analyst - polytechnique montréal | polytechnique montréal est … · 2014-07-09 · additionally,...
TRANSCRIPT

Analyst
MINIREVIEW
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article OnlineView Journal | View Issue
Bacteriophages:
NitMMEtB(edba
aRegroupement Quebecois de Materiaux de P
Ecole Polytechnique de Montreal, Case
Montreal, Quebec H3C 3A7, Canada. E-maibBiophage Pharma, 6100 Royalmount, MontcLaser Processing and Plasmonics Laborat
Ecole Polytechnique de Montreal, Case
Montreal, Quebec H3C 3A7, Canada
Cite this: Analyst, 2014, 139, 1224
Received 22nd October 2013Accepted 8th January 2014
DOI: 10.1039/c3an01989f
www.rsc.org/analyst
1224 | Analyst, 2014, 139, 1224–1236
biosensing tools for multi-drugresistant pathogens
N. Tawil,*abc E. Sacher,a R. Mandevilleb and M. Meunierc
Pathogen detection is of utmost importance in many sectors, such as in the food industry, environmental
quality control, clinical diagnostics, bio-defence and counter-terrorism. Failure to appropriately, and
specifically, detect pathogenic bacteria can lead to serious consequences, and may ultimately be lethal.
Public safety, new legislation, recent outbreaks in food contamination, and the ever-increasing
prevalence of multidrug-resistant infections have fostered a worldwide research effort targeting novel
biosensing strategies. This review concerns phage-based analytical and biosensing methods targeted
towards theranostic applications. We discuss and review phage-based assays, notably phage
amplification, reporter phage, phage lysis, and bioluminescence assays for the detection of bacterial
species, as well as phage-based biosensors, including optical (comprising SPR sensors and fiber optic
assays), electrochemical (comprising amperometric, potentiometric, and impedimetric sensors), acoustic
wave and magnetoelastic sensors.
1. Introduction to virulent pathogenicbacteria
Bacterial pathogens have developed several mechanisms thataid in circumventing the host defences. The pathogenicity andvirulence of a microorganism are highly complex and are gov-erned by the bacterium–host interactions. Classical bacterialvirulence factors include toxins and molecules essential for
ancy Tawil obtained her Ph.D.n Biomedical Engineering fromhe Ecole Polytechnique deontreal in 2013. She holds aaster's degree in Chemicalngineering (Ecole Poly-echnique de Montreal) and aachelor's degree in PhysiologyMcGill University). Her inter-sts include biosensors, drug-elivery systems, systemsiology, phage therapy andntibiotics.
ointe, Department of Engineering Physics,
Postale 6079, succursale Centre-ville,
real, Quebec H4P 2R2, Canada
ory, Department of Engineering Physics,
Postale 6079, succursale Centre-ville,
adhering and invading the host, as well as evading the host'sdetection mechanisms and immune response.
Classically, these virulent bacteria have been treated withantibiotics since the discovery of penicillin in 1928.1 Majormechanisms of antibiotic action include the inhibition of thesynthesis of DNA (quinolones), RNA (rifamycins), and cellwalls (b-lactams and glycopeptides) or proteins (macrolides,lincosamides, streptogramins, amphenicols, oxazolidinones,tetracyclines, aminocyclitols and aminoglycosides).2 Quino-lones interfere with DNA replication by introducing DNAbreaks and trapping topoisomerase II (for Gram-negativebacteria) and topoisomerase IV (primarily in Gram-positivebacteria) at the cleavage stage, thus compromising strandrejoining.2 Similarly, rifamycins inhibit DNA transcription bybinding to the b-subunit of an active RNA polymerase.
Edward Sacher is a Professor inthe Department of EngineeringPhysics at the Ecole Poly-technique de Montreal. He is aFellow of the Royal Society ofChemistry (London) and theIEEE. His research focuses onbiomedical engineering andapplied physics, with an interestin surfaces and interfaces. He isalso the founder and Co-Directorof the Surface Analysis Labora-tory at the Ecole Polytechnique.
This journal is © The Royal Society of Chemistry 2014

Minireview Analyst
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
Additionally, bactericidal activity can be achieved by inhibi-tion of cell wall synthesis.
Bacterial cell walls are composed of peptidoglycan layers thatare extended by transglycosylases and cross-linked by penicillin-binding proteins (PBPs).3 b-lactams bind to the active site ofPBP and inhibit the formation of the peptide bond that cross-links peptidoglycan subunits, inducing changes in cell-wallmorphology and subsequent bacterial death. Moreover, abroad-range of drugs inhibits protein synthesis by affectingeither the 50S or the 30S ribonucleoprotein subunits. Drugs thatinhibit the 50S ribosome subunit either block the initiation ofprotein transcription or the elongation of the nascent peptidechain. Furthermore, 30S inhibitors either prevent aminoacyltRNA from binding to the ribosome or induce tRNA mis-matching, either of which result in protein mistranslation.2
Although the development of antibiotics stands as one of themost substantial medical innovations, the ever-increasingprevalence of multidrug-resistant infections has created aglobal public health crisis.4 In the United States, excess healthcosts attributable to antibiotic-resistant infections annuallyreach $20 billion, with 8 million days of additional hospitaloccupancy, and $35 billion in societal costs.5
Antibiotic resistance can be acquired through severalmechanisms, including mutations due to selective pressure andhorizontal transfer of genetic material through transformation,conjugation and transduction. These genes include a variety ofenzymes and efflux pumps that are detrimental to the antibac-terial agent. Moreover, these genes can alter the metabolicpathway, resulting in the production of altered cell-wallproteins that evade antibiotic targeting.
Although substantial efforts are directed toward producingantibiotic and non-antibiotic derivatives, such as antibacterialvaccines, phage therapy, immunostimulants, adjuvants, pro-biotics, and antivirulence therapies, the gold standard forbacterial detection remains traditional culture-based assays,which are cumbersome and time-consuming. Other conven-tional techniques, such as the polymerase chain reaction (PCR),immunological assays, uorescence and spectrophotometry,
Rosemonde Mandeville studiedmedicine at the University ofAlexandria and holds a doctoraldegree in Medical Microbiologyfrom the University of Manitoba.She is the President and ChiefScientic Officer of BiophagePharma, Inc., whose main focusis the development of phage-based therapeutical approachesfor biomedical and environ-mental applications. Dr Mande-ville is a board member ofseveral companies and therecipient of many prestigiousawards.
This journal is © The Royal Society of Chemistry 2014
suffer from several drawbacks: they are time-consuming,expensive, laborious, and require enrichment steps and highlytrained personnel. There is, therefore, an increasing need formethods that eliminate the necessity for microbial cultures,reduce the time of diagnosis, and increase the sensitivity andspecicity of detection.
The advent of biosensors has permittedmajor advancementsin microbial detection. Depending on their transductionelements, these can be classied as optical, electrochemical oracoustical. Moreover, they can be classied according to theirbiorecognition elements. Most biosensors rely on the use ofantibodies. However, polyclonal antibodies suffer from highcosts of production and environmental instability, thus limitingtheir shelf-life, storage and applicability in non-regulated envi-ronments such as humid and high temperature climates.Moreover, polyclonal antibodies have the ability to recognizedifferent epitopes present on the same pathogen or on relatednon-pathogenic organisms, thus hindering their selectivity.6 Onthe other hand, monoclonal antibodies can be produced morerapidly, and are selective, but their production is still costly andthey are fragile to environmental uctuations, such as temper-ature, pH, ionic strength, and cross-reactivity.
We now turn our attention to bacteriophages, which are akey factor in microbial homeostasis on Earth, outnumberingtheir bacterial hosts by tenfold. Throughout history, bacterialpathogens have developed multiple strategies to combatviruses, which were circumvented by the ability of phages toevolve and survive in adverse environments. The specicity ofbacteriophages to their bacterial hosts, robustness, low-costand ease of production make them ideal recognition elementsfor bacterial biodetection purposes.
Here we delineate the clinically relevant bacterial patho-gens, of worldwide concern, that are targeted by today's bio-detection research and development efforts. We then describebacteriophages, their mechanisms of action, and relevance intherapeutics and diagnostics. We nally review phage-basedanalytical and biosensing methods, notably phage-amplication,reporter phages, phage lysis, bioluminescence assays, as well as
Michel Meunier is a Professor inthe Department of EngineeringPhysics at the Ecole Poly-technique de Montreal and aFellow of the Optical Society ofAmerica, SPIE, and the Cana-dian Academy of Engineering.He is the Director of the LaserProcessing and PlasmonicsLaboratory (LP2L) and theholder of the Canada ResearchChair in Materials Micro/Nano-engineering Using Lasers.
Analyst, 2014, 139, 1224–1236 | 1225

Analyst Minireview
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
the novel optical, electrochemical and mass-sensitive biosensortechnologies using bacteriophages as a recognition element.
