adhesion bacteriana
TRANSCRIPT

EXPLOTACIÓN DE LA ADHESIÓN BACTERIANA
PATOGÉNESIS
Autora: A. Vanesa Macías CañoSeminario: Mecanismos de Patogeneidad bacteriana
Supervisión: Dra. Dña. Carmen AmaroCurso: 2010 - 2011
Fecha: 22/12/101

inicio
• INTRODUCCIÓN
• IMPORTANCIA DE LA ADHERENCIA
• MOLÉCULAS DE ADHESIÓN
• ADHESINAS BACTERIANAS
• COLONIZACIÓN E INVASIÓN DEL HUÉSPED
• YERSINIA
• ADHESIÓN
• RESULTADO
• PESTE NEGRA
• LYSTERIA MONOCYTOGENES
• ADHESIÓN
• RESULTADOS
• NEISSERIA GONORROEHEAE
• ADHESIÓN
• RESULTADOS
• HELCOBACTER PYLORI
• ADHESIÓN
• RESULTADOS
2

Importancia de la adherencia
3

MOLÉCULAS DE ADHESIÓN
4

5

Adhesinas bacterianas
6
Las adhesinas fimbriales son parte constitutiva de una fimbria y las moléculas encargadasde asegurar la adhesión de esa estructura a su receptor en la célula hospedera.

Colonización e invasión del huésped
7

Integrina y yersinia
• Género de bacterias que pertenece a la familia de las Enterobacteriaceae.
• Patógenos de animales, de donde pasan al ser humano produciendo enfermedades.
• Bácilos del tipo gramnegativos aerobios y anaerobios facultativos;
• mótiles a 22°C, pero no a 37°C, por flagelos anfitricos, o peritricos,
• forman pilis, y fimbrias.
• No forman cápsulas de gran espesor ni esporas.
• Las especies son: Yersinia pestis, Yersinia enterocolitica y Yersinia pseudotuberculosis.
8

9

ADHESIÓN DE YERSINIA
10

RESULTADO DE YERSINIA
11

CURIOSIDAD DE LA PESTE NEGRA
12

CADHERINAS Y LISTERIA MONOCYTOGENES
13
• Bacteria intracelular facultativa
• Causa la Listeriosis
• Infecciones alimentarias más virulentas, con una tasa de mortalidad entre un 20 a 30%,.
• Bacilo Gram positivo (Joseph Lister)
• Es pequeño, no ramificado y anaerobio facultativo
• capaz de proliferar en un amplio rango de temperaturas (1°C a 45°C) y una elevada concentración de sal.
• Catalasa positivo y no presenta cápsula ni espora.
• Tiene flagelos perítricos, gracias a los cuales presenta movilidad a 30ºC o menos, pero es inmóvil a 37 ºC,

