ampk i mtor
DESCRIPTION
asdfTRANSCRIPT

Adenosin monofosfat cinasa (AMPK):
un important regulador metabòlic.
L’AMPK (Adenosine 5’-monophosphate (AMP)-activated protein kinase) és un enzim clau per a la regulació global del metabolisme energètic de l’organisme. S’expressa en òrgans metabòlicament rellevants (fetge, pàncrees, múscul, adipòcits, etc.) i la seva activitat depèn de les condicions energètiques cel·lulars (és un sensor energètic).
Quan els nivells energètics de la cèl·lula són alts (concentracions altes d’ATP), l’AMPK es troba inactiva. En canvi, quan els nivells energètics de la cèl·lula són baixos (baixes concentracions d’ATP i altes concentracions d’AMP), aquest enzim s’activa al·lostèricament per AMP i s’encarrega de fosforilar múltiples proteïnes diana relacionades amb la producció i el consum d’energia.
En línies generals, l’AMPK activa vies productores d’energia (catabòliques) i inhibeix vies de consum energètic (anabòliques):
• Estimula l’absorció i oxidació de glucosa en el múscul. • Inhibeix la síntesi de glucosa. • Estimula l’oxidació hepàtica d’àcids grassos. • Estimula l’oxidació d’àcids grassos al múscul. • Inhibeix la síntesi d’àcids grassos i colesterol. • Inhibeix la síntesi proteica i el creixement cel·lular (inhibeix mTOR). • Activa p53 • Afavoreix l’autofàgia • Estimula la biogènesi mitocondrial i el metabolisme oxidatiu.
1
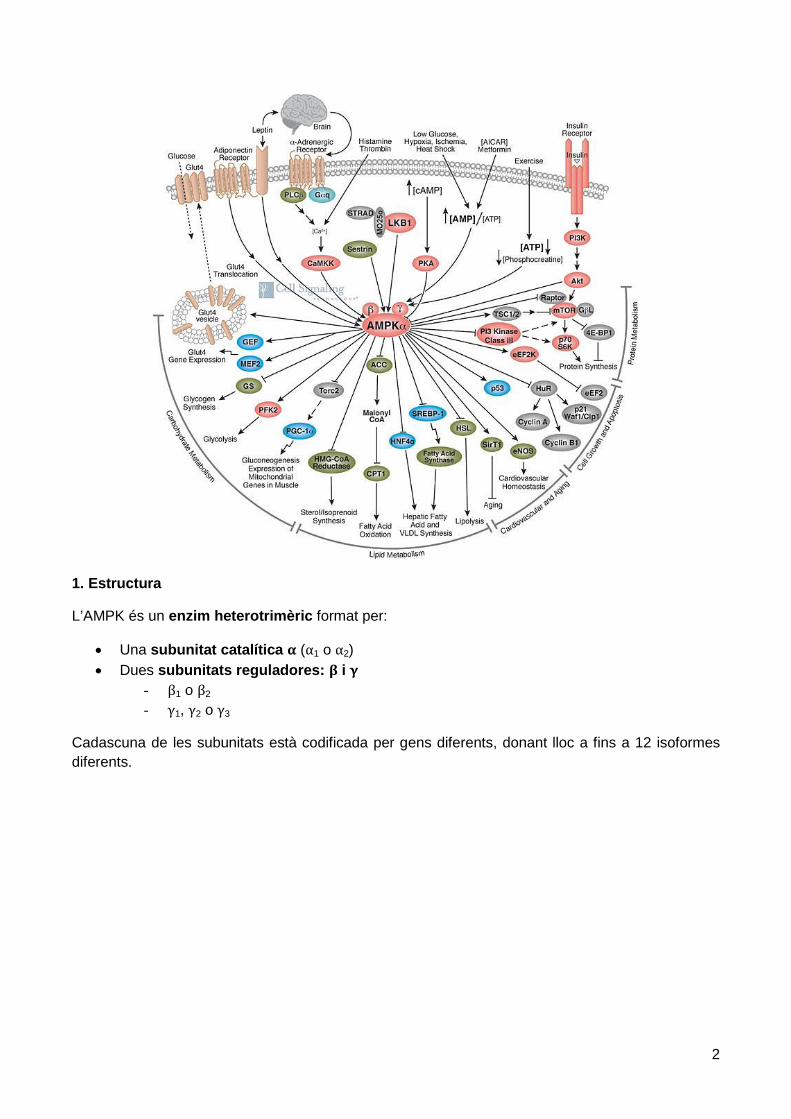
1. Estructura
L’AMPK és un enzim heterotrimèric format per:
• Una subunitat catalítica α (α1 o α2) • Dues subunitats reguladores: β i γ
- β1 o β2 - γ1, γ2 o γ3
Cadascuna de les subunitats està codificada per gens diferents, donant lloc a fins a 12 isoformes diferents.