2. Clinically relevant bacterialpathogens
Microorganisms are an intrinsic part of our environment,and play a fundamental role in human hemostasis. Problemsarise when immunocompromised patients come into contactwith certain pathogens that have developed resistance toantimicrobial agent in our health care system. The mostpredominant nosocomial agents are Enterococcus faecium,Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacterbaumannii, Pseudomonas aeruginosa, and Enterobacter species,commonly known as “the ESKAPE bugs”.7 In the followingsection, we briey describe some of these bacteria, as well asother virulent bacteria that cause major problems in hospitalsin the developed and developing countries. Most of thesebacteria have been targeted in biosensing technologiesdepicted in this review.
2.1. Staphylococcus aureus: the case of the golden killerbacterium
S. aureus is a ubiquitous Gram-positive microorganism thatcolonizes the nasal passages and skin of approximately 50% ofhealthy individuals.8 S. aureus can cause a broad spectrum ofinfections, ranging from common skin infections to sepsis. Itis the leading cause of nosocomial pneumonia, bloodstreaminfection, and infections caused by surgical wounds andprosthetic implants.9,10 S. aureus has quickly acquired resis-tance against b-lactam antibiotics by producing a specicpenicillinase enzyme11 encoded by a plasmid. This promptedthe introduction, in 1959, of methicillin, a modied penicillinthat was unaffected by the staphylococcal penicillinase.10 Itssuccess was, unfortunately, short-lived. b-lactams, aresubstrate analogs of a penicillin-binding protein, which, aspreviously mentioned, catalyzes the formation of peptidecrosslinks between glycan chains of the bacterial cell wall.This time, S. aureus acquired a mecA gene that encodes PBP2a,that confers a broad spectrum of resistance against methi-cillin, as well as all other b-lactams (i.e. synthetic penicillins,cephalosporins and carbapenems).11 Recently, strains resis-tant to glycopeptides, such as vancomycin, have emerged andacquired a vanA gene cluster.12 These strains are very hard todetect and are oen associated with therapeutic failure. MRSAhas become an increasingly alarming problem, as it is nolonger contained in hospital settings, but has emerged intothe community, and is now pandemic. According the Centersfor Disease Control and Prevention (CDC), more patients diedfrom MRSA-related infections in the United States, in 2005,13
than did from acquired immunodeciency syndrome (AIDS)complications.14
2.2. Y. Pestis: from “black death” to biological warfare
Yersinia pestis is the causative organism of plague, which isoen thought about as an ancient disease, but which is, to this
1226 | Analyst, 2014, 139, 1224–1236
day, endemic in parts of Asia, Africa, and America, causing areported three thousand deaths annually.15 Y. pestis is a Gram-negative bacterium of the Enterobacteriaceae family. It infectshuman macrophages, which are transported to draining lymphnodes. It becomes extracellular upon apoptosis of the infectedhost macrophage, colonizing major organs and causingsystemic failure, leading to death.16 The lack of proper diag-nostic tools that can effectively and rapidly detect this pathogenhas contributed to underreporting of cases and major short-comings in case-management and surveillance.15
2.3. E. coli O157:H7: an emerging lethal food-bornepathogen
Enterohemorrhagic E. coli is a zoonotic life-threatening bacte-rium that was rst described in 1982, and leads to diarrhea,hemorrhagic colitis and hemolytic-uremic syndrome inhumans. O157:H7 is a pathogenic subset of the Shiga toxin-producing E. coli (STEC) that is mainly transmitted byconsumption of undercooked meat, unpasteurized dairy prod-ucts, and produce contaminated with bovine feces. Detectionand control of these pathogens represent a challengingproblem, with regard to their high virulence and the extremelylow infectious dose required to induce disease.17
2.4. Salmonella: a non-negligible threat
Salmonella bacteria colonize a broad spectrum of host organ-isms, efficiently infecting both animals and plants. Among theSalmonella species, Salmonella enterica is the primary cause offood-borne poisoning in humans, contaminating a wide rangeof produce, such as pre-cut meats, poultry, eggs, vine vegeta-bles, fruits, nuts, sprouts, leafy greens, roots and beans.18 TheCDC reports that approximately 15% of United States citizenswill become infected with food-borne pathogens, with salmo-nellosis ultimately responsible for the largest number ofhospitalizations and related deaths.19
2.5. Listeria monocytogenes: an emerging pathogen
L. monocytogenes is a Gram-positive rod responsible for listeri-osis, a u-like disease, inducing gastroenteritis and fever inhealthy individuals. However, listeriosis can be deadly, withsymptoms ranging from septicemia, meningitis or spontaneousabortion, in immune-compromised patients, children, theelderly, and pregnant women.20 Listeria is one of the leadingemerging pathogens associated with food-borne diseases, withan estimated 1600 cases yearly in the United States.21
2.6. M. tuberculosis: the modern “white plague”
Tuberculosis (TB) is a disease caused by Mycobacterium tubercu-losis, and is one of themost destructive bacterial diseases to afflicthumans. The World Health Organization reports that approxi-mately 33% of mankind is latently infected with TB, with 1.8million annual deaths and 9.8 million new infections every year.
Following the inhalation of the bacilli, the bacteria depositin the lung alveolar space, and multiply in the alveolar macro-phages. They are then carried to the lymph nodes by dendritic
This journal is © The Royal Society of Chemistry 2014

Fig. 1 TEM photomicrograph of bacteriophages adhering specificallyto the MRSA bacterium.
Minireview Analyst
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
cells (DCs), which undergo programmed cell death and releasemycobacterial antigenic peptides. These antigens activate T-cells that will proliferate and become effector T cells, whichreturn to the lungs to ght the infection. This is followed by thegeneration of granuloma, which can ultimately contain theinfection. Many factors, such as a poor diet, immunosuppres-sion, steroid use and HIV therapy, can reactivate the latent stateof the infection and cause bacteria to become metabolicallyactive and reactivate the disease.
Drug-susceptible TB can be successfully treated by dailyadministrating a regimen of drugs to the patient, for an initialtreatment phase of two months, followed by a continuationphase ranging from four to seven months.22 However, majorcomplications arose following the emergence of multidrug-resistant and extensively drug-resistant strains, around theworld. As with MRSA, there are very few compounds in clinicaldevelopment for anti-TB drugs; thus, fast detection becomesparamount for a better management of the disease.
3. The potent bacteriophage
Bacteriophages serve as the natural counterbalance to bacteria,and have become the most ubiquitous source of life on earth(1030–1032 phage species). They are commonly found in humanintestines, having been consumed by humans via foods (e.g.,ground beef, pork sausage, chicken, farmed freshwater sh, oilsardines, raw skim milk, cheese, and drinking water),23 and arewidely distributed in our ecosystem.
Modern phage research was initiated by Frederick W. Twortin 191523,24 and carried on by Felix d'Herelle, in 1917,25 whouncovered the natural ability of the phages to combat bacterialpathogens and prevent infections.26–28 Bacteriophages areviruses that infect both Gram-negative and -positive bacteria byusing their pili as receptors. Phages are parasites that use theirbacterial host machinery to direct their replication. Each phageparticle is constituted of genomic material enclosed in a proteinor lipoprotein coat, called a capsid.23 Phages are subdivided inmultiple genera, based on their morphology, nucleic acidhomology and serology, and two groups, based on their lifecycles, replication, and propagation.29 Lytic bacteriophages arevirulent phages that specically recognize and infect their hostbacterium, use the bacterial machinery to replicate theirgenomes, produce and assemble their structural components,and ultimately lyse and kill the bacterium to release the newlyassembled virions.
Temperate phages differ from lytic phages as they are capableof both lytic and lysogenic propagation. These phages have thecapacity of incorporating their genomic material into bacterialchromosomes and assuming a quiescent state (prophage), whichallows both the bacterium and the prophage to co-exist for aperiod of time without lysis,29 shielding the host bacterium fromfurther phagic infection, and introducing novel genetic materialsuch as resistance genes and restriction systems.23
Currently, phages are grouped in nineteen families, of which twopossess an RNA genome and ve are enveloped. Tailed phages, ofthe order Caudovirales, account for approximately 95% of present-day isolated bacteriophages. The virions have an icosahedral head,
This journal is © The Royal Society of Chemistry 2014
composed of repeating units of one or two proteins, which enclosethe phage's genomic double-stranded DNA.23 Tailed phages aregrouped according to their morphology: those possessing a longexible tail belong to the Siphoviridae, while the Myoviridaecomprise those with contractile tails, and the Podoviridea arephages with very small non-contractile tail.23
Ten more families account for a vast variety of taillessphages, which are polymorphic in shape, may have a lipid coat,and differ in their genomic material, with the nucleocapsidcontaining ssDNA, dsDNA or RNA. Moreover, these phages candiffer on the basis of their means of propagation, and bywhether they are segmented or not.23 The difference betweentailed and tailless phages is of major importance for hostrecognition and adsorption. The interaction between the phageand the receptor of the bacteria is an on-and-off process, ulti-mately followed by the irreversible binding of the phage to itshost.30 Tailed-phages specically recognize their bacterial hoststhrough the interaction of a single central ber called adhesin,or a cluster of 3, 6 or 12 bers found on their caudal extremities.These caudal proteins will recognise and adhere to specicepitopes on their host membrane, and achieve irreversiblebinding through the interaction of a secondary tail protein toanother bacterial membrane receptor.
