alk-1-enyl ether phospholipids (plasmalogens) and glycolipids of treponema hyodysenteriae: analysis...
TRANSCRIPT

273
Biochimica et Biophysics Acta, 618 (1980) 273-281 @ Elsevier/North-Holland Biomedical Press
BBA 57547
ALK-l-ENYL ETHER PHOSPHOLIPIDS (PLASMALOGENS) AND GLYCOLIPIDS OF TREPONEMA HYODYSENTERZAE
ANALYSIS OF ACYL AND ALK-l-ENYL MOIETIES *
HERBERT M. MATTHEWS **, TZE-KEN YANG and HOWARD M. JENKIN ***
The Hormel Institute, University of Minnesota, Austin, MN 55912 (U.S.A.)
(Received June lst, 1979) (Revised manuscript received November 14th, 1979)
Key words: Plasmalogen; Glycolipid Alk-1-enyl ether; (T. hyodysenteriae)
Summary
The lipids of Treponema hyodysenteriae B78, the etiologic agent of swine dysentery, comprised 16.4% of the cell dry weight, and consisted of 37.4% glycolipids, 28.6% phospholipids, and 34.0% neutral lipids. Monogalactosyldi- acylglycerol, a major lipid in all Treponema except Treponema pallidurn, comprised 80% of the glycolipids. An unidentified galactolipid less polar than monogalactosyldiacylglycerol was also detected. Phosphatidylglycerol (19.5% of the total lipids) was the major phospholipid. Phosphatidylcholine, characteristically the major phospholipid of treponemes, comprised 6.1% of the total lipids. Cardiolipin and lysophosphatidylcholine were minor components.
The alk-lenyl ether forms of both the phospholipids (plasmalogens) and glycolipids predominated. The alk-lenyl ether forms of monogalactosyldiacyl- glycerol, the unidentified galactolipid, phosphatidylglycerol, cardiolipin, and phosphatidylcholine were 88.3,96.4, 74.8, 60.6, and 6.3%, respectively.
The acyl and alk-l-enyl chains of the organism were qualitatively similar and differed dramatically from those of the medium, indicating a capability for fatty acid synthesis that most Treponema do not possess. Saturated C14, C15, and Cl6 chains comprised more than 95% of the acyl and alk-lenyl groups.
About 25% of the chains were iso- : 0, anteiso-15 : 0, and other branched moieties.
* A preliminary report of this investigation. Abstract D71, was presented at the 79th Annual Meeting of the American Society for Microbiology, Los Angeles. CA, U.S.A.. May. 1979.
** Present address: Department of Microbiology and Immunology, University of Arkansas for Medical Sciences, Little Rock. AR 72205, U.S.A.
*** To whom reprint requests should be addressed.

274
Introduction
In 1972, Harris et al. [l] isolated a spirochete in pure culture from the intestine of a pig affected with dysentery. A new species, Treponema hyodysenteriae, was proposed for the organism since it morphologically resembled treponemes [ 1,2] and induced swine dysentery when given orally to pigs [1,3]. It is the only pathogenic Treponema which has been serially cultivated in vitro.
Lipids comprise about 20% of the cellular dry weight of Tre~onema [4,5] and, except in Treponema pallidurn [6,7], 25-50s of the lipid consists of a single glycolipid, monoglycosyldiacylglycerol [4,5,8]. The major phospholipid is phosphatidylcholine [ 5,6,8], a characteristic lipid of eucaryotic cells more so than of bacteria [ 91. Most Z’reponema require long-chain fatty acids for growth [5,10] although a few oral [11,12] and intestinal [13] strains require short- chain volatile fatty acids. Nothing is known about the lipid composition or metabolism of T. hyodysenteriae.
Plasmalogens, alk-lenyl ether lipids, are present in mammals, birds, marine invertebrates and vertebrates, and some protozoa [14]. Significant amounts have only recently been reported in fungi in the soil yeast Puliularia [15]. Various anaerobic bacteria contain plasm~ogens [16--271 whereas aerobic and facultatively anaerobic bacteria, except for the marine bacterium MB 45 1281, do not contain these ether lipids [14,20]. Although the lipids of numerous species and strains of Treponema, anaerobic spirochetes, have been investigated [ 4-6,8,29--311, only choline plasmalogen (about 20% of choline phospho- lipid) has been reported in one strain [S] of this genus.
