alterations in neurobehaviors and inflammation in hippocampus of rats induced by oral administration...
TRANSCRIPT

RESEARCH ARTICLE
Alterations in neurobehaviors and inflammation in hippocampusof rats induced by oral administration of microcystin-LR
Xiao-Bo Li & Xin Zhang & Jingjuan Ju & Yunhui Li &Lihong Yin & Yuepu Pu
Received: 27 March 2014 /Accepted: 3 June 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Microcystin-LR (MC-LR) is a widely studied toxicpeptide secreted by certain water blooms of cyanobacteria thatexhibit hepatotoxicity and neural toxicity. This study aimed toobserve the neurotoxic effects of low-dose MC-LR exposureby oral administration. Male Sprague-Dawley (SD) rats wereadministered orally every 2 days for 8 weeks with pure waterand 0.2, 1.0, and 5.0 μg/kg MC-LR. The Morris water mazetest was used to assess the spatial learning and memorycapability of rats. The activation of astrocytes and nitric oxidesynthase (NOS) was evaluated by immunohistochemistry, andconcentrations of nitric oxide (NO) in rat hippocampus wereanalyzed. Slight liver dysfunction was observed in the 5.0 μg/kg MC-LR-treated rats. Impairment of spatial learning andmemory was also observed in the 5.0 μg/kg MC-LR-treatedrats. Astrocytes in the hippocampus of the 5.0 μg/kg MC-LR-treated rats showed enhanced activation and cell density; theinflammatory indicators, NOS and NO, increased in accor-dance with astrocyte activation. This study showed that oralexposure of MC-LR had adverse affects on neurobehaviors,and induced inflammation in memory-related brain regions.
Keywords Microcystin-LR . Neurobehavior . Astrocytes .
Nitric oxide . Inflammation .Morris water maze
Introduction
Microcystin-LR (MC-LR) is a monocyclic peptide toxin pro-duced by blue-green algae proliferating in eutrophic
freshwater bodies and usually found in more than half of thefreshwater bloom (Graham et al. 2010). The concentration ofdissolved MCs in eutrophic water body varies from 0.1 to20 μg/L (Xu et al. 2008; Ye et al. 2009). This toxic peptidetests positive in aquatic plants (Mitrovic et al. 2005), fish(Schmidt et al. 2013), shrimp (Papadimitriou et al. 2012),turtles, ducks, and water birds (Chen et al. 2009) after out-break of cyanobacterial blooms. Thus, this phenomenon in-creases the exposure risk of humans through the gastrointes-tinal tract, not only fromMC-LR-contaminated water but alsofrom MC-LR-containing food.
The major toxic effect of MC-LR is hepatotoxicity, butpotential neurotoxicity has also been reported by several stud-ies. This toxic chemical may enter through the brain bloodbarrier, which is mediated by organic anion-transporting poly-peptides (Fischer et al. 2005). Intrahippocampal injection ofMC-LR can impair the neurobehavior and histology of ratbrains (Li et al. 2014). An in vivomodel showed that exposureto 0.1 μg−1 MC-LR results in the loss of GABAergic toincrease significantly in Caenorhabditis elegans (Ju et al.2013; Li et al. 2009a).
Astrocytes are subtype of glial cells that exert manyessential complex functions in the central nervous system(CNS). Reactive astrocytes respond to all forms of CNSinjury or disease, which has become a pathological hall-mark of CNS structural lesions (Sofroniew and Vinters2009). The reactive astrocytes can produce various in-flammatory cytokines and chemokines, which are specu-lated to be involved in reparative processes at earlystages of neuroinflammation (Sharma et al. 2007; Choi et al.2002). However, several inflammatory mediators andchemokines secreted by astrocytes have the potential tocontribute to inflammation-mediated damage of the CNS(Meeuwsen et al. 2003; Williams et al. 2009). NO is oneof the inflammatory mediators derived from astrocytes(Stewart et al. 2000; Buskila et al. 2007).
Responsible editor: Philippe Garrigues
X.<B. Li (*) :X. Zhang : J. Ju :Y. Li : L. Yin :Y. PuKey Laboratory of Environmental Medicine Engineering, Ministryof Education, School of Public Health, Southeast University,Dingjiaqiao 87, Nanjing 210009, Chinae-mail: [email protected]
Environ Sci Pollut ResDOI 10.1007/s11356-014-3151-x

NO is a gaseous free radical synthesized from L-arginine bynitric oxide synthase (NOS) and produced by neurons, astro-cytes, and endothelial cells (Buskila et al. 2007). NOS havethree different isoforms, and their activity depends on the celltype involved. NOS1-expressing neurons can rapidly releasesmall amounts of NO functioning as neurotransmitters(Boehning and Snyder 2003; Prast and Philippu 2001).NOS2 can synthesize large amounts of NO, which is derivedby astrocytes on a slower time scale. Astrocyte-derived NOhas been demonstrated as a reaction to stimuli, such as in-flammation (LopezLopez-Figueroa et al. 2000; Moro et al.2004; Murphy 2000). This NOS2 synthesized NO destroysneurons by mitochondrial dysfunction (Stewart et al. 2000;Bolanos and Heales 2010).