Some types of bacteriophages, such as P22, have short tailbers, called tail spike proteins, which recognize and bind thehost bacteria. Furthermore, the phage tail oen exhibits enzy-matic activity capable of locally breaking down the cell wall.Filamentous phages, on the other hand, consist of a longstructure made of repeating pVIII proteins. The length of thephage is proportional to the number of nucleotides in itsgenome (there are 0.435 pVIII proteins per nucleotide, whichcorresponds to 0.1435 nm per nucleotide).31
Fig. 1 shows a transmission electron microscope (TEM)photomicrograph of bacteriophages specically interactingwith a MRSA bacterium.
Analyst, 2014, 139, 1224–1236 | 1227

Analyst Minireview
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
Their specicity, ability to persist in a vast range of envi-ronmental conditions, and ease of production in suitable hosts,make bacteriophages critical vehicles for the coevolution andhorizontal gene transfer between different bacterial species.23
Moreover, their many advantageous properties make them animportant tool for the diagnosis and the control (preventionand therapy) of bacterial diseases.
Their therapeutic potential for the treatment of bacterialinfections in humans was recognized in the early 1920s with thepioneering work of d'Herelle, who successfully treated patientsinfected with the bubonic plague while stationed in Egypt, andpatients suffering from cholera in India.23 However, phageresearch came to a standstill with the advent of antibiotics andtheir mass production during World War II. Recent interest inbacteriophages was brought about by the alarming incidence ofmulti-drug resistant pathogens. Today, the commercial andmedical opportunities of phage-based products are vast owingto their benecial properties. The characterization of novelphages targeting specic bacterial pathogens is achieved muchfaster, and at a lower cost, than with antibiotics. Moreover, theirspecicity permits targeted action against their host, withminimal effects on the otherwise desirable microbial ora, andtarget problematic bacterial states such as biolms andpersister cells.32,33 The ability of phages to distinguish betweenlive and dead cells aids in overcoming major limitations ofroutinely used techniques for microbial diagnostics, such asPCR and hybridization-based assays. Finally, bioengineeredphages with novel functionalities34,35 and phage-functionalizednanoparticles36 can be used for sensing, as well as other relevanttechniques, such as molecular imaging.
4. Phage-based assays
Since their discovery by Twort and d'Herelle, bacteriophageshave been extensively exploited to specically target and iden-tify bacteria. To this day, phage typing is used to differentiatebetween diverse strains of particular species of bacteria. Phagetyping exploits their ability to specically recognize, infect and,ultimately, lyse their host. However, characterization andepidemiological identication of bacteria, using this method,
Table 1 Phage-based amplification assays for the detection of bacterial
Bacterial Species Phage
E. coli O157:H7 LG1E. coli O157:H7 AR1E. coli MS2Salmonella P22Salmonella SJ2Salmonella enterica SJ2Mycobacterium avium paratuberculosis D29M. tuberculosis D29Y. pestis FA1122Y. pestis FA11221.1.1.1.1.1.1.1 Y. pestis L-413CPseudomonas aeruginosa NCIMB 10116Salmonella Felix-01S. aureus N-15
1228 | Analyst, 2014, 139, 1224–1236
requires the maintenance of a large phage library, thusrestricting its use to a limited number of reference laboratories.Therefore, there is a paramount need for phage-based tech-nologies that can be used for the detection of pathogenicbacteria in a clinical setting. Three main methods exploitingbacteriophages are used to detect clinically relevant bacterialpathogens: (1) bacterial amplication of phages followed by thedetection of progeny phages, (2) detection of light produced byreporter phages or uorescent reporter phages followinginfection of bacteria, and (3) detection of intracellular compo-nents following infection and lysis of the bacterial host.
4.1. Phage amplication assays
Lytic phage propagation consists of adhesion to the bacterialhost, transfection and replication of the phage genome, fol-lowed by the assembly of the virion structural components. Thisculminates in the release of the progeny phages, as a result ofbacterial cell lysis. The increased number of phages in solutioncan be used as a marker of the specic infection and phage lyticpropagation in a target bacterium. In the past, this techniquewas hampered by the necessity of a large bacterial titer, as wellas the cost and complexity of identifying the progeny phageswith high-performance liquid chromatography (HPLC). Noveladvances in phage amplication assays have obviated the needfor complicated instruments, and require a low abundance ofinitial target pathogens. Phages are incubated with their targetbacteria, and phagicidal reagent is added before lysis can occur,to inactivate free phages. The virucide is then neutralized andhelper cells, susceptible to be infected by the progeny phages,are added to the solution and plated onto a so agar overlay.The phages are allowed to replicate within the previously infectedbacterial pathogens and can, in turn, infect the helper cells,resulting in the formation of lysis plaques. This technique hasthe advantages of being able to detect initially low concentrationsof bacteria, as well as target bacteria in a heterogeneous bacterialsample. Diagnostic kits and novel assays based on phageamplication are available for M. tuberculosis,37–42 Staphylococcusaureus,43–45 and Yersinia Pestis.38,46 Table 1 lists phage-basedamplication assays for the detection of a large number ofbacterial species. Phage amplication assays, combined with the
species
Time of detection Sensitivity Ref.
23 h 2 CFU per 25 g 55, 563.5 h 1 CFU mL�1 56, 572 h 104 CFU mL�1 584 h 104 CFU g�1 5920 h 3 CFU per 25 g 555 h 104 CFU mL�1 6048 h 10 CFU mL�1 6148 h 102 CFU mL�1 615 h 106 CFU mL�1 624 h 103 CFU mL�1 624 h 105 CFU mL�1 384 h 10 CFU mL�1 434 h 102 CFU mL�1 435 h 105 CFU mL�1 44
This journal is © The Royal Society of Chemistry 2014

Minireview Analyst
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
addition of antibiotics, can help distinguish between resistantand susceptible strains, such as antibiotic resistant M. tubercu-losis47–50 and MRSA.51 Moreover, phage typing is oen combinedwith other techniques, such as PCR,52,53 mass spectrometry44,46
and competitive ELISA.54
4.2. Reporter phage assays
It is possible to engineer recombinant phages for pathogendetection. This can be done by inserting the reporter gene intothe target cell and monitoring the production of the proteinencoded by the inserted genomic material. The method of geneinsertion into the phage varies considerably. Cloning can onlybe used with rather small phage genomes and is made difficultby the need of customized genetic manipulation, systems andnucleic acid packaging. An easier, more efficient and cost-effective method is the insertion of the reporter gene into theintergenic parts of the phage genome, resulting in multiple anddiverse particles that can later be selected, depending on thedesired properties. Cloning of the reporter phage into a plasmidis another method of insertion. The introduction of a plasmidinto a phage host will result into double-crossover homologousrecombination between the plasmid and the phage DNA, andthe introduction of the plasmid into the phage genome. Again,the expression of the reporter gene in the host cell will result ina measurable signal. The use of a reporter phage providesan economic and time-saving method of detection of patho-genic bacteria. A variety of reporter genes, such as reyluciferase (Lux), green uorescent protein (GFP), and yellowuorescent protein (ZsYellow), have been inserted into non-coding regions of a phage and were subsequently used forbacterial detection (Table 2).