In this report we present data on the lipid composition as well as the acyl and alk-lenyl compositions of the individual lipids of T. hyodysenteriue. Evidence is provided that the organism differs significantly from the other Treponema regarding lipid composition and metabolism.
Materials and Methods
Propagation of organism and lipid extraction. T. hyodysenteriae, type strain B78 [l], was obtained from D.L. Harris, Iowa State University, Ames, IA (U.S.A.) and grown under deoxygenated HZ/CO2 (50 : 50, v/v) in Trypticase soy broth with 10% fetal calf serum [32]. Cells in the early stationary phase were centrifuged at 12 000 X g for 30 min at 4°C and washed three times with 20 mM phospha~-buffered saline (pH 7.3). Lipids were immediately extracted [61.
Lipid fractionation. Lipid classes were separated by preparative thin-layer chromatography using Silica gel H (500 pm thick) on 20 cm X 20 cm glass plates. Total lipids (10 mg/plate) iYere first chromatographed with CHCIJ CH,OH/28% NH,OH (60 : 15 : 2, v/v/v) to isolate two glycolipids that were eluted from the gel with acetone. Total neutral lipids were recovered with CHC13. Total phospholipids were eluded with CH30H/CHC13/Hz0 (2 : 1 : 0.8, v/v/v) Lind recovered in the CHC13 layer after adjusting the ratio to 1 : 1 : 0.9 (v/v/v). Individual phospholipids were then isolated using CHC13/CH30H/H,0 (65 : 25 : 4, v/v/v).

275
Analysis of lipids. Lipids were tentatively identified by comparison of mobil- ities on Silica gel H plates with authentic standards employing CHC13/CH30H/ 28% NHaOH (60 : 35 : 5, 60 : 15 : 2, and 60 : 10 : 1, v/v/v), CHC1JCH30H/ HZ0 (65 : 25 : 4, v/v/v), ‘and CHCl&H,OH/acetic acid/H,0 (25 : 15 : 4 : 2, by vol.). ‘Twodimensional chromatography was performed using boric acid- containing Silica gel G plates with the solvent systems described by Poorthius et al. [ 331. Silica gel H plates developed first with CHC1&H30H/28% NH40H (60 : 30 : 4, v/v/v) and in the second dimension with CHClJCH,OH/H,O (65 : 25 : 4, v/v/v) were also used. Lipids were detected with cu-naphthol, molybdate, ninhydrin, 2,4dinitrophenylhydrazine, H,S04, and dichloro- fluorescein [34]. The latter was removed from lipid fractions as described by Clarke et al. [ 181.
Acid methanolysis was performed at 100°C for 2 h using nonadecanoic acid and 1 ,ldimethoxyheptadecane as internal standards [6]. The fatty acid methyl esters and dimethyl acetals were separated by thin-layer chromatography using benzene and eluted from the silica gel with light petroleum (38-45”C)/diethyl ether (80 : 20, v/v). Catalytic hydrogenation followed by bromination was used to determine the presence of unsaturated and cyclopropane constituents [34]. Methyl esters of non-hydroxy and hydroxy fatty acids were separated using light petroleum (38-45”C)/diethyl ether (3 : 1, v/v). Methyl glycosides were prepared by acid methanolysis at 80°C for 20 h [35] using mannitol or glucose as internal standards, and converted to trimethylsilyl derivatives [ 361.
Dioxanes of the alk-lenyl chains were prepared by magnetically stirring the isolated lipids and 1,ldimethoxyheptadecane as internal standard with 1,3- propanediol in an anhydrous solution of p-toluene-sulfonic acid in benzene for 4 h at 75°C [37]. Dioxanes recovered in the upper phase after addition of 5 ml light petroleum (38-45”C)/diethyl ether (2 : 1, v/v) and 0.5 ml water were purified on Silica gel H by developing twice with light petroleum (38-45’C)/ diethyl ether (95 : 5, v/v) and eluting with diethyl ether.