This study investigated the effects of MC-LR exposure onthe spatial learning and memory capacity of rats, as well as thepotential mechanisms associated with reactive astrocytes andinflammatory mediator NO.
Methods
Animals and chemicals
SD rats (28 days old) were purchased from Shanghai SLRCLaboratory Animal Co. Ltd., China. MC-LR (purity≥95 %)was obtained from Enzo Life Sciences International, USA(ALX-350-012). MC-LR (1 mg) was dissolved in 1 mL ofmethanol and diluted to 100 μg/mL with pure water as stocksolution and diluted to 0.2, 1.0, and 5.0 μg/mL with purewater before application. Monoclonal antibody of N-20 andglial fibrillary acidic protein (GFAP) were purchased fromBeijing Zhongshan Golden Bridge Biotechnology Co Ltd.,China (ZS-651) and Sigma–Aldrich Inc., USA (G9269),respectively.
Animal treatment
SD rats were housed in the animal facility of the SoutheastUniversity, China. Food and water were available ad libitumunder a 12-h dark/light cycle. All procedures were conductedin conformity with approved institutional protocols. Male rats(28 days old) were divided into four groups (with eight rats ineach group), namely, the vehicle control (pure water) and 0.2,1.0, and 5.0 μg/kg MC-LR by intragastric administrationevery 2 days for 8 weeks. The weights of rats were measuredevery 2 days.
Morris water maze test
The Morris water maze test was carried out 24 h after the endof MC-LR administration to measure the spatial learning andmemory abilities of rats. A 1.6-m diameter water maze was
filled with water to a depth of 26.5 cm, and the water temper-ature was adjusted to 26±0.5 °C. The circle platform, whichwas 25 cm high and 12 cm in diameter, was placed at a fixedlocation in the center of one of the four imaginary quadrants.Ink (20mL) was used tomake the water opaque. The rats werereleased into the water at one of the three different startingpositions and allowed to swim for up to 90 s to locate theplatform. Escape latencies were defined as the time betweenthe start of swimming and climbing the platform. Distancetraveled, swim speed, and escape latency were measured oneach trial. On day 6, we performed a probe trial starting fromthe opposite of the platform quadrants and removed the hid-den platform.We further assessed the time periods spent in thecircle segments, frequencies in the platform zone, enlargedplatform zone (Loureiro et al. 2012), velocity, and total dis-tance moved. The enlarged platform zone, which was 18 cmin diameter, was defined as the circle with the same centerpoint with the platform.
Serum biochemical parameter analysis
Animals were deeply euthanized under ether anesthesia 24 hafter the behavioral studies. Blood was drawn from the orbitalsinus of each animal. Serum samples were obtained from thecollected blood samples by incubation at room temperaturefor 30 min and centrifugation at 3,000 rpm for 10 min. Theobtained serum samples were subjected to serum aspartateaminotransaminase (AST), alanine amino transferase (ALT),alkaline phosphatase (ALP), cholinesterase (ChE), total pro-tein (TP), and albumin (ALB) concentration assay using anautomated clinical chemistry analyzer (Selectra Junior, VitalScientific, The Netherlands).
Histopathological analysis
Animals were deeply euthanized under ether anesthesia 24 hafter the behavioral studies. Four rats from each group wereperfused intracardially with 4 % paraformaldehyde (PFA) inphosphate-buffered saline (PBS) at 4 °C. Brains and livers ofrats were immediately removed, post-fixed in 4 % PFA over-night at 4 °C, embedded in paraffin, serially sectioned (5 μm),and mounted on silane-covered slides. After dewaxing, thesections selected at the same layers from each rat were stainedwith hematoxylin and eosin (HE) and evaluated under a lightmicroscope (×400) to examine the morphological alterationsin neurons in the hippocampal region and histology of theliver. Undamaged neurons were defined as those with round-or oval-shaped nuclei and without shrinkage or edema. Thenormal histology of liver is composed of innumerable lobules,each of which is a hexagonal structure consisting of a centralvein surrounded by radiating hepatocyte plates.
Environ Sci Pollut Res
Immunohistochemistry staining
After dewaxing, sections were boiled in an 850-Wmicrowaveoven for 15 min in citrate buffer (2.1 g of citric acidmonohydrate/L, pH 6) (Carl Roth). Endogenous peroxidasewas inhibited by 1 % hydrogen peroxide in pure methanol(Merck) for 15 min. Sections were incubated with 10 %normal pig serum in Tris-buffered saline (Biochrom, Berlin,Germany) to block nonspecific binding of immunoglobulins,and then with mouse monoclonal antibodies GFAP (1:80) orN-20 (1:50) overnight at 4 °C. Antibody binding to tissuesections was visualized with a biotinylated rabbit anti-mouseIgG F(ab)2 antibody fragment (1:400; DAKO). Subsequently,sections were incubated with a horseradish peroxidase-conjugated streptavidin complex for 30 min (1:100; DAKO)and developed by diaminobenzidine substrate (Fluka). Final-ly, sections were counterstained with Mayer’s hemalum. Re-garding the negative controls, the primary antibodies wereomitted.
