advances in microbial
TRANSCRIPT

ELSEVIER
Review Advances in microbial steroid biotransformation Shashi B. Mahato and Subhadra Garai
Indian Institute of Chemical Biology, Calcutta, India
Microbial biotransformations of various steroids are reviewed. Developmental studies on hydroxylation, carbon- carbon bond cleavage, enzymatic catalysis in nonaqueous solvents, use of cyclodextrin medium, cell immobili- zation, and new microbial reactions are highlighted. Various steroid substrates, their metabolites and the microorganisms used for the transformations are compiled covering the literature for the period 1992-1995. (Steroids 62:332-345, 1997) © 1997 by Elsevier Science Inc.
Keywords: microbial biotransformation; review; substrate; metabolite; compilation
Introduction
The importance of microbial biotechnology in the produc- tion of steroid drugs and hormones was realized for the first time in 1952 when Murray and Peterson of Upjohn Com- pany patented the process of 1 la-hydroxylation of proges- terone by a Rhizopus speciesJ Since then, microbial reac- tions for the transformation of steroids have proliferated, and specific microbial transformation steps have been in- corporated into numerous partial syntheses of new steroids for evaluation as drugs and hormones. A variety of steroids are widely used as anti-inflammatory, diuretic, anabolic, contraceptive, antiandrogenic, progestational, and antican- cer agents as well as in other applications. Although intro- ductions of new steroids into commerce is now limited, our interest lies in improvement in the yields of desired metabo- lites as well as preparation of novel steroids that are difficult to synthesize by chemical means. The areas of microbial biotechnology that are now receiving attention are: applica- tion of the newer concepts of genetic engineering of micro- organisms for their improvement as steroid-transforming agents; solubility improvement for carrying out biotransfor- mation of substrates that are sparingly soluble in water; immobilization of enzymes or whole cells in a suitable ma- trix for repetitive economic utilization of enzymes; devel- opment of a continuous process for economic product re- covery; and manipulation of culture media for improvement in product yields by use of cyclodextrin. Our previous re-
25 v i e w s - covered literature of the period 1979-mid-1992.
Address reprint requests to Dr. Shashi B. Mahato, Indian Institute of Chemical Biology, 4 Raja S.C. Mullick Road, Jadavpur, Calcutta-700032, India. Received August 19, 1996; accepted November 14, 1996.
Steroids 62:332-345, 1997 © 1997 by Elsevier Science Inc. 655 Avenue of the Americas, New York, NY 10010
This review attempts to present the situation during the period from late 1992 to 1995.
Hydroxylation
The lloL-, 1113-, and 16c~-hydroxylations are now exclu- sively achieved in the steroid industry by microbial trans- formations operating at high yield and controlled costs. However, developmental activities for further cost reduc- tion in these transformations are sometimes reported. The other hydroxylations that seem to have potential for indus- trial exploitation are 9o~- and 14ot-hydroxylations. Russian workers 6 studied the 1113-hydroxylation of cortexolone by the mycellium of Curvularia lunata VKMF-644 in the pres- ence of 13-cyclodextrin (13-CD). The ratios of cortexolone to [3-CD varying from 1.0:0.1 to 1.0:1.0 were used. The yield of hydrocortisone by cortexolone biotransformation at a concentration of 4 g/L (the ratio of cortexolone to [3-CD was 1.0:0.6) was 70-75%. When 13-CD was not used, the yield was 40-45%. No stimulation of the process was observed when the mycell ium immobilized in Ca-alginate gel was used as a biocatalyst. Stabilization of steroid 1 l- hydroxylation activity of Cunninghamella elegans l~roto- plasts in organic osmotic stabilizers has been reported.' Pro- toplasts of C. elegans showing 1 lo~-, and 1113-hydroxylating ability of substance S (cortexolone) preserved high trans- formation activity when dispersed in glucose-enriched or- ganic osmotic stabilizers. The observation is interesting and requires further investigation.
14o~-hydroxyandrost-4-ene-3,17-dione is a useful inter- mediate for the preparation of hormones. A patent has been taken out 8 for the manufacture of this product by fermen- tation of androst-4-ene-3,17-dione (AD) with a strain of Myrothecium striatisporum. Another patent was taken 9 for
0039-128X]97/$17.00 PII S0039-128X(96)00251-6

manufacture of the same product by fermentation of the same substrate but using a number of different microbial strains. 613,14ot-Dihydroxyandrost-4-ene-3,17-dione and 14ct-hydroxyandrost-4-ene-3,6,17-trione possess androgen activity and are useful inhibitors for breast cancer cells. The process for production of the diketone as well as the trik- etone has been patented. 1° The process involves culturing a microorganism belonging to the genus Myrothecium in a medium supplemented with AD and isolating the diketone from the culture medium. The diketone is then converted to the triketone in a simple manner. Another process for the preparation of the triketone has also been patented. 1~ The process comprises fermentation of AD with Acremonium strictum NN 106 to yield 14o~-hydroxy-AD. Alkoxylation of ketone group at the 3 position of the latter led to the for- mation of 3-alkoxy-14a-hydroxyandrosta-3,5-dien-17-one, which was subsequently oxidized by oxidizing agents to the desired product.
9oL-Hydroxyandrost-4-ene-3,17-dione is a useful inter- mediate that is easily dehydrated into the 9,11-dehydro system. Such 9-halogenated corticoids as well as 11-keto structures can be produced without the traditional l l - hydroxylations. Microbial processes are now available for production of this intermediate. A process for manufacture of the compound by fermentation of a sterol mixture con- taining [3-sitosterol, campesterol, and stigmasterol with Mycobacterium fortuitum has been patented. ~2 Thus, the microorganism was cultured aerobically in 2.5 L medium containing 22.5 g sterol mixture and polyethylene polypro- pylene glycol for 72 h. The product (yield 31.1%) was iso- lated from the culture medium using adsorbent at the end of the fermentation. Another patent of similar nature has been taken for the production of the compound. ~3 According to this process, Mycobacterium fortuitum was cultured for 72 h in a medium containing sitosterol and an ethylvinyl ben- zene-divinylbenzene resin coated with the polyethylene- polypropylene oxide adduct of N, N"-tetra (oxypropylenox- ide) N'-(2-oxo-propyl) diethylene-triamine. The isolation and purification of the compound from the fermentation broth was facilitated by including a surfactant-coated adsor- bent in the medium.
Microb ia l t r ans fo rma t ion of 3[3-hydroxy-5c~-H- pregnanes to their 9et-hydroxy-4-ene-3-keto derivatives has also been reported. 14 Biotransformation of 5et-H-steroids at 0.5-3.0 g/L by a wild Rhodococcus species strain isolated from the soil resulted in the formation of the corresponding 4-ene-3-keto and 9et-hydroxy-4-ene-3-keto analogs. The conversion of 5(x-H-steroids into 9et-hydroxy derivatives as final products depended on the functionality of the D-ring. Transformation of sitosterol to 9ot-hydroxyandrost-4-ene- 3,17-dione by Mycobacterium sp. 207 cells in the presence of an adsorption resin has been published. ~5 A bioreactor containing an adsorption resin and with a two-stage axial- flow turbine as a stirrer was used. The preparative yield of the product was 51%, and the adsorbent could be used re- peatedly.
Carbon-Carbon bond cleavage
Although stigmasterol has long been used as a starting ma- terial for its chemical modification by oxidative cleavage of
Biotransformation of steroids: Mahato and Garai
the 22-double bond to suitable starting structures, sitosterol, campesterol, and cholesterol were considered waste prod- ucts because of the lack of economic routes for the cleavage of their saturated side chain. Development of the selective microbial degradation of the sterol side chain helped the utilization of these cheap and readily available natural prod- ucts. However, continued attempts are being made for fur- ther cost reduction in the process. A process for conversion of sterols to androsta-1,4-diene-3,17-dione (ADD) has been patented.16 The process involves two-phase culture of Mv- cobacterium vaccae ZIMET 11094. Strains and culture con- dition were carefully standardized to ensure the prevention of inhibition of the sterol side chain degradation by ADD. An adsorbent resin was used in the final culture medium, and an aqueous suspension of sterol (10 g/L) was added. The product ADD was recovered from the adsorbent by elution with MeOH.
Liu reported the transformation of cholesterol to testos- terone by a Mycobacterium sp. 17 A supplement of 4% glu- cose and 1% peptone into a synthetic medium, pH control of 6.0, and continuous feeding of cholesterol were the most important parameters. Under optimal conditions, the maxi- mum molar conversion rate of testosterone from cholesterol was upto 42.68% in a 2.5 L surface-aerated fermentor after 100 h cultivation. Testosterone was isolated from the fer- mentation broth by the addition of XAD-7 resin. Testoster- one production by Mycobacterium sp. NRRLB-3683 has been discussed by Hung et al. TM It has been shown that testosterone is not formed by single reduction of the 17-keto group of AD but by a double reduction of both the 17-keto group and the 1-2 double bond of ADD.
Bioconversion of sitosterol to AD by mutants of Myco- bacterium vaccae has been reported by Spassov et al. ~9 These mutants were resistant to rifampicin and streptomy- cin. Production of ADD from cholesterol using two-step microbial transformation has been reported. 2° Cholesterol (20 g/L) was initially converted to cholestenone by an in- ducible cholesterol oxidase-producing bacterium, Arthro- bacter simplex U-S-A-18. The maximum productivity of cholestenone was 8 g/L-day, and the molar conversion rate was 80%. Subsequently, a fine suspension of cholestenone (50 g/L), which was prepared directly from the fermentation broth of A. simplex to ADD by Mycobacterium sp. NRRLB- 3683 in the presence of an androstenone adsorbent, Amber- lite XAD-7. The maximum productivity of ADD was 0.91 g/L-day, and the molar conversion rate was 35%.
Mixed culture
Developmental studies are continuing for conducting two or more microbial steps in one fermentation using mixed cul- tures. In mixed culture fermentation, it is essential that the two microorganisms are capable of inducing the desired enzymes in one another's presence. Transformation of cor- texolone into prednisolone by mixed culture has been re- ported. 2~ Curvularia lunata was the most active organism for 11 [3-hydroxylation of cortexolone to cortisol; whereas, Mycobacterium smegmatis was the most potent bacterium converting the latter to prednisolone. A mixed culture of C. lunata grown for 48 h and allowed to transform cortexolone for another 24 h and M. smegmatis was a combination that
Steroids, 1997, vol. 62, April 333

Review
gave 51% yield of prednisolone. The yield of the product was markedly affected by the composition of the fermenta- tion medium. Some nutritional requirements for the mixed culture have been described. 22 Transformation of cortexo- lone into prednisolone in a single-step fermentation using immobilized mixed culture, entrapped in different gels or adsorbed on clay particles has also been reportedY Calcium alginate at a gel concentration of 2% gave the highest trans- formation activities and prednisolone yields. The entrapped mixed cultures could be repeatedly used in batch-wise trans- formation for at least six times when suspended in diluted nutrient medium and for three times when suspended in distilled water. In the latter case, the entrapped cultures had to be reactivated in nutrient medium for future use. Mixed cultures adsorbed on clay particles were successfully reused 18 times with reactivation after the ninth and fifteenth uses. Continuous transformation of cortexolone into prednisolone by mixed cultures adsorbed on clay particles was more ef- ficient than batch-wise reused adsorbed cultures.