2

2. Activació
AMPK s’activa per dues senyals diferents:
• Unió al·lostèrica d’AMP • Fosforilació: una treonina en posició 172 (Thr172) es fosforila mitjançant dues vies:
- Via de CAMKKβ, calmodulin cinasa depenent de calci. - Via de LKB1 (Liver Kinase B1), depenent d’AMP. És un supressor tumoral que es
troba mutat en la síndrome de Peutz-Jeghers, amb alta susceptibilitat al càncer.
Com veurem l’AMPK s’activa per múltiples processos, incloent aquells que provoquen alteracions en el ratio ATP/AMP intracel·lular (hipòxia), les concentracions de calci, així com l’acció de certes hormones, citoquines, adipoquines o exercici.
L’AMP s’uneix a la subunitat gamma i activa l’enzim. Al mateix temps, afavoreix que LKB1 fosforili la subunitat alfa (T172), produint un augment d’activitat de més de 100 vegades. En aquesta situació, AMPK fosforila tots els substrats que troba. Quan hi ha concentracions altes d’ATP, s’uneix a la subunitat gamma, afavorint l’enllaç amb la fosfataxa PP2C que defosforila l’AMPK inactivant.la. La fosforilació de l’AMPK per PI3K-Akt en la Serina485/491 en resposta a insulina causa un canvi de conformació que impedeix l’activació per LKB1.
Activadors
Activen AMPK de manera indirecta, modificant la cadena respiratòria mitocondrial o l’estat cel·lular de la cèl·lula.
• Metformina: és un fàrmac antihiperglucèmic que inhibeix l’alliberament de glucosa pel fetge, molt estès en el tractament de la diabetis. Inhibeix la CRM (cadena respiratòria), augmenta AMP i activa l’AMPK i LKB1.
• Tiazolidinediones (TZDs): augmenten la sensibilitat a glucosa. Algunes d’elles activen AMPK. També estimulen l’alliberament d’adiponectina per part dels adipòcits, la qual activa AMPK en el fetge, reduint la producció de glucosa.
• Adiponectina: activa AMPK al múscul esquelètic i al fetge. • Leptina: activa AMPK al múscul i fetge. Inhibeix AMPK Hipotàlem.
3

• Factor neurotròfic ciliar (CNTF): la senyalització desencadenada per aquest factor incrementa l’oxidació d’àcids grassos i la sensibilitat a la insulina mitjançant l’activació d’AMPK.
• Grelina/Cannabinoides: tant la Grelina com els Cannabinoides són potents orexígens que incrementen l’activitat d’AMPK en l’hipotàlem i el cor, mentre que l’inhibeixen al fetge i al teixit adipós. Efectes contraris a la Leptina.
• Productes naturals: alcaloides, extracte de meló amarg (Momordica charantia), gal·lat d’epigalocatequina (present en el tè verd), etc.
• Àcid α-lipoic: inactiva AMPK en l’hipotàlem, l’activa en el múscul. • Resveratrol/SIRT1: L’activació de SIRT1 (una desacetilasa) per resveratrol incrementa
l’activitat AMPK. • AICAR: és un intermediari nucleotídic, anàleg de l’AMP, capaç d’estimular AMPK. • Altres fàrmacs: A-769662, PT1
3. AMPK en el fetge i el múscul
En la diabetis de tipus 2 (T2DM), els òrgans amb major resistència a la insulina són el fetge, el múscul i el teixit adipós, la qual cosa dificulta l’absorció i utilització de glucosa per part d’aquests, conduint així a un estat de constant hiperglucèmia. Com veurem a continuació, l’AMPK podria representar un potencial agent farmacològic per revertir aquestes anomalies metabòliques associades a la diabetis tipus 2.