After immunostaining, the brain sections were examinedunder light microscopy by three histologists blinded to treat-ment conditions. The immunoreactivities of GFAP and iNOSwere evaluated in the hippocampus. The number of GFAP+
and iNOS+ cells of each rat brain section was counted in threenonoverlapping high-power fields (HPFs; ×400 magnifica-tion) and analyzed. The HPFs were selected from hippocam-pal areas that had a maximum of positive cells. In each studiedfield, only positive cells with the nucleus at the focal planewere counted.
NO concentration assay
After anesthesia was administered, four rats from each groupwere decapitated on an iced table. Brain tissues were collectedand washed in ice-cold saline, and hippocampi were separatedand stored in liquid nitrogen. Prior to assay, the hippocampiwere homogenized in ice-cold 50 mM Tris–HCl buffer(pH 7.55). The homogenate was centrifuged for 15 min at3,000 rpm, and the supernatant was used for NO concentrationassay by nitrate reductase methods according to kit protocols(Nanjing Jiancheng Bioengineering Institute).
Statistical analyses
Indicators of the probe trial were analyzed by ANOVA,followed by student’s t test for parametric comparison be-tween groups. Repeated measures ANOVA was conductedto compare the changes in escape latencies for the Morriswater maze test. The immunostaining results were statisticallyanalyzed by ANOVA, followed by student’s t test or nonpara-metric t test statistics. Results are shown as arithmetic meansand standard errors of means. SPSS 12.0 software was used
for statistical analysis, and the significance level was set atP<0.05.
Results
Effects on learning and memory ability
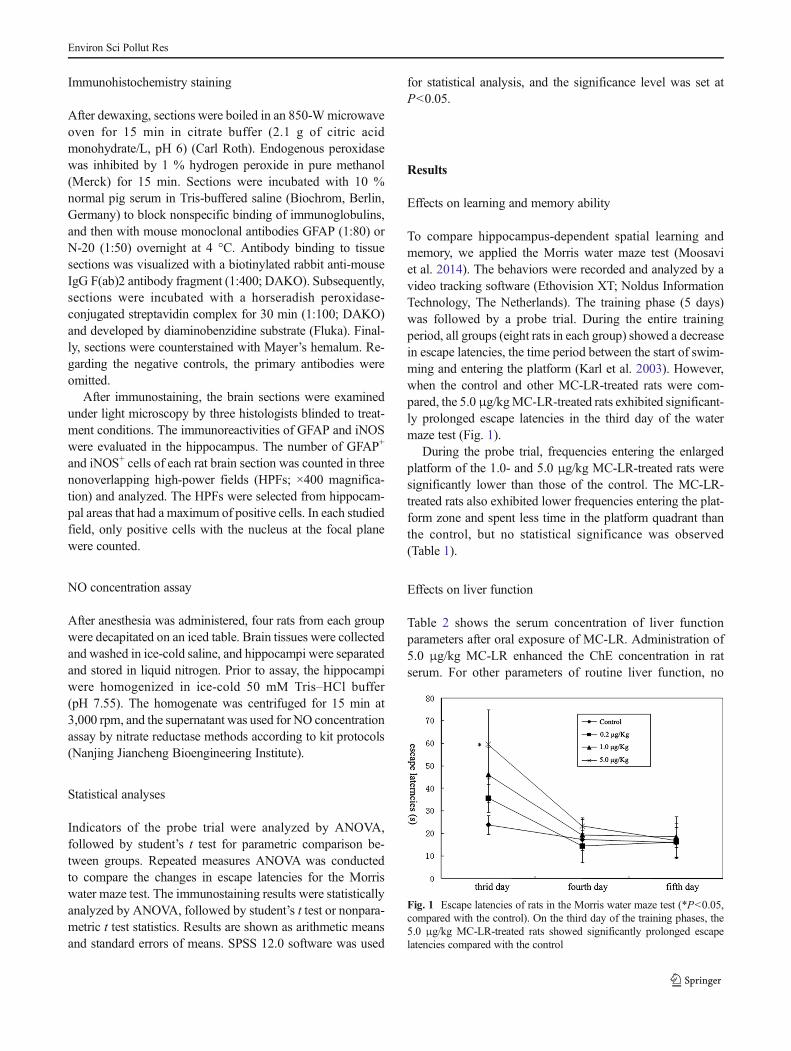
To compare hippocampus-dependent spatial learning andmemory, we applied the Morris water maze test (Moosaviet al. 2014). The behaviors were recorded and analyzed by avideo tracking software (Ethovision XT; Noldus InformationTechnology, The Netherlands). The training phase (5 days)was followed by a probe trial. During the entire trainingperiod, all groups (eight rats in each group) showed a decreasein escape latencies, the time period between the start of swim-ming and entering the platform (Karl et al. 2003). However,when the control and other MC-LR-treated rats were com-pared, the 5.0 μg/kgMC-LR-treated rats exhibited significant-ly prolonged escape latencies in the third day of the watermaze test (Fig. 1).