Part of the total lipids was reduced with 70% Vitride reagent in benzene [ 381 and then chromatographed using diethyl ether/acetic acid (100 : 0.5, v/v) to detect alkyl- and alk-lenylglycerols [39]. To determine the position of attachment of the ether moieties, the alk-lenylglycerols were hydrogenated and the total alkylglycerols obtained were chromatographed on boric acid- impregnated Silica gel G plates using CHCl,/CH,OH (98 : 2, v/v) [40] and l-O- alkyl- and 20alkylglycerol ether standards.
Gas-liquid chromatography was performed with a Fisher-Victoreen 4000 instrument equipped with flame ionization detectors. Trimethylsilyl derivatives of the methyl glycosides were separated at 160°C in a stainless steel column (0.32 cm X 183 cm) packed with 3% SE-30 on 100/120 mesh Gas-Chrom Q (Applied Science Laboratories, Inc., State College, PA, U.S.A.). An aluminum column (0.32 cm X 244 cm) packed with 10% SP-2330 on 100/120 mesh Chromosorb W AW (Supelco, Inc., Bellefonte, PA, U.S.A.) was used at 155°C for methyl esters, 145°C for dimethyl acetals, and 165% for dioxanes. Gas chromatography-mass spectrometry of fatty acid methyl esters and dimethyl acetals was performed with a DuPont DP-102 mass spectrometer system equipped with a 30 m X 0.25 mm inner diameter glass column containing SP- 1000 (Supelco). The temperature was programmed at 6’C/min from 90 to

276
220°C. Tentative identifications and quantitation of isolated peaks were accomplished as previously described [6,41] using reference and internal standards. Quantities of individual lipids and their alk-lenyl forms were estimated from the average chain lengths and amounts of the acyl and alk-l- enyl moieties in relation to the weights of internal standards [34,42].
Lipid quantities were also determined gravimetrically after drying in vacua over CaCl, and P,05 and by phosphorus analysis [43]. Glycerol was determined with chromotropic acid after periodate oxidation of a total 2 N HCl hydroly- sate of the lipid [ 441.
Results
Identification of lipids Lipid components were initially identified by staining reactions and mobili-
ties in several solvent systems compared with standards as described in Materials and Methods. The molybdate reagent gave positive reactions indica- tive of phosphatidylglycerol, phosphatidylcholine, cardiolipin and lysophospha- tidylcholine. Positive reactions indicating the occurrence of monoglycosyldi- acylglycerol and an unidentified glycolipid less polar than monoglycosyldiacyl- glycerol were obtained with cw-naphthol. Sterols and sterol esters appeared as red spots after heating with H,SO+ No ninhydrin-positive spots were detected. The only other detectable lipid was traces of a component observed on some plates that migrated with sphingomyelin. Alk-l-enyl groups were detected in both glycolipids and phospholipids (plasmalogens) after spraying plates with dinitrophenylhydrazine in 2 N HCl.
The individual lipids were isolated and identifications substantiated by quantitative analyses of glycerol, phosphorus, sugar, and acyl plus aIk-l- enyl contents. The lipids identified as phosphatidylglycerol, phosphatidyl- choline, and cardiolipin contained glycerol, phosphorus, and acyl plus alk-l- enyl chains in the molar ratios of 2.00 : 0.97 : 1.81, 1.00 : 1.11 : 1.71, and 3.00 : 1.74 : 3.97, respectively. Analysis of lysophosphatidylcholine was not attempted.
Both glycolipids contained galactose as the only sugar. Negative reactions with ninhydrin and molybdate reagents indicated the absence of free amino groups and phosphate esters in both lipids. Identification of monogalactosyldi- acylglycerol was substantiated by analyses of glycerol, galactose, and acyl plus alk-lenyl chains which were present in the molar ratios of 1.00 : 0.91 : 1.72. The ratios for the unidentified galactolipid were 1.00 : 0.84 : 1.48. The possible presence of sterol in the unidentified galactolipid [45] was not indicated by any pink coloration of the lipid on chromatographic plates heated with H,SO+ Also, no sterol was detected on plates developed with light petroleum, (38-45”C)/diethyl ether (50 : 20, v/v) after acid methanolysis of the lipid . .