3.1. AMPK en el fetge
• En el fetge l’AMPK controla l’homeòstasi de la glucosa a través de la inhibició de la gluconeogènesi. Concretament, l’AMPK inhibeix l’expressió d’enzims de la gluconeogènesi com la fosfoenolpiruvat carboxiquinasa i la glucosa-6-fosfatasa, i ho fa inactivant factors de transcripció com CRTC2 o FOXO1.
• També disminueix la lipogènesi hepàtica. AMPK inactiva l’acetil-CoA carboxilasa 1 (ACC1) i la HMG-CoA reductasa, inhibint així les vies de síntesi d’àcids grassos i colesterol. També inactiva la glicerol-3-fosfat aciltransferasa, inhibint la síntesi de triglicèrids.
• A més, augmenta la β-oxidació d’àcids grassos. També augmenta l’activitat de la malonil-CoA descarboxilasa (MCD), disminuint així els nivells de malonil-CoA, que és un substrat de la lipogènesi i inhibidor de la lipòlisi.
4

Metabolisme lipídic i la seva regulació per AMPK.
3.2. AMPK en el múscul
L’AMPK muscular s’activa durant la contracció, per la qual cosa probablement actua com a mitjancera en molts dels efectes beneficiosos de l’exercici.
• Com en el fetge, l’activació d’AMPK en el múscul augmenta l’oxidació d’àcids grassos disminuint l’activitat d’ACC1 i augmentant la de MCD.
• L’augment de la β-oxidació resulta en una menor acumulació de lípids en el múscul, la qual cosa augmenta la sensibilitat a la insulina.
• S’ha observat que l’activació d’AMPK condueix a un augment dels transportadors de glucosa GLUT4 en les cèl·lules musculars, així com una major concentració de l’enzim glicolític hexoquinasa.
• AMPK incrementa l’expressió de gens mitocondrials i biogènesi mitocondrial. Aquest fet és rellevant en tant que la resistència a la insulina i les alteracions metabòliques (incapacitat per obtenir energia a partir de substrats diversos) estan associades a disfuncions mitocondrials musculars.
En definitiva, l’activació d’AMPK tant al múscul com al fetge pot desencadenar tot un espectre d’efectes metabòlics beneficiosos que podrien millorar els defectes associats a la diabetis de tipus 2 o a la síndrome metabòlica.
4. Perspectiva terapèutica d’AMPK
Veient els efectes de l’AMPK sobre el metabolisme de la glucosa i el metabolisme lipídic, l’activació d’aquest enzim facilita una estratègia terapèutica efectiva per tractar la diabetis de tipus 2 (NIDDM) i altres desordres metabòlics relacionats. No obstant, abans d’establir AMPK com a diana farmacològica s’haurien de tenir en compte certs aspectes com:
• Quina de les isoformes de l’AMPK ha de ser activada. No està clar quina isoforma o combinacions d’isoformes s’han d’activar per obtenir el màxim benefici i, a més, les concentracions tissulars de cada isoforma varien entre espècies.
5

• Efectes secundaris indesitjats. L’activació d’AMPK en l’hipotàlem condueix a un augment de la gana, i la seva inhibició a una disminució de la gana. Un altre exemple és que l’activació constitutiva d’AMPK en ratolins condueix a l’apoptosi de cèl·lules β pancreàtiques.
• Activació d’AMPK específica per teixit. Donats els possibles efectes secundaris, s’obre la possibilitat de tractar d’activar AMPK en un o diversos teixits determinats (probablement, l’opció òptima per als diabètics seria en fetge i múscul). Però activar AMPK només en aquests teixits i evitar que s’activi de manera sistèmica no és una tasca fàcil.
