandular hairs and glandular trichome. Below the epidermis there are several layers of chlorenchima cells. In the primary growth phase of the pedicel, characteristic only for the flower stage, vascular system consists of bicolateral bundles separated with pith rays. As a result of secondary growth, simultaneously with the fruit growth, there is significant increase of the cross sectional area of the pedicels. Secondary growth begins with initiation of the vascular cambium. Secondary xylem is already well developed in fruit size 6-10 mm (in Fig. 3 and 4) and at the cross sections of pedicels is evident ring of xylem tissue with narrow one-cell secondary pith rays. At the outer and inner side of xylem is phloem tissue forming discontinued ring, separated from each other and from xylem with parenchyma. New formed elements of secondary xylem and phloem pushed primary xylem towards the centre and primary phloem toward the outer part of pedicel. As a result of forming secondary phloem, in the later stages of pedicel development outer phloem becomes more or less continuous. In the outer region of pericicle and somewhat less frequently at the inner side of inner phloem are located groups of mechanical fibres. A central part of the pedicel is composed of large parenchimatous cells.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
854 ©FORMATEX 2010
______________________________________________
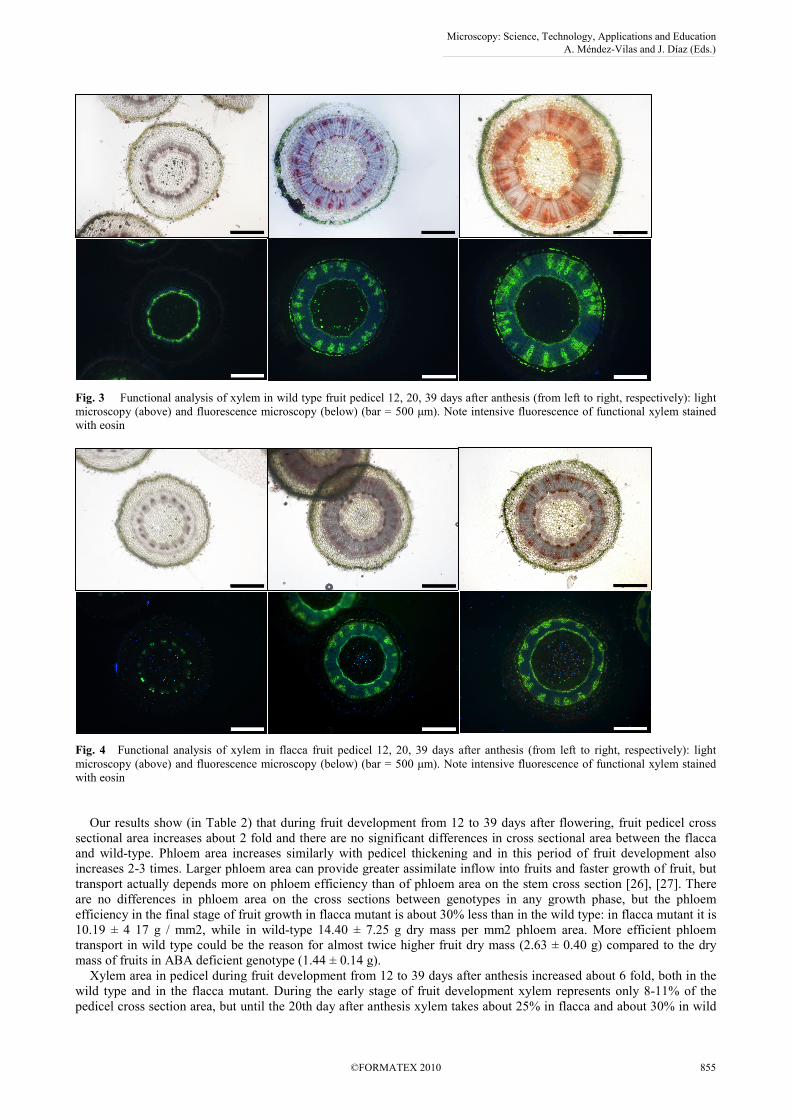
Fig. 3 Functional analysis of xylem in wild type fruit pedicel 12, 20, 39 days after anthesis (from left to right, respectively): light microscopy (above) and fluorescence microscopy (below) (bar = 500 µm). Note intensive fluorescence of functional xylem stained with eosin
Fig. 4 Functional analysis of xylem in flacca fruit pedicel 12, 20, 39 days after anthesis (from left to right, respectively): light microscopy (above) and fluorescence microscopy (below) (bar = 500 µm). Note intensive fluorescence of functional xylem stained with eosin Our results show (in Table 2) that during fruit development from 12 to 39 days after flowering, fruit pedicel cross sectional area increases about 2 fold and there are no significant differences in cross sectional area between the flacca and wild-type. Phloem area increases similarly with pedicel thickening and in this period of fruit development also increases 2-3 times. Larger phloem area can provide greater assimilate inflow into fruits and faster growth of fruit, but transport actually depends more on phloem efficiency than of phloem area on the stem cross section [26], [27]. There are no differences in phloem area on the cross sections between genotypes in any growth phase, but the phloem efficiency in the final stage of fruit growth in flacca mutant is about 30% less than in the wild type: in flacca mutant it is 10.19 ± 4 17 g / mm2, while in wild-type 14.40 ± 7.25 g dry mass per mm2 phloem area. More efficient phloem transport in wild type could be the