4.3. Phage lysis assays
The lytic propagation of bacteriophages results in the lysis andrelease of bacterial intracellular components, as well as the
Table 2 Reporter phage assays for the detection of bacterial species
Reporter gene Bacterial specie Phage
luxI and luxR E. coli l
luxI and luxR E. coli O157:H7 PP01luxI and luxR E. coli O157:H7 PP01luxI and luxR E. coli O157:H7 PP01luxAB E. coli O157:H7 AR1luxAB Y. Pestis FA1122luxAB Salmonella enteritidis P22luxAB Salmonella Typhimurium P22luxAB Salmonella enterica P22luxAB L. monocytogenes A511luxAB Enteric bacteria CocktailluxAB B. anthracis Wb
lacZ E. coli T4inaW Salmonella P22luc M. tuberculosis phAE142luc M. tuberculosis Che/phAETRCGFP E. coli IP008e-/2xGFPGFP E. coli l
GFP M. tuberculosis TM4
This journal is © The Royal Society of Chemistry 2014
liberation of progeny virions into the extracellular environment.Their lytic properties are attributed to two phagic proteinsimplicated in the break-down of the bacterial cell wall. The rstfunctions as a hole-forming protein, providing small pores inthe bacterial cytoplasmic membrane, while the second protein,an endolysin, degrades peptidoglycans, resulting in the releaseof multiple intracellular markers that can easily be detected.77
Table 3 lists phage-mediated cell lysis assays. The most popularmethod is the measurement of bacterial intracellular adenosinetriphosphate (ATP). The amount of ATP metabolized by abacterium declines quickly, following cell death. The reyluciferase/luciferin enzyme system permits the detection andenumeration of viable bacterial cells, by sensing intracellularATP.78 The enzyme system permits the production of light thatcan be measured by a small hand-held luminometer and isdirectly proportional to the quantity of ATP in the assay.However, this system is limited by the fact that the detection ofspecic pathogens is not possible. The desired specicity canonly be achieved by specic cell lysis, which can be accom-plished by phages.77 Therefore, pathogen-specic phages andADP are added to the sample, where the phage will only infectthe target bacteria and not interact with other microorganismsthat may be present in the sample. The sensitivity of this assaycan be improved by measuring the amount of released ade-nylate kinase (AK). Similarly, intracellular AK will be releasedaer phage lysis. When ADP is present in excess, the equilib-rium of the reaction shis toward the creation of ATP, and theoverall quantity of uorescent light emitted from rey lucif-erase will be proportional to the number of viable cells inthe sample. Table 4 lists bioluminescence assays, includingthose utilizing AK. So as to further improve the sensitivity ofthe assay, AK is combined with an immunomagnetic separa-tion (IMS) step, where the pathogens of interest can becaptured on antibody-coated magnetic beads.77 Anothermethod for the identication and quantication of bacterialpathogens relies on the amperometric detection of enzymatic
Time of detection Sensitivity Ref.
10 h 1 CFU mL�1 6322 h 1 CFU mL�1 646 h 1 CFU mL�1 6412.5 h 1 CFU mL�1 643 h 10 CFU mL�1 5730 min 106 CFU mL�1 6524 h 63 CFU mL�1 6616 h 106 CFU mL�1 676 h 10 CFU mL�1 6620 h 1 CFU g�1 685 h 10 cells g�1 691 h 103 CFU mL�1 658 h 10 CFU mL�1 702 h 10 CFU mL�1 711–2 weeks 103 CFU mL�1 72
16 24 h 108 CFU mL�1 736 h Not reported 746 h 102 CFU mL�1 754 h 100 cells 76
Analyst, 2014, 139, 1224–1236 | 1229

Table 3 Phage-mediated cell lysis assays for the detection of bacterial species
Bacterial species Phage Time of detection Sensitivity Ref.
E. coli O157:H7 CSLO157 10 h 10 cells per 25 g 79E. coli PhiX174 2 h 105 CFU mL�1 80E. coli NCIMB 10359 1 h 104 CFU mL�1 81E. coli AT-20 2 h 103 CFU mL�1 82E. coli T4 2 h 103 CFU mL�1 83E. coli l 6 h 102 CFU mL�1 84B. cereus B1-7064 8 h 10 CFU mL�1 85Salmonella SJ2 2 h 103 CFU mL�1 82Salmonella newport Felix-01 and newport 2 h 104 CFU mL�1 81
Table 4 Bioluminescence assays for the detection of bacterial species
Assay type Bacterial species Phage Time of detection Sensitivity Ref.
Adenylate kinase assay E. coli E. coli phage 1 h 103 CFU mL�1 81YOYO-1 uorescent dye E. coli O157:H7 LG1 6 h 2 CFU g�1 56DAPI uorescent dye E. coli T4 6 h Not reported 86Adenylate kinase assay Salmonella newport Newport phage 2 h 103 CFU mL�1 81Adenylate kinase assay Salmonella enteritidis SJ2 2 h 103 CFU mL�1 82Adenylate kinase assay E. coli G2-2 AT20 2 h 103 CFU mL�1 82Bioluminescent phage Bacillus anthracis Wb:luxAB 60 min 103 CFU mL�1 65
Analyst Minireview
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
activity induced by intracellular bacterial enzymes liberated,following phage-induced lysis. Betagalactosidase and endoly-sins can also be monitored.
5. Phage-based biosensors
Recent years have seen the increased use of biosensors as anovel method for pathogen detection. These new systems aredesigned to overcome the many limitations of conventionaldetection platforms, such as reverse transcriptase PCR (RT-PCR)and enzyme-linked immunosorbent assays (ELISA). AlthoughRT-PCR and ELISA are popular, and prove effective in detectingthe presence of pathogens, they still require an enrichment stepprior to analysis, as well as highly trained personnel. On theother hand, biosensors are selective, sensitive, cost-effective,rapid and portable alternatives to conventional analyticalmethods. Various recognition elements, such as nucleic acidsand antibodies, can be used in pathogen detection. There are,however, very few biosensing technologies using bacterio-phages in the clinical, agro-food or environmental elds. Thelack of attention paid to phages may result from themisconception that their study is too specialized an under-taking for most engineering laboratories. However, viraldiversity and novelty are astounding, and including them insensing platforms has several benets. Here we outline thebasic principles and recent advances in biosensing technol-ogies using optical, electrochemical and mass monitoring forphage-based diagnostics.
5.1. Optical biosensors
Optical biosensors have been extensively investigated as ameans of detecting pathogens, due to their well-known
1230 | Analyst, 2014, 139, 1224–1236
selectivity and sensitivity. Of the various optical techniquesused for the detection of pathogens, surface plasmon reso-nance and evanescent eld optical ber systems dominate theindustry (e.g. Biacore, Biosensing Instruments, Inc. andResearch International).
5.1.1. Surface plasmon resonance biosensors. Surfaceplasmon resonance (SPR) is a method that can be suitablymodied for bacterial detection, as it is designed for the real-time monitoring of all dynamic processes, without labeling andcomplex sample preparation. Therefore, adsorption rate, asso-ciation and dissociation kinetics, as well as affinity constantsand ligand interactions, can be easily and rapidly examined.87
In 1902, Wood observed slender, dark bands when illumi-nating a metallic diffraction grating with polychromatic light,which he called anomalies.88 Later, Fano came to the conclusionthat the excitation of electromagnetic surface waves on thediffraction grating caused these anomalies.89 Moreover, whileilluminating thin metal lms on a substrate in 1958, Turbadarnoticed a decrease in reectivity.90 A decade later, Ottoexplained these results and demonstrated that this drop inreectivity is due to the excitation of surface plasmons observedin attenuated total reection (ATR).91 This phenomena was alsoreported the same year by Kretschmann and Raether, whileusing another ATR conguration.92
A surface plasmon is a specic mode of the electromagneticeld (i.e. a longitudinal charge density wave) that propagates at theinterface between a metal and a dielectric.93 Surface plasmons arecharacterized by a eld distribution and a complex propagatingconstant that is extremely sensitive to uctuations in the refractiveindex distribution.93 It is important to use the right kind of metal,as this metal must fulll certain requirements. The metal ofchoice must exhibit free electron behavior: the metal must followthe free electron model and its permittivity component, in the
This journal is © The Royal Society of Chemistry 2014

Fig. 2 Schematic representation of phage-based biosensing tech-niques for the detection of bacterial pathogens using (a) optical, (b)electrochemical, and (c) mass monitoring techniques.
Minireview Analyst
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
visible and near infrared regions of the spectrum, must be nega-tive.93 This is due to the fact that a surface plasmon exists at themetal-dielectric interface only at wavelengths longer than a certaincritical wavelength. This critical wavelength is contingent on theplasma frequency, and is specic to the metal. Metals, such asgold, silver and aluminum, are all suited for surface plasmonresonance because their critical wavelengths lie in the UV orvisible region of the spectrum. Gold is onemetal of choice becauseof its stability, and silver, because it provides a sharp SPR reso-nance peak.
Surface plasmons are excited by light waves, using (1) prismcoupling and attenuated total reection, (2) diffraction on ametal diffraction grating, and (3) coupling among paralleloptical waveguides.93
An SPR optical sensor generally comprises an opticalsystem, a transducing medium and an electronic systempermitting data processing. Analysis is performed bycapturing a recognition element on the sensor surface, and byinjecting the analyte over the sensor surface. A measurablesignal will be generated in the event of analyte binding to theimmobilized ligand on the sensor surface, resulting from arefractive index change in the evanescent eld, shiing theSPR angle. Fig. 2a is a schematic representation of the SPRdirect detection of captured bacteria, via immobilized bacte-riophages, on a gold sensing platform.