Enzymatic catalysis in nonaqueous solvents
The application of nonaqueous environment for enzymatic catalysis has received much attention in recentyears. Ex-
24 2 J cellent reviews are available on this aspect. ' Enzymes acquire remarkable novel properties such as enhanced sta- bility, altered substrate specificity, and ability to catalyze new reactions. 26 However, progress on the application of these promising features is restricted so far, because our fundamental understanding of this area is very limited. Zaks and Klibanov 27 studied the dependence of catalytic activity of several unrelated enzymes in a number of organic sol- vents as a function of the water content. From their experi- ment on replacement of water with other hydrogen bond- forming additives, and titration of enzyme amino acids in an organic medium, as well as the literature on dehydrated enzymes, they concluded that the water required by en- zymes in nonaqueous solvents provides them with sufficient conformational flexibility needed for catalysis. The results of a study on the control of enzyme enantioselectivity by the reaction medium by Sakurai et al. 28 opined that predictable and rational control of enantioselectivity of enzymes by changing the reaction medium rather than the enzyme itself is possible.
The effect of cell immobilization and organic solvents on sulfoxidation and steroid hydroxylation by Mortierella isa- bellina ATCC 42613 was investigated by Holland et al. 29 They observed that the 14a-hydroxylation of progesterone and the sulfoxidation of thioanisole proceeded in high yield using resting-cell bioconversions but were not carried out by alginate bead preparations in the absence of an organic solvent. The best results were obtained with 5 or 10% aque- ous methanol, and the stereoselectivity of sulfoxidation of thioanisole was found to be dependent upon the nature and concentration of organic co-solvent. A review on the recent development of microbial sterol biotransformation sys- tems 3° particularly emphasizes the new enzymic approach of investigating the cleavage of sterol side chains, new de- velopments in the area of immobilized cell systems, and use of organic media. Sterol side-chain cleavage with immobi- lized Mycobacterium cells in water-immiscible organic sol-
vents was investigated. 31 The highest degradation activities were obtained with cells immobilized in celite with bis (2- ethylhexyl) phthalate as the conversion medium.
A review has been published 32 that deals with biocataly- sis with enzymes and microorganisms immobilized by en- trapment in lyotropic liquid crystals and substrates dis- solved in a co-existing solvent phase from which products may be isolated. Examples are given for the continuous enzymic production of (R)-cyanohydrins and for the micro- bial degradation of steroids.
A new method for the microbial transformation on in- terface between hydrophilic carriers and hydrophobic or- ganic solvents has been proposed. 33 Microorganisms were placed on the interface between such hydrophilic carriers as agar and hydrophobic organic solvents. These microorgan- isms could catalyze various transformations of such syn- thetic substrates as hydrolysis, esterification, oxidation, and reduction more efficiently than emulsion systems. This new type of bioreactor was tentatively named an "interface bio- reactor.' '
Cyclodextrin medium
Cyclodextrins (CD) form inclusion compounds that are bio- compatible with microbes. This property prompted their use in microbial transformation of water-insoluble organic com- pounds. An intensive and systematic investigation of the oxidation of cholesterol to cholest-4-en-3-one by Rhodococ- cus erythropolis was undertaken in the presence of natural and chemically modified CDs in a stirred bioreactor. 34 The biotransformation was strongly affected by the mode of ad- dition of the natural CDs. Although simultaneous addition of cholesterol with either 13- or -/-CD led to a limited en- hancement effect, the microbial oxidation of [3- or ~-CD complexes of cholesterol was totally inhibited. In contrast, the alkylated CDs, dimethyl-, trimethyl-, and hydroxy-, pro- pyl-13-CD exhibited a remarkable enhancement of the mi- crobial oxidation, irrespective of their mode of addition.
A comparative study was conducted by the same au- thors 35 on two biocatalysts, resting R. erythropolis cells and soluble cholesterol oxidase, both catalyzing cholesterol oxi- dation in cyclodextrin medium. The enzyme-mediated ste- rol oxidation was clearly enhanced by the dimethylated 13-CD, as in the microbial oxidation. However, the micro- bial transformation was subject to a larger enhancement effect than the enzymic one with respect to corresponding transformation without CD. The larger dimethylated 13-CD- induced effect exerted on the microbial system was attrib- uted to the stronger affinity of dimethylated [3-CD to the microbial cell. This CD--cell interaction was thought to be manifested through a slightly inhibited microbial growth and a limited leakage of cellular proteins and cholesterol oxidase, Alekhina et al. 36 studied microbiological transfor- mation of steroid-13-CD inclusion compounds. They ob- served that [3-CD at 7-8 g/L promoted 1,2-dehydrogenation of hydrocortisone to prednisolone and of 6a-methyl hydro- cortisone to 6c~-methylprednisolone by Corynbacterium simplex.
A review on the use of cyclodextrins in bioconversions in plant cell biotechnology has appeared. 37 The application of cyclodextrins as precursor solubilizers in biotechnology
334 Steroids, 1997, vol. 62, April

processes, in which plant cells are used, is new. The paper describes the use of CD in the bioconversions by freely suspended and/or immobilized plant cells or plant enzymes. After complexation with [3-CD, the phenolic steroid, 1713- estradiol could be ortho-hydroxylated into a catechol, mainly 4-hydroxyestradiol by a phenoloxidase from grown cells of Mucuna pruriens. The glucosylation of podophyl- lotoxin by cell cultures from Linumflavum was also inves- tigated. Four cyclodextrins, [3-CD, ~,-CD, hydroxypropyl- 13-CD, and dimethyl-[3-CD were used to improve solubility of podophyllotoxin. Dimethyl-13-CD met the needs the best, and the solubility of podophyllotoxin could be enhanced. Podophyllotoxin-13-D-glucoside was formed by the L. fla- vam cells growing in the presence of podophyllotoxin, com- plexed with dimethyl-[3-CD.
Acetone-dried vegetative cells
Cells from microorganisms dried with acetone are useful for special preparations, although this process does not compete with vegetative cell fermentations for manufacture. Al- though fermentation of AD with acetone-dried cells of a fungal strain, Aspergillus fumigatus yielded ADD as the major product and testosterone and its A ~ anolog as minor ones, usual fermentation of AD with the strain furnished a new dihydroxy derivative, 7[3, l la-dihydroxy AD (1) and another steroid 6[3, 1 l~x-dihydroxy AD (2) along with minor metabolites, ADD, A ~ testosterone, and testosterone. 38 Ap- parently in acetone-dried cells, the hydroxylase enzymes are inactivated, but the dehydrogenase and reductase enzymes remains active. Selective 1-dehydrogenation of progester- one by a strain of Aspergillus fumigatus was achieved by fermentation of progesterone with acetone-dried cells of the fungus in the presence of [3-cyclodextrin. 39 Usual fermen- tation of progesterone with the strain yielded four isolable metaboli tes: 3[3-hydroxy-5ot-pregnan-20-one; 1513- hydroxypregna- 1,4-diene-3,20-dione; 713,1513-dihydroxy- pregn-4-ene-3, 20-dione; and 11 et,1513-dihydroxypregn-4- ene-3,20-dione. However, fermentation of the substrate with acetone-dried cells furnished pregna-l,4-diene-3, 20- dione and AD. On the other hand, fermentation with ac- etone-dried cells in presence of [3-cyclodextrin afforded pregna-1,4-diene-3,20-dione as the only isolable metabolite. The results demonstrated that in acetone-dried cells, the hydroxylase and A4-reductase enzymes are inactivated. The activity of the enzyme system responsible for the degrada- tion of the side chain of progesterone is inhibited in the presence of 13-cyclodextrin. Inhibition of certain enzymes in presence of a cyclodextrin is a phenomenon of consider- able interest.
New cell-immobilization techniques
A novel cell-immobilization technique was developed by Houng e t a l . 4° that involves increase of substrate partition to the gel matrix by coating a polyurea thin layer on the sur- face of the calcium alginate beads. The bioconversion of progesterone to 1 le~-hydroxyprogesterone with these poly- urea-coating alignate-entrapped Aspergillus ochraceous cells was investigated using different organic solvents in biphasic media. The reaction medium of ethyl acetate could
Biotransformation of steroids: Mahato and Garai
markedly enhance the bioconversion rate with the existence of a hydrophobic layer, most likely resulting from the in- creasing portion of substrate to gel matrix. Bioconversion with higher substrate concentration was possible using ethyl acetate water medium. The conversion rate increased almost linearly with increasing substrate concentration from 10 to 80 g/L.
The rate with 80 g/L progesterone increased up to six times greater than the rate with the immobilized cells with- out coating. The method could be performed under mild conditions. It has potential for industrial application.
New microbial reactions and novel metabolites
Although introduction of new steroids into commerce is now limited, there are continuing reports on new microbial reactions as well as the production of new steroids by mi- crobial transformation that are otherwise difficult to synthe- size. Murohisa and lida 41 reported production of four new key intermediates in the microbial degradation of choles- terol and campesterol sitosterol side chains. Exposure of 19-hydroxycholesterol and 19-hydroxycampesterol to Rho- dococcus mutant K-3 afforded four new steroid carboxylic acids (3-6) in addition to estrone. The formation of these products helped to propose a degradation pathway for 19- hydroxysterols. Two novel microbial conversion products of progesterone derivatives were obtained by Krisch- enowski and Kieslich 42 during screening of 131 microor- ganisms under aerobic conditions. While fermentation of 17~-hydroxyprogesterone with Co~nespora melonis CBS 16260 yielded 8[3,17~x-dihydroxyprogesterone (7), fermen- tation of 17o~-acetoxyprogesterone furnished 1113,1213- dihydroxy-17e~-acetoxyprogesterone (8) in addition to the known l113-hydroxy compound. Incubation of 3[3- acetoxycholest-5-ene-19-ol with Moraxella sp. gave three neutral metabolites (9-11) of which compound 9 was un- known. 43 Acidic metabolites were not formed. Thus, the authors suggested that complete removal of C17-side chain takes place prior to aromatization of the ring A in 11.
Murohisa and Iida 44 isolated a mutant H-45 of Rho- dococcus equi K-3, which produced the new compound 3-oxostigmasta-1, 4-dien-26-oic acid (12) in high yield from sitosterol (containing 40% campesterol) and three other new compounds 3-oxocampesta- 1, 4-dien-26-oic acid (13), 3-oxostigmast-4-en-26-oic acid (14), and 3-oxo- campest-4-en-26-oic acid (15) along with a number of known compounds. It should be noted that in the original paper, 44 compounds (13) and (15) have wrongly been men- tioned as ergostane derivatives. The formation of the inter- mediates helped the authors to propose a degradation path- way of sitosterol side chain by Rhodococcus equi.