reason for almost twice higher fruit dry mass (2.63 ± 0.40 g) compared to the dry mass of fruits in ABA deficient genotype (1.44 ± 0.14 g). Xylem area in pedicel during fruit development from 12 to 39 days after anthesis increased about 6 fold, both in the wild type and in the flacca mutant. During the early stage of fruit development xylem represents only 8-11% of the pedicel cross section area, but until the 20th day after anthesis xylem takes about 25% in flacca and about 30% in wild
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 855
______________________________________________
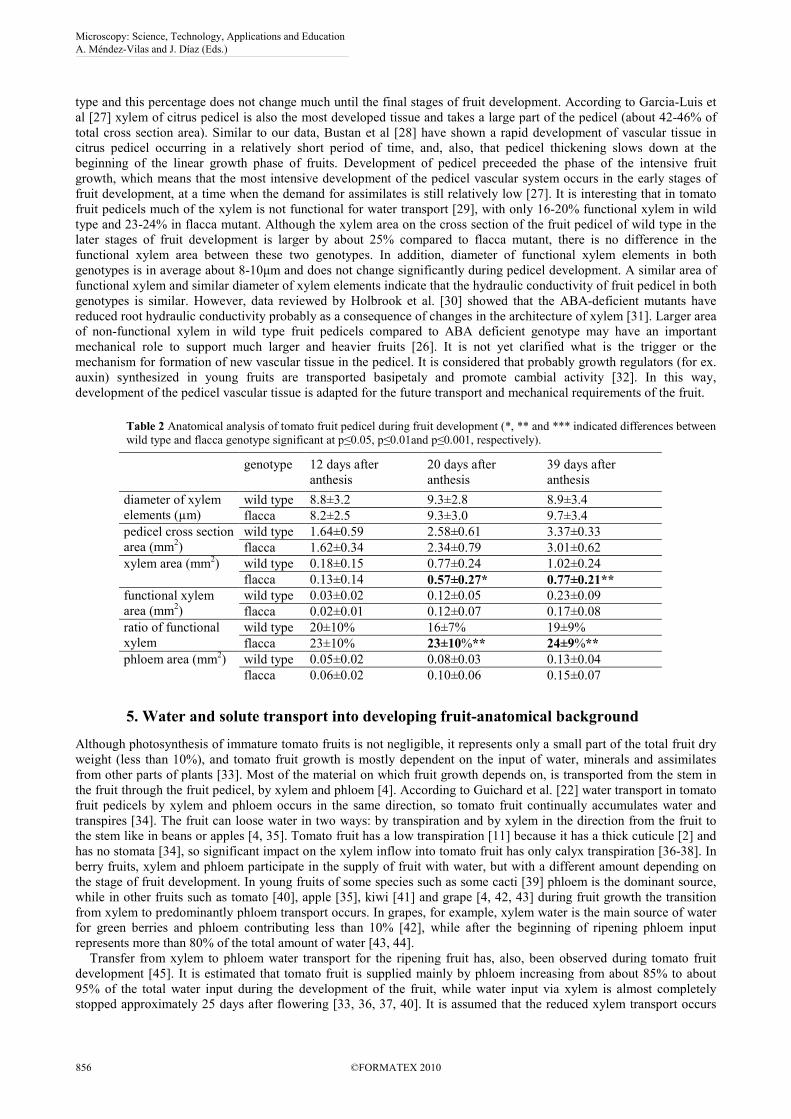
type and this percentage does not change much until the final stages of fruit development. According to Garcia-Luis et al [27] xylem of citrus pedicel is also the most developed tissue and takes a large part of the pedicel (about 42-46% of total cross section area). Similar to our data, Bustan et al [28] have shown a rapid development of vascular tissue in citrus pedicel occurring in a relatively short period of time, and, also, that pedicel thickening slows down at the beginning of the linear growth phase of fruits. Development of pedicel preceeded the phase of the intensive fruit growth, which means that the most intensive development of the pedicel vascular system occurs in the early stages of fruit development, at a time when the demand for assimilates is still relatively low [27]. It is interesting that in tomato fruit pedicels much of the xylem is not functional for water transport [29], with only 16-20% functional xylem in wild type and 23-24% in flacca mutant. Although the xylem area on the cross section of the fruit pedicel of wild type in the later stages of fruit development is larger by about 25% compared to flacca mutant, there is no difference in the functional xylem area between these two genotypes. In addition, diameter of functional xylem elements in both genotypes is in average about 8-10µm and does not change significantly during pedicel development. A similar area of functional xylem and similar diameter of xylem elements indicate that the hydraulic conductivity of fruit pedicel in both genotypes is similar. However, data reviewed by Holbrook et al. [30] showed that the ABA-deficient mutants have reduced root hydraulic conductivity probably as a consequence of changes in the architecture of xylem [31]. Larger area of non-functional xylem in wild type fruit pedicels compared to ABA deficient genotype may have an important mechanical role to support much larger and heavier fruits [26]. It is not yet clarified what is the trigger or the mechanism for formation of new vascular tissue in the pedicel. It is considered that probably growth regulators (for ex. auxin) synthesized in young fruits are transported basipetaly and promote cambial activity [32]. In this way, development of the pedicel vascular tissue is adapted for the future transport and mechanical requirements of the fruit.