• Inhibició de la proliferació i creixement cel.lular. La via LKB1-AMPK actua com supressora tumoral, disminuint la proliferació i creixement de cèl.lules preneoplàstiques. Això explica que les mutacions de LKB1 afavoreixin el càncer. Paradoxalment, LKB1-AMPK també pot afavorir els tumors ja establerts, activant la reprogramació metabòlica de les cèl.lules canceroses.
6

mTOR I ENVELLIMENT
Del treball: “A New path to longevity” David Stipp, Scientific American 306 (1): 32 -39, 2012.
Al novembre de 1964 un grup de 38 científics liderats pel professor Stanley Skoryna van viatjar fins la illa de Pasqua, situada a 2200 milles de Xile, amb la intenció d’estudiar la flora, fauna i la seva població. L’equip de Skoryna va recollir diverses mostres, de les quals va destacar un tub amb terra que contenia un bacteri (Streptomyces hygroscopicus) que produïa una substància química que varen demostrar prolongava la vida a diverses espècies.
Aquesta substància, la Rapamicina, incrementa l’esperança de vida màxima en ratolins de laboratori. Durant molts anys, els gerontòlegs han volgut descobrir compostos antienvelliment. Van descobrir que mutacions gèniques en animals poden fer més llarga la seva vida (gens de la superòxid dismutasa, receptor d’IGF-1...), i que la restricció calòrica produïa els mateixos efectes en moltes espècies.
Al 2006, es va demostrar que el Resveratrol, ingredient que conté el vi negre, semblava un bon fàrmac antienvelliment perquè reduïa els efectes negatius de les dietes riques en greixos en ratolins. Es pensa que això ho aconseguia actuant sobre uns enzims anomenats sirtuïnes (histones desacetilases). No obstant, va fallar al aplicar-se a ratolins amb dietes normals. Al 2009, els científics van demostrar que la Rapamicina, que fins llavors es coneixia com inhibidora de la proliferació cel·lular, augmentava la vida màxima en un 12% en ratolins i la supervivència fins a un terç en ratolins d’edat avançada.
La rapamicina actua inhibint una proteïna anomenada TOR (target of Rapamycin) i el seu gen. TOR ha estat objecte d’estudi perquè s’ha suggerit que la supressió de la versió en mamífers (mTOR) podria disminuir el risc de malalties relacionades amb l’edat, com Parkinson, Alzheimer, càncer, diabetis tipus 2, osteoporosi, degeneració muscular cardíaca, etc.
Història de TOR
La rapamicina, nomenada així per la tribu que viu a l’illa de Pasqua: els Rapa Nui, es va descobrir que podia disminuir la proliferació de cèl·lules immunitàries, la qual cosa va permetre disminuir el rebuig que es genera en els trasplantaments. La Rapamicina inhibeix el sistema immune bloquejant la transició de la fase G1 a S en els limfòcits T. Aquest efecte ha permès el seu ús com immunosupressors després dels transplants d’òrgans. Al 1980, es va observar que la droga inhibia el creixement tumoral i des de 2007 derivats de la rapamicina són usats per combatre alguns tipus de càncer.
Al 1991, Michael N. Hall va identificar que la rapamicina inhibeix els efectes de dos gens del llevat relacionats amb el creixement: TOR1 i TOR2. Més tard, es van arribar a aïllar els gens TOR en mamífers, plantes i insectes. Els gens TOR codifiquen un enzim que s’uneix amb altres proteïnes citoplasmàtiques per formar un complex anomenat TORC1, que supervisa moltes activitats cel·lulars relacionades amb el creixement cel.lular i metabolisme. També hi ha un altre complex anomenat TORC2.
mTOR és una serina/treonina proteïna cinasa que regula la proliferació, motilitat, supervivència, síntesi de proteïnes, transcripció i metabolisme. mTOR pertany a la família de proteïna cinases relacionades amb les fosfatidilinositol 3-kinases.
