adherence of vibrio parahaemolyticus to rabbit intestinal epithelial cells in vitro
TRANSCRIPT

FEMS Microbiology Letters 84 ( 1991) 113-1 l 8 © 1991 Federation of European Microbiological Societies 0378-1097/91/$03.50 Published by Elsevier
113
FEMSLE 04675
Adherence of Vibrio parahaemolyticus to rabbit intestinal epithelial cells in vitro
M.K. Chakrabarti , A.K. Sinha and T. Biswas
National Institute of Cholera and Enteric Diseases, Calcutta, India
Received 11 June 1991 Revision received 12 August 1991
Accepted 16 August 1991
Key words: Vibrio parahaemolyticus; Kanagawa phenomenon; Adhesion; Hemolysin; Cell surface hydrophobicity; Rabbit intestinal epithelial cell
SUMMARY
Kanagawa positive strains of Vibrio para- haemolyticus showed adherence whereas most of the Kanagawa negative strains were non-adhering to rabbit intestinal epithelial cells. Anti-hemo- lysin antisera did not inhibit the adherence of V. parahaemolyticus strains. Moreover, the adhesive capacity of non-adhering strains was not en- hanced by purified hemolysin. Cell surface hy- drophobicity remained the same in both Kana- gawa positive and negative strains of V. para- haemolyticus. Fetuin strongly inhibited the adher- ence to rabbit intestinal epithelial cells followed by -mannose and D-glucose.
Correspondence to: M.K. Chakrabarti, Division of Pathophysi- ology, National Institute of Cholera and Enteric Diseases, P-33, CIT Road, Scheme XM, Beliaghata, Calcutta, 700010, India.
2. INTRODUCTION
Vibrio parahaemolyticus are divided into two groups on the basis of their ability to hemolyse a high-salt blood agar medium (Kanagawa phe- nomenon) [1]. Hemolytic strains, known as Kana- gawa positive (KP+), are generally isolated from patients whereas Kanagawa negative (KP-) strains are generally environmental [2]. Kanagawa hemolysin (thermostable direct hemolysin) may be one of the virulence factors of V. para- haemolyticus [3].
Bacterial adherence and proliferation within the intestine are prerequisites for virulence. Sev- eral investigators have sought evidence for the relationship between Kanagawa phenomenon of V. parahaemolyticus and their ability to adhere to isolated cells. Carruthers et al. [4] found a corre- lation between adhesion to HeLa cells and pro- duction of the Kanagawa hemolysin whereas other workers found no significant difference in the adhesive capacity of KP + and KP- strains [2,5,6].

114
In the present communication we report a differ- ence in adherence properties of KP + and KP strains, isolated from different sources in Cal- cutta, to rabbit intestinal epithelial cells (RIEC).
3. M A T E R I A L S A N D M E T H O D S
3.1. Bacterial strains Twenty s t r a i n s o f V. parahaemolyticus were
used in this study. Ten were KP + and were isolated from diarrhoeal patients admitted to the Pediatric and General Ward of the Infectious Diseases Hospital, Calcutta. The remander were K P - and were environmental isolates.
3.2. Preparation of rabbit intestinal epithelial cell (RIEC)
R I E C was prepared by the method of Dean and Isaacson [7]. Rabbits weighing 1.2-1.5 kg were killed and the small intestine was removed, rinsed with cold 150 mM PBS (pH 7.2) and kept in PBS for 30 min at 4 ° C. The intestine was then opened and the epithelium removed by scraping. Epithelial cells were collected by centrifugation at 200 × g for 15 min, washed in PBS and resus- pended in PBS to a concentration of 1 × 1(t ~ cel ls /ml. The number of epithelial cell was counted with a heamocytometer .
3.3. Adherence assay The adhesive capacity of radiolabelled bacteria
was measured according to the method of Sugar- man and Donta [8]. Bacteria were grown in Brain Hear t Infusion broth (Difco) supplemented with 1.5% NaCI containing 1 /xCi [laC]-leucine for 18 h at 37 °C with constant shaking. 200 /xl of pre- washed bacterial suspension (1 × 10 m C F U / m l ) in PBS was incubated with an equal volume of RIEC (1 × 108/ml) in a shaking waterbath for 1 h at 37 ° C. Mixtures were then filtered through a membrane filter (pore size 8 p.m, Millipore, U.S.A.) fitted in a disc-filter holder attached to a glass syringe. Non-adherent bacteria were re- moved by washing the filter with 30 ml of PBS through the syringe. Filters were then dried, placed in scintillation fluid and counted in a
Beckman liquid scintillation counter. The results were expressed as disintegration per minute (dpm).