Table 2 Anatomical analysis of tomato fruit pedicel during fruit development (*, ** and *** indicated differences between wild type and flacca genotype significant at p≤0.05, p≤0.01and p≤0.001, respectively).
genotype 12 days after anthesis
20 days after anthesis
39 days after anthesis
diameter of xylem elements (µm)
wild type 8.8±3.2 9.3±2.8 8.9±3.4 flacca 8.2±2.5 9.3±3.0 9.7±3.4
pedicel cross section area (mm2)
wild type 1.64±0.59 2.58±0.61 3.37±0.33 flacca 1.62±0.34 2.34±0.79 3.01±0.62
xylem area (mm2) wild type 0.18±0.15 0.77±0.24 1.02±0.24 flacca 0.13±0.14 0.57±0.27* 0.77±0.21**
functional xylem area (mm2)
wild type 0.03±0.02 0.12±0.05 0.23±0.09
flacca 0.02±0.01 0.12±0.07 0.17±0.08
ratio of functional xylem
wild type 20±10% 16±7% 19±9% flacca 23±10% 23±10%** 24±9%**
phloem area (mm2) wild type 0.05±0.02 0.08±0.03 0.13±0.04
flacca 0.06±0.02 0.10±0.06 0.15±0.07
5. Water and solute transport into developing fruit-anatomical background
Although photosynthesis of immature tomato fruits is not negligible, it represents only a small part of the total fruit dry weight (less than 10%), and tomato fruit growth is mostly dependent on the input of water, minerals and assimilates from other parts of plants [33]. Most of the material on which fruit growth depends on, is transported from the stem in the fruit through the fruit pedicel, by xylem and phloem [4]. According to Guichard et al. [22] water transport in tomato fruit pedicels by xylem and phloem occurs in the same direction, so tomato fruit continually accumulates water and transpires [34]. The fruit can loose water in two ways: by transpiration and by xylem in the direction from the fruit to the stem like in beans or apples [4, 35]. Tomato fruit has a low transpiration [11] because it has a thick cuticule [2] and has no stomata [34], so significant impact on the xylem inflow into tomato fruit has only calyx transpiration [36-38]. In berry fruits, xylem and phloem participate in the supply of fruit with water, but with a different amount depending on the stage of fruit development. In young fruits of some species such as some cacti [39] phloem is the dominant source, while in other fruits such as tomato [40], apple [35], kiwi [41] and grape [4, 42, 43] during fruit growth the transition from xylem to predominantly phloem transport occurs. In grapes, for example, xylem water is the main source of water for green berries and phloem contributing less than 10% [42], while after the beginning of ripening phloem input represents more than 80% of the total amount of water [43, 44]. Transfer from xylem to phloem water transport for the ripening fruit has, also, been observed during tomato fruit development [45]. It is estimated that tomato fruit is supplied mainly by phloem increasing from about 85% to about 95% of the total water input during the development of the fruit, while water input via xylem is almost completely stopped approximately 25 days after flowering [33, 36, 37, 40]. It is assumed that the reduced xylem transport occurs
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
856 ©FORMATEX 2010
______________________________________________

because somewhere along the way between the stem and the fruit there is a place with a high hydraulic resistance [33] - the abscission zone, often considered as the cause of the high hydraulic resistance for xylem water flow. Results of several anatomical studies of xylem in fruit pedicels have shown that there is reduced xylem area at, both, cross and longitudinal sections in the abscission zone in tomato pedicels [29, 46-48]. Near the abscission zone of tomato fruit ended more than 90% of vessels, and in addition there are a significantly reducing their diameter [33, 47]. According to Rancic et al [48] hydraulic resistance of the abscission zone, calculated based on anatomical sections, increases at least two orders of magnitude compared to the pedicel zone near the stem. Direct measurement of hydraulic conductivity [33] showed that in total xylem hydraulic resistance in tomato fruit pedicel, dominant role has hydraulic resistance in the abscission zone. In the studies of xylem in grape berry pedicels in experiments with dye infusion different authors have shown that dye inflow in mature grape berries is much less than in green ones. Before the stage of maturity, if the cut end of the grape pedicel were immersed in dye, the dye inflows in berries for about 30 minutes, crossing at the distance of 5-10cm, but after the beginning of ripening, the dye flows through the pedicel xylem to the fruit, but does not enter into the fruit, which led the general conclusion that the xylem in berries at the stage of maturation is non-functional. This is most often explained as a result of rapid growth and consequential breaking, stretching or damaging vessels [35, 42, 44, 49, 50, 51]. Phenomenon that after stopping transport water by xylem into fruit, transport by phloem still continues could be explained by the fact that phloem remains functional due to its structure. Phloem is, namely, composed of living cells [52] with elastic cell walls which could follow the growth without being damaged. Even in the case of damage, such damage may be easily exceeded by transport through simplast of parenhimatous cells [35]. However, experiments with visualization of the xylem transport demonstrated that vascular elements of xylem remain intact and apparently functional during the development of grape berries because apoplast dye may enter the pedicel through the central and peripheral vessels by the end of xylem ripening grape berries using the applied hydrostatic gradient, applying pressure or using a absorbing material on the part of the fruit contrary to the pedicel [53]. In addition, the switch in grape from phloem to xylem transport is fast, and occurs in 2 days [54], which is a relatively short period of time for major changes in the anatomical level, especially if they are a consequence