Yoshihama obtained new steroids in his investigation on the preparation of steroids by microbial transformation of AD and progesterone and evaluation of their pharmaceuti- cal properties. 45 Microbial hydroxylation of AD by Acre- monium strictum NN 106 gave three known compounds, 14 ot-hydroxy-AD, 11 ~-hydroxy-AD, and 713,1 lc~-dihydroxy- AD, and two new steroids, 6[3,14~-dihydroxy-AD (16), and 613,1 lc~-dihydroxy-AD (2). 14~-Hydroxyandrost-4-ene- 3,6,17-trione (17) was prepared from 16 by oxidation. Com- pound 17 showed the highest inhibition against estrogen
Steroids, 1997, vol. 62, April 335

Review
Table 1 Hydroxylation
Substrate Microorganism Product Reference
21-Acetoxy-17cx-hydroxypregn-4- Arthrobacter simplex (i) 11t3, 21-Dihydroxypregna-1, 4,17(20)-trien-3-one 55 ene-3,11,20-trione (ii) 11 [3-Hydroxypregna-1,4,17
(20)-trien-3-one-21-hemisuccinate 11 [3,17e,21-Trihydroxypregn-4-ene-3, 20-dione 21-Acetoxy-17~-hydroxypregn-4-
ene-3,20-dione 17~-acetoxy-21-hydroxypregna-1,
4-diene-3,20-dione 17e-Acetoxypregn-4-ene-3,20-dione
Absidia orchidis 56
57
42
17c~-Acetoxy- 11 [3,21 -dihyd roxyp reg n a- 1, 4-diene,3,20-dione
(i) 17e-Acetoxy-1113-hyd roxypreg n-4-ene-3,20-dione (ii) 17e-Acetoxy-1113,1213-dihydroxypregn-4-ene-3,
20-dione &x-Androstan-3-one Cephalosporium 17e-Hydroxy-5~-androstan-3-one 58
aphidicola 5e-And rostane-3,6-dione Cephalosporium (i) &x,1713-Dihydroxy-513-androstan-6-one 58
aphidicola (ii) 1713-Hydroxyandrost-4-ene-3, 6-dione (iii) 313,5~-Dihydroxyandrost-6,17-dione (iv) 313,5~,1713-Trihydroxya ndrostan-6-one
Androsta-1,4-diene-3,17-dione 1 lcx-Hyd roxyandrosta-1,4-diene-3,17-dione
Androst-4-ene-3,17-dione Androst-4-ene-3,17-dione Androst-4-ene-3,17-dione
Androst-4-ene-3,17-dione
Androst-4-ene-3,17-dione
Androst-4-ene-3,17-dione Androst-4-ene-3,17-dione Androst-4-ene-3,17-dione
1713-(N-Butylcarbamoyl)-4-aza-5e- androst-l-en-3-one
17e,21-Dihydroxypregn-4-ene-3,20- dione
17e,21-Dihydroxypregn-4-ene-3,20- dione
17e,21-Dihydroxypregn-4-ene-3,20- dione
17e,21-Dihydroxypregn-4-ene-3,20- dione
17cx,21-Dihyd roxypreg n-4-ene-3,20- dione
24-Epibrassinolide
24-Epicastasterone
13-Ethylgon-4-ene-3,17-dione 1713-Hydroxyandrosta- 1,4-dien-3-one 17[3-Hyd roxya nd rost-4-en-3-one 313-Hydroxyandrost-5-en-17-one
1713-Hydroxyandrost-4-en-3-one
17c~-Hydroxyp reg n-4-ene-3,20-dio ne
Cochiobolus lunatus
Pycnosporium sp ATCC 12231
Aspergillus ochraceus
Curvularia lunata Acremonium stricture Acremonium
strictum NN106
59
Myrothecium striatisporum
Eurotium sp. Myrothecium Aspergillus fumigatus
14c~-Hydroxya nd rost-4-ene-3,17-dione 14~-Hydroxyand rost-4-ene-3,6,17-trione (i) 14{x-Hydroxyandrost-4-ene-3,17-dione (ii) 11cx-Hydroxyandrost-4-ene-3,17-dione (iii) 713,11cx-Dihydroxyandrost-4-ene-3,17-dione (iv) 613,14e-Dihydroxyandrost-4-ene-3,17-dione (v) 613,11c~-Dihydroxyandrost-4-ene-3,17-dione
Mucorpiriformis (i) 1713-Hyd roxya ndrost-4-en-3-one 61 (ii) 1413-Hydroxyandrost-4-ene-3,17-dione (iii) 7~-Hyd roxyandrost-4-ene-3,17-dione (iv) 1413,1713-Dihydroxyandrost-4-en-3-one (v) 7e,1413,1713-Trihyd roxyandrost-4-en-3-one 14e-Hydroxyandrost-4-ene-3,17-dione
Selenastrum capricornutum
Cunninghamella elegans
Curvularia lunata and Mycobacterium
smegmatis Curvularia lunata
Curvularia lunata and
Mycobacterium smegmatis
Beauveria bassiana
Cunninghamella echinulata
Cunninghamella echinulata
Penicillium raistrickfi Mucor piriformis Mucor piriformis Mucor piriformis
Marchantia polymorpha
Mucor piriformis
14~x-Hyd roxya nd rost-4-ene-3,17-dione 613,14cx-Dihydroxyandrost-4-ene-3,17-dione
(i) 713,11~-Dihydroxyandrost-4-ene-3,17-dione (ii) 613,11e-Dihydroxyandrost-4-ene-3,17-dione 1713-(N-butylcarbamoyl)-1 lcx-hydroxy-4-aza-&x-
androst-l-en-3-one (i) 11c~,17~,21 Trihydroxypregn-4-ene-3,20-dione (ii) 1113,17~,21-Trihydroxypregn-4-ene-3,20-dione 11 [3,17c~,21 -Trihyd roxypreg na-1,4-diene-3, 20-dione
1113,17e,21-Trihydroxypregn-4-ene-3,20-dione
11 [3,17e,21-Trihydroxypregna- 1,4-diene-3, 20-dione
11cx,17~,21-Trihydroxypregn-4-ene-3,20-dione
121~-Hydroxy-24-epibrassinolide
1213-Hydroxy-24-epicastasterone
15e-Hydroxy-13-ethylgon-4-ene-3,17-dione 14~,1713-Dihydroxyandrosta-1,4-dien-3-one 14~x-Hydroxya ndrost-4-ene-3,17-dione
(i) 3[3,7e-Dihydroxyandrost-5-en-17-one (ii) 313,7e,171~-Trihydroxyandrost-5-ene
613,1713-Dihyd roxya nd rost-4-en-3-one
(i) 7e,17o~-Dihydroxypregn-4-ene-3,20-dione (ii) 613,17e,20e-Trihyd roxypreg n-4-en-3-o ne (iii) 11e,17c~,20-Trihydroxypregn-4-en-3-one 813,17e-Dihydroxypregn-4-ene-3,20-dione
(i) 3[3,7c~-Dihyd roxypreg n-5-en-20-one (ii) 313,7e,1 le-Trihydroxypregn-5-en-20-one
17e-Hyd roxypreg n-4-ene-3,20-dione
3[3-Hydroxypregn-5-en-20-one
Corynespora Melonis CBS 16260
Pycnosporium sp. ATCC 12231
60 11 45
8
9 10 38
62
7
21
6, 63-65
22
66
67
67
68-72 73 73 61
74
61
42
42
336 S te ro ids , 1997, vo l . 62, Ap r i l

Bio t rans format ion o f steroids: Mahato and Garai
Table 1 (continued)
Substrate Microorganism Product Reference
11c(-Hydroxypregn-4-ene-3,20-dione Cephalosporium (i) 6#,11(x-Dihyd roxypreg n-4-ene-3,20-dione 75 aphidicola (ii) 66,11c(,2017,-Trihydroxypreg n-4-en-3-one
17(x-Hydroxypreg n-4-ene-3,20-dione
17(x-Hyd roxypreg n-4-ene-3,20-dione
Preg na-4,16-diene-3,20-dione
Preg na-4,16-diene-3,20-dione
(iii) 1 l(~,2013-Dihydroxypregn-4-en-3-one (i) 1213,17(x-Dihyd roxypregn-4-ene-3,20-dione 75
(ii) 66,17c(-Dihydroxypregn-4-ene-3,20-dione (iii) 613,11(x,17(x-Trihydroxypregn-4-ene-3,20-dione
(i) 7c~,17(x-Dihydroxypregn-4-ene-3,20-dione 48 (ii) 6#,17(x,20(x-Trihydroxypreg n-4-en-3-one (iii) 11e~,17e,20(x-Trihydroxypregn-4-en-3-one
(i) 14(x-Hydroxypregna-4,16-diene-3,20-dione 48 (ii) 7(x,14(x-Dihydroxypregn-4,16-diene-3,20-dione
(iii) 36,7c(,14c~-Trihydroxy-5(x-pregn-16-en-20-one (iv) 3(x,7(x,14(x-Trihyd roxy-5(x-preg n- 16-en-20-o ne
(i) 7(x,14(x-Dihyd roxypregna-4,16-diene-3,20-dione 61 (ii) 14(x-Hyd roxypregna-4,16-diene-3,20-dione
(iii) 313,7(x,14c~-Trihydroxy-5(x-preg n-16-en-20-one
Pregna-4,16-diene-3,20-dione Preg n-4-ene-3,20-dione
Pregn-4-ene-3,20-dione
Preg n-4-ene-3,20-dione
Preg n-4-ene-3,20-dione Preg n-4-ene-3,20-dione Preg n-4-ene-3,20-dione
Preg n-4-ene-3,20-dione
Pregn-4-ene-3,20-dione
Pregn-4-ene-3,20-dione
Pregn-4-ene-3,20-dione Pregn-4-ene-3,20-dione Preg n-4-ene-3,20-dione
Pregn-4-ene-3,20-dione
Pregn-4-ene-3,20-dione
Pregn-4-ene-3,20-dione
Pregn-4-ene-3,20-dione Pregn-4-ene-3,20-dione
Cephalosporium aphidicola
Mucor piriformis
Mucor piriformis
Mucor piriformis
Mucor piriformis Mucor piriformis
Rhizopus nigricans
Aspergillus ochraceus
Mortierella isobellina Rhizopus nigricans Aspergillus
ochraceus Acremonium stricture
Cochliobolus lunata
Aspergillus fumigatus
Aspergillus ochraceus Mucor piriformis Cochliobolus specifer
Cephalosporium aphidicola
Cyanidiophyceac ernersonii
Muriella aurantiaca
Galdierla sulphuraria Scemedesmus
capricortum
(iv) 3c(,7(x,14~-Trihydroxy-5(x-preg n-16-en-20-one 14(x-Hydroxypregna-4,16-diene-3,20-dione 73 (i) 513,14c(-Dihydroxypreg nane-3,2O-dione 61
(ii) 14(x-Hydroxypreg n-4-ene-3,20-dione (iii) 66,14c~-Dihydroxypreg n-4-ene-3,20-dione (iv) 7(x,14(x-Dihydroxypreg n-4-ene-3,20-dione (v) 713,14(x-Dihydroxypregn-4-ene-3,20-dione (i) 1 l(x-Hydroxypregn-4-ene-3,20-dione 76
(ii) 1 l(x-Hydroxy-5(x-pregnane-3,20-dione (iii) 66,1 l(x-Dihydroxypregn-4-ene-3,20-dione 1 l(x-Hydroxypreg n-4-ene-3,20-dione 77
14(x-Hydroxypreg n-4-ene-3,20-dione 1 l(x-Hydroxypreg n-4-ene-3,20-dione 1 ltx-Hyd roxypreg n-4-ene-3,20-dione
29 78 40
(i) 713,156-Dihydroxypregn-4-ene-3,20-dione 45 (ii) 66,1 l(x-Dihydroxypregn-4-ene-3,20-dione
(iii) 11c~,1513-Dihydroxypregn-4-ene-3,20-dione (iv) 613,11(x,17c~-Trihydroxypregn-4-ene-3,20-dione (v) 11 e,156,17(x-Trihydroxypregn-4-ene-3,20-dione
(vi) 7{3,1513,17(x-Trihydroxypreg n-4-ene-3,20-dione (i) 7c~,1 ll3-Dihydroxypregn-4-ene-3,20-dione 79
(ii) 14(x-Hydroxypregn-4-ene-3,11,20-trione (i) 1 lc~-Hydroxypregn-4-ene-3,20-dione 80
(ii) 156-Hydroxypregn-4-ene-3,20-dione (iii) 76-Hydroxypregn-4-ene-3,20-dione (iv) 76,15i~-Dihydroxypregn-4-ene-3,20-dione (v) 1 lc¢,156-Dihydroxypregn-4-ene-3,20-dione 1 lc~-Hydroxypregn-4-ene-3,20-dione 81 14(x-Hydroxypreg n-4-ene-3,20-dione 73