7
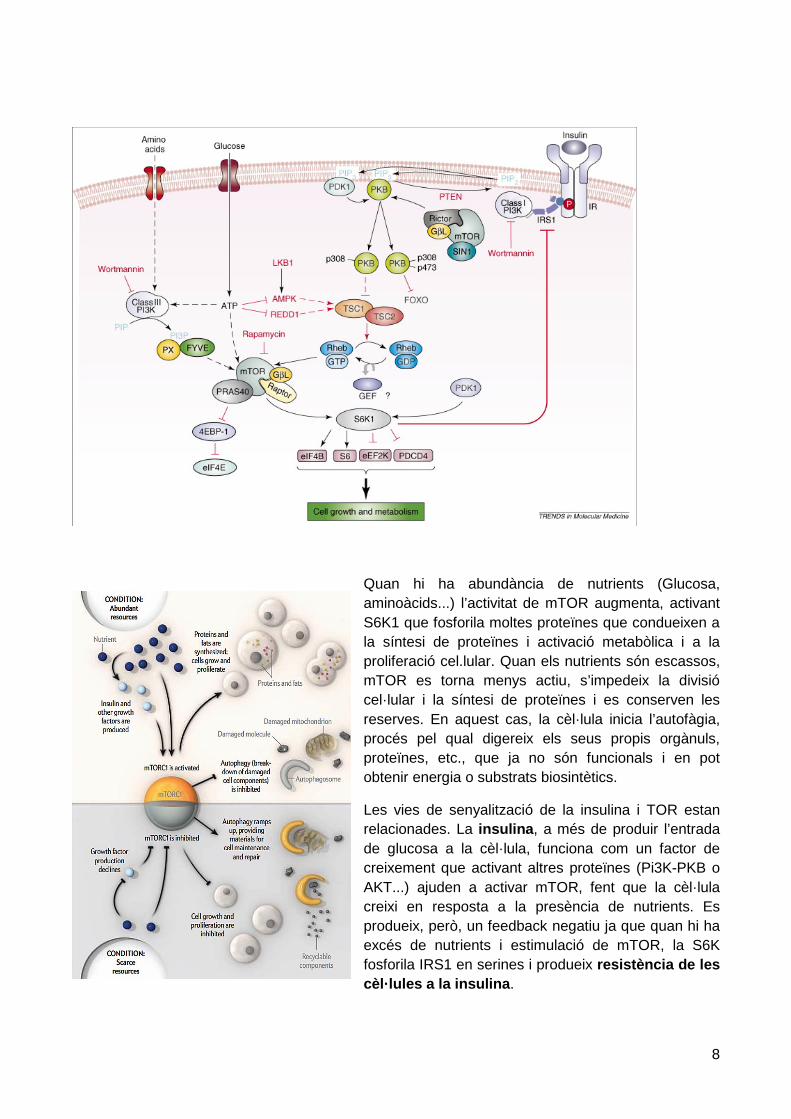
Quan hi ha abundància de nutrients (Glucosa, aminoàcids...) l’activitat de mTOR augmenta, activant S6K1 que fosforila moltes proteïnes que condueixen a la síntesi de proteïnes i activació metabòlica i a la proliferació cel.lular. Quan els nutrients són escassos, mTOR es torna menys actiu, s’impedeix la divisió cel·lular i la síntesi de proteïnes i es conserven les reserves. En aquest cas, la cèl·lula inicia l’autofàgia, procés pel qual digereix els seus propis orgànuls, proteïnes, etc., que ja no són funcionals i en pot obtenir energia o substrats biosintètics.
Les vies de senyalització de la insulina i TOR estan relacionades. La insulina, a més de produir l’entrada de glucosa a la cèl·lula, funciona com un factor de creixement que activant altres proteïnes (Pi3K-PKB o AKT...) ajuden a activar mTOR, fent que la cèl·lula creixi en resposta a la presència de nutrients. Es produeix, però, un feedback negatiu ja que quan hi ha excés de nutrients i estimulació de mTOR, la S6K fosforila IRS1 en serines i produeix resistència de les cèl·lules a la insulina.
8

Per tant, una alimentació excessiva comporta una estimulació de mTOR, que pot arribar a produir resistència a insulina.