Current instruments have a detection limit of 10�7 refrac-tive index units (RIU), corresponding to 0.1 pg mm�2 ofmolecules gathering on the sensor surface.94 When applied topathogen detection, the current detection limit is about 105
CFU mL�1 for Listeria monocytogenes,95 E. coli O157:H7,96
Campylobacter jejuni,96 S. typhimurium,96 and S. aureus,97 usingdifferent recognition elements. Balasubramanian et al.showed that it was possible to specically detect Staphylo-coccus aureus, using a Spreeta biosensor, by physicallyadsorbing bacteriophages on the gold surface. However, thesurface coverage of the platform by bacteriophages did nothave a signicant effect on the sensitivity of detection.98 Theauthors assumed that the non-favorable orientation of thephages resulted in blocking the bacterial adhesion sites.Moreover, their results showed that it was difficult to detectbacteria in solution, due to the size of the bacterium cell (0.5–1mm). Because the evanescent wave of the SPR is limited to 300nm in the z-direction, only a portion of the bacterium willgenerate a response by interfering with the evanescent wave.Improving immobilization strategies and phage recognitionelements allowed for a more sensitive approach (i.e. detectionlimit ranging from 102–103 CFU mL�1) for the detection of S.aureus99–101 and E. coli.100 Table 5 compiles recent advances inphage-based SPR detection of pathogens.
5.1.2. Fiber optic biosensors. As with SPR biosensors, beroptic sensors react to changes in the evanescent eld due tomodications of the refractive index at the sensor surfacefollowing analyte binding.108 In general, ber-optic sensorsutilize lasers to generate evanescent waves that excite uo-rescently labeled pathogens present on the surface of thewaveguide.109 This generates uorescent signals that can bedetected in real time. Fluorescein isothiocyanate (FITC) is
This journal is © The Royal Society of Chemistry 2014 Analyst, 2014, 139, 1224–1236 | 1231
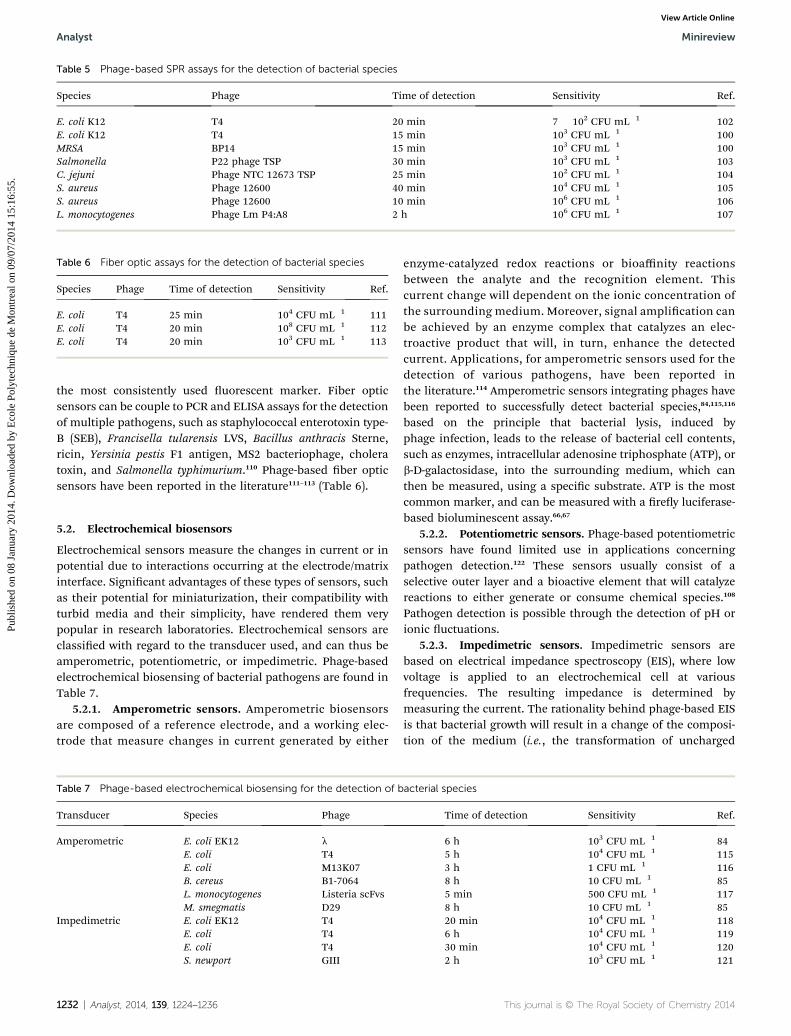
Table 5 Phage-based SPR assays for the detection of bacterial species
Species Phage Time of detection Sensitivity Ref.
E. coli K12 T4 20 min 7 � 102 CFU mL�1 102E. coli K12 T4 15 min 103 CFU mL�1 100MRSA BP14 15 min 103 CFU mL�1 100Salmonella P22 phage TSP 30 min 103 CFU mL�1 103C. jejuni Phage NTC 12673 TSP 25 min 102 CFU mL�1 104S. aureus Phage 12600 40 min 104 CFU mL�1 105S. aureus Phage 12600 10 min 106 CFU mL�1 106L. monocytogenes Phage Lm P4:A8 2 h 106 CFU mL�1 107
Table 6 Fiber optic assays for the detection of bacterial species
Species Phage Time of detection Sensitivity Ref.
E. coli T4 25 min 104 CFU mL�1 111E. coli T4 20 min 108 CFU mL�1 112E. coli T4 20 min 103 CFU mL�1 113
Analyst Minireview
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
the most consistently used uorescent marker. Fiber opticsensors can be couple to PCR and ELISA assays for the detectionof multiple pathogens, such as staphylococcal enterotoxin type-B (SEB), Francisella tularensis LVS, Bacillus anthracis Sterne,ricin, Yersinia pestis F1 antigen, MS2 bacteriophage, choleratoxin, and Salmonella typhimurium.110 Phage-based ber opticsensors have been reported in the literature111–113 (Table 6).
5.2. Electrochemical biosensors
Electrochemical sensors measure the changes in current or inpotential due to interactions occurring at the electrode/matrixinterface. Signicant advantages of these types of sensors, suchas their potential for miniaturization, their compatibility withturbid media and their simplicity, have rendered them verypopular in research laboratories. Electrochemical sensors areclassied with regard to the transducer used, and can thus beamperometric, potentiometric, or impedimetric. Phage-basedelectrochemical biosensing of bacterial pathogens are found inTable 7.
5.2.1. Amperometric sensors. Amperometric biosensorsare composed of a reference electrode, and a working elec-trode that measure changes in current generated by either
Table 7 Phage-based electrochemical biosensing for the detection of b
Transducer Species Phage
Amperometric E. coli EK12 l
E. coli T4E. coli M13K07B. cereus B1-7064L. monocytogenes Listeria scFvsM. smegmatis D29
Impedimetric E. coli EK12 T4E. coli T4E. coli T4S. newport GIII
1232 | Analyst, 2014, 139, 1224–1236
enzyme-catalyzed redox reactions or bioaffinity reactionsbetween the analyte and the recognition element. Thiscurrent change will dependent on the ionic concentration ofthe surrounding medium. Moreover, signal amplication canbe achieved by an enzyme complex that catalyzes an elec-troactive product that will, in turn, enhance the detectedcurrent. Applications, for amperometric sensors used for thedetection of various pathogens, have been reported inthe literature.114 Amperometric sensors integrating phages havebeen reported to successfully detect bacterial species,84,115,116
based on the principle that bacterial lysis, induced byphage infection, leads to the release of bacterial cell contents,such as enzymes, intracellular adenosine triphosphate (ATP), orb-D-galactosidase, into the surrounding medium, which canthen be measured, using a specic substrate. ATP is the mostcommon marker, and can be measured with a rey luciferase-based bioluminescent assay.66,67
5.2.2. Potentiometric sensors. Phage-based potentiometricsensors have found limited use in applications concerningpathogen detection.122 These sensors usually consist of aselective outer layer and a bioactive element that will catalyzereactions to either generate or consume chemical species.108
Pathogen detection is possible through the detection of pH orionic uctuations.
5.2.3. Impedimetric sensors. Impedimetric sensors arebased on electrical impedance spectroscopy (EIS), where lowvoltage is applied to an electrochemical cell at variousfrequencies. The resulting impedance is determined bymeasuring the current. The rationality behind phage-based EISis that bacterial growth will result in a change of the composi-tion of the medium (i.e., the transformation of uncharged
acterial species
Time of detection Sensitivity Ref.
6 h 103 CFU mL�1 845 h 104 CFU mL�1 1153 h 1 CFU mL�1 1168 h 10 CFU mL�1 855 min 500 CFU mL�1 1178 h 10 CFU mL�1 8520 min 104 CFU mL�1 1186 h 104 CFU mL�1 11930 min 104 CFU mL�1 1202 h 103 CFU mL�1 121
This journal is © The Royal Society of Chemistry 2014

Table 8 Phage-based magnetoelastic biosensing for the detection of bacterial species
Species Phage Time of detection Sensitivity Ref.