of cell growth, so therefore, a simple anatomical explanation (such as xylem interruption) for change from xylem to phloem transport which appears in the phase of berry ripening is unlikely [4]. In our experiment with tomato, the dye is transported by xylem and passed through a few cm long pedicel relatively fastly reaching the opposite end of the pedicel (near the fruit) in 15-30 minutes. Similar results to ours in experiment with tomato were obtained by van Ieperen et al. [33]. In their experiment of functional vessel staining, the most intense colour was near the place where the dye is added to the petiole (part of the pedicel near the stem), while the pedicel near the fruit was less coloured, but there are still was a lot of vessels [33]. The experiment with dye infusion in tomato pedicel performed by Malone and Andrews [56] also showed that there is a continuous functional xylem along the pedicel and the pericarp and according to these authors, over 90% of hydraulic resistance between stem and fruit is in the fruit pericarp and not in the abscission zone. The results from our experiments also showed that in any stage of pedicel development there are no disruption of xylem transport since the data presented on table 2 show that functional xylem increased during fruit development in both genotypes. It should, also, be taken into account that the experiment was carried out with the pedicels that were cut off from the parent plants and all fruits were bigger than 0.5 cm when removed before immersion pedicel in staining solution. In these circumstances the only driving force for the xylem flow was transpiration of calyx as in the case of the removal of sepals transport was negligible. In the case when the fruit is on the pedicel, an additional driving force for the xylem flow from stem into the fruit is, besides sepal transpiration cuticular transpiration of the fruit. However because of thick tomato fruit cuticle [2] and the lack of stomata [34] tomato fruit has weak transpiration [11]. On the other hand, the presence of the fruit could possibly have a negative impact on the flow of water by xylem into fruit. In direct measurements of hydraulic conductivity of tomato pedicels Van Ieperen et al. [33] concluded that in the zones before and after abscission zone there are increased conductivity during fruit development which is in line with the formation of the new xylem elements during secondary growth. However, thanks to the high resistance in the abscission zone, due to the small xylem area and absence of secondary growth in this area [33, 46, 47], the total hydraulic resistance of the fruit pedicels between 11 and 31 days after anthesis is not changed and does not increase. It is known that in the pericarp of tomato fruit 13-14 days after flowering accumulates starch, and the rapid accumulation of hexose begins 23-25 days after flowering [45]. It is interesting that at the same time with the accumulation of hexose, approximately 25 days after anthesis fruit water intake by xylem almost completely stops [33, 36, 37, 40]. The facts that the hydraulic conductivity of pedicel does not change during the period from 11 to 35 days after anthesis [33], and that the stop of xylem transport occurs 25 days after anthesis [36, 37, 40], indicates that although the hydraulic resistance in the abscission zone is much less than in other parts of the pedicel, it is constant during fruit development and apparently did not cause interruption of water xylem transport. Rapid transition from significant to minimal contribution of xylem to the fruit water balance at the initial stages of fruit ripening phase in grape also coincides with an increased intake of sugar in the fruit [53, 43], while phloem input increases about 10 times [42]. Since our results have shown that in tomato fruit petiole from 12 to 39 days after anthesis there is no interruption in the xylem functionality, in a situation where the fruits are intact on the plant, the cessation of water xylem input into the fruit can be explained by the fact that there are flow through xylem but that it occurs in the reverse direction. Considering that the fruit is a plant organ with a
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 857
______________________________________________

great sink activity, it is possible that the excess of water which enters in fruit by phloem is forwarded by xylem into the stem. In this way the water that comes out from the fruit by xylem, prevents xylem transport in the direction toward the fruit. In line with this hypothesis are the data Ehret & Ho [36, 37] and Ho et al. [40] that significant xylem transport in tomato fruit occurs only at night when the phloem transport is on minimum [34]. Tomato fruit growth is also limited by the elasticity of cell walls of egzocarp [57]. Parenchymatous mezocarp cells which grow rapidly make pressure on egzocarp, and this is the reason why the mechanical properties of egzocarp are important in controlling growth of fruits. It is thought that enzymes such as peroxidase, which reduce the elasticity of cell walls, may play an important role in controlling growth of the fruit, since it has been observed that the cessation of growth of the fruit is associated with an increase in peroxidase activity in egzocarp [2, 58]. Increased phloem water intake may increase the pressure inside the fruit due to the limited extensibility egzocarp of fruits [57], which leads to the return of water by xylem especially if the excess of entered water by phloem exceeds the needs of berries for growth and transpiration [43]. Restoring water by xylem makes fruit less susceptible to plant water status [33] and can reduce their vulnerability to cracking as it serves as a overflow mechanism [59]. A similar conclusion gave Keller et al. [43] based on grape experiment. While in the grapevine there are apparently intact and functional xylem vessels, at the beginning of ripening