TOR també respon a situacions estressants com hipòxia i dany cel·lular. En general, davant de dany cel·lular l’activitat de TOR es veu frenada, per tal de que la cèl·lula pugui dedicar tots els seus recursos en la reparació, com per exemple produint proteïnes mitocondrials i així ajudant a recuperar els seus sistemes energètics.
Accions de mTOR:
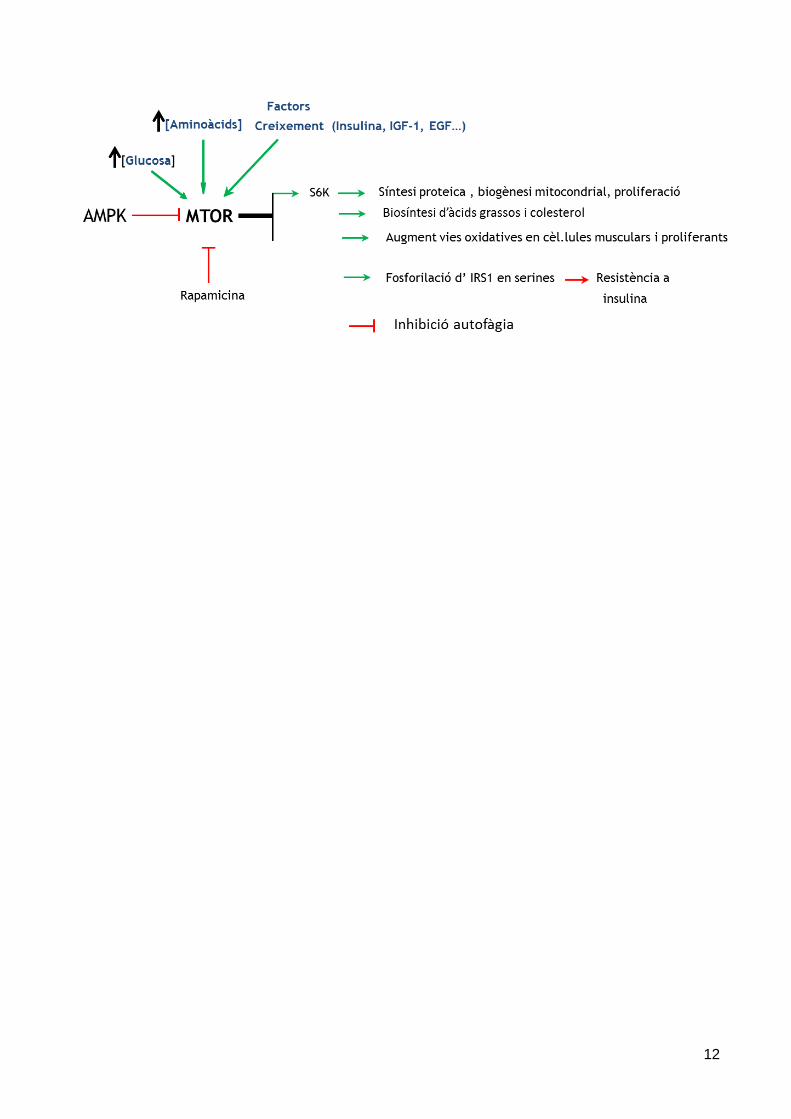
En general, mTOR s’oposa a bona part de les accions de l’AMPK. Augmenta els processos que consumeixen energia (transcripció, síntesi de proteïnes, biogènesi mitocondrial, proliferació) i també metabòlics, com biosíntesi de lípids (àcids grassos i colesterol). Per dur a terme aquests processos requereix abundància de nutrients (aminoàcids –Leucina- i glucosa) i activació de les vies d’obtenció d’energia (glicòlisi, via pentoses, COT i CRM). L’autofàgia resta frenada. Cal recordar que AMPK inhibeix mTOR (fosforila i activaTSC2 i inhibeix Raptor).
