adaptation strategies in wetland plants: links between ecology and physiology. proceedings of a...
TRANSCRIPT

A Role for Phytotoxins in the Phragmites Die-Back Syndrome?Author(s): Jean Armstrong, William Armstrong, Zenbin Wu and Fawzia Afreen-ZobayedSource: Folia Geobotanica & Phytotaxonomica, Vol. 31, No. 1, Adaptation Strategies in WetlandPlants: Links between Ecology and Physiology. Proceedings of a Workshop (1996), pp. 127-142+III-VIPublished by: SpringerStable URL: http://www.jstor.org/stable/4181424 .
Accessed: 15/06/2014 16:58
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Springer is collaborating with JSTOR to digitize, preserve and extend access to Folia Geobotanica&Phytotaxonomica.
http://www.jstor.org
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

Folia Geobot. Phytotax. 31: 127-142, 1996
A ROLE FOR PHYTOTOXINS IN THE PHRAGMITES DIE-BACK SYNDROME ?
Jean Armstrong1), William Armstrong 1), Zenbin Wu2) & Fawzia Afreen-Zobayed')
1) Department of Applied Biology, University of Hull, Hull, HU6 7RX, U.K.; tel. +44 1482 465527, fax +44 1482 465458
2) Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China
Keywords: Aeration, Callus, Lignification, Roots, Senescence, Sulphide
Abstract: Die-back and healthy stands of Phragmites australis (CAV.) TRIN. ex