Adhesión de LISTERIA MONOCYTOGENES
14
La internalina bacteriana se une a la cadherina . Α y β cadherina actúan como adaptadores entre la cola citoplásmica de la E-cadherina y el citoesqueleto de actina. Este vínculo con el citoesqueleto de actina se requiere para la invasión de L. monocytogenes
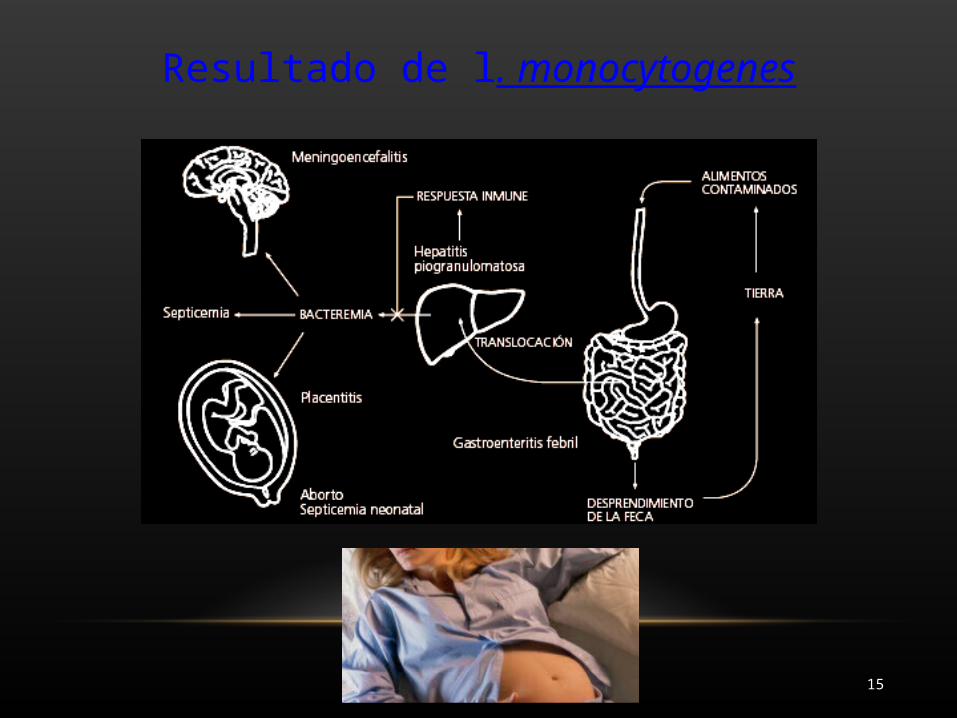
Resultado de l. monocytogenes
15

Ig - CAMs y Neisseria gonorrhoeae
16
• Diplococo Gram negativo, oxidasa positivo,
• Causa la gonococia
• Se diferencia de otros tipos de Neisseria ya que fermenta solo a la glucosa.
• Se caracteriza por ser de difícil cultivo, siendo muy exigente a nivel nutricional y a la vez muy sensible a sustancias que se encuentran en los medios de cultivo corrientes.
• Requiere una atmósfera con 5-10% de CO2.

17
Vías de señalización (a) mediada por la invasión CEACAM3 por N. gonorrhoeae y (b) la fagocitosis mediada por receptores Fc.
Quinasas de la familia Src (Hck y FGR en [a] y Lyn y Hck en [b]) fosforilan residuos de tirosina en el ITAM motivo el receptor. Syk tirosina quinasa es reclutados para el motivo del ITAM fosforilados, donde se activa. Syk después transmite una señal descendente que conduce a la activación de Rac y Cdc42, lo que podría activar la N-WASP, dando lugar a la polimerización de actina a través de la Arp2 / 3 complejos.
18
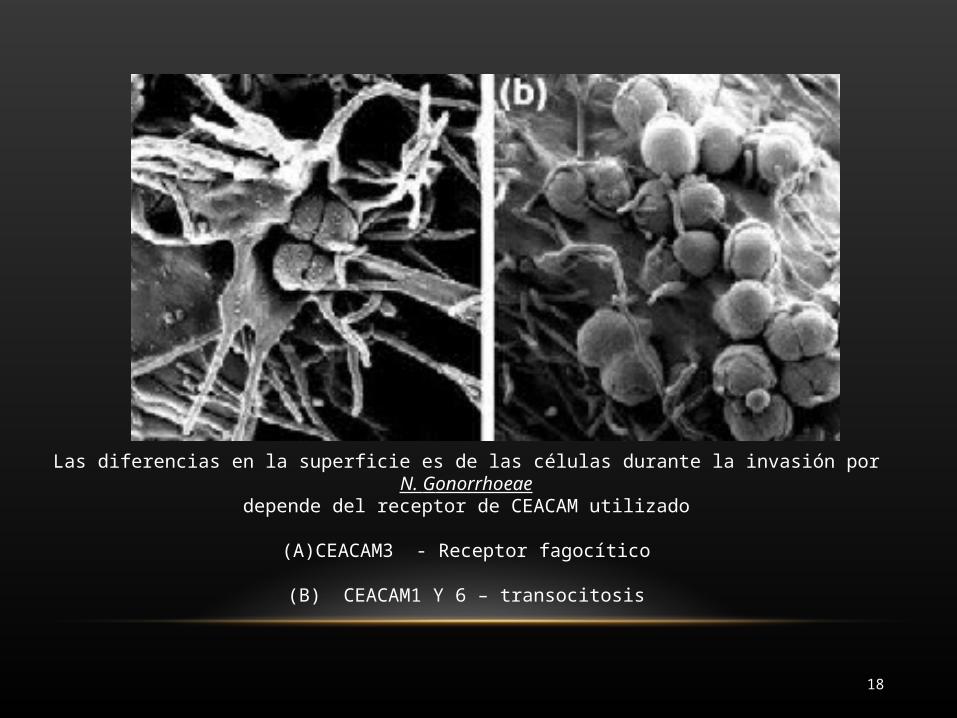
Las diferencias en la superficie es de las células durante la invasión por N. Gonorrhoeaedepende del receptor de CEACAM utilizado
(A)CEACAM3 - Receptor fagocítico
(B) CEACAM1 Y 6 – transocitosis