grape berries there are significant reduction of water transported to the fruit through xylem comparing to the phloem [42, 53, 60]. Keller et al. [43] have explained this phenomenon by excising phloem water by xylem recirculation. It is known that the apoplast pressure in grapes fruits is higher than in plants [61], so the entry of water by xylem in the berries is probably inhibited by the positive pressure within the mature berries [43]. In accordance with this theory are data by Choat et al. [59] who made direct measurements of hydraulic conductivity of grapevine berry stems from 20-100 days after flowering. They found that indeed there is a significant increase in resistance to pedicel in the later stages of ripening (80-100 days after flowering), but this increase was not strong enough and occurs too late, that could explain the interruption of xylem transport that happens between 60-75 days after flowering. Malone and Andrews [56] have experimentally demonstrated the flow of fruit to the leaves, which also indicates the presence of functional xylem connection between stem and fruit. Although the use of xylem transport as a way for transport water into the fruit can be mush reduced, it does not necessarily mean that it is consequence of the physical blockade or other direct reduction of hydraulic conductivity of xylem, and it is more probably a consequence of the loss of the driving force (hydrostatic gradient) in berry apoplast. There are examples that, during development of the fruit, the relative importance of flow by xylem and phloem can be reversed and is probably more dependent on the sink/source relationships, and not on the physical loss of continuity or xylem conductivity [4]. In plum fruit, xylem entry is for example, dominant during phase II (mid-June), phloem transport becomes dominant at the start of phase III (early July), while later in the phase III (mid-July) intensity of both flows is similar, which again indicates the presence of functional xylem [4]. Temporary reduction in size of plant organs is a common occurrence during the drought and it reflects the water loss by transpiration directly or water divert from the plant organ to the rest of the plant by xylem [42]. During the day, conditions which enhance leaf transpiration led to decreased size of the immature fruit of grapes, apples and other species [35, 4]. In organs with low transpiration rate, like many fleshy fruits, fruit shrinkening during the day indicates that transport of water from the organ by xylem, exceeds water intake by phloem. Shrinkening of grape berries during the soil drying occurs in green immature fruits, but fruits that are in maturation phase are much less sensitive to plant water status [42]. Although this was interpreted as a break in functional xylem as a consequence of rapid growth [4, 35, 44, 50, 51] we believe that the insensitivity of ripening berries on the plant water deficit is actually evidence of strong phloem component. Tomato fruit does not exhibit or has very little variation in fruit size due to temporary lack of water in the stem [34, 36, 37]. Fruit shrinking is low even when the water potential gradient between fruit and stem apoplasts clearly favours the transport of water in the direction to the stem during the drought period [34]. Lack of fruits shrinkening during periods of drought does not necessarily imply lack of diverting water by xylem [33], but water intake by phloem may run contrary to gradient of water potential and could easily exceed the amount of water that is returned from the fruit in stem by xylem, resulting in a net import of water.
6. Role of ABA in fruit development
It is believed that ABA stimulates assimilate intake into fruit and accelerates the fruit maturation [62, 63]. Important role of ABA in the fruit sink activity is confirmed by our data showing the lower efficiency of phloem and almost twice lower dry weight of fruit of ABA deficient genotype in comparison with the wild type. It is possible that ABA acts by increasing sugar intake by phloem or releases glucose from stored carbohydrates [64]. Investigations based on ABA deficient mutants have indicated that ABA has little effect on long distance transport of photoassimilates, but has a role in regulating distribution of photoassimilates in the place of their accumulation [65]. Some authors believe that the ABA has a role in redirecting assimilates from phloem to the fruit by enhancing the sink activity of pericarp and locular tissue during the phase of the most rapid growth [65, 66]. Experiments with injection of egzogenous ABA in citrus fruits showed that ABA increased concentrations of glucose and fructose, but did not affect concentration of organic acids [64]. It is possible that ABA increases the accumulation of sugars in sink organs by activation of enzyme (acid invertase) which catalyzes the irreversible hydrolysis of sucrose into glucose and fructose [67]. Similarly, results of Cheikh and Brenner [68] indicate that the ABA can affect the accumulation of starch through the action on the activity
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
858 ©FORMATEX 2010
______________________________________________

of fructoso-1,6-biphosphatase [65]. The concentration of ABA in the pericarp of tomato in the later stages of fruit development in comparing with the initial value first decreases [9], and then rapidly increases [63] reaching the maximum five weeks after flowering, which coincides with the phase of final fruit growth [69]. Fruit maturation in apples and cherries correspond with increases concentration of endogenous ABA in fruits [70]. Since the greatest concentration of free ABA coincides with the phase of final fruit size and changing colour of fruits [62] implies that ABA is a regulator in ripening tomato fruits [63] and it is considered that ABA enables the initiation of maturation by increasing the fruit sensitivity to ethylene [71].