3.4. Preparation of anti-hemolysin antiserum Antiserum against purified hemolysin (Sigma,
U.S.A.) was prepared by injecting 5 #g hemolysin with Freund's complete adjuvant (Difco, USA) intramuscularly into the rabbits. Subsequently, booster doses of 20 # g hemolysin with Freund's incomplete adjuvant were injected at 2 week in- tervals for a period of 2 months. The antiserum was collected, inactivated by heating at 56 ° C for 30 rain and stored at - 7 0 o C.
3.5. Neutralisation of" hemolysin by anti-hemolysin antisera
Antisera (0.25 ml) was serially diluted two-fold with an equal volume of 0.9% NaCl containing 5.0 mM CaCI= and 10.0 mM Tris buffer (pH 7.0). Hemolys in solution (0.5 ml containing 2 haemolytic units) in buffered saline was added to
3 ° each tube and incubated at .7 C for 1 h. The remaining hemolytic activities were then deter- mined by adding 1 ml of 1% defibrinated rabbit erythrocytes suspended in buffered saline to each tube of the above series [9].
The highest dilution at which complete inhibi- tion of hemolysis occured was taken as the anti- body titre and this was found to be 1 • 128.
To determine the effect of antihemolysin anti- sera on bacterial adherence to the epithelial cell, 0.2 ml of antiserum or 0.2 ml of diluted antiserum (1 in 100) was mixed with RIEC. An equal vol- ume of a suspension of radiolabelled bacteria (1 × 1() 1~ C F U / m l ) was added and the adherence assays were performed as described above.
To determine the effect of hemolysin on the adherence, 0.2 ml of RIEC was incubated with 0.4 ml hemolysin (5.0 /xg/ml) for 15 min at 37°C. Cells were then washed with PBS and used for adherence assays.
3.6. Salt aggregation test Cell surface hydrophobicity of all these strains
was tested using the salt aggregation test (10). One drop of a bacterial suspension (5 x 109 bac- teria per ml in 0.02 M sodium phosphate buffer, pH 6.8) and one drop of each ammonium sulfate

solut ion was added and the bac ter ia mixed on
glass slides. Bacter ia l aggregat ions were read vi-
sually against a whi te background.
3. 7. The effect of monosaccharides and glycopro- tein
T h e effect of d i f ferent monosaccha r ides and
glycoprote in was d e t e r m i n e d by t rea t ing the bac-
ter ia with 2 0 0 / z l of each test c o m p o u n d at 37 ° C
for 1 h unde r constant shaking. The t rea ted cells
were then washed and r e suspended in PBS for
a d h e r e n c e assays.
T h e fol lowing test compounds were used per
200 /zl: o - m a n n o s e 200 /xg, o-g lucose 200 /xg,
L-fucose 200 /xg , N-ace ty l -o -g lucosamine 200 #g ,
N-ace ty l -o -ga lac tosamine 200 /zg, Fe tu in 25-200 /zg.
To d e t e r m i n e the background bacter ia l adhe-
sion to the f i l ter alone, which might result f rom
aggrega t ion in the p resence or absence of test
compounds , controls wi thout R I E C were always
taken. Background bacter ia l adhes ion did not ex-
ceed 0.02% adhesion. R I E C were examined un-
der mic roscope to d e t e r m i n e whe the r morpho log-
ical abnormal i t i es occur red due to t r ea tment . Re-
sults were r eco rded only when the control R I E C
rema ined unaffec ted .
4. R E S U L T S
4.1. Adhesive capacity of different strains The adhesive capaci ty of d i f ferent KP + and
K P - strains were compared . All the tes ted KP ÷
strains showed adherence , the pe r cen t age of ad-
he rence varying f rom 22.0-38.8 (Table 1). How-
ever, most of the K P - strains did not adhe re to
the R I E C and out of 10 strains only 2 exhibi ted
adhe rence which r e m a i n e d 0 .4 -10 .5% and was
much lower than that of KP + strains (Table 1).
4.2. Effect of treatment of RIEC with hemolysin and antihemolysin antisera
P r e - t r e a t m e n t o f R I E C with hemolysin had no
effect on the adhes ion o f V. parahaemolyticus strains (Table 2). Add i t i on o f ant ihemolys in antis-
era to the incubat ion mixture could not inhibit
adhesive capaci t ies of t es ted strains (Table 3).