Resultados de N. gonorroehae
19

Selectina y helicobacter pylori
20
• Gram negativa de forma espiral,
• Tiene unos 4–6 flagelos.
• Microaerófila, es decir, requiere oxígeno pero a más bajas concentraciones de las encontradas en la atmósfera.
• Usa hidrógeno y metanogénesis como fuente de energía.
• Además es oxidasa y catalasa positiva.
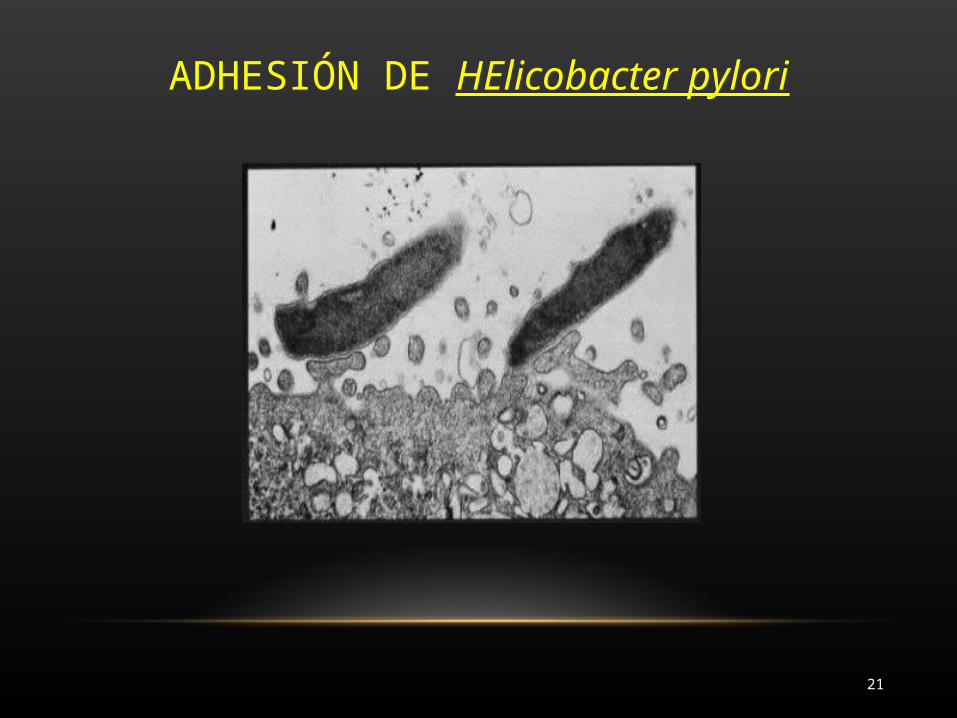
ADHESIÓN DE HElicobacter pylori
21