(i) 21-Hyd roxypregn-4-ene-3,2O-dione 82 (ii) 17(x-Hydroxypregn-4-ene-3,20-dione
(iii) 116-Hydroxypregn-4-ene-3,20-dione (iv) 11c(,17c~-Dihydroxypregn-4-ene-3,20-dione (v) 1113,17c(,21-Trihydroxypregn-4-ene-3,20-dione (vi) 11(x,17(x,21-Trihydroxypregn-4-ene-3,20-dione
(i) 613,1 l~x-Dihydroxypregn-4-ene-3,20-dione 75 (ii) 1 l(x-Hydroxypregn-4-ene-3,20-dione
(iii) 126,17-Dihydroxypregn-4-ene-3,20-dione (iv) 206-Hydroxypreg n-4-en-3-one (v) 66,1 l(x,2013-Trihydroxypregn-4-en-3-one (i) 26-Hydroxypregn-4-ene-3,20-dione 83
(ii) 66-Hydroxypregn-4-ene-3,20-dione (iii) 9(x-Hydroxypreg n-4-ene-3,20-dione (iv) 14(x-Hydroxypreg n-4-ene-3,20-dione (v) 16(x-Hydroxypregn-4-ene-3,20-dione (i) 613-Hydroxypregn-4-ene-3,20-dione 83
(ii) 15(x-Hydroxypregn-4-ene-3,20-dione 6{3-Hydroxypregn-4-ene-3,20-dione 83
(i) 26-Hydroxypregn-4-ene-3,20-dione 83 (ii) 66-Hydroxypreg n-4-ene-3,20-dione
S te ro ids , 1997, vo l . 62, Ap r i l 337

Review
Table 2 Side-chain cleavage
Substrate Microorganism Product Reference
313-Acetoxy-19-hydroxy- Moraxella sp. (i) 19-Hydroxy-5(~-androst-1-ene-3,17-dione 43 cholest-5-ene (ii) 19-Hydroxyandrost-4-ene-3,17-dione
313-Acetoxy-19-hydroxy- cholest-5-ene
(iii) 9(x,19-Dihydroxyandrost-4-ene-3,17-dione (iv) 3-Hyd roxyestra-1,3,5(10)-trien-17-one
(i) 3-Hydroxyestra-1,3,5(10)-trien-17-one 84 (ii) 19-Hydroxy-5(x-androst-1-ene-3,17-dione
(iii) 19-Hydroxyandrost-4-ene-3,17-dione (iv) 9(x,19-Dihydroxyand rost-4-ene-3,17-dione (v) 19-Hydroxya ndrosta ne-3,17-dione
Androst-4-ene-3,17-dione 85 Androsta-1,4-diene-3,17-dione 86 Androsta-1,4-diene-3,17-dione 20
Moraxella sp.
Cholesterol Mycobacterium sp. NRRLB-3805 Cholesterol Rhodococcus corallina Cholesterol Arthrobacter simplex and
Mycobacterium sp. NRRLB-3683 Cholesterol Rhodococcus equi (i) Androstane-3,17-dione 87
(ii) Androsta-1,4-diene-3,17-dione Cholesterol Mycobacterium sp. (i) Androst-4-ene-3,17-dione 88
(ii) Androsta-1,4-diene-3,17-dione (iii) 17#-Hyd roxyandrosta-l,4-dien-3-one (iv) 17#-Hydroxyandrosta-1,4-dien-3-one
313-Hyd roxya ndrost-5-en-17-one Cholesterol
Cholesterol Cholesterol
2e,3e-Dihydroxy- 5(~-cholestan-6-one
19-Hyd roxy-cholesterol
1713-Hydroxyandrost-4-en-3-one (i) Androsta-1,4-diene-3,17-dione
(ii) Androst-4-ene-3,17-dione (i) 2(x,3(x,6(x-Trihyd roxy-5(x-a nd rosta n- 17-o ne
(ii) 2c(-Hydroxyandrost-4-ene-3,17-dione (i) Estra-l,3,5(10)-trien-3-ol
(ii) 2(3-Hyd roxy-1,3,5(10)-estra-trien-17-yl)- propionic acid
(iii) 2-Methyl-6(3-hydroxy-1,3,5(10)-estratriene-17-yl)- heptanoic acid
(iv) 2(3-Hyd roxy-1,3,5(10),17-estra-tetraen-17-yl)- propionic acid (i) 2(3-Hyd roxy-1,3,5(10),17-estra-tetraen-17-yl)- propionic acid (ii) 2,3-Dimethyl-6-(3-hyd roxy-1,3,5(10)-estratriene- 17-yl)- heptanoic acid
19-Hydroxy-campesterol
Mycobacterium fortuitum NRRLB-8153
Mycobacterium sp. Rhodococcus equi
Mycobacterium vaccae
Rhodococcus mutant k-3
Rhodococcus mutant k-3
89
17 90
91
41
41
Lithocholic acid Mycobacter ium sp. 20(x-Hydroxymethylpregn-4-en-3-one 92 313-Methoxymetho- M y c o b a c t e r i u m 315-Methoxymethoxy-21 -hydroxy- 93
xyergosta-5,7,22-triene 20-methylpregna-5,7-diene Pregn-4-ene-3,20-dione Pseudomonas sp. (i) Androsta-1,4-diene-3,17-dione 94
(ii) 17L3-Hydroxyandrosta-1,4-dien-3-one Pregn-4-en-3,20-dione Cephalosporium aphidicola 1713-Acetoxyandrost-4-en-3-one 75 Sitosterol Mycobacterium fortuitum 9(x-Hydroxyandrost-4-ene-3,17-dione 13 Sitosterol Arthrobacter oxydans (i) 3-Oxochol-4-en-24-oic acid 95
(ii) 27-Norcholest-4-ene-3,24-dione Sitosterol Rhodococcus equi k-3 (i) 3-Oxo ergosta-1,4-dien-26-oic acid 44
(ii) 3-Oxo ergost-4-en-26-oic acid (iii) 20-Ca rboxypreg n-4-en-3-one (iv) 20-Ca rboxypreg na-l,4-dien-3-one (v) Androst-4-en-3,17-dione
(vi) And rosta-1,4-diene-3,17-dione (vii) Propionic acid
(viii) Acetic acid Sitosterol Mycobacterium sp. 9c~-Hydroxya ndrost-4-ene-3,17-dione 15 Sitosterol Mycobacterium NRRLB-3683 Androsta-1,4-diene-3,17-dione 96 Sitosterol Arthrobacter simplex (i) Androst-4-ene-3,17-dione 97
(ii) Androsta-1,4-diene-3,17-dione Sitosterol M y c o b a c t e r i u m Androst-4-ene-3,17-dione 98 Sitosterol Mycobacterium v a c c a e Androst-4-ene-3,17-dione 19 Solasodiene Mycobacterium sp. NRRLB-3805 Androst-4-ene-3,17-dione 99 Sterol Mycobacterium v a c c a e Androsta-1,4-diene-3,17-dione 16 Sterol Mycobacterium fortuitum 9(x-Hydroxyandrost-4-ene-3,17-dione 12, 100 Sterol Mycobacterium NRRLB-3805 Androst-4-ene-3,17-dione 101 Sterol Mycobacterium fortuitum 9(x-Hydroxyandrost-4-ene-3,17-dione 102 Sterol Mycobacterium v a c c a e Androsta-1,4-diene-3,17-dione 103 Sterol Arthrobacter simplex Androsta-1,4-diene-3,17-dione 104 Sterol Mycobacterium sp. (i) Androst-4-ene-3,17-dione 105
(ii) Androsta-1,4-diene-3,17-dione
338 Steroids, 1997, vol. 62, April

Biotransformation of steroids: Mahato and Garai
Table 3 Dehydrogenation
Substrate Microorganism Product Reference
31~,-Acetoxy-513-preg n- Mycobacterium Pregna-4,16-diene-3,20-dione 106 16-en-20 -one N R R L B - 3 8 0 5
21-Acetoxy-1113,17(x- Arthrobactersimp/ex 21-Acetoxy-111%17~-dihydroxy-6~-methylpreg na- 107 dihydroxy-6c~-methyl 1,4-diene-3,20-dione preg n-4-ene-3,20-dione
513-Androstane-3,17-dione Mycobacterium sp. (i) Androst-4-ene-3,17-dione 108 (ii) 313-Hydroxy-5c~-androstan-17-one
Androsta-1,4-diene-3,17-dione Androsta-1,4-diene-3,17-dione
And rost-4-ene-3,17-dione And rost-4-ene-3,17-dione
Aspergillus fumigatus Acetone dried cell of
A. fumigatus Chenodeoxycholic acid B a c i l l u s 3a-Hydroxy-7-oxo-513-cholan-24-oic acid 109 Chenodeoxycholic acid Bacillus sp. 3ct-Hydroxy-7-oxo-513-cholan-24-oic acid 110 Cholesterol Mycobacterium sp. Cholest-4-en-3-one 88 Cholesterol Rhodococcus e q u i Cholest-4-en-3-one 90 Cholesterol Escherichia (i) Cholest-4-en-3-one 111
(ii) Cholest-5-en-3-one Cholesterol Nocardia r u b r a Cholest-4-en-3-one 112 Cholesterol Arthrobacter simplex Cholest-4-en-3-one 113 Cholesterol Rhodococcus Cholest-4-en-3-one 34
erythropolis Cholesterol Rhodococcus e q u i Cholest-4-en-3-one 114 Cholic acid Bacillus (i) 3(x-Hydroxy-7,12-dioxo-513-cholan-24-oic acid 109
(ii) 3c(,12ct-Dihydroxy-7-oxo-513-cholan-24-oic acid Cholic acid Arthrobacter simplex (i) 3c~,7(x-Dihydroxy-12-oxo-513-chotan-24-oic acid 52
(ii) 7c(,12ct-Dihydroxy-3-oxo-4-cholenoic acid Cholic acid Bacillus sp. TTUR-2 3(x,7a-Dihydroxy-12-oxo-5~-cholan-24-oic acid 115 Cholic acid Bacillus sp. (i) 3(x,7(x-Dihydroxy-12-oxo-513-cholan-24-oic acid 110
313,1713-Dihydroxy-5ct-androstane 3ct-Hydroxy-5(x-androstan-17-one
3,3-(2,2-Dimethyltrimethyl- enedioxy)-5-10ct-epoxy-5(x- estr-9(11 )-en-1713-ol
3(x-Hydroxy-513-androstan-17-one 313-Hydroxypregn-5-en-20-one Lithocholic acid 19-Nor-1713-hydroxy-androst-
4-en-3-one Sitosterol
(ii) 3a,12ct-Dihydroxy-7-oxo-513-cholan-24-oic acid (iii) 3ct-Hydroxy-7,12-dioxo-513-cholan-24-oic acid Androst-4-ene-3,17-dione
(i) Androst-4-ene-3,17-dione (ii) 5a-Androsta ne-3,17-dione
3,3-(2,2-Dimethyltrimethylenedioxy)-5,10tx- epoxy-5a-estr-9(11 )-en-17-one
1113,17(x,21-Trihydroxypregn- 4-ene-3,20-dione
1113,17(x-21-Trihydroxypregn- 4-ene-3,20-dione
1113,17(x,21-Trihydroxy-6c~- methyl-pregn-4-ene-3,20-dione
1113,17ct,21 -Trihydroxypreg n- 4-ene-3,20-dione
1113,17ct,21-Trihydroxypreg n- 4-ene-3,20-dione
Ursodeoxycholic acid
38 38
Nocardia rubra 112 Mycobacterium sp. 108
Mycobacterium NRRLB-3683
Mycobacterium sp. Nocardia rubra Mycobacterium sp. Rhodococcus
sp. DSM 92344 Rhodococcus equi k-3
Arthrobacter simplex
Corynebacterium simplex
Corynebacterium simplex
Corynebacterium equi
Arthrobacter simplex
(i) And rost-4-ene-3,17-dione Pregn-4-ene-3,20-dione 3-Oxo-4-cholenoic acid
(i) 3-Hydroxyandrosta-1,3,5(10)-trien-17-one (ii) Androsta-1,3,5(10)-triene-3,1713-diol (i) Sitosta-l,4-dien-3-one
(ii) Sitos-4-en-3-one 1113,17(~,21-Trihydroxypregna-1,4-diene-3,
20-dione 1113,17a,21-Trihydroxypregna-1,4-diene-3,
20-dione 1113,17(x,21 -Trihydroxy-6(x-methyl-pregna-1,
4-diene-3,20-dione 1113,17(x,21-Trihydroxypregna-1,4-diene-3,
20-dione 1113,17c~,21-Trihydroxypregna-1,4-diene-3,
20-dione 713-Hydroxy-3-oxo-513-cholan-24-oic acid Bacillus sp.