115
Table 1
Adhesive capacity of different strains of V. parahaemolyticus to rabbit intestinal epithelial cell (Mean_+ SD, n = 6)
Strain Source Kanagawa Sero~pe % Adhesion phenom- enon
N10 Clinical + 05 :K15 38.5 -+2.5 N17 Clinical + 04:K55 32.1 _+ 1.2 N22 Clinical + 02:K3 28.4 _+1.1 N23 Clinical + 03 :K58 22.0 _+3.2 N24 Clinical + 05 : K15 35.8 _+ 2.2 N37 Clinical + 05:K17 35.0 _+2.2 N38 Clinical + 04:K55 23.8 _+2.1 N108 Clinical + 05 :K68 26.4 + 1.9 Nll0 Clinical + 05 : K68 31.0 -+l.5 Nl12 Clinical + 03:K58 29.0 -+1.9 N40 Environment - 04:K42 0.4 -+0.1 N42 Environment - 07:K19 2.2 _+ 1.0 N43 Environment - 03:K30 1.5 +0.4 N44 Environment - 04:K42 8.0 -+ 1.2 N72 Environment - 05:K17 0.8 +0.4 N76 Environment - 04 : K42 0.42-+ 0.22 N82 Environment - 04 : K42 0.68-+ 0.14 N93 Environment - 04:K42 10.5 -+ 1.6 N97 Environment - 03:K31 0.5 +_0.2 N124 Environment - 06:K46 1.0 _+0.3
% Adhesion = Counts (DPM) of bacteria to epithelial cell
Total counts added to the mixture x 100.
P r e - t r e a tm en t o f non -adhe ren t bacter ia with
hemolysin did not s t imulate thei r adhesive capac-
ity (results not shown).
Table 2
Effect of pre-treatment of rabbit intestinal epithelial cell with hemolysin on the adhesion of V. parahaemolyticus strains (Mean -+ SD, n = 6)
Strain Kanagawa % Relative phenomenon adhesion
N10 + 99_+5.8 (1) N17 + 96_+4.6 (4) N22 + 102 _+ 4.0 (2) N24 + 92 + 4.5 (8)
% Relative adhesion = % adhesion of treated bacteria
× 100. % adhesion of untreated bacteria
Figures in the parenthesis indicate percent inhibition or stim- ulation of adhesion.
116
Table 3
Effect of antihemolysin antisera on the adhesion of V.. para- haemolyticus strains to isolated RIEC (Mean_+ SD, n = 6)
Strain Kanagawa % Relative adhesion phenomenon
N10 + 101 _+ 4.4 (1) N17 + 98 _+ 5.2 (2) N22 + 95 _+ 3.8 (5) N24 + 102 _+ 6.0 (2)
% Relative adhesion =-
% adhesion of treated bacteria x 100.
% adhesion of untreated bacteria
Figures in the parenthesis indicate percent inhibition or stim- ulation of adhesion.
4.3. Cell surface hydrophobicity of different stra#ts All the KP + and KP strains of V. para-
haemolyticus tested aggregated at ammonium sul- fate concentrations of 1.8-2.5 M, which indicated that the strains were not highly hydrophobic. Surface hydrophobicity was similar for both KP + and K P - strains and was not correlated with adherence propert ies of these strains.
4.4. Effect of pre-treatment of bacteria with monosaccharides and glycoprotein
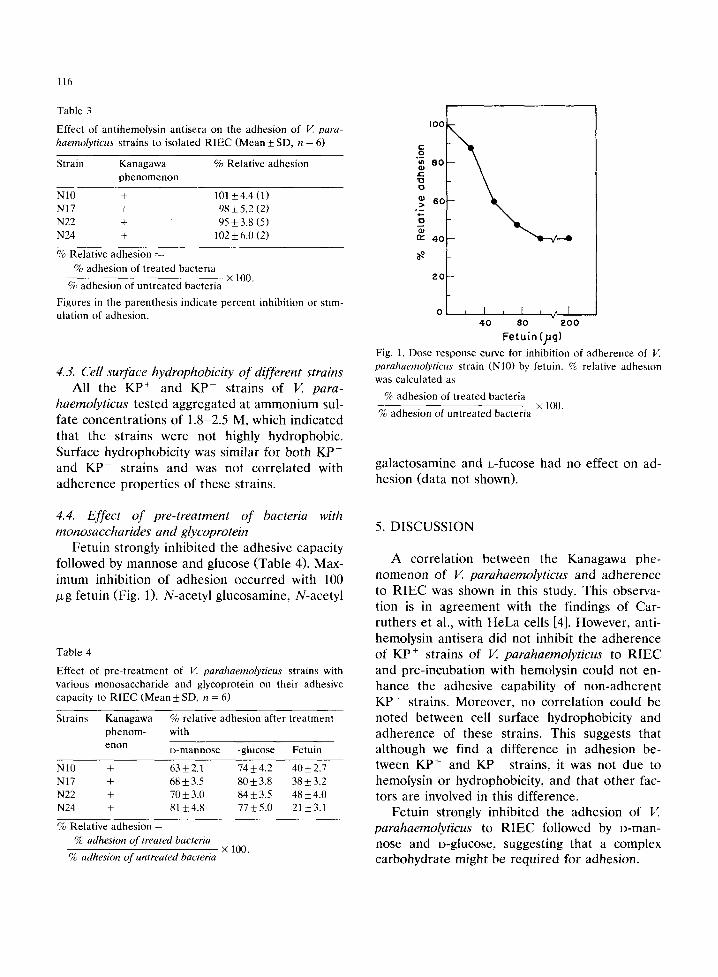
Fetuin strongly inhibited the adhesive capacity followed by mannose and glucose (Table 4). Max- imum inhibition of adhesion occurred with 100 /xg fetuin (Fig. 1). N-acetyl glucosamine, N-acetyl
Table 4
Effect of pre-treatment of V.. parahaemolyticus strains with various monosaccharide and glycoprotein on their adhesive capacity to RIEC (Mean_+ SD, n = 6)