The mobility of the unidentified galactolipid was faster than monogalacto- syldiacylglycerol in all solvent systems. Differences in mobilities were greatest in the presence of boric acid which retards lipids containing vicinal hydroxyl groups [33]. On boric acid-containing plates developed in solvent system I of Poorthius et al. [33], the RF values for the unidentified galactolipid and mono-

277
galactosyldiacylglycerol were 0.55 and 0.19, respectively; without boric acid, the RF values were 0.77 and 0.64, respectively.
Quantitative composition of lipids The lipids consisted of 37.4% glycolipids, 28.6% phospholipids, and 34.0%
neutral lipids (Table I). Sterols comprised the bulk of the neutral lipids which were not characterized further. Glycolipids consisted of about 80% mono- galactosyldiacylglycerol and 20% of the unidentified component. Phosphatidyl- glycerol was the principal phospholipid whereas phosphatidylcholine, the major phospholipid of other Treponema [5,6], constituted about one-third of the amount of phosphatidylglycerol. Cardiolipin and lysophosphatidylcholine were present in small amounts.
The presence of the alk-1-enylacyl form of the lipids was confirmed and quantitated by the preparation and analysis of the dimethyl acetals and diox- anes of the alk-lenyl moieties (Table I). Both glycolipids existed almost entirely as the alk-l-enylacyl analogue. The alk-lenylacyl (plasmalogen) form accounted for 74.8% of phosphatidylglycerol and 60.6% of cardiolipin whereas very little phosphatidylcholine and none of its lyso-form existed as plasma- logens.
No evidence for alkylglycerolipids was obtained. Although alk-1-enylglycerols derived from the alk-lenyl ether lipids were detected after Vitride reduction [ 38,391 of the total lipids, no alkylglycerols were found.
Hydrogenation of the aIk-lenylglycerols obtained after Vitride reduction
TABLE I
LIPID COMPOSITION OF T. HYODYSENTERIAE Bl8 AND PROPORTION OF THE LIPIDS IN THE
ALK-I-ENYLACYL FORM
30 mg lipids were extracted from the spirochete. fractionated, and quantitated gravimetrically and by analyses of phosphorus, carbohydrate. and acyl plus alk-1-enyl contents as described in Materials and Methods. Lipids amounted to 15.4 mg/l of culture and 16.4% of the organism’s dry weight. All values are the mean of at least two determinations from duplicate samples. Mel% values were calculated from the molar ratio of the alk-1-enyl chains, analyzed as dimethyl acetals. to the acyl groups, analyzed as methyl esters [421. Values estimated from the ratio of the dioxanes of the alk-1-enyl chains to the fatty acid methyl esters were 3.6-12.1% higher. n.d., not detected.
Lipid Relative % by weight mol% in alk-l-enylacyl form
Total lipids
Polar lipids
Phospholipids 28.6
Phosphatidylglycerol 19.5 Phosphatidylcholine 6.1 Cardiollpin 2.0 Lysophosphatidylcholiine 1.0
Glycolipids 37.4
Monogalactosyldiacylglycerol 30.0 Unidentified galactolipid 7.4
Neutral lipids 34.0
43.3 -
29.6 74.8 9.2 6.3 3.0 60.6 * 1.5 n.d.
56.7 -
45.5 88.3 11.2 96.4
- -
* The content of cardiolipin in the alk-l-enylacyl form was calculated on the assumption that both ends of the molecule existed in the plasmalogen form. A moIecular species possibly exists, however, which contains an alk-1-enyl chain in only one end of the molecule.

278
resulted in alkylglycerols that cochromatographed with 1-0-alkylglycerol on boric acid-containing plates [ 401. This indicates that the aIk-l-enyl moieties in the ether lipids are attached at the C-l position of glycerol.