During the probe trial, frequencies entering the enlargedplatform of the 1.0- and 5.0 μg/kg MC-LR-treated rats weresignificantly lower than those of the control. The MC-LR-treated rats also exhibited lower frequencies entering the plat-form zone and spent less time in the platform quadrant thanthe control, but no statistical significance was observed(Table 1).
Effects on liver function
Table 2 shows the serum concentration of liver functionparameters after oral exposure of MC-LR. Administration of5.0 μg/kg MC-LR enhanced the ChE concentration in ratserum. For other parameters of routine liver function, no
Fig. 1 Escape latencies of rats in the Morris water maze test (*P<0.05,compared with the control). On the third day of the training phases, the5.0 μg/kg MC-LR-treated rats showed significantly prolonged escapelatencies compared with the control
Environ Sci Pollut Res
significant alterations were observed between the control andtreated groups.
Pathological examination of brain and liver tissues
The HE staining sections were evaluated by an examinerblinded to experimental conditions. Several neurons withshrinkage or edema were observed both in the control andMC-LR-treated groups, but the number of “damaged” neu-rons was not significant. No pathological alterations, such asedema and inflammation, were observed in the liver section.
Astrocyte activation in hippocampus
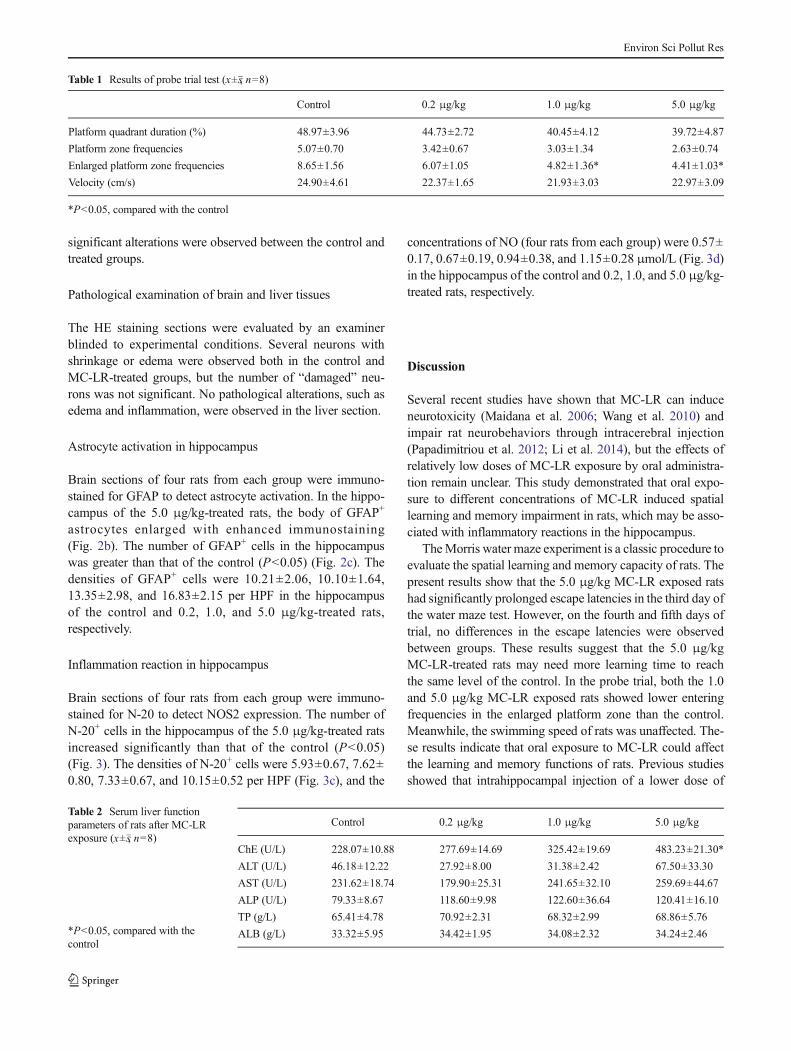
Brain sections of four rats from each group were immuno-stained for GFAP to detect astrocyte activation. In the hippo-campus of the 5.0 μg/kg-treated rats, the body of GFAP+
astrocytes enlarged with enhanced immunostaining(Fig. 2b). The number of GFAP+ cells in the hippocampuswas greater than that of the control (P<0.05) (Fig. 2c). Thedensities of GFAP+ cells were 10.21±2.06, 10.10±1.64,13.35±2.98, and 16.83±2.15 per HPF in the hippocampusof the control and 0.2, 1.0, and 5.0 μg/kg-treated rats,respectively.