Aminoàcids Glucosa
9

1. La connexió de TOR amb l’envelliment S’ha demostrat que la restricció calòrica retarda l’envelliment en ratolins, mosques.... Reduint un terç la ingesta de calories diàries es pot augmentar un 30-40% la vida màxima. Estudis en cucs i mosques de la fruita han demostrat que suprimint TOR s’obtenen efectes semblants als de la restricció calòrica: augmentar el període de vida màxim.
Però TOR no només mimetitza els efectes de la restricció calòrica, sinó també els dels gerontogens, gens coneguts que augmenten la longevitat. Alguns d’aquests gerontogens tenen una mutació que condueix a un bloqueig de les senyals de creixement, com les mutacions en el receptor del IGF1 (insulin growth factor 1). Aquesta mutació provocaria que els animals augmentessin la seva esperança de vida màxima.
El 2009, es va fer un estudi que concloïa que la rapamicina augmentava l’esperança de vida en un 28% en ratolins mascles i un 38% en ratolins femelles. Un altre estudi reportava que la inhabilitació del gen S6K1, que codifica l’enzim (S6Kinase) activat per mTOR, feia els ratolins resistents a malalties relacionades amb l’edat i augmentava la seva esperança de vida màxima.
La possible causa d’aquest augment de longevitat és que, al inhibir mTOR, augmenta l’autofàgia (la qual cosa permet renovar orgànuls envellits) i disminueix la proliferació cel·lular (reduint així mutacions i dany cel·lular associats a aquesta proliferació). Aquests processos retarden l’envelliment dels teixits.
2. L’enigma evolutiu de mTOR L’existència d’un sistema que funcioni com antienvelliment cel·lular no té cabuda per als biòlegs evolucionistes, ja que s’oposaria a la selecció natural. Una de les solucions a aquest enigma seria que la restricció calòrica activés una resposta evolutiva enfront la fam, disminuint l’envelliment dels organismes joves per tal que puguin reproduir-se en temps d’abundància de nutrients.
Hi ha una altra teoria descrita per Blagosklonny, que postula que mTOR, essencial pel desenvolupament i reproducció, funciona com un agent d’envelliment una vegada s’ha acabat l’etapa reproductiva. Això faria augmentar la proliferació de múscul llis a les arteries (un pas clau en l’arterioesclerosi), l’acumulació de greix, el desenvolupament de resistència a insulina, multiplicació
10

d’osteoclasts i creixement de tumors. Al disminuir l’autofàgia (que es dóna quan mTOR està actiu) es crearien radicals lliures que provocarien danys en el DNA i els mitocondris, afectant al metabolisme energètic cel·lular. A més, mTOR contribuiria a l’acumulació de proteïnes de difícil degradació en les cèl.lules (neurones...), produint les malalties neurodegeneratives.
Per tant, els efectes de la inhibició de mTOR serien merament accidents de la natura i, en realitat, mTOR actuaria com un mecanisme proliferatiu cel·lular durant la joventut en fases d’abundància d’aliments, però com a agent envellidor a partir de la maduresa.
3. mTOR en Medicina Malgrat els efectes beneficiosos de la rapamicina, cal destacar que també incrementa el colesterol plasmàtic, causa anèmia i interfereix en la cicatrització de les ferides, a més dels efectes degeneratius esmentats.
na alternativa a la rapamicina podria ser la metformina, prescrita a diabètics per disminuir la glucosa en sang. Tot i que no està ben entès, es coneix que inhibeix la via de mTOR i activa l’enzim AMPK, que promou la resposta a l’estrès de les cèl·lules. Alhora, AMPK també inhibeix mTOR. També s’ha mostrat que emula els efectes de la restricció calòrica en l’activació gènica.
Si els efectes de la rapamicina a ratolins es poguessin aplicar a humans, augmentaria la nostra esperança de vida màxima uns 10 anys. No només això, es podrien prevenir malalties tals com osteoporosi, Alzheimer, càncer etc., que representen una despesa important en matèria de sanitat. Però el desenvolupament d’una droga així no serà fàcil.
Cal recordar que l’exercici, via AMPK, o la restricció calòrica poden modular la via de mTOR.
A continuació es resumeixen els efectes i interaccions d’AMPK i mTOR.
11
12