Composition of fatty acyl moieties The fatty acids of the total lipids and the major polar lipids are shown in
Table II. The growth medium contained 10% fetal calf serum which accounted for all of the medium fatty acids: 14 : 0 (1.3%), 15 : 0 (0.8%), 15 : 1 (0.4%), 16 : 0 (24.0%), 16 : 1 (5.7%), 17 : 0 (l.O%), 17 : 1 (1.4%), 18 : 0 (10.3%), 18 : 1 (25.2%), 18 : 2 (6.1%), and 22.7% of C&, and larger fatty acids. The fatty acid content of T. hyodysenteriae and the culture medium were drasti- cahy different. Saturated fatty acids comprised 99% of those of the total lipids of the organism and only 38% of those of the medium. Major fatty acids of the organism had 14, 15, or 16 carbon chains. Significant amounts of 18 : 0 and 18 : 1 fatty acids were found in phosphatidylcholine, cardiolipin, and lysophos- phatidylcholine. Branchedchain fatty acids accounted for about 25% of the acyl contituents of the total lipids and consisted mainly of iso- and anteiso- 15 : 0 acids. Minor amounts of iso- : 0, iso- : 0 and anteiso-17 : 0 acids were found. No hydroxy or cyclopropane fatty acids were detected.
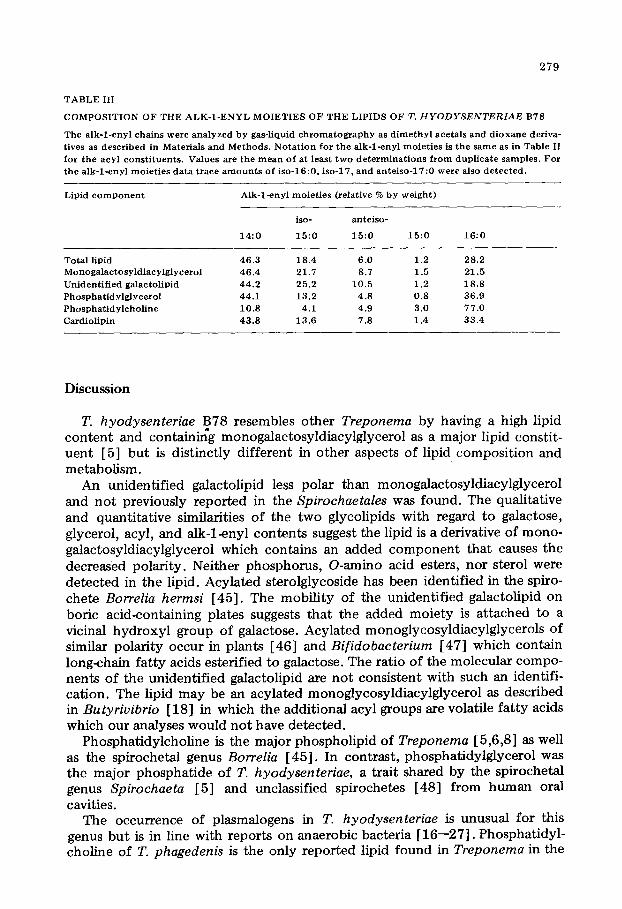
Composition of alk-l-enyl moieties The composition of the alk-lenyl chains (Table III) was qualitatively similar
to the acyl moieties although 17 : 0,18 : 0, and 18 : 1 aIk-1-enyl moieties were not present in detectable amounts. The iso-, anteiso-, and n-15 : 0 aIk-1-enyl moieties had about the same distribution as the acyl moieties. The greatest difference in the distributions of the aIk-l-enyl and acyl chains was that 14 : 0 was the principal aIk-lenyl moiety whereas 16 : 0 predominated among the acyl groups.
TABLE II
COMPOSITION OF THE ACYL MOIETIES OF THE LIPIDS OF T. HYODYSENTERIAE B78
Methyl esters of the fatty acids were prepared, separated from the dimethyl acetals, and analyzed by gas- liquid chromatography as described in Materials and Methods. The fatty acids are designated by the num- ber of carbon atoms followed by the number of double bonds with the prefixes anteiso or iso to indicate the type of branching. Values are the mean of at least two determinations from duplicate samples. For the acyl moieties data, trace amounts of iso-16:O. iso-17:O and anteiso-17:O were also detected.