Inflammation reaction in hippocampus
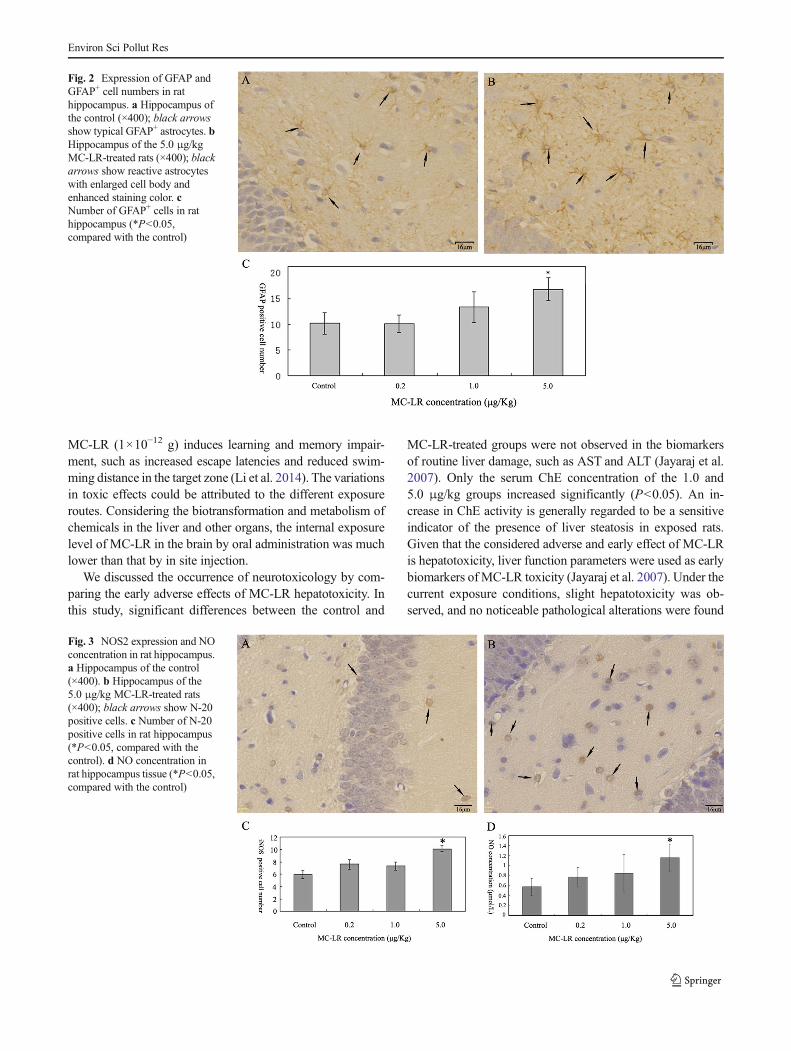
Brain sections of four rats from each group were immuno-stained for N-20 to detect NOS2 expression. The number ofN-20+ cells in the hippocampus of the 5.0 μg/kg-treated ratsincreased significantly than that of the control (P<0.05)(Fig. 3). The densities of N-20+ cells were 5.93±0.67, 7.62±0.80, 7.33±0.67, and 10.15±0.52 per HPF (Fig. 3c), and the
concentrations of NO (four rats from each group) were 0.57±0.17, 0.67±0.19, 0.94±0.38, and 1.15±0.28 μmol/L (Fig. 3d)in the hippocampus of the control and 0.2, 1.0, and 5.0 μg/kg-treated rats, respectively.
Discussion
Several recent studies have shown that MC-LR can induceneurotoxicity (Maidana et al. 2006; Wang et al. 2010) andimpair rat neurobehaviors through intracerebral injection(Papadimitriou et al. 2012; Li et al. 2014), but the effects ofrelatively low doses of MC-LR exposure by oral administra-tion remain unclear. This study demonstrated that oral expo-sure to different concentrations of MC-LR induced spatiallearning and memory impairment in rats, which may be asso-ciated with inflammatory reactions in the hippocampus.