Acknowledgements We wish to thank the CM Rick Tomato Genetics Resource Center, USA for seeds used in this study and to Project CROPWAT for financial support. We also acknowledge invaluable help and technical support of Maja Terzic and Radenko Radosevic.
References
[1] Bussieres P. Potential Dry Matter and Water Import Rates in the Tomato Fruit in Relationship to Fruit Size. Annals of Botany 1993;72:63-72.
[2] Andrews J, Adams SR, Burton KS and Evered CE. Subcellular localization of peroxidase in tomato fruit skin and the possible implications for the regulation of fruit growth. Journal of Experimental Botany, 2002;53:2185-2191.
[3] Kulkarni M, Deshpande U. Comparative Studies in Stem Anatomy and Morphology in Relation to Drought Resistance in Tomato (Lycopersicon esculentum). Am. J. Plant Physiol. 2006;1:82-88.
[4] Matthews MA, KA Shackel. Growth and Water Transport in Fleshy Fruit. In, Vascular Transport in Plants, N.M. Holbrook and M.A. Zwieniecki (Eds.) 2005 pp 181-197.
[5] Luković J, Maksimović I, Zorić L, Nagl N, Perčić M, Polić D, Putnik-Delić M. Histological characteristics of sugar beet leaves potentially linked to drought tolerance. Industrial Crops and Products. 2009; 30:281–286.
[6] Stertz SC, Espirito Santo AP, Bona C, Freitas RJS. Comparative morphological analysis of cherry tomato fruits from three cropping systems. Scientia Agricola 2005; 62: 296-298.
[7] Lemaire-Chamley M, Petit J, Garcia V, Just D, Baldet P, Germain V, Fagard M, Mouassite M, Cheniclet C, and Rothan C. Changes in Transcriptional Profiles Are Associated with Early Fruit Tissue Specialization in Tomato, Plant Physiology. 2005;139:750–769.
[8] Xiao H, Radovich C, Welty N, Hsu J, Li D, Meulia T, van der Knaap E. Integration of tomato reproductive developmental landmarks and expression profiles, and the effect of SUN on fruit shape. BMC Plant Biology. 2009;9:49 doi:10.1186/1471-2229-9-49
[9] Gillaspy G, Ben-David H and Gruissem W. Fruits: a developmental perspective. Plant Cell 1993;5: 1439–1451. [10] Bertin N, Gautier H, Roche C. Number of cells in tomato fruit depending on fruit position and source–sink balance during
plant development. Plant Growth Regulation 36, 2002. 105–112. [11] Guichard S, Gary C, Leonardi C, and Bertin N. Analysis of Growth and Water Relations of Tomato Fruits in Relation to Air
Vapor Pressure Deficit and Plant Fruit Load. J Plant Growth Regul. 2005;24:201–213. [12] Jones HG, Sharp CS, Higgs KH. Growth and Water Relations of Wilty Mutants of Tomato (Lycopersicon esculentum Mill.)
Journal of Experimental Botany. 1987;38:1848-1856. [13] Liu Y, Bino RJ, Karssen CM, Hilhorst HMW. Water relations of GA- and ABA-deficient tomato mutants during seed and fruit
development and their influence on germination. Physiologia plantarum 2006;96:425 – 432. [14] Cornish K, Zeevaart JAD. Phenotypic Expression of Wild-Type Tomato and Three Wilty Mutants in Relation to Abscisic
Acid Accumulation in Roots and Leaflets of Reciprocal Grafts. Plant Physiology 1988; 87:190-194. [15] Neill SJ, Horgan R. Abscisic Acid Production and Water Relations in Wilty Tomato Mutants Subjected to Water Deficiency.
Journal of experimental botany. 1985;36:1222-1231. [16] Baum SF, PN Tran, WK Silk. Effects of salinity on xylem structure and water use in growing leaves of sorghum. New. Phytol.