Lipid component Acyl moieties (relative % by weight)
iso- anteiso-
14:o 15:o 15:o 15:o 16:0 17:o 18:0 18:l
Total lipid Monogalactocyldiacylglycerol Unidentified galactolipid Phosphatidylglycerol Phosphatidylcholine Cardiolipin Lysophosphatidylcholine
10.8 18.5 5.9 0.8 58.9 0.9 2.6 0.8 9.6 23.0 9.8 0.9 54.9 1.2 7.7 27.4 10.1 0.8 51.9 1.2 -
12.4 19.3 7.2 0.8 58.2 0.9 -
2.6 6.6 3.1 0.8 69.0 1.5 11.5 3.5 7.8 14.3 6.2 0.7 47.8 1.1 17.0 3.8
1.3 1.1 0.6 0.4 27.0 1.7 51.5 14.7
279
TABLE III
COMPOSITION OF THE ALK-l-ENYL MOIETIES OF THE LIPIDS OF T. HYODYSENTERIAE B78
The alk-1-enyl chains were analyzed by gas-liquid chromatography as dimethyl acetals and dioxane deriva- tives as described in Materials and Methods. Notation for the alk-l-enyl moieties is the same as in Table II
for the acyl constituents. Values are the mean of at least two determinations from duplicate samples. For the alk-l-enyl moieties data trace amounts of iso-16:O. iso-17, and anteiso-17:O were also detected.
Lipid component Alk-lenyl moieties (relative % by weight)
14:o
iso-
15:o
anteiso-
15:o 15:o 16:0
Total lipid 46.3 18.4 6.0 1.2 28.2 Monogalactosyldiacylglycerol 46.4 21.7 8.7 1.5 21.5 Unidentified galactolipid 44.2 25.2 10.5 1.2 18.8 Phosphatidylglycerol 44.1 13.2 4.8 0.8 36.9 Phosphatidylcholine 10.8 4.1 4.9 3.0 77.0 Cardiolipin 43.8 13.6 7.8 1.4 33.4
Discussion
T. hyodysenteriae F78 resembles other Treponema by having a high lipid content and containing monogalactosyldiacylglycerol as a major lipid con&it- uent [5] but is distinctly different in other aspects of lipid composition and metabolism.
An unidentified galactolipid less polar than monogalactosyldiacylglycerol and not previously reported in the Spirochaetules was found. The qualitative and quantitative similarities of the two glycolipids with regard to galactose, glycerol, acyl, and alk-lenyl contents suggest the lipid is a derivative of mono- galactosyldiacylglycerol which contains an added component that causes the decreased polarity. Neither phosphorus, O-amino acid esters, nor sterol were detected in the lipid. Acylated sterolglycoside has been identified in the spiro- chete Borreliu hermsi [45]. The mobility of the unidentified galactolipid on boric acid-containing plates suggests that the added moiety is attached to a vicinal hydroxyl group of galactose. Acylated monoglycosyldiacylglycerols of similar polarity occur in plants [46] and Bifidobacterium [47] which contain longchain fatty acids esterified to galactose. The ratio of the molecular compo- nents of the unidentified galactolipid are not consistent with such an identifi- cation. The lipid may be an acylated monoglycosyldiacylglycerol as described in Bu tyriuibrio [ 18 ] in which the additional acyl groups are volatile fatty acids which our analyses would not have detected.
Phosphatidylcholine is the major phospholipid of Treponema [ 5,6,8] as well as the spirochetal genus Borreliu [45]. In contrast, phosphatidylglycerol was the major phosphatide of T. hyodysenteriae, a trait shared by the spirochetal genus Spirochaeta [ 51 and unclassified spirochetes [48] from human oral cavities.
The occurrence of plasmalogens in T. hyodysenteriae is unusual for this genus but is in line with reports on anaerobic bacteria [ 16-271. Phosphatidyl- choline of T. phagedenis is the only reported lipid found in Treponema in the

plasmalogen form [8]. Interestingly, this lipid of T. hyodysenteriae existed as only 6.3% plasmalogen although other polar lipids were mainly of the plasma- logen type. The plasmalogen form of phosphatidylglycerol, the major phospha- tide of T. hyodysenteriue (74.8% plasmalogen), has previously been described in Clostridium (9%) [17], Sphaerophorus (7.8%) [19], Anaeroplusmu (58.3%) [23], and Butyrivibrio (about 100%) [18]. Cardiolipin plasmalogen, reported in minor amounts (5.2%) in Sphuerophorus [19], amounted to 60.6% in T. hyodysen teriae.
The finding that the glycolipids of T. hyodysenteriue also existed in the plasmalogen-like form is unusual but not unique. Glycolipids of Butyrivibrio also exist in the ‘plasmalogen form’ [18].