TheMorris water maze experiment is a classic procedure toevaluate the spatial learning and memory capacity of rats. Thepresent results show that the 5.0 μg/kg MC-LR exposed ratshad significantly prolonged escape latencies in the third day ofthe water maze test. However, on the fourth and fifth days oftrial, no differences in the escape latencies were observedbetween groups. These results suggest that the 5.0 μg/kgMC-LR-treated rats may need more learning time to reachthe same level of the control. In the probe trial, both the 1.0and 5.0 μg/kg MC-LR exposed rats showed lower enteringfrequencies in the enlarged platform zone than the control.Meanwhile, the swimming speed of rats was unaffected. The-se results indicate that oral exposure to MC-LR could affectthe learning and memory functions of rats. Previous studiesshowed that intrahippocampal injection of a lower dose of
Table 1 Results of probe trial test (x±s, n=8)
Control 0.2 μg/kg 1.0 μg/kg 5.0 μg/kg
Platform quadrant duration (%) 48.97±3.96 44.73±2.72 40.45±4.12 39.72±4.87
Platform zone frequencies 5.07±0.70 3.42±0.67 3.03±1.34 2.63±0.74
Enlarged platform zone frequencies 8.65±1.56 6.07±1.05 4.82±1.36* 4.41±1.03*
Velocity (cm/s) 24.90±4.61 22.37±1.65 21.93±3.03 22.97±3.09
*P<0.05, compared with the control
Table 2 Serum liver functionparameters of rats after MC-LRexposure (x±s, n=8)
*P<0.05, compared with thecontrol
Control 0.2 μg/kg 1.0 μg/kg 5.0 μg/kg
ChE (U/L) 228.07±10.88 277.69±14.69 325.42±19.69 483.23±21.30*
ALT (U/L) 46.18±12.22 27.92±8.00 31.38±2.42 67.50±33.30
AST (U/L) 231.62±18.74 179.90±25.31 241.65±32.10 259.69±44.67
ALP (U/L) 79.33±8.67 118.60±9.98 122.60±36.64 120.41±16.10
TP (g/L) 65.41±4.78 70.92±2.31 68.32±2.99 68.86±5.76
ALB (g/L) 33.32±5.95 34.42±1.95 34.08±2.32 34.24±2.46
Environ Sci Pollut Res
MC-LR (1×10−12 g) induces learning and memory impair-ment, such as increased escape latencies and reduced swim-ming distance in the target zone (Li et al. 2014). The variationsin toxic effects could be attributed to the different exposureroutes. Considering the biotransformation and metabolism ofchemicals in the liver and other organs, the internal exposurelevel of MC-LR in the brain by oral administration was muchlower than that by in site injection.
We discussed the occurrence of neurotoxicology by com-paring the early adverse effects of MC-LR hepatotoxicity. Inthis study, significant differences between the control and
MC-LR-treated groups were not observed in the biomarkersof routine liver damage, such as AST and ALT (Jayaraj et al.2007). Only the serum ChE concentration of the 1.0 and5.0 μg/kg groups increased significantly (P<0.05). An in-crease in ChE activity is generally regarded to be a sensitiveindicator of the presence of liver steatosis in exposed rats.Given that the considered adverse and early effect of MC-LRis hepatotoxicity, liver function parameters were used as earlybiomarkers of MC-LR toxicity (Jayaraj et al. 2007). Under thecurrent exposure conditions, slight hepatotoxicity was ob-served, and no noticeable pathological alterations were found
Fig. 2 Expression of GFAP andGFAP+ cell numbers in rathippocampus. a Hippocampus ofthe control (×400); black arrowsshow typical GFAP+ astrocytes. bHippocampus of the 5.0 μg/kgMC-LR-treated rats (×400); blackarrows show reactive astrocyteswith enlarged cell body andenhanced staining color. cNumber of GFAP+ cells in rathippocampus (*P<0.05,compared with the control)
Fig. 3 NOS2 expression and NOconcentration in rat hippocampus.a Hippocampus of the control(×400). b Hippocampus of the5.0 μg/kg MC-LR-treated rats(×400); black arrows show N-20positive cells. c Number of N-20positive cells in rat hippocampus(*P<0.05, compared with thecontrol). d NO concentration inrat hippocampus tissue (*P<0.05,compared with the control)
Environ Sci Pollut Res

in liver histology, indicating that the impairment of learningandmemory functions appeared at the very early stage ofMC-LR toxic effects.
The hippocampus is a structure predominantly involved inspatial navigation (Stella et al. 2012). We further assessed thepathological alteration in the hippocampus to investigate thepotential mechanism of neurobehavior impairment. No no-ticeable neuronal damage was observed in the MC-LR-treatedhippocampus by histological analysis. Li et al. reported histo-logical changes in neurons of the CA1 region induced byMC-LR (Papadimitriou et al. 2012; Li et al. 2014). We speculatedthat direct intrahippocampal injection induced more severedamage to brain histology than oral exposure.
Astrocytes are sensitive to alterations in the CNS micro-environment and rapidly activate in all conditions that affectnormal neuronal functions (Li et al. 2009b). The enhancedactivation of astrocytes is induced by different environmentalneurotoxic chemicals (Rai et al. 2010; Bai et al. 2014). Reac-tive astrocytes are key pathological features, and astrocyte-derived NO possesses critical functions in CNS inflammatoryreactions (Hamby and Sofroniew 2010). In the present study,hyperplasia of astrocytes and the positive expression of GFAPin rat hippocampus, which are indicators of astrocyte activa-tion, were demonstrated after MC-LR exposure. Astrocyte-derived NO differs from neuron-derived NO because it existsin large amounts and is synthesized by NOS2 (Buskila et al.2007). MC-LR may affect the expression of NOS2 in astro-cytes in vitro because of the inhibition of the functions ofprotein phosphatase 1 and 2A (Pahan et al. 1998). Therefore,we analyzed the expression of GFAP and NOS2, as well asNO concentration, in brains of the control andMC-LR-treatedrats. In the hippocampus of the 5.0 μg/kgMC-LR treated rats,significant accumulations of GFAP+ and NOS2+ cells wereobserved. Moreover, the NO concentration in the hippocam-pal tissues increased in accordance with NOS2+ cells. Theseresults show enhanced inflammatory reactions induced byMC-LR in rat brains. Endogenous NO is involved in spatialmemory stages (Moosavi et al. 2014; Arnaiz et al. 2004), sothe increase in NO level could be considered associated withspatial memory impairment.