B. anthracis JRB-7 30 min 103 CFU mL�1 138B. anthracis Filamentous landscape
phage1 h 1 cell Per spore 132
B. anthracis JRB-7 2 h 104 CFU mL�1 128S. typhimurium Filamentous phage 20 min 103 CFU mL�1 133S. typhimurium JRB-7 30 min 103 CFU mL�1 139
Minireview Analyst
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
metabolites, such as carbohydrates, into charged products,mainly acids), causing an increase in medium conductivityFig. 2b. The lysis of host bacteria by phages reduces microbialgrowth. The conductances of phage-treated and untreatedsamples are then compared. Limitations of direct impedimetricdetection include the necessity of optimizing the medium forelectrical measurements, as well as selecting microorganismsthat produce sufficient amounts of ionized metabolites topermit their detection.123 Successful phage-base impedimetricdetections of E. coli,MRSA and Salmonella have been reported inthe literature.118,120,124–126
5.3. Phage-based mass biosensors
5.3.1. Acoustic wave biosensors. Acoustic wave biosensorsare mass-sensitive detectors, using quartz-crystal resonators.These oscillating crystals resonate at a frequency that dependson their geometry. There is a direct relationship between themass adsorbed on the surface of the crystal and its resonantfrequency in air. Most acoustic biosensors use piezoelectrictransducers that generate and transmit acoustic waves with avibrating crystal, which resonates in a frequency-dependentmanner. Of the acoustic wave sensors using a piezoelectrictransducer, the quartz crystal microbalance (QCM) is largelyused for pathogen detection. Applying an electrical eld toelectrodes situated on each side of the crystal creates a potentialdifference across the two sides. This produces a mechanicaloscillation with a characteristic vibrational frequency, and issensitive to mass Fig. 2c. Olsen et al. reported the detection ofSalmonella typhimurium, using specic bacteriophage, with adetection limit of 102 CFU mL.127
5.3.2. Magnetoelastic sensors. Magnetoelastic sensorsare composed of a magneto-restrictive material thatcontracts or lengthens when exposed to an external magneticeld. The resonant frequency is dependent on the massbound to the surface of the resonator. The phage-baseddetection of Salmonella typhimurium and Bacillus anthracishas been reported with a detection limit in the range of 103
CFU mL (ref. 128–138) (Table 8).
6. Conclusions
Novel diagnostic technologies are of great importance for thecontrol of drug-resistant pathogens. Rapid, specic, andsensitive means should aim at facilitating the diagnosis andenabling an accurate and efficient decision-making in indi-vidual point-of-care settings. The selectivity, specicity,
This journal is © The Royal Society of Chemistry 2014
robustness, and low production cost of phages make them idealrecognition elements for bacterial detection. However, despiteefforts at commercializing phage-based diagnostic devices,traditional microbiological methods remain the gold standardfor diagnosis in a clinical setting. Although vast efforts havereduced identication times from days to hours, when usingmodern molecular techniques, such as PCR, most assays stillrequire an amplication period to increase the bacterial load ofthe specimen to be tested. There is, therefore, a requirement forbiosensing technologies with sensitivities permitting thedetection of concentrations below 102 CFU mL�1. In addition,future research efforts should be aimed at detecting bacteriawithin complex matrices, in order to effectively move phagediagnostics from the laboratory to the clinic.
Acknowledgements
We thank the Natural Sciences and Engineering ResearchCouncil of Canada and Biophage Pharma for funding this work.
Notes and references
1 H. Pennington, Nature, 2007, 446, 981.2 M. A. Kohanski, D. J. Dwyer and J. J. Collins, Nat. Rev.Microbiol., 2010, 8, 423–435.
3 A. Typas, M. Banzhaf, C. A. Gross and W. Vollmer, Nat. Rev.Microbiol., 2012, 10, 123–136.
4 C. A. Arias and B. E. Murray, N. Engl. J. Med., 2009, 360, 439–443.
5 R. R. Roberts, B. Hota, I. Ahmad, R. D. Scott, II, S. D. Foster,F. Abbasi, S. Schabowski, L. M. Kampe, G. G. Ciavarella,M. Supino, J. Naples, R. Cordell, S. B. Levy andR. A. Weinstein, Clin. Infect. Dis., 2009, 49, 1175–1184.
6 J. E. Dover, G. M. Hwang, E. H. Mullen, B. C. Prorok andS.-J. Suh, J. Microbiol. Methods, 2009, 78, 10–19.
7 H. W. Boucher, G. H. Talbot, J. S. Bradley, J. E. Edwards, Jr,D. Gilbert, L. B. Rice, M. Scheld, B. Spellberg and J. Bartlett,Clin. Infect. Dis., 2009, 48, 1–12.
8 H. F. L. Wertheim, D. C. Melles, M. C. Vos, W. van Leeuwen,A. van Belkum, H. A. Verbrugh and J. L. Nouwen, LancetInfect. Dis., 2005, 5, 751–762.
9 D. J. Biedenbach, G. J. Moet and R. N. Jones, Diagn.Microbiol. Infect. Dis., 2004, 50, 59–69.
10 A. Pantosti and M. Venditti, Eur. Respir. J., 2009, 34,1190–1196.
Analyst, 2014, 139, 1224–1236 | 1233

Analyst Minireview
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
11 D. Lim and N. C. J. Strynadka, Nat. Struct. Biol., 2002, 9,870–876.
12 D. A. Schoeld, N. J. Sharp and C. Westwater, PubMedCentral, 2012, 2, 105–283.
13 R. M. Klevens, M. A. Morrison, J. Nadle, S. Petit, K. Gershman,S. Ray, L. H. Harrison, R. Lyneld, G. Dumyati, J. M. Townes,A. S. Craig, E. R. Zell, G. E. Fosheim, L. K. McDougal,R. B. Carey, S. K. Fridkin and A. B. M. Investigators, JAMA,J. Am. Med. Assoc., 2007, 298, 1763–1771.
14 HIV Surveillance Report, C. f. D. C. a. Prevention, ed. C. f. D.C. a. P, US Department of Health and Human Services,Atlanta, 2007, vol. 17.
15 W. H. Organization, Weekly epidemiological record, 2010,vol. 85, pp. 37–48.
16 E. D. Williamson and P. C. F. Oyston, Clin. Exp. Immunol.,2013, 172, 1–8.
17 M. B. dos Santos, J. P. Agusil, B. Prieto-Simon, C. Sporer,V. Teixeira and J. Samitier, Biosens. Bioelectron., 2013, 45,174–180.
18 C. Hernandez-Reyes and A. Schikora, FEMS Microbiol. Lett.,2013, 343, 1–7.
19 C. f. D. C. a. Prevention, Morbidity and Mortality WeeklyReport (MMWR), 2011, vol. 60, pp. 749–755.
20 E. P. da Silva and E. C. P. De Martinis, Appl. Microbiol.Biotechnol., 2013, 97, 957–968.
21 Risk assessment of Listeria monocytogenes in ready-to-eatfoods: interpretative summary, Microbial Risk assessmentseries, W. H. Organization, 2013, http://www.who.int/foodsafety/publications/micro/en/mra4.pdf.
22 G. Griffiths, B. Nystroem, S. B. Sable and G. K. Khuller, Nat.Rev. Microbiol., 2010, 8, 827–834.
23 E. Kutter and A. Sulakvelidze, Bacteriophages: Biology andApplications, CRC Press, Boca Raton, Florida, 2005.
24 F. Twort, Lancet, 1915, 11, 1241.25 F. D'Herelle, Comptes rendusde l'Academie des Sciences,
1917, 165, 373–375.26 F. D'Herelle, Compte rendu de l'Academie des Sciences, 1919,
168, 631.27 F. D'Herelle, Compte rendu de l'Academie des Sciences, 1919,
169, 817.28 W. B. Cherry, B. R. Davis, P. R. Edwards and R. B. Hogan,
J. Lab. Clin. Med., 1954, 44, 51–55.29 R. Mandeville, M. Griffiths, L. Goodridge, L. McIntyre and
T. T. Henchuk, Anal. Lett., 2003, 36, 3241–3259.30 R. Moldovan, E. Chapman-McQuiston and X. L. Wu,
Biophys. J., 2007, 93, 303–315.31 S. Sidhu, Phage display in biotechnology and drug discovery,
CRC Press, 2005.32 T. K. Lu and J. J. Collins, Proc. Natl. Acad. Sci. U. S. A., 2009,
106, 4629–4634.33 T. K. Lu and J. J. Collins, IEEE, Engineering Synthetic
Bacteriophage to Combat Antibiotic-Resistant Bacteria, 2009.34 T. K. Lu, J. Bowers and M. S. Koeris, Trends Biotechnol.,
2013, 31, 325–327.35 B. Zakeri and T. K. Lu, ACS Synth. Biol., 2013, 2, 358–372.36 N. Tawil, E. Sacher, E. Boulais, R. Mandeville and
M. Meunier, J. Phys. Chem. C, 2013, 117, 20656–20665.
1234 | Analyst, 2014, 139, 1224–1236
37 J. Minion andM. Pai, Int. J. Tuberc. Lung Dis., 2010, 14, 941–951.38 K. V. Sergueev, Y. X. He, R. H. Borschel, M. P. Nikolich and
A. A. Filippov, PLoS One, 2010, 5, DOI: 10.1371/journal.pone.0011337.
39 C. T. Zhu, Z. L. Cui, R. J. Zheng, H. Yang, R. L. Jin, L. H. Qin,Z. H. Liu, J. Wang and Z. Y. Hu, PLoS One, 2011, 6, DOI:10.1371/journal.pone.0024435.