The fact that the acyl and alk-lenyl moieties of T. hyodysenteriue were qualitatively similar but differed quantitatively is in agreement with other studies [ 15,23,26,49]. The presence of anteiso-isomers is interesting since these have previously been identified in spirochetes only in Spirochuetu [ 51. Other intestinal spirochetes from swine contain only the normal and iso-isomers [48].
The fatty acids of most treponemes are similar to those of the medium since the organisms require longchain fatty acids for growth [5,10] due to the inability to synthesize, P-oxidize, and desaturate them [4,8]. In contrast, the fact that the fatty acids of T. hyodysenteriae were drastically unlike those of the medium indicates the organism is able to synthesize long-chain fatty acids. Growth in serum-containing medium without added short-chain fatty acids suggests the latter are not required precursors for synthesis of long-chain fatty acids as appears to be the case for T. microdentium [11,12], Treponemu strain PR-7 [ 131, and certain unclassified spirochetes [48].
Miao et al. [50] has suggested that differences in the DNA of T. hyody- senteriue and other Treponema argue for a separate genus for this spirochete. Our studies have shown that the lipid composition and metabolism of T. hyodysenteriae are also significantly different from other Treponema. The taxonomy of this intestinal pathogen appears therefore to be in a similar state of uncertainty as that of other intestinal and oral spirochetes [48].
Acknowledgements
This investigation was supported in part by Public Health Service contract NO1 AI 42537 from the National Institute of Allergy and Infectious Diseases; grant HL 08214 from the Program Projects Branch, Extramural Programs, National Heart, Lung and Blood Institute, and by The Hormel Foundation. H.M.M. was the recipient of Public Health Service Research Fellowship Award 5 F32 AI05354 from the National Institute of Allergy and Infectious Diseases. We are grateful to D.L. Harris and J.M. Kinyon for the organism, P.C. Schmid for advice on plasmalogen analyses, Wayne Heimermann for mass spectrometric analyses, and H.H.O. Schmid and W.J. Baumann for lipid standards.
References
1 Harris, D.L., Glock, R.D.. Christensen, C.R. and Kinyon. J.M. (19’72) Vet. Med. Small Anim. Clin. 67,
61-64

281
2 Glock. R.D. and Harris, D.L. (1972) Vet. Med. SmaII Anim. Clin. 67, 65-68 3 Kinyon. J.M., Harris, D.L. and Glock. R.D. (1977) Infect. Immun. 15, 638-646 4 Johnson, R.C., Livermore, BP., Jenkin, H.M. and Eggebraten. L. (1970) Infect. Immun. 2, 6066609
5 Livermore, B.P. and Johnson, R.C. (1974) J. Bacterial. 120. 1268-1273 6 Matthews, H.M.. Yang, T-K. and Jenkin, H.M. (1979) Infect. Immun. 24, 713-719 7 Dupouey, P. and Betz. A. (1969) Ann. Inst. Pasteur (Paris) 117. 395407 8 Meyer, H. and Meyer, F. (1971) Biochim. Biophys. Acta 231. 93-106 9 Goldfine, H. (1972) Adv. Microb. Phys. 8. l-58
10 Johnson, R.C. and Eggebraten. L.M. (1971) Infect. Immun. 3, 723-726 11 Socransky, S.S., Loesche, W.J., Hubersak, C. and MacDonald, J.B. (1964) J. Bacterial. 88. 200-209
12 Hardy, P.H., Jr. and Munro. C.O. (1966) J. Bacterial. 91. 27-32 13 Smibert. R.M. and Claterbaugh. R.L., Jr. (1972) Can. J. Microbial. 18, 1073-1078 14 Snyder, F. (ed.) (1972) in Ether Lipids, Chemistry and Biology, pp. 177-350, Academic Press, New
York
15 Goni. F.M., Dominquez, J.B. and Uruburu, F. (1978) Chem. Phys. Lipids 22. 79-81 16 Allison. M.J.. Bryant, M.P., Katz. I. and Keeney, M. (1962) J. Bacterial. 83, 1084-1093 17 Baumann, N.A.. Hagen, P-O. and Goldfine, H. (1965) J. Biol.. Chem. 240.1559-1567 18 Clarke, N.G., Ha&wood, G.P. and Dawson, R.M.C. (1976) Chem. Phys. Lipids 17. 222-232 19 Hagen, P.-O. (1974) J. Bacterial. 119, 643445 20 Kamio, Y., Kanegasaki. S. and Takahashi, H. (1969) J. Gen. Appl. Microbial. 15. 439451 21 Kanegasaki, S. and Takahashi. H. (1968) Biochim. Biophys. Acta 152, 40-49
22 Katz, I. and Keeney, M. (1964) Biochim. Biophys. Acta 84. 128-132 23 Langworthy, T.A., Mayberry, W.R.. Smith, P.F. and Robinson, I.M. (1975) J. Bacterial. 122, 785-
787 24 Van Golde. L.M.G., Akkermans-Kruyswijk, J.. Franklin-Klein, W., Lankhorst, A. and Prim.. R.A.