In brief, the results demonstrate that exposure to MC-LRimpaired the spatial learning and memory capacity of rats,which appeared at the very early stage of MC-LR toxiceffects, and produced significant inflammatory effects in therat brain. In addition to inflammation, oxidative stress is also amajor mechanism of MC-LR-induced toxicity (Sedan et al.2013; Meng et al. 2013). Thus, the neurotoxic effects ofenvironmental pollutant MC-LR should be considered to pro-tect the health of susceptible populations.
Acknowledgments This study was funded by the National NaturalScience Foundation of China (Nos. 81072258 and 30972440), NaturalScience Foundation of Jiangsu Province (No. BK2010402), Research and
Innovation Project for College Graduates of Jiangsu Province (No.CXLX_0156) and supported by the Fundamental Research Funds for theCentral Universities.
References
Arnaiz SL, D'Amico G, Paglia N, Arismendi M, Basso N, del RosarioLores Arnaiz M (2004) Enriched environment, nitric oxide produc-tion and synaptic plasticity prevent the aging-dependent impairmentof spatial cognition. Mol Aspects Med 25:91–101
Bai R, Zhang L, Liu Y, Li B, Wang L, Wang P et al (2014) Integratedanalytical techniques with high sensitivity for studying brain trans-location and potential impairment induced by intranasally instilledcopper nanoparticles. Toxicol Lett 226:70–80
Boehning D, Snyder SH (2003) Novel neural modulators. Annu RevNeurosci 26:105–131
Bolanos JP, Heales SJ (2010) Persistent mitochondrial damage by nitricoxide and its derivatives: neuropathological implications. FrontNeuroenergetics 2:1
Buskila Y, Abu-Ghanem Y, Levi Y, Moran A, Grauer E, Amitai Y (2007)Enhanced astrocytic nitric oxide production and neuronal modifica-tions in the neocortex of a NOS2 mutant mouse. PLoS One 2:e843
Chen J, Zhang D, Xie P, Wang Q, Ma Z (2009) Simultaneous determi-nation of microcystin contaminations in various vertebrates (fish,turtle, duck and water bird) from a large eutrophic Chinese lake,Lake Taihu, with toxic Microcystis blooms. Sci Total Environ 407:3317–3322
Choi JS, Jung SW, Ju JC, Lee SW, Kim KY, Kim HM (2002) Cytokineproduction regulation in human astrocytes by a herbal combination(Yuldahansotang). Immunopharmacol Immunotoxicol 24:55–67
Fischer WJ, Altheimer S, Cattori V, Meier PJ, Dietrich DR, Hagenbuch B(2005) Organic anion transporting polypeptides expressed in liverand brain mediate uptake of microcystin. Toxicol Appl Pharmacol203:257–263
Graham JL, Loftin KA, Meyer MT, Ziegler AC (2010) Cyanotoxinmixtures and taste-and-odor compounds in cyanobacterial bloomsfrom the Midwestern United States. Environ Sci Technol 44:7361–7368
Hamby ME, Sofroniew MV (2010) Reactive astrocytes as therapeutictargets for CNS disorders. Neurotherapeutics 7:494–506
Jayaraj R, Deb U, Bhaskar AS, Prasad GB, Rao PV (2007)Hepatoprotective efficacy of certain flavonoids against microcystininduced toxicity in mice. Environ Toxicol 22:472–479
Ju J, Ruan Q, Li X, Liu R, Li Y, Pu Y et al (2013) Neurotoxicologicalevaluation of microcystin-LR exposure at environmental relevantconcentrations on nematode Caenorhabditis elegans. Environ SciPollut Res Int 20:1823–1830
Karl T, Pabst R, von Horsten S (2003) Behavioral phenotyping of mice inpharmacological and toxicological research. Exp Toxicol Pathol 55:69–83
Li Y, Ye H, Du M, Zhang Y, Ye B, Pu Y et al (2009a) Induction ofchemotaxis to sodium chloride and diacetyl and thermotaxis defectsby microcystin-LR exposure in nematode Caenorhabditis elegans. JEnviron Sci (China) 21:971–979
Li XB, Zheng H, Zhang ZR, Li M, Huang ZY, Schluesener HJ et al(2009b) Glia activation induced by peripheral administration ofaluminum oxide nanoparticles in rat brains. Nanomedicine 5:473–479
Li G, Yan W, Cai F, Li C, Chen N, Wang J (2014) Spatial learning andmemory impairment and pathological change in rats induced byacute exposure to microcystin-LR. Environ Toxicol 29:261–268
Lopez-FigueroaMO, Day HE, Lee S, Rivier C, Akil H,Watson SJ (2000)Temporal and anatomical distribution of nitric oxide synthase