40 D. Biswas, A. Deb, P. Gupta, R. Prasad and K. S. Negi, IndianJ. Med. Microbiol., 2008, 26, 75–78.
41 M. E. S. Zaki and T. Goda, Tuberculosis, 2007, 87, 102–108.42 N. Simboli, H. Takiff, R. McNerney, B. Lopez, A. Martin,
J. C. Palomino, L. Barrera and V. Ritacco, Antimicrob.Agents Chemother., 2005, 49, 425–427.
43 G. Stewart, S. A. A. Jassim, S. P. Denyer, P. Newby, K. Linleyand V. K. Dhir, J. Appl. Microbiol., 1998, 84, 777–783.
44 C. L. Pierce, J. C. Rees, F. M. Fernandez and J. R. Barr, Anal.Chem., 2011, 83, 2286–2293.
45 C. L. Pierce, J. C. Rees, F. M. Fernandez and J. R. Barr, Mol.Cell. Proteomics, 2012, 11, DOI: 10.1074/mcp.M111.012849.
46 C. R. Cox, J. C. Rees and K. J. Voorhees, J. Mass Spectrom.,2012, 47, 1435–1441.
47 H. Albert, A. Heydenrych, R. Mole, A. Trollip andL. Blumberg, Int. J. Tuberc. Lung Dis., 2001, 5, 906–911.
48 R. J. Mole and T. W. O. Maskell, J. Chem. Technol.Biotechnol., 2001, 76, 683–688.
49 H. Albert, A. Trollip, I. Seaman and R. J. Mole, Int. J. Tuberc.Lung Dis., 2004, 8, 1114–1119.
50 I. J. Eltringham, S. M. Wilson and F. A. Drobniewski, J. Clin.Microbiol., 1999, 37, 3528–3532.
51 K. R. Hance, B. C. Smith, T. D. Steinmark and J. C. Rees, Am.J. Clin. Pathol., 2005, 124, 644–645.
52 R. K. Kutin, A. Alvarez and D. M. Jenkins, J. Microbiol.Methods, 2009, 76, 241–246.
53 R. W. Reiman, D. H. Atchley and K. J. Voorhees, J. Microbiol.Methods, 2007, 68, 651–653.
54 R. P. Tiwari, N. S. Hattikudur, R. N. Bharmal, S. Kartikeyan,N. M. Deshmukh and P. S. Bisen, Tuberculosis, 2007, 87,193–201.
55 S. J. Favrin, S. A. Jassim and M. W. Griffiths, Int. J. FoodMicrobiol., 2003, 85, 63–71.
56 L. Goodridge, A. Gallaccio and M. W. Griffiths, Appl.Environ. Microbiol., 2003, 69, 5364–5371.
57 N. Ulitzur and S. Ulitzur, Appl. Environ. Microbiol., 2006, 72,7455–7459.
58 A. J. Madonna, S. Van Cuyk and K. J. Voorhees, RapidCommun. Mass Spectrom., 2003, 17, 257–263.
59 R. S. de Siqueira, C. E. R. Dodd and C. E. D. Rees, Braz.J. Microbiol., 2003, 34, 118–120.
60 S. J. Favrin, S. A. Jassim and M. W. Griffiths, Appl. Environ.Microbiol., 2001, 67, 217–224.
61 A. Foddai, C. T. Elliott and I. R. Grant, Appl. Environ.Microbiol., 2009, 75, 3896–3902.
62 D. A. Schoeld, I. J. Molineux and C. Westwater, J. Clin.Microbiol., 2009, 47, 3887–3894.
63 S. Ripp, P. Jegier, M. Birmele, C. M. Johnson, K. A. Daumer,J. L. Garland and G. S. Sayler, J. Appl. Microbiol., 2006, 100,488–499.
This journal is © The Royal Society of Chemistry 2014

Minireview Analyst
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
64 S. Ripp, P. Jegier, C. M. Johnson, J. R. Brigati andG. S. Sayler, Anal. Bioanal. Chem., 2008, 391, 507–514.
65 D. A. Schoeld and C. Westwater, J. Appl. Microbiol., 2009,107, 1468–1478.
66 J. Chen and M. W. Griffiths, J. Food Prot., 1996, 59, 908–914.67 G. Thouand, P. Vachon, S. Liu, M. Dayre and
M. W. Griffiths, J. Food Prot., 2008, 71, 380–385.68 M. J. Loessner, M. Rudolf and S. Scherer, Appl. Environ.
Microbiol., 1997, 63, 2961–2965.69 C. P. Kodikara, H. H. Crew and G. Stewart, FEMS Microbiol.
Lett., 1991, 83, 261–266.70 F. Tetart, F. Repoila, C. Monod and H. M. Krisch, J. Mol.
Biol., 1996, 258, 726–731.71 P. K. Wolber and R. L. Green, Trends Biotechnol., 1990, 8,
276–279.72 N. Banaiee, M. Bobadilla-Del-Valle, P. F. Riska, S. Bardarov,
P. M. Small, A. Ponce-De-Leon, W. R. Jacobs, G. F. Hatfulland J. Sifuentes-Osornio, J. Med. Microbiol., 2003, 52, 557–561.
73 V. Kumar, P. Loganathan, G. Sivaramakrishnan, J. Kriakov,A. Dusthakeer, B. Subramanyam, J. Chan, W. R. Jacobs andN. P. Rama, Tuberculosis, 2008, 88, 616–623.
74 M. Namura, T. Hijikata, K. Miyanaga and Y. Tanji,Biotechnol. Prog., 2008, 24, 481–486.
75 T. Funatsu, T. Taniyama, T. Tajima, H. Tadakuma andH. Namiki, Microbiol. Immunol., 2002, 46, 365–369.
76 M. Piuri, W. R. Jacobs and G. F. Hatfull, PLoS One, 2009, 4,DOI: 10.1371/journal.pone.0004870.
77 M. Zourob, S. Elwary and A. Turner, Principles of BacterialDetection, Springer, New-York, 2008.
78 P. E. Stanley, J. Biolumin. Chemilumin., 1989, 4, 375–380.79 P. Kannan, H. Y. Yong, L. Reiman, C. Cleaver, P. Patel and
A. A. Bhagwat, Foodborne Pathog. Dis., 2010, 7, 1551–1558.80 O. Laczka, C. Garcia-Aljaro, F. J. del Campo,
F. X. M. Pascual, J. Mas-Gordi and E. Baldrich, Anal.Chim. Acta, 2010, 677, 156–161.
81 R. Blasco, M. J. Murphy, M. F. Sanders and D. J. Squirrell,J. Appl. Microbiol., 1998, 84, 661–666.
82 Y. Wu, L. Brovko and M. W. Griffiths, Lett. Appl. Microbiol.,2001, 33, 311–315.
83 O. Minikh, M. Tolba, L. Y. Brovko and M. W. Griffiths,J. Microbiol. Methods, 2010, 82, 177–183.
84 T. Neufeld, A. Schwartz-Mittelmann, D. Biran, E. Z. Ron andJ. Rishpon, Anal. Chem., 2003, 75, 580–585.
85 M. Yemini, Y. Levi, E. Yagil and J. Rishpon,Bioelectrochemistry, 2007, 70, 180–184.
86 T. Kenzaka, F. Utrarachkij, O. Suthienkul and M. Nasu,J. Health Sci., 2006, 52, 666–671.
87 J. Davies, Nanobiology, 1994, 3, 5–16.88 R. W. Wood, Philos. Mag., 1902, 3, 396–410.89 U. Fano, J. Opt. Soc. Am., 1941, 31, 213–222.90 T. Turbadar, Proc. Phys. Soc., London, 1959, 73, 5.91 A. Otto, Zeitschri fur Physik A Hadrons and Nuclei, 1968,
216, 398–410.92 H. R. E. Kretschmann, Z. Naturforsch., 1968, 23A, 2135–
2136.93 J. Homola, Surface Plasmon Resonance Based Sensors,
Springer Berlin Heidelberg, Berlin, 2006.