2000;146:119-127. [17] Bussieres P. Water Import Rate in Tomato Fruit: A Resistance Model. Annals of Botany. 1994;73:75-82. [18] Roth I. Fruits of angiosperms 1977; Gebrüder Borntraeger, Berlin 675 pp. [19] Hayward HE. The structure of economic plants, 1938 New York, Macmillan, [20] Wang F, A Sanz, M Brenner, A Smith. Sucrose Synthase, Starch Accumulation, and Tomato Fruit Sink Strength. Plant
Physiology 1993;101:321-327. [21] Carrari F, Fernie AR. Metabolic regulation underlying tomato fruit development. Journal of Experimental Botany. 2006;57:
1883–1897. [22] Guichard S, Bertin N, Leonardi C, Gary C. Tomato fruit quality in relation to water and carbon fluxes Agronomie 2001;21:
385-392. [23] Cutler JM, Rains DW, Loomis RS. The importance of cell size in the water relations of plants. Physiol. Plant. 1977;40:255–
260. [24] Hsiao T. Plant responses to water stress. Annual Review of Plant Physiology 1973;24:519–80. [25] Kulkarni M, Deshpande U. In Vitro screening of tomato genotypes for drought resistance using polyethylene glycol. African
Journal of Biotechnology, 2007;6:691-696. [26] Zhang C, Tanabe K, Tamura F, Matsumoto K and Yoshida A. 13C-photosynthate accumulation in Japanese pear fruit during
the period of rapid fruit growth is limited by the sink strength of fruit rather than by the transport capacity of the pedicel. Journal of Experimental Botany. 2005;56:2713–2719.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 859
______________________________________________

[27] Garcia-Luis A, Oliveira MEM, Bordon Y, Siqueira DL, Tominaga S, Guardiola JL.Dry matter accumulation in citrus fruit is not limited by transport capacity of the pedicel. Annals of Botany 2002;90:755–764.
[28] Bustan A, Erner Y, Goldschmidt EE. Interactions between developing Citrus fruit and their supportive vascular system. Annals of Botany 1995;76:657–666.
[29] Rančić D, Pekić Quarrie S, Terzić M, Savić S, Stikić R Comparison of light and fluorescence microscopy for xylem analysis in tomato pedicels during fruit development. Journal of Microscopy. 2008;232: 618–622.
[30] Holbrook NM, Shashidhar VR, James RA, Munns R. Stomatal control in tomato with ABA-deficient roots: response of grafted plants to soil drying. Journal of Experimental Botany (2002) 53:1503–1514
[31] Hose E, Steudle E, Hartung W. Abscisic acid and hydraulic conductivity of maize root: a study using cell- and root-pressure probes. Planta 2000;211: 874–882.
[32] Sachs T. The control of patterned differentiation of vascular tissues Advances in Botanical Research 1981;9: 151-262. [33] van Ieperen W, Volkov VS and van Meeteren U. Distribution of xylem hydraulic resistance in fruiting truss of tomato
influenced by water stress. Journal of Experimental Botany. 2003;54:317-324 . [34] Johnson RW, Dixon MA, Lee DR. Water relations of the tomato during fruit growth Plant, Cell and Environment 1992;15:
947-953. [35] Lang A. Xylem, phloem and transpiration flows in developing apple fruits. Journal of Experimental Botany 1990;41:645–651 [36] Ehret DL, Ho LC. Effects of osmotic potential in nutrient solution on diurnal growth in tomato fruit. Journal of Experimental
Botany. 1986;37:1294-1302. [37] Ehret DL, Ho LC. Translocation of Ca in relation to tomato fruit growth. Annals of Botany 1986;58: 679-688. [38] Araki T, Kitano M, Eguchi H. Respiration, sap flux, water balance and expansive growth in tomato fruit. Biotronics 1997; 26:
95–102. [39] Nobel PS, De la Barrera E. Carbon and water balances for young fruits of platyopunitas. Physiologia Plantarum 2000;109:
160–166. [40] Ho LC, Grange RI, Picken AJ. An analysis of the accumulation of water and dry matter in tomato fruit. Plant, Cell and
Environment 1987;10:157–162. [41] Dichio B, Remorini D, Lang S. Developmental changes in xylem functionality in kiwifruit: implications for fruit calcium
accumulation. Acta Horticulturae 2003;610:191–195. [42] Greenspan MD, Shackel KA, Matthews MA. Developmental changes in the diurnal water budget of the grape berry exposed to
water deficits. Plant, Cell and Environment. 1994;17: 811–820. [43] Keller M, Smith JP and Bondada BR. Ripening grape berries remain hydraulically connected to the shoot. Journal of
Experimental Botany, 2006;57:2577–2587. [44] Creasy GL, Price SF, Lombard PB. Evidence for xylem discontinuity in Pinot noir and Merlot grapes: dye uptake and mineral
composition during berry maturation. American Journal of Enology and Viticulture 1993;44:187–192. [45] Ruan YL, Patrick JW. The cellular pathway of post phloem sugar transport in developing tomato fruit. Planta 1995;196: 434–
444. [46] Lee DR. Vasculature of the abscission zone of tomato fruit: implications for transport. Canadian Journal of Botany.