Environ Sci Pollut Res

mRNA expression and nitric oxide production during central ner-vous system inflammation. Brain Res 852:239–246
LoureiroM, Cholvin T, Lopez J,Merienne N, Latreche A, Cosquer B et al(2012) The ventral midline thalamus (reuniens and rhomboid nuclei)contributes to the persistence of spatial memory in rats. J Neurosci32:9947–9959
Maidana M, Carlis V, Galhardi FG, Yunes JS, Geracitano LA, MonserratJM et al (2006) Effects of microcystins over short- and long-termmemory and oxidative stress generation in hippocampus of rats.Chem Biol Interact 159:223–234
Meeuwsen S, Persoon-Deen C, Bsibsi M, Ravid R, van Noort JM (2003)Cytokine, chemokine and growth factor gene profiling of culturedhuman astrocytes after exposure to proinflammatory stimuli. Glia43:243–253
MengG, Liu J, Lin S, GuoZ andXuL (2013)Microcystin-LR-CausedROSgeneration involved in p38 activation and tau hyperphosphorylation inneuroendocrine (PC12) cells. Environ Toxicol
Mitrovic SM, Allis O, Furey A, James KJ (2005) Bioaccumulation andharmful effects of microcystin-LR in the aquatic plants Lemnaminor and Wolffia arrhiza and the filamentous alga Chladophorafracta. Ecotoxicol Environ Saf 61:345–352
Moosavi M, Abbasi L, Zarifkar A, Rastegar K (2014) The role of nitricoxide in spatial memory stages, hippocampal ERK and CaMKIIphosphorylation. Pharmacol, Biochem Behav 122C:164–172
Moro MA, Cardenas A, Hurtado O, Leza JC, Lizasoain I (2004) Role ofnitric oxide after brain ischaemia. Cell Calcium 36:265–275
Murphy S (2000) Production of nitric oxide by glial cells: regulation andpotential roles in the CNS. Glia 29:1–13
Pahan K, Sheikh FG, Namboodiri AM, Singh I (1998) Inhibitors ofprotein phosphatase 1 and 2A differentially regulate the expressionof inducible nitric-oxide synthase in rat astrocytes and macrophages.J Biol Chem 273:12219–12226
Papadimitriou T, Kagalou I, Stalikas C, Pilidis G, Leonardos ID (2012)Assessment ofmicrocystin distribution and biomagnification in tissuesof aquatic food web compartments from a shallow lake and evaluationof potential risks to public health. Ecotoxicology 21:1155–1166
Prast H, Philippu A (2001) Nitric oxide as modulator of neuronal func-tion. Prog Neurobiol 64:51–68
Rai A, Maurya SK, Khare P, Srivastava A, Bandyopadhyay S (2010)Characterization of developmental neurotoxicity of As, Cd, and Pbmixture: synergistic action of metal mixture in glial and neuronalfunctions. Toxicol Sci 118:586–601
Schmidt JR, Shaskus M, Estenik JF, Oesch C, Khidekel R, Boyer GL(2013) Variations in the microcystin content of different fish speciescollected from a eutrophic lake. Toxins (Basel) 5:992–1009
Sedan D, Giannuzzi L, Rosso L, Marra CA, Andrinolo D (2013)Biomarkers of prolonged exposure to microcystin-LR in mice.Toxicon 68:9–17
Sharma V, Mishra M, Ghosh S, Tewari R, Basu A, Seth P et al (2007)Modulation of interleukin-1beta mediated inflammatory response inhuman astrocytes by flavonoids: implications in neuroprotection.Brain Res Bull 73:55–63
Sofroniew MV, Vinters HV (2009) Astrocytes: biology and pathology.Acta Neuropathol 119:7–35
Stella F, Cerasti E, Si B, Jezek K, Treves A (2012) Self-organization ofmultiple spatial and context memories in the hippocampus. NeurosciBiobehav Rev 36:1609–1625
Stewart VC, Sharpe MA, Clark JB, Heales SJ (2000) Astrocyte-derived nitric oxide causes both reversible and irreversibledamage to the neuronal mitochondrial respiratory chain. JNeurochem 75:694–700
Wang M, Wang D, Lin L, Hong H (2010) Protein profiles in zebrafish(Danio rerio) brains exposed to chronic microcystin-LR.Chemosphere 81:716–724
Williams R, Yao H, Dhillon NK, Buch SJ (2009) HIV-1 Tat co-operateswith IFN-gamma and TNF-alpha to increase CXCL10 in humanastrocytes. PLoS One 4:e5709
Xu Q, Chen W, Gao G (2008) Seasonal variations in microcystin con-centrations in Lake Taihu, China. Environ Monit Assess 145:75–79
Ye W, Liu X, Tan J, Li D, Yang H (2009) Diversity and dynamics ofmicrocystin—producing cyanobacteria in China’s third largest lake,Lake Taihu. Harmful Algae 8:637–644
Environ Sci Pollut Res