116
92 112 92
117
44
118
36
36
119, 120
121,122
123
synthetase (aromatase). It also exhibited aromatase inhibi- tion in rat ovaries, depression in serum estrogen levels, and strong antitumor activity against human ovarian cancer on nude mice. Hydroxylation of progesterone by the same fun- gal strain yielded three dihydroxyprogesterones and three t r ihydroxyprogesterones, among which 713,1513,17a- trihydroxyprogesterone (18) was claimed to be a new com- pound. The compound (18) has also been prepared by Mahato et al. 46 by microbial hydroxylation of 17c~- hydroxyprogesterone by Aspergillus fumigatus. It is re-
ported to have the property of suppressing ovulation and showing high differentiation induction activity in bone mar- row leukemia MI cells. 47 The compound also showed strong immunosuppressive activity on mouse lymphocytes and weak toxicity against lymphocytes compared to progester- one. 45
Madyastha and Joseph 48 obtained two new steroids, 3 13,7c~, l 4e t - t r ihydroxy-5 ot-pregn- 16-en-20-one (19) 3o~,7o~, 14et-trihydroxy-5o~-pregn- 16-en-20-one (20) by transformation of 16-dehydroprogesterone by Mucor piri-
Ste ro ids , 1997, vo l . 62, Ap r i l 339

Review
Table 4 Miscellaneous reaction
Substrate Microorganism Product Reference
313-Acetoxy-19-hydroxy- Moraxella sp. (i) 19-Hydroxycholestan-3-one 84 cholest-5-ene ~, c~'-bipyridyl (ii) 19-Hydroxycholest-4-en-3-one
5c~-Androstane-3,17-dione
Androsta-1,4-diene-3,17-dione
5~-Androstan-17-one
(iii) Cholest-5-ene-313,19-diol (i) 1713-Hyd roxy-5a-androstan-3-one 58
(ii) 17~-Acetoxyandrosta n-3-one (i) 1713-Hydroxyandrosta-1,4-dien-3-one 124
(ii) Androst-4-ene-3,17-dione (iii) 1713-Hydroxya ndrost-4-en-3-one (i) 5e-Androstan-1713-ol 58
Androsta-1,4-diene-3,17-dione Androst-4-ene-3,11,17-trione Androst-4-ene-3,17-dione Androst-4-ene-3,17-dione
Chenodeoxycholic acid
Cholesterol Cholic acid
Cholic acid
Cholic acid
Deoxycholic acid
Digitoxin Digitoxigenin
3~-Hyd roxy-5~-androstan-17-one 313-Hydroxyandrost-5-en-17-one
17~-Hydroxypregn-4-ene- 3,20-dione
17~-Hydroxypregn-4-ene- 3,20-dione
Lithocholic acid
Lithocholic acid 3-Methoxy-8,14-secoestra-
1,3,5(10),9(11 )-tetraene- 14,17-dione
I~-Methyldigitoxin Pregn-4-ene-3,20-dione Preg n-4-ene-3,20-dio ne
Sitosterol
Cephalosporium aphidicola
Marchantia polymorpha
Cephalosporium aphidicola
Mycobacterium sp. NRRLB-3683 Marchantia polymorpha Aspergillus fumigatus Acetone dried cells
of A. fumigatus Alcaligen recti
Arthrobacter-82 A/ca/igen recti
Arthrobacter simplex
Corynebacterium
Arthrobacter simplex
Digitalis lunata Pergularia tomentosa
Mycobacterium sp, Mucorpiriformis sp.
Nocardia DSM-43298
Mucor piriformis
Arthrobacter simplex
Cunninghamela blackesleena Kloeekera magma
Digitalis Cyanidiophyceae caldium Scenedesmus
quadricauda Rhodococcus equi k-3
(ii) 5c~-And rosta ne- 1713-acetate 1713-Hydroxyandrost-4-en-3-one 18 17c~-Hydroxyandrost-4-ene-3,11 -dione 124 1717,-Hydroxyandrost-4-en-3-one 38
(i) 1713-Hydroxyandrost-4-en-3-one 38 (ii) 1713-Hydroxyand rosta-l,4-dien-3-one (i) 3~-Methoxy-7~,12~-dihydroxy-5~-cholan- 125
24-oic acid (ii) 3e-Methoxy-7~-hydroxy-5~-cholan-24-oic acid (iii) 3cx-Methoxy-713-hydroxy-513-cholan-24-oic acid (iv) 3cx-Methoxy-12ct-hydroxy-513-cholan-24-oic
acid (v) 7~-Hydroxy-3-oxo-4-cholenoic acid (vi) 7c~-Hydroxy-3-oxo-4-cholenoic acid
3-Oxo-23,24-dinorchola-1,4-diene-22-oic acid 126 (i) 3~-Methoxy-7~,12~-dihydroxy-513-cholan- 125
24-oic acid (ii) 3tx-Methoxy-7c~-hydroxy-513-cholan-24-oic acid (iii) 3~-Methoxy-713-hydroxy-513-cholan-24-oic acid (iv) 3e-Methoxy-12~-hydroxy-513-cholan-24-oic acid (v) 7e,12~-Dihydroxy-3-oxo-4-cholenoic acid (i) 3,12-Dioxo-23,24-dinorchola-4,6-dienoic acid 52
(i i) 7c~-Hydroxy-3,12-dioxo-23,24-dinorcholenoic acid (iii) 3~,7e-Dihydroxy-12-oxo-513-23,24-
dinorcholan-22-oic acid (iv) 7tx,12~-Dihydroxy-3-oxo-513-cholan-24-oic acid (v) 7~,12~-Dihydroxy-3-oxo-23,24-dinorchol-
4-enoic acid (vi) Methyl-3~,7~,12~-trihydroxy-513-cholan-24-oate (vii) 21~-Hydroxy-3,12-dioxo-23,24-dinorcholan-
4,6-dienoic acid 3e,12~-Dihydroxy-7-oxo-23,24-dinor-513-cholan- 127
22-oic acid (i) Methyl-3~,12~-dihydroxy-513-cholan-24-oate 128
(ii) 3,12-Dioxo-513-cholan-24-oic acid (iii) 3cx-Hydroxy-12-oxo-51~-cholan-24-oic acid Digoxin 129
(i) Periplogenin 130 (ii) l~-Hydroxy digitoxigenin
(iii) Uzarigenin 3~-hydroxy-5¢x-androstan-17-one 108
(i) 3~,1713-Dihydroxyandrost-5-ene 61 (ii) 313-Hydroxyandrost-5-ene-7,17-dione
(iii) 313,1713-Dihyd roxyandrost-5-ene-7-one (i) 9,10-Seco-3,17~-dihydroxypregna-1,3,5 131
(10)-triene-9,20-dione (ii} 9,10-Seco-3,17c~,20-trihydroxy-pregna-1,
3,5(10)-trien-9-one 17e,20{x-Dihyd roxypreg n-4-en-3-one
(i) Methyl-3~-acetoxy-5~-cholan-24-oate (ii) n-Butyl 3~-acetoxy-513-cholan-24-oate
Lithocholic acid 3~-O-13-D-glucopyranoside 17~-Hyd roxy-3-met hoxy-8,14-secoestra-1,
3,5(10),9(11 )-tetraen-14-0ne
13-Methyl digoxin 20c~-Hyd roxypreg n-4-en-3-o ne 3-Hydroxy-9,10-secopregna-1,3,5(10)-
triene-9,20-dione (i) Stigmasta-1,4-dien-3-on-26-oic acid
(ii) Stigmast-4-en-3-on-26-oic acid
48, 61
128
50 132
133 83 83
44
3 4 0 Ste ro ids , 1997, vo l . 62, A p r i l
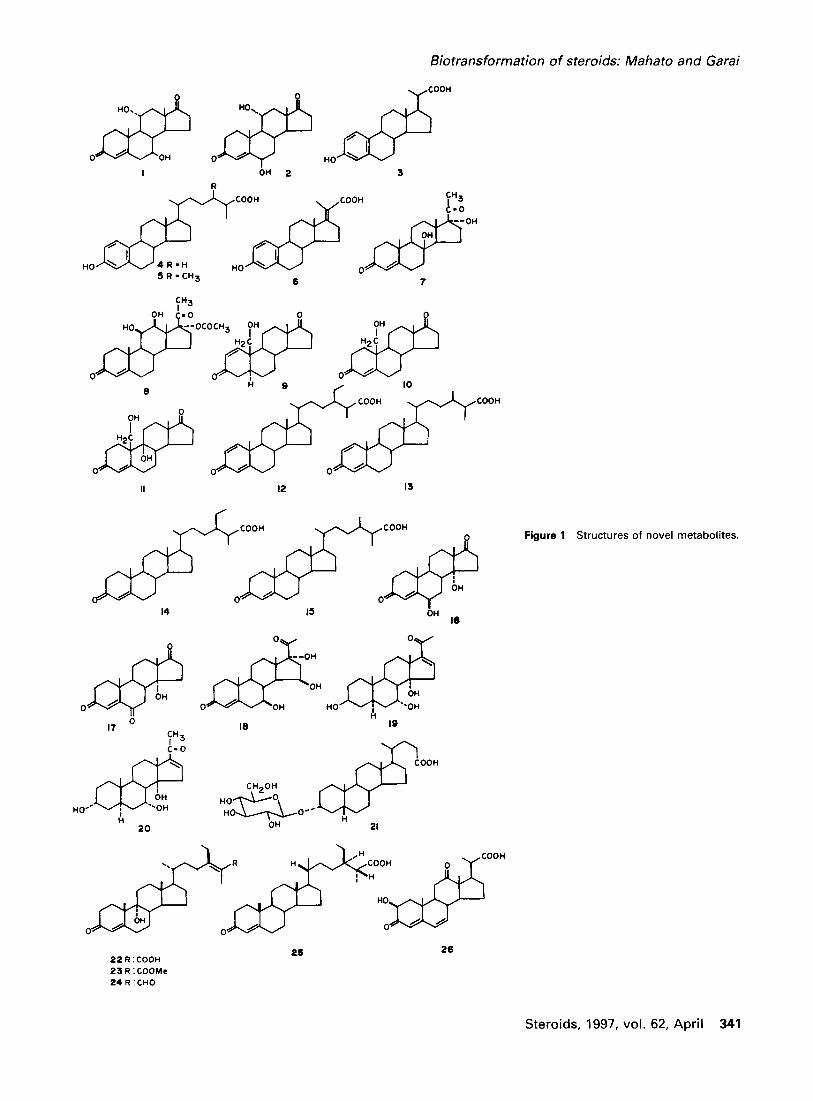
Biotransformation of steroids: Mahato and Garai
I OH 2 3
R
~ J ~ C O O H ~llr~C00H i730
5 R • CH 3 6 7
ICH 3
8 H 9 I0
o~'"~v ~" g 0-~ ~'..~ V 0 "~ ~ V
II 12 13
0
0 17 ~H3 C-O
H 0 " ~ H 20
~ ~ ~ : 0 M COOH COOH 0
14 15 OH 16
--OH
OH OH 0 HO
18 19
COOH
CH20H ,,, . , , '" , . , , ,~ ~ . o ~ O , L I J
OH 21
"~ . ~ .H . ICOOH
2 5 26 2 2 R :COOH 2 3 R : COOMe 2 4 R : CHO
Figure 1 Structures of novel metabolites.