Strains Kanagawa % relative adhesion after treatment phenom- with e r l o n D-mannose -glucose Fetuin
N10 + 63+2.1 74_+4.2 40_+2.7 NI7 + 68_+3.5 80_+3.8 38_+3.2 N22 + 70_+3.0 84_+3.5 48_+4.0 N24 + 81_+4.8 77_+5.0 21-+3.1
% Relative adhesion =
% adhesion of treated bacteria
I00
t - o
8 0 .c "EJ o
2 0
i I l [ r ~ l 40 80 2 0 0
Fetuin (jig) Fig. 1. Dose response curve for inhibition of adherence of V. parahaemolyticus strain (NIO) by fetuin. % relative adhesion was calculated as
% adhesion of treated bacteria × 100,
% adhesion of untreated bacteria
galactosamine and L-fucose had no effect on ad- hesion (data not shown).
5. DISCUSSION
A correlation between the Kanagawa phe- nomenon of V. parahaemolyticus and adherence to R I E C was shown in this study. This observa- tion is in agreement with the findings of Car- ruthers et al., with HeLa cells [4]. However, anti- hemolysin antisera did not inhibit the adherence of KP + strains of V. parahaemolyticus to R I E C and pre-incubation with hemolysin could not en- hance the adhesive capability of non-adherent K P - strains. Moreover, no correlation could be noted between cell surface hydrophobicity and adherence of these strains. This suggests that although we find a difference in adhesion be- tween KP + and K P - strains, it was not due to hemolysin or hydrophobicity, and that other fac- tors are involved in this difference.
Fetuin strongly inhibited the adhesion of V. parahaemolyticus to R I E C followed by D-man- nose and o-glucose, suggesting that a complex carbohydrate might be required for adhesion.
× 100. % adhesion o f untreated bacteria

ACKNOWLEDGEMENTS
We thank Dr. S.C. Pal, Director of this insti- tute, for his interest and encouragement through- out these studies. The secretarial assistance of Mr. S. Karmakar is thankfully acknowledged.
REFERENCES
[1] Wagatsuma, S. (1968) Media Circle 13, 159-161. [2] Reyes, A.L., Crawford, R.G., Spaulding, P.L., Peeler,
J.T. and Twedt, R.M. (1983) Infect. Immun. 39, 721-725. [3] Shirai, H., lto, H., Hirayama, T., Nakamoto, Y.
117
Nakabayashi, N., Kumagai, K., Takeda, Y. and Nishibuchi, M. (1990) Infect. lmmun. 58, 3568-3573.
[4] Carruthers, M.M. (1977) J. Infect. Dis. 136, 588-592. [5] Ijima, Y., Yamada, H. and Sinoda, S. (1981) Can. J.
Microbiol. 27, 1252-1259. [6] Gingras, S.P. and Howard, L.V. (1980) Appl. Environ.
Microbiol. 39, 369-371. [7l Dean, E.A. and Isaacson, R.E. (1982) Infect. lmmun. 36,
1192-1198. [8] Sugarman, B. and Donta, S.T. (1979) J. Med. Microbiol.
12, 373-378. [9] Cherwonogrodzky, J.W. and Clark, A.G. (1982) FEMS
Microbiol Lett., 15, 175-179. [10] Rozgonyi, F., Szitha, K.R., Ljungh, A., Baloda, S.B.,
Hjerten, S. and Wadstrom, T. (1985) FEMS Microbiol. Lett. 30, 131-138.