This journal is © The Royal Society of Chemistry 2014
94 G. De Crescenzo, C. Boucher, Y. Durocher and M. Jolicoeur,Cell. Mol. Bioeng., 2008, 1, 204–215.
95 P. Leonard, S. Hearty, J. Quinn and R. O'Kennedy, Biosens.Bioelectron., 2004, 19, 1331–1335.
96 A. D. Taylor, J. Ladd, Q. M. Yu, S. F. Chen, J. Homola andS. Y. Jiang, Biosens. Bioelectron., 2006, 22, 752–758.
97 A. Subramanian, J. Irudayaraj and T. Ryan, Sens. Actuators,B, 2006, 114, 192–198.
98 S. Balasubramanian, I. B. Sorokulova, V. J. Vodyanoy andA. L. Simonian, Biosens. Bioelectron., 2007, 22, 948–955.
99 R. Guntupalli, I. Sorokulova, A. Krumnow, O. Pustovyy,E. Olsen and V. Vodyanoy, Biosens. Bioelectron., 2008,151–154.
100 N. Tawil, E. Sacher, R. Mandeville and M. Meunier, Biosens.Bioelectron., 2012, 37, 24–29.
101 N. Tawil, E. Sacher, R. Mandeville and M. Meunier, J. Phys.Chem. C, 2013, 6686–6691.
102 S. K. Arya, A. Singh, R. Naidoo, P. Wu, M. T. McDermott andS. Evoy, Analyst, 2011, 136, 486–492.
103 A. Singh, S. K. Arya, N. Glass, P. Hani-Moghaddam,R. Naidoo, C. M. Szymanski, J. Tanha and S. Evoy,Biosens. Bioelectron., 2010, 26, 131–138.
104 A. Singh, D. Arutyunov, M. T. McDermott, C. M. Szymanskiand S. Evoy, Analyst, 2011, 136, 4780–4786.
105 S. Balasubramanian, I. B. Sorokulova, V. J. Vodyanoy andA. L. Simonian, Biosens. Bioelectron., 2007, 22, 948–955.
106 R. Guntupalli, I. Sorokulova, A. Krumnow, O. Pustovyy,E. Olsen and V. Vodyanoy, Biosens. Bioelectron., 2008, 24,151–154.
107 V. Nanduri, A. K. Bhunia, S. I. Tu, G. C. Paoli andJ. D. Brewster, Biosens. Bioelectron., 2007, 23, 248–252.
108 H. Sharma and R.Mutharasan, Sens. Actuators, B, 2013, 183,535–549.
109 X. D.Wang andO. S. Woleis, Anal. Chem., 2013, 85, 487–508.110 G. P. Anderson, B. M. Lingerfelt and C. R. Taitt, Sens. Lett.,
2004, 2, 18–24.111 A. Mazhorova, A. Markov, A. Ng, R. Chinnappan,
O. Skorobogata, M. Zourob and M. Skorobogatiy, Opt.Express, 2012, 20, 5344–5355.
112 M. Smietana, W. J. Bock, P. Mikulic, A. Ng, R. Chinnappanand M. Zourob, Opt. Express, 2011, 19, 7971–7978.
113 S. M. Tripathi, W. J. Bock, P. Mikulic, R. Chinnappan, A. Ng,M. Tolba and M. Zourob, Biosens. Bioelectron., 2012, 35,308–312.
114 J. D. Brewster, A. G. Gehring, R. S. Mazenko,L. J. VanHouten and C. J. Crawford, Anal. Chem., 1996, 68,4153–4159.
115 L. Gervais, M. Gel, B. Allain, M. Tolba, L. Brovko,M. Zourob, R. Mandeville, M. Griffiths and S. Evoy, Sens.Actuators, B, 2007, 125, 615–621.
116 T. Neufeld, A. S. Mittelman, V. Buchner and J. Rishpon,Anal. Chem., 2005, 77, 652–657.
117 I. Benhar, I. Eshkenazi, T. Neufeld, J. Opatowsky, S. Shakyand J. Rishpon, Talanta, 2001, 55, 899–907.
118 A. Shabani, M. Zourob, B. Allain, C. A. Marquette,M. F. Lawrence and R. Mandeville, Anal. Chem., 2008, 80,9475–9482.
Analyst, 2014, 139, 1224–1236 | 1235

Analyst Minireview
Publ
ishe
d on
08
Janu
ary
2014
. Dow
nloa
ded
by E
cole
Pol
ytec
hniq
ue d
e M
ontr
eal o
n 09
/07/
2014
15:
16:5
5.
View Article Online
119 L. Yao, P. Lamarche, N. Tawil, R. Khan, A. M. Aliakbar,M. H. Hassan, V. P. Chodavarapu and R. Mandeville, IEEETrans Biomed Circuits Syst., 2011, 5, 223–230.
120 M. B. Mejri, H. Baccar, E. Baldrich, F. J. Del Campo,S. Helali, T. Ktari, A. Simonian, M. Aouni andA. Abdelghani, Biosens. Bioelectron., 2010, 26, 1261–1267.
121 R. Dadarwal, A. Namvar, D. F. Thomas, J. C. Hall andK. Warriner, Mater. Sci. Eng., C, 2009, 29, 761–765.
122 H. K. Zhang, X. Li, Y. P. Bai, R. F. Niu, Y. F. Jia, C. Z. Zhang,L. Zhang, X. Z. Feng and Y. J. Cao, Biotechnol. Appl.Biochem., 2009, 53, 185–192.
123 M. B. Falahee, S. F. Park and M. R. Adams, J. Food Prot.,2003, 66, 1724–1726.
124 A. Shabani, M. Zourob, B. Allain, M. Lawrence andR. Mandeville, IEEE, Electrochemical detection of bacteriausing bacteriophage, 2007.
125 L. Yao, M. Haj-Hassan, E. Ghafar-Zadeh, A. Shabani,V. Chodavarapu and M. Zourob, in IEEE 2008 CanadianConference on Electrical and Computer Engineering, vol. 1–4, 2008, pp. 836–839.
126 N. Tawil, E. Sacher, R. Mandeville and M. Meunier, Biosens.Bioelectron., 2012, 37, 24–29.
127 E. V. Olsen, I. B. Sorokulova, V. A. Petrenko, I. H. Chen,J. M. Barbaree and V. J. Vodyanoy, Biosens. Bioelectron.,2006, 21, 1434–1442.
128 L. L. Fu, S. Q. Li, K. W. Zhang, I. H. Chen, J. M. Barbaree,A. X. Zhang and Z. Y. Cheng, IEEE Sens. J., 2011, 11,1684–1691.
129 L. L. Fu, S. Q. Li, K. W. Zhang, I. H. Chen, V. A. Petrenko andZ. Y. Cheng, Sensors, 2007, 7, 2929–2941.
130 S. Horikawa, D. Bedi, S. Q. Li, W. Shen, S. C. Huang,I. H. Chen, Y. T. Chai, M. L. Auad, M. J. Bozack,
1236 | Analyst, 2014, 139, 1224–1236
J. M. Barbaree, V. A. Petrenko and B. A. Chin, Biosens.Bioelectron., 2011, 26, 2361–2367.
131 S. Horikawa, K. A. Vaglenov, D. M. Gerken, Y. T. Chai,M. K. Park, S. Q. Li, V. A. Petrenko and B. A. Chin, inSensing for Agriculture and Food Quality and Safety IV, ed.M. S. Kim, S. I. Tu and K. Chao, 2012, vol. 8369.
132 M. L. Johnson, J. H. Wan, S. C. Huang, Z. Y. Cheng,V. A. Petrenko, D. J. Kim, I. H. Chen, J. M. Barbaree,J. W. Hong and B. A. Chin, Sens. Actuators, A, 2008, 144, 38–47.
133 R. S. Lakshmanan, R. Guntupalli, J. Hu, D. J. Kim,V. A. Petrenko, J. M. Barbaree and B. A. Chin, J. Microbiol.Methods, 2007, 71, 55–60.
134 R. S. Lakshmanan, R. Guntupalli, J. Hu, V. A. Petrenko,J. M. Barbaree and B. A. Chin, Sens. Actuators, B, 2007,126, 544–550.
135 R. S. Lakshmanan, R. Guntupalli, S. Huang, M. L. Johnson,L. C. Mathison, I. H. Chen, V. A. Petrenko, Z. Y. Cheng andB. A. Chin, inMaterials and Devices for Smart Systems III, ed.J. Su, L. P. Wang, Y. Furuya, S. TrolierMcKinstry and J. Leng,2009, vol. 1129, pp. 131–136.
136 R. S. Lakshmanan, J. Hu, R. Guntupalli, J. H. Wan,S. C. Huang, H. Yang, V. A. Petrenko, J. M. Barbaree andB. A. Chin, in Chemical and Biological Sensing VII, ed. P. J.Gardner and A.W. Fountain, 2006, vol. 6218, pp. Z2180–Z2180.
137 M. K. Park, J. H. Oh and B. A. Chin, Sens. Actuators, B, 2011,160, 1427–1433.
138 J. H. Wan, M. L. Johnson, R. Guntupalli, V. A. Petrenko andB. A. Chin, Sens. Actuators, B, 2007, 127, 559–566.
139 S. Huang, H. Yang, R. S. Lakshmanan, M. L. Johnson,J. Wan, I. H. Chen, H. C. Wikle, V. A. Petrenko,J. M. Barbaree and B. A. Chin, Biosens. Bioelectron., 2009,24, 1730–1736.
This journal is © The Royal Society of Chemistry 2014