Steroids, 1997, vol. 62, April 341

Review
formis. The structure of 20 was determined by X-ray crys- tallography. 49 A new bile acid glycoside lithocholic acid 3a-O-13-D-glucopyranoside (21) was produced by Cunning- hamella blakesleena ST-22 from lithocholic acid. 5° In a study on the sterol-transforming ability of fast growing My- cobacterium strains obtained by in vivo genetic recombina- tion experiment Ambrus et al. 5 ~isolated novel intermediates of microbial side-chain degradation of sitosterol. They reported the structure and stereochemistry of the new 26-oxygenated steroid derivatives 22-25, The molecular structure of compound 25 was determined by X-ray crys- tallography, and its absolute configuration was deduced to be 24(R) and 25(R). A novel metabolite, 213-hydroxy-3,12- dioxo-23,24-dinorchola-4,6-dienoic acid 26 was produced along with a few other metabolites by transformation of cholic acid by Arthrobacter simplex. The pathway of for- mation of these metabolites of cholic acid has also been proposed. 52'53 Cholic acid-induced N-methylhydantoin syn- thesis by a strain ofAlcaligens recti in nutrient medium has been reported. 54 The formation of this product has been attributed to the amino acids contained in the nutrient me- dium under the catalysis of various enzymes at different stages in the presence of cholic acid. It is noteworthy that cholic acid and other bile acids are present in the mamma- lian liver where creatine is synthesized. It has been sug- gested that in mammalian liver, cholic acid acts as an inducer for the synthesis of creatinine, the immediate pre- cursor of N-methylhydantoin.
Future possibilities
Since 1950, steroids have been the traditional field for in- dustrially used microbial transformation, and remarkable progress already has been made. However, further develop- mental activities relating to controlled multiple transforma- tion for the purpose of cost reduction are anticipated. Con- tinuous emphasis on the application of genetic engineering of microorganisms for their improvement as steroid- transforming agents is expected. Development of new ste- roids possessing useful biological activity may receive greater attention. Continuing attempts are expected for uti- lization of sterol fractions obtained as industrial by-product in the production of steroid intermediates. There is a distinct possibility of further development on conducting two or more microbial steps in a single-step fermentation using mixed culture or immobilized mixed culture. The commer- cial success of microbial steroid transformation so far has been achieved by flourishing vegetative cell cultures. Among the alternative processes, the present interest centers on the development of process using immobilized cells for industrial exploitation. Further emphasis on this process is expected to achieve the objective. Further developmental studies on the use of cyclodextrins, nonaqueous, low water, or mixed solvent systems in microbial transformations of water insoluble organic compounds are anticipated.
Microbial hydroxylations of various steroid substrates are shown in Table 1. Tables 2 and 3 show the side-chain cleavages and the dehydrogenations, respectively. Table 4 shows miscellaneous reactions. In each table, the substrates are arranged alphabetically.
Acknowledgments Financial supports from the Council of Scientific and In- dustrial Research (CSIR), New Delhi, India in the form of Senior Research Fellowship (SG) and Emeritus Scientist (SBM) are gratefully acknowledged.
References 1. Murray HC, Peterson DH (1952). Oxygenation of steroids by Mu-
corales fungi. U.S. Patent 2602769 (Upjohn Co., Kalamazoo, Michigan, USA).
2. Mahato SB, Mukherjee A (1984). Steroid transformations by mi- croorganisms. Phytochemistry 23:2131-2154.
3. Mahato SB, Banerjee S (1985). Steroid transformation by micro- organisms II. Phytochemistry 24:1403-1421.
4. Mahato SB, Banerjee S, Podder S (1989). Steroid transformations by microorganisms III. Phytochemistry 28:7-40.
5. Mahato SB, Mazumder I (1995). Current trends in microbial ste- roid biotransformation. Phytochemistry 34:883-898.
6. Chincholkar SB, Sukhodol'skaya GV, Baklashova TG, Kosch- cheenko KA (1992). Peculiarities of the 11 [3-hydroxylation of ste- roid compounds by the mycellium of Curvularia lunata VKMF- 644 in the presence of [3-cyclodextrin. Prikl Biokhim Mikrobiol 28:685-693.
7. Dtugonski J, Bartnicku K, Choyeeka V, Sedlaezek L (1992). Sta- bilization of steroid 11-hydroxylation activity of Cunninghamella elegans protoplasts in organic osmotic stabilizers. World J Micro- biol Biotechnol 8:50(0504.
8. Yoshioka H, Asada S (1994). 14ct-Hydroxy-4-androstene-3,17- dione manufacture with Myrothecium. Japanese Patent 06, 153. 987 (Nippon Kayaku, KK).
9. Asada S (1994). Microbial manufacture of 14ct-hydroxy-4- androstene-3,17-dione. Japanese Patent 06, 225, 791 (Nippon Kay- aku, KK).
10. Yoshioka H, Asada H, Fujita S (1994). Process for production of 6[3, 14c~-dihydroxy-4-androstene-3,17-dione. European Patent 599, 658 (Nippon Kayaku, KK).
11. Nakakoshi M, Yoshihama M, Nakamura H, Kumazawa E, Kabaya- shi N, Kawashima T, Ishiguro S, Seya M (1993). Preparation of 14c~-hydroxy-4-androstene-3,6,17-trione from 4-androstene-3,17- dione. Japanese Patent 0576, 387 (Snow Brand Milk Prod. Co. Ltd., Tokyo).
12. Birke M, Hoerhold C, Groh H, Deppmeyer V, Naumann H, Schild- batch A, Daehnhardt S, Schaner F (1992). Microbial manufacture of 9a-hydroxyandorst-4-ene-3,17-dione from sterols. (East) Ger- man Patent 298, 278 (Jena Pharm. Gmbh., Jena).
13. Birke M, Schildbach A, Heller I, Hoerhold C, Seidel L, Atart P (1992). Microbial manufacture and recovery of 9c~-hydroxy- androst-4-ene-3,17-dione. (East) German Patent 298, 279 (Jena Pharm. Gmbh., Jena).
14. Voishvillo NE, Turuta AM, Kamemitskii AV, Dzhlantiashvili ND, Daeheva-Spasova VK (1992). Microbiological transformations of 3[3-hydroxy-5a-H-pregnanes to their A4-3-keto-9a-hydroxy de- rivatives. Khim-Farm Zh 26:64-68.
15. Borman EA, Redikulifsev Yu V, Koshcheenko KA, Turuta AM, Kamernitskii AV (1992). Transformation of sitosterol to 9a- hydroxy-androstene-dione by Microbacterium sp. 207 cells in the presence of an adsorption resin. Prikl Biokhim Mikrobiol 28:551- 556.
16. Reiche R, Heller I, Hoerhold C, Gotlsehaldf B (1992). Manufac- ture of androsta-l,4-diene-3,17-dione from sterols with Mycobac- terium. (East) German Patent 300, 364 (VEBJENA PHARM).
17. Liu WH, Kuo CW, Wu KL, Lee CY, Hsu WY (1994). Transfor- mation of cholesterol to testosterone by Mycobacterium sp. J lnd Microbiol 13:167-171.
18. Hung B, Falero A, Llanes N, Perez C, Ramirez MA (1994). Tes- tosterone as biotransformation product in steroid conversion by Mycobacterium sp. Biotechnol Lett 16:497-500.
19. Spassov G, Pramatarova V, Vlahov R, Reinhold G (1993). Bio- conversion of sitosterol to androsteue-dione by mutants of Myco- bacterium vaccae. Dokl Bulg Acad Nauk 46:123-126.
20. Lee CY, Chen CD, Liu WH (1993). Production of androsta-l,4-
342 Steroids, 1997, vol. 62, April

diene-3,17-dione from cholesterol using two-step microbial trans- formation, Appl Microbial Biotechnol 38:447-452.
21. Ghanem KM, Yusef HH (1992). Transformation of Reichstein's compound S into prednisolone by a mixed culture under some physiological conditions. Rev Latinoam Microbial 34:107-114.
22. Ghanem KM, Yusef HH (1992). Some nutritional requirements of a mixed culture transforming Reichstein's compound S into pred- nisolone. Can J Microbial 38:753-757.
23. Ghanem KM, EL-Aassar SA, Yusef HH (1992). Transformation of Reichstein's compound S into prednisolone by immobilized mixed cultures. J. Chem Technol Biotechnol 54:115-121.
24. Klibanov AM (1986). Enzymes that work in organic solvents. Chemtech 16:354-359.
25. Hailing PJ (1987). Biocatalysis in multiphase reaction mixtures containing organic liquids. Biotechnol Adv 5:47-84.
26. Zaks A, Klibanov AM (1985). Enzyme-catalyzed processes in or- ganic solvents. Proc Natl Acad Sci USA 82:3192-3196.
27. Zaks A, Klibanov AM (1988). The effect of water on enzyme action in organic media. J Biol Chem 263:8017-8021.
28. Sakuvai T, Margolin AL, Rkussell AJ, Klibanov AM (1988). Con- trol of enzyme enantioselectivity by the reaction medium. J Am Chem Sac 110:7236-7237.