22

RESULTADOS DE HElicobacter pylori
23
Infección por Infección por H. pyloriH. pylori Infección por Infección por H. pyloriH. pylori
GastritisGastritisSuperficialSuperficial
AntritisAntritisDifusaDifusa
PangastritisPangastritis(G. atrófica (G. atrófica multifocal)multifocal)
Ulcera DuodenalUlcera DuodenalUlcera GástricaUlcera GástricaCáncer Gástrico Cáncer Gástrico
AsintomáticosAsintomáticos
BIBLIOGRAFÍA
• Christiaan van OoBacterial pathogenesis: Opening the gates of type III secretion. Nat Rev Microbiol 8, Jun 2010;389-389 doi:10.1038/nrmicro2378
• Eisenreich W, Dandekar T, Heesemann J, Goebel W. Carbon metabolism of intracellular bacterial pathogens and possible links to virulence. Nat Rev Microbiol, Jun 2010;8:401-412 | doi:10.1038/nrmicro2351Xiu-Jun Yu, Kieran McGourty, Mei Liu, Kate E. Unsworth, David W. Holden. pH Sensing by Intracellular Salmonella Induces Effector Translocation. Science. 21 May 2010;328:1040-1043.
• Flannagan RS, Cosío G, Grinstein S. Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat Rev Microbiol, May 2009;7:355-366 doi:10.1038/nrmicro2128
• Juhas M, van der Meer JR, Gaillard M, Harding RM, Hood DW, Crook DW. Genomic islands: tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol Rev. 2009 Mar;33(2):376-93.
• Shames SR, Auweter SD, Finlay BB. Review. Co-evolution and exploitation of host cell signaling pathways by bacterial pathogens. Int J Biochem Cell B, Feb 2009; 41(2):380-389. doi:10.1016/j.biocel.2008.08.013
• Diacovich L, Gorvel J-P. Bacterial manipulation of innate immunity to promote infection. Nat Rev Microbiol, Feb 2010;8:117-128 doi:10.1038/nrmicro2295
• Spallek T, Robatzek S, Göhre V. How microbes utilize host ubiquitination. Cell Microbiol, 2009;11(10):1425-1434. DOI:10.1111/j.1462-5822.2009.01346.x
• The Mechanisms of Bacterial Pathogenicity. Todar's Online Textbook of Bacteriology, 2009.
• Lambris JD, Ricklin D, Geisbrecht BV. Complement evasion by human pathogens. Nat Rev Microbiol, 2008;6 (2):132-142. doi:10.1038/nrmicro1824
• Wu H-J, Wang AH-J, Jennings MP. Discovery of virulence factors of pathogenic bacteria. Curr Opin Chem Biol 2008 ;12 (1):93-101. doi:10.1016/j.cbpa.2008.01.023
• Fraser JD, Proft T. The bacterial superantigen and superantigen-like proteins. Immunol Rev, 2008; 225(1):226 – 243.
• Munford RS. Sensing gram-negative bacterial lipopolysaccharides: A human disease determinant? Infect Immun, 2008;76(2):454-465.
• Zipfel PF, Würzner R, Skerk C. Review. Complement evasion of pathogens: Common strategies are shared by diverse organisms. Mol Immunol, Sept 2007; 44(16):3850-3857. XIth European meeting on Complement in Human Disease. doi:10.1016/j.molimm.2007.06.149 doi:10.1016/j.molimm.2007.06.149
• Pallen, M.J., Wren, B.W. Bacterial pathogenomics. Nature 2007;449(7164):835-842. doi:10.1038/nature06248
• Schaible UE, Kaufmann SHE. Iron and microbial infection. Nat Rev Microbiol, Dec 2004;2:946-953 doi:10.1038/nrmicro1046
• Hathaway LJ, Kraehenbuhl JP. The role of M cells in mucosal immunity. Cell Mol Life Sci. 2000 Feb;57(2):323-32. DOI: 10.1007/PL00000693 24

GRACIAS POR SU ATENCIÓN
25