1989;67:1898–1902. [47] Andre´ JP, Catesson AM, Liberman M. Characters andorigin of vessels with heterogenous structure in leaf andflower
abscission zones. Canadian Journal of Botany 1999;77:253–261. [48] Rančić D, Pekić Quarrie S, Radošević R, Terzić M, Pećinar I , Stikić R, Jansen S. The application of various anatomical
techniques for studying the hydraulic network in tomato fruit pedicels. Protoplasma 2010;DOI 10.1007/s00709-010-0115-y. [49] Châtelet DS, Rost TL, Matthews MA and Shackel KA. The peripheral xylem of grapevine (Vitis vinifera) berries. 2. Anatomy
and development. Journal of Experimental Botany. 2008;59:1997–2007. [50] Findlay N, Oliver KJ, Nii N, Coombe BG. Solute accumulation by grape pericarp cells. IV. Perfusion of pericarp apoplast via
the pedicel and evidence for xylem malfunction in ripening berries. Journal of Experimental Botany. 1987;38:668–679. [51] Lang A, During H. Partitioning control by water potential gradient: evidence for compartmentation breakdown in grape
berries. Journal of Experimental Botany. 1991;42:1117–1122. [52] Esau K. Anatomy of seed plants, New York, USA. John Wiley and Sons 1977 [53] Bondada BR, Matthews MA and Shackel KA. Functional xylem in the post-veraison grape berry. Journal of Experimental
Botany. 2005; 56:2949–2957. [54] Düring, H, Lang A, Oggionni F. Patterns of water flow in Riesling berries in relation to developmental changes in their xylem
morphology. Vitis 1987;26:123-131. [56] Malone M, Andrews J. The distribution of xylem hydraulic resistance in the fruiting truss of tomato. Plant, Cell and
Environment. 2001;24:565–570. [57] Matthews MA, Cheng G, Weinbaum SA. Changes in water potential and dermal extensibility during grape berry development.
Journal of the American Society of Horticultural Science 1987;112:314–319. [58] Savić S, Stikić R, B Vucelić Radović, B Bogičević, Z Jovanović and V Hadži-Tašković Šukalović. Comparative effects of
regulated deficit irrigation (RDI) and partial root-zone drying (PRD) on growth and cell wall peroxidase activity in tomato fruits. Scientia Horticulturae 2008;117:15-20.
[59] Choat B, GA Gambetta, KA Shackel, MA Matthews. Vascular Function in Grape Berries across Development and Its Relevance to Apparent Hydraulic Isolation. Plant Physiology 2009;151:1677-1687
[60] Rogiers SY, Smith JA, White R, Keller M, Holzapfel BP, Virgona JM. Vascular function in berries of Vitis vinifera (L) cv. Shiraz. Australian Journal of Grape and Wine Research 2001;7: 46–51.
[61] Tyerman SD, Tilbrook J, Pardo C, Kotula L, Sullivan W, Steudle E. Direct measurements of hydraulic properties in developing berries of Vitis vinifera L. cv. Shiraz and Chardonnay. Australian Journal of Grape and Wine Research 2004;10:170–181.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
860 ©FORMATEX 2010
______________________________________________

[62] Kojima K, S Kuraishi, N Sakurai, K Fusao. Distribution of abscisic acid in different parts of the reproductive organs of tomato. Scientia Hortic. 1993;56: 23-30.
[63] Buta JG, Spaulding DW. Changes in indole-3-acetic acid and abscisic acid levels during tomato (Lycopersicon esculentum Mill.) fruit development and ripening. Journal of Plant Growth Regulation. 1994;13:163-166.
[64] Kojima K, Yamada Y, Yamamoto M. Effectso f Abscisic Acid Injection on Sugar and Organic Acid Contents of Citrus Fruit. J.Japan Soc.Hort. Sci. 1995;64:17-21.
[65] Smith GS, Klages KU, Green TGA, Walton EF. Changes in abscisic acid concentration, surface conductance, and water content of developing kiwifruit. Scientia Horticulturae 1995;62:13-27.
[66] Kojima K. Simultaneous measurement of ABA, IAA and GAs in citrus: Role of ABA in relation to sink ability. Plant physiology and biochemistry 1995;29:179-185.
[67] Pan Q-H, Li M-J, Peng C-C, Zhang N, Zou X, Zou K-Q, Wang X-L, Yu X-C, Wang X-F and Zhang D-P. Abscisic acid activates acid invertases in developing grape berry. Physiologia Plantarum 2005;125:157–170.
[68] Cheikh N, Brenner ML. Regulation of Key Enzymes of Sucrose Biosynthesis in Soybean Leaves. Plant Physiol. 1992; 100:1230-1237.
[69] Abdel-Rahman M, Thomas TH, Dossand GJ, Howell L.Changes in endogenous plant hormones in cherry tomato fruit during development and maturation. Physiologia plantarum 1975; 34:39-43.
[70] Setha S, Kondo S, Hirai N, Ohigashi H. Xanthoxin, abscisic acid and its metabolite levels associated with apple fruit development. Plant Science 2004;166:493–499.
[71] Jiang Y, DC Joyce, AJ Macnish. Effect of Abscisic Acid on Banana Fruit Ripening in Relation to the Role of Ethylene. Journal of Plant Growth Regulation
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 861
______________________________________________