29. Holland HL, Poddar S, Tripet B (1992). Effect of cell immobili- zation and organic solvents on sulfoxidation and steroid hydrox- ylation by Mortierella isabellina. J lnd Microbial 10:195-197.
30. Ahmad S, Garg SK, Johri BN (1992). Biotransformation of sterols: Selective cleavage of the side chain. Biotechnol Adv 10:1-67.
31. Dias ACP, Cabral JMS, Pinheiro HM (1994). Sterol side-chain cleavage with immobilized Mycobacterium cells in water immis- cible organic solvents. Enzyme Microb Technol 16:708-714.
32. Miethe P, Gruber R, Mohr KH (1992). Lyotropic liquid crys- tals--A new possibility of biocatalysis in organic media. Bioengi- neering 8:38-4 1.
33. Oda S, Ohta H (1992). Microbial transformation on interface be- tween lyophilic carriers and hydrophobic organic solvents. Biosci Biotechno! Biochem 56:2041-2045.
34. Jadoun J, Bar R (1993). Microbial transformations in a cyclodex- trin medium. Part 3. Cholesterol oxidation by Rhodococcus erythropolis. Appl Microbial Biotechnol 40:230-240.
35. Jadoun J, Bar R (1993). Microbial transformation in a cyclodextrin medium. Part 4. Enzyme vs Microbial oxidation of cholesterol. Appl Microbial Biotechnol 40:477-482.
36. Alekhina TM, Ryzhkova VM, Gusarova TI, Kurakov VV, Klab- nichkina GA (1993). Microbial transformation of steroid 13-cyclo- dextrin inclusion compounds. Khim-Farm Zh 27:59-62.
37. Uden WV, Woeydenbag H, Prar N (1994). Cyclodextrin as a use- ful tool for the bioconversion in plant cell biotechnology. Plant Cell Tissue Organ Cult 38:103-113.
38. Banerjee S, Mukherjee E, Mahato SB (1993). Metabolism of an- drost-4-ene-3,17-dione by A.wergillus fumigatus. J Chem Res (Suppl): 236-237.
39. Garai S, Banerjee S, Mahato SB (1995). Selective 1-dehydroge- nation of progesterone by Aspergillus fumigatus. J Chem Res (Suppl): 408-409.
40. Houng JY, Chiang WP, Chen KC, Tiu C (1994). lla-Hydroxy- lation of progesterone in bi-phasic media using alginate-entrapped Aspergillus ochraceus gel beads coated with polyurea. Enzyme Microb Technol 16:485-491.
41. Murohisa T, lida M (1993). Studies on microbial transformation (XXVI). Microbial degradation of 19-hydroxysterol side-chains. J Ferment Bioeng 75:13-17.
42. Krischenoski D, Kieslich K (1993). Two novel microbial conver- sion products of progesterone derivatives. Steroids 58:278-281.
43. Sankar VN, Row TNG, Madyastha KM (1993). Evidence for a new pathway in the microbial conversion of 3[3-acetoxycholest-5- en-19-ol into estrone. J Chem Sac Perkin Trans 1:2233-2236.
44. Murohisa T, Iida M (1993). Studies on microbial transformation (XXVII). Some new intermediates in microbial side-chain degra- dation. J Ferment Bioeng 76:174-177,
45. Yoshihama M (1993). Microbial hydroxylation of steroid hor- mones and their pharmaceutical applications. Yukijirushi Nyugyo Kenkyusho Hokoku 99:1-70.
46. Mahato SB, Mukherjee E, Banerjee S (1994). Pharmaceutically important steroids by microbial biotransformations with Aspergil-
Biotransformation of steroids: Mahato and Garai
lus fumigatus. Proceedings of the 16th International Congress of Biochemistry and Molecular Biology, New Delhi, India, Abstracts, Vol II. p. 150.
47. Yoshihama M, Tamura K, Miyata N, Nakakoshi M (1988). Manu- facture of 7[3-hydroxy-4-pregnene-3,20-dione derivatives from pregn-4-ene-3,20-dione with Acremonium. World Patent 8805, 782 (Snow Brand Milk Products Co. Ltd).
48. Madyastha KM, Joseph T (1994). Transformation of 16- dehydroprogesterone and 17a-hydroxyprogesterone by Mucor piriformis. Appl Microbial Biotechnol 41:170-177.
49. Dastidar P, Joseph T, Madyastha KM, Row TNG 11993). Structure of a steroid fungal metabolite. Acta Co'st Sect C. Cryst Structure Commun C49:273-275.
50. Nihira T, Okada Y, Yamada Y (1992), A new bile acid glycoside produced by Cunninghamella blakesleeana ST-22 from lithocholic acid. J Ferment Bioeng 74:19 l - 193.
5 l. Ambrus G, likoy E, Horvath G, Podauyi B, Bocskei Z, Gyurky S, Jekkel A (1992). Novel intermediates of microbial side-chain deg- radation of sitosterol. Tetrahedron Lett 33:5267-5268.
52. Mukherjee E, Banerjee S, Mahato SB (1993). Transformation of cholic acid by Arthrobacter simplex. Steroids 58:484-490.
53. Mahato SB, Mukherjee E, Banerjee S (1994). Advances in micro- bial biotechnology of bile acids. Biotech Adv 12: 357-391.
54. Mahato SB, Mazumder I, Luger P, Weber M (1994). Cholic acid induced N-methylhydantoin synthesis in microbial system. J hl- dian Chem Sac 71:401-408.
55. Kominek LA, Wolf JH, Steiert PS (1993). 1,2-dehydrogenation of steroidal 21-esters with Arthrobacter simplex, or, Bacterium cy- clooxidans. U.S. Patent 5, 225, 335 (Upjohn Co.I.
56. Zeng B, Chen S, Chen B (1993). Improvement of enzyme activity in microbial l ll3-hydroxylation of steroids. Zhongguo Yiyao Gongye Zazhi 24:529-532.
57. Undisz K, Groh H, Stopsach H, Hoerhold SC (1992). Bioconver- sion of steroids by Cochliobolus lunatus. II. 1113-Hydroxylation of 17c~,21-dihydroxypregna-l,4-diene-3,20-dione-17-acetate in de- pendence of the inducer structure. J Steroid Biochem Mol Biol 43:543-547.
58. Hanson JR, Nasir H (1993). The biotransformation of some ste- roids by Cephalosporium aphidicola. Phytochemistr 3, 33:831-834.
59. Shi J, Sun X, Zhen L, Niu J (1993). 1 la-Hydroxylation of andro- sta- 1,4-diene-3,17-diane (ADD) by Aspergillus ochraceus. Shang- hai Yike Daxue Xuebao 20:373-376.
60. Weber A, Kewnnecke M (1993). Preparation of 14a-hydroxy-4- androstene-3,17-diane by fermentation, German Patent 4,129,005 (Schering A, G).
61. Madyastha KM (1994). Preparatively usethl transformation of ste- roids and morphine alkaloids by Mucor piriformis. Proc Indian Acad Sci Chem Sci 106:1203-1212.
62. Arison BH, Carlin JR, Venkataramant ES (1993). Manufacture of testosterone-5a-reductase inhibitors with Selenustrum capricornu- rum. U.S. Patent 5, 215, 894 (Merck).
63. Santhanam HK, Shreke GS (1994). Solvent selection and produc- tivity in multiphase biotransformation systems. Biotechnol Prog 10:187-192.
64. Sedlaczek L, Milczarek K, Wilmanska D, Parniewski P, Lubisz K, Filip A, Majdziak K, Szczepaniak J, Korbel J (1993). Method for manufacturing hydrocortison from Reichstein's S substance. Po- land Patent 161, 090 (Pabianickie Zaklady Farmaceutyczne "'Polfa").
65. Wilzmanska D, Milczarek K, Rumijowksa A, Bartnicka K, Sed- laczek L (1992). Elimination of byproducts in 11 ~-hydroxylation of substance S using Curvularia lunata clon~s regenerated from NTG-treated protoplasts. Appl Microbial Bioteehnol 37:626-63(I.
66. Zeng B, Zhang B (1992). The relationship between the growing characterization and steroid l la-hydroxylation of Beauveria bassiana AS69 during fermentation. Weishengwuzue ~zhi 12:43- 45.
67. Voigt B, Porzel A, Naumann H, Hoerhold SC, Adam G (1993). Hydroxylation of the native brassinosteroids 24-epicastasterone and 24-epibrassinolide by the fungus Cunninghamella echinulata. Steroids 58:320-323.
68. Schmauder HP, Irrgang S, Schlosser D (1991). Steroid hydroxyl- ation with an immobilized filamentus fungus. Meded Fac Land- bouwwet, Rijksuniv Gent 56:1785 1787.
Steroids, 1997, vol. 62, April 343

Review
69. Schlosser D, Irrgang S, Schmauder HP (1992). Immobilization of Penicillium raistrickii spores and mycellium growth for 15a- hydroxylation of 13-ethylgon-4-ene-3,17-dione. Folia Microbial (Prague) 37:249-255.
70. Schlosser D, Schmauder HP (1992). Catalyst capacity factor in steroid biotransformation. Chem Eng Tech 64:357-3611.
71. Schlosser D, Irrgang S, Schmauder HP (1993). Steroid hydroxyl- ation with free and immobilized cells of Penicillium raistrickii in the presence of 13-cyclodextrin. Appl Microbiol Biotechnol 39:16- 20.
72. Schlosser D, Irrgang S, Schmauder HP, Dautzenberg H, Tomas- chewski G, Zimelka W, Baumgartl H (1992). 15a-Hydroxylation of 13-ethyl-gon-4-ene-3,17-dione with free and immobilized Pen# cillium raistrickii. Proceedings of the DECHEMA Biotechnol Conference, 231-234.
73. Madyastha KM, Joseph T (1993). Studies on the 14a-hydroxy- lation of progesterone in Mucor pir(['ormis. J Steroid Biochem Mol Biol 45:563-569.
74. Hamada H, Takiguchi K, Konishi H (1992). Manufacture of 613- hydroxytestosterone from testosterone by plant tissue culture of Marchantia. Japanese Patent 04, 139, 193 (JGC Corp.).
75. Farooq AH, James R, Iqbal Z (1994). Hydroxylation of progester- one by Cephalosporium aphidicola. Phytochemistry 37:723-726.
76. Kim MH, Kim MN (1991). Transformation pathway of the pro- gesterone by Rhizopus nigricans. Misaengmul Hakhoechi 29:111- 116.
77. Dutta TK, Sarnanta TB (1992). An improved method for separation and estimation of bioconverted steroids. J Gen Appl Microbiol 38:283-288.
78. Lee JJ, Kim MN (1993). Ion effect on steroid bioconversion in Rhizopus nigricans Hart 'guk kyunhakhoechi 21:23-27.
79. Vitas M, Smith K, Rozman D, Komel R (1994). Progesterone metabolism by the filamentous fungus Cochliobolus lunata. J Ste- roid Biochem Mol Biol 49:87-92.
80. Smith KE, Ahmed F, Williams RAD, Kelly SL (1994). Microbial transformations of steroids, VIII. Transformation of progesterone by whole cells and microsomes ofAspergillusfumigatus. J Steroid Biochem Mol Biol 49:93-100.
81. Chen KC, Yin Ws, Tiu C, Houng JY (1994). 1 la-Hydroxylation of progesterone using modified alginate immobilized cells. Enzyme Microbiol Teehnol 16:551-555.