(1975) FEBS Lett. 53, 57-60 25 Van Golde, L.M.G.. Prins, R.A., Franklin-Klein, W. and Akkermans-Kruyswijk, J. (1973) Biochim.
Biophys. Acta 326. 314-323 26 Verkiey, A.J.. Ververgaert, P.H.J.Th., Prins, R.A. and Van Golde. L.M.G. (1975) J. Bacterial. 124,
1522-1528
27 Wegner, G.H. and Foster, E.M. (1963) J. Bacterial. 85. 53-61 28 McAllister, D.J. and De Siervo, A.J. (1975) J. Bacterial. 123, 302-307 29 Coulon-Morelec. M.-J., Dupouey, M.P. and Marechal, J. (1969) C.R. Acad. Sci. Paris 269. 854-857 30 Livermore, B.P. and Johnson, R.C. (1970) Biochim. Biophys. Acta 210, 315-318
31 Vaczi. L., Kiraly. K. and Rethy, A. (1966) Acta Microbial. Acad. Sci. Hung. 13. 79-84 32 Kinyon, J.M. and Harris, D.L. (1974) Vet. Rec. 95, 219-220 33 Poorthius, J.H.M.. Yazaki, P.J. and Hostetler, K.Y. (1976) J. Lipid Res. 17, 433-437 34 Kates. M. (1972) Techniques of Lipidology; Isolation, Analysis and Identification of Lipids, American
Eisevier Publishing Co., Inc., New York 35 Vance, D.E. and Sweeley, CC. (1967) J. Lipid Res. 8, 621-630 36 Desnick, R.J., Sweeley, CC. and Krivit, W. (1970) J. Lipid Res. 11, 31-37 37 Venkata Rae. P., Ramachandran, S. and Cornwell, D.G. (1967) J. Lipid Res. 8, 380-390 38 Snyder. F., Blank. M.L. and WykIe, R.L. (1971) J. Biol. Chem. 246. 3639.-3645 39 Snyder, F. (1976) in Lipid Chromatographic Analysis (Marinetti, G.V., ed.), 2nd edn.. Vol. 1, Marcel
Dekker Inc., New York 40 Wood, R. and Snyder. F. (1967) Lipids 2, 161.-171 41 Farquhar, J.W. (1962) J. Lipid Res. 3, 21-30 42 Viswanathan. C.V., Basilio. M.. Hoevet, S.P. and Lundberg, W.O. (1968) J. Chromatogr. 34. 241-245 43 Rouser, G.. Fleicher, S. and Yamamoto, A. (1970) Lipids 5, 494. -496
44 Dittmer, J.C. and Wells. M.A. (1969) Methods Enzymol. 14.482-530 45 Livermore, B.P., Bey, R.F. and Johnson. R.C. (1978) Infect. Immun. 20, 215-220 46 Clayton, T.A.. MaeMurray, T.A. and Morrison. W.R. (1970) J. Chromatogr. 47. 277-281
47 Veerkamp, J.H. (1972) Biochim. Biophys. Acta 273,359-367 48 Livermore. B. and Johnson, R.C. (1975) Can. J. Microbial. 21, 1877-1880 49 Goldfine, H. and Panos, C. (1971) J. Lipid Res. 12. 214-220 50 Miao, R.M., Fieldsteel, A.H. and Harris, D.L. (1978) Infect. Immun. 22, 736-739