82. Atta FM, El-kady IA (1994). Transformation of progesterone by Cochliobolus spicifer. Bull Fac Sci, Assiut Univ, B23:83-91.
83. Pollio A, Pinto G, Greca MD, De Mario A, Previtera L (1994). Progesterone bioconversion by microalgal cultures. Phytochemis- try 37:126%1272.
84. Madyastha KM, Shankar VN (1994). Role of neutral metabolites in microbial conversion of 313-acetoxy-19-hydroxycholest-5-ene into estrone. Appl Environ Microbiol 60:1500-1511.
85. Liu W, Lee CY (1992). Production of androst-4-ene-3,17-dione from cholesterol by Mycobacterium sp. in a synthetic medium. Zhongguo Nongye Hyaxue Huizhi 30:52-58.
86, Shi J, Chu Z, Ju Y, Mo G, Cheng W (1992). Degradation of cholesterol to androsta- 1,4-diene-3,17-dione by Rhodococcus cor- allina. Zhongguo Yiyao Gongye Zazhi 23:204-207.
87. Ahmed S, Roy PK, Basu SK (1993). Cholesterol side-chain cleav- age by immobilized cells of Rhodococcus equi DSM 89-133. In- dian J Exp Biol 31:319-322.
88, Smith M, Zahnley J, Pfeifer D, Goff D (1993). Growth and cho- lesterol oxidation by Mycobacterium species in Tween 80 medium. Appl Environ Microbiol 59:1425-1429.
89, Srivastava A, Patil S (1994). Investigation of some physicochem- ical parameters involved in the biotransformation of cholesterol to 17-ketosteroids. J Microb Biotechnol 9:101-112.
90. Ahmed S, Johri BN (1993). Microbial transformation of steroids in organic media. Indian J Chem (sect B) 32B:67-69.
91. Vorbrodt HM, Adam G, Porzel A, Hoerhold C, Daenhardt S, Boehme KH (1991). Microbial degradation of 2a,3a-dihydroxy- 5c~-cholestan-6-one by Mycobacterium vaccae. Steroids 56:586- 588.
92. Wang KC, Chiang N, Su CC, (1994). Microbial transformation of 5[3-steroids with Mycobacterium sp. (NRRLB-3805). Chin Pharm J (Taipei) 46:227-232.
93. Weber A, Kennecke M, Neef G (1992). 20-Methyl-5,7-pregna-
diene-313,2 l-diol derivatives and their manufacture with Mw:obac- terium. World Patent 0903, 465 (Schering A-G).
94. Dhar A, Samanta TB (1993). Novel oxidative cleavage of C~v-C2o bond in pregnane by a Pseudomonas sp. J Steroid Biochem Mol Biol 44:101-104.
95. Dutta RK, Roy M, Singh HD (1992). Metabolic blocks in the degradation of [3-sitosterol by a plasmid-cured strain of Arthro- batter oxydans. J Basic Microbiol 32:167-176.
96. Roy PK, Khan AW, Kumar J, Chopra SDK, Basu SK (1992). Steroid transformation in a laboratory-scale glass airlift fermentor. World J Microbiol Biotechnol 8:399-401.
97. Mathur S, Bhatia MC, Mathur SN (1992). Biotransformation of 13-sitosterol into 17-ketosteroids by some strains of Arthrobacter. Role Biotechnol Agric 99-102.
98. Kurakov VV, Korovova YN, Ryzhkova VM (1993). Effect of the initial concentration of 13-sitosterol on its microbiological transfor- mation to androst-4-ene-3,17-dione. Khim Farm Zh 27:65~57.
99. Shukla A, Patil S, Bharti S, (1992). Microbial conversion of sola- sodine to 4-androstene-3,17-dione (AD), a key intermediate for androgen synthesis. Lett Appl Microbiol 15:86-88.
100. Seidel L, Hoerhold C (1992). Selection and characterization of new microorganisms for the manufacture of 9-OH-AD from sterols. J Basic Microbiol 32:49-55.
101. Lee KU (1990). Conversion of plant sterol to androst-4-ene-3,17- dione by a mutant of Mycobacterium sp. NRRLB-3805. Misaeng- mul Hakhoechi 28:351-363.
102. Atrat PG, Koch B, Szekalla B, Hoerhold SC (1992). Application of newly synthesized detergents in the side-chain degradation of plant sterols by Myobacterium fortuitum. J Basic Microbio132:147-157.
103. Gottschaldt B, Grosse HH, Hoerhold C, Welzker M, Naumann H, Heller I, Birke M, Plonka G (1993). German Patent 301, 740 (Institute ftir Microbiologie and Experimentelle Therapie; Je- napharm GmbH JENA).
104. Oda S (1994). Manufacture of 1,4-androstadiene-3,17-dione with immobilized microorganism. Japanese Patent 0600, 095 (Kansai Paint Co. Ltd).
105. Zhang LQ, Bian EP, Wang Y (1992). Side-chain cleavage of ste- rols by h{vcobacterium sp. MI2. Yaox3'e Xuebao 27:903-907.
106. Weber A, Kenneck M (1992). Manufacture of 4-pregnene-3,20- dione and its derivative by microbial transformation of pregnanes. World Patent 9203, 571 (Schering A-G).
107. Pinheiro HM, Cabral JMS, Adlercreutz P (1993). Quinones as external electron acceptors in steroid ALdehydrogenation with en- trapped cells in organic medium. Biocatalysis 7:83-96.
108. Wang KC, Chaing N, Lee SS (1993). Microbial transformation of 5-c~-androstan-3~-ol-17-one with Mycobacterium sp. Zhonghua Yaoxue Zazhi 45:215-221.
109. Kimura H, Okamura A, Kawaide H, Yamaura T (1992). Oxidation of bile acids with Bacillus. European Patent 518, 661 (Tokyo Tan- abe Co. Ltd).
110. Kimura H, Okamura A, Kawide H (1994). Oxidation of 3,7 and 12-hydroxyl groups of cholic acid by an alkalophilic Bacillus sp. Biosci Biotechnol Biochem 58:1002-1006.
111. Panchishina MV (1992). Transformation of sterols by Escherichia. Zh Mikrobiol Epidemiol lmmunobiol, 8-11.
112. Osipowicz 13, Krezel Z, Siewinski A (1992). Biotransformation XXXIII. Oxidation of 313- and 17c~-hydroxysteroids by Nocardia rubra cells in heptane-water system. J Basic Microbiol 32:215- 216.
113. Liu WH, Tsai MS, Lee CY (1994). Bioconversion of cholesterol to cholest-4-ene-3-one using free and immobilized growing cells of Arthrobacter simplex. Zhongguo Huaxue Huizhi 32:190-198.
114. Myamato I, Toyoda K (1994). Cholestenone manufacture with cholesterol oxidase of Rhodococcus. Japanese Patent 06, 157, 585 (Nippon Oils and Fats Co. Ltd).
115. Kimura H, Okamura A, Kawaide H (1993). Bacillus for manufac- ture of 3c~,7e~-dihydroxy-12-keto-513-cholanic acid. European Pat- ent 539, 216 (Tokyo Tanabe Co. Ltd).
116, Weber A, Kennecke M, Klages U, Nickisch K, Rohde R (1992). Preparation of 17-oxosteroids by microbial oxidation of 1713- hydroxysteroids. World Patent 9203, 572 (Schering A-G).
117, Sallam LA, EI-Abyad MS, EI-Refai AMH, El-Menofi HA, Adham NZ (1995). Bioconversion of 19-nortestosterone by Rhodococcus
344 Steroids, 1997, vol. 62, April

sp. DSM 92-344. I: Optimization of transformation parameters. Process Biochem (Oxford) 30:25-34.
118. Grigorova P, Krysteva M, Pendulovska V, Bashinovsky S, Mitova J, Krysteva S (1992). Activity of immobilized cells from Arthro- bacter simplex included in polymer matrix. Tr Nauchnoizsled KhimikoCarm lnst 18:94-102.
119. Abou EI-Hawa M, Mahfouz W, Taha O, Sallam LAR (1993). AI-Dehydrogenation of cortisol with bacteria II. Some aspects of the enzymic-dehydrogenation of cortisol with Co~nebacterium equi. Egypt J Microbiol 28:215-222.
120. Abou-E1-Hawa M, Mahfouz W, Taha O, Sallam LAR (1993). A I- Dehydrogenation of cortisol with bacteria 1-screening experiments and some physiological studies. Egypt J Microbiol 28:119-130.
121. Pinheiro HM, Cabral JMS (1992). Activity and stability of an entrapped-cell system for the ALdehydrogenation of steroids in organic media. Biotechnol Bioeng 40:1123-1127.
122. Pinheiro HM, Cabral JMS (1992). Microenvironmental effects on steroid ,~-dehydrogenation in organic media using immobilized whole cells. Prog Biotechnol 8:129-136.
123. Kimura H, Okamura A (1992). Manufacture of 3-keto-7[3- hydroxycholanic acid with Bacillus. Japanese Patent 04, 278, 096 (Tokyo Tanabe Co. Ltd).
124. Hamada H, Naka S, Kurban H (1993). Stereoselective reduction in the biotransformation of androstane derivatives by cell suspension cultures of Marehantia polymorpha. Chem Lett :2111-2112.
125. Mazumder I, Mahato SB (1993). Bile acid transformation by Al- caligens recti. Steroids 58:78-86.
Biotransformation of steroids: Mahato and Garai
126. Fa Y, Su Q (1992). Microbial production of 3-oxo-bisnorchola- 1,4-diene-22-oic acid from cholesterol by an Arthrobacter 82. Weishengwu Xuebao 32:17-22.
127. Sugiura T, Tsuji M, Fumino M, lchihara Y (1992). Manufacture of 3a,12a-dihydroxy-7-keto-513-bisnorcholanic acid (salts) with Co- rynebacterium. Japanese Patent 04, 211,386 (Kuraray Co. Ltd).
128. Mukherjee E, Banerjee S, Mahato SB (1993). Microbial transfor- mation of deoxycholic acid and lithocholic acid by Arthrobacter simplex. J Chem Res (suppl): 416-417.
129. Kreis W, Reinhard E (1992). 1213-Hydroxylation of digitoxin by suspension-cultured Digitalis lunata cells. Production of digoxin in 20 lit and 300-1itre air-lift bioreactors. J Biotechnol 26:257-273.
130. El Olemy MM, Elhag H, El Domiaty M, Sattar EA, AI Azizi MM, AI Said MS (1994). Biotransformation of digitoxigenin by cell suspension cultures of Pergularia tomentosa and Ammi visnaga. Saudi Pharm J 2:76-83.
131. Choudhury S, Halos S, Krischenowski D, Schmeda-Hirschmann G (1993). Two new 17a-hydroxy progesterone transformation prod- ucts from Nocardia DSM 43298. World J Microbiol Biotechnol 9:56-58.
132. Weber A, Kennecke M, Vidic HJ (1993). Manufacture of 17c~- hydroxy-3-methoxy-8,14-seco-l,3,5 (10),9(11) estratetraene-14- one by reduction in aqueous suspension with KIoeckera magma. World Patent 9320, 222 (Schering A- G).
133. lonkova I (1991). Biotransformation of 13-methyldigitoxin by sur- face cultures of Digitalis purpurea L. in vitro. Probl Farmakol Farm 5:3843.
Steroids, 1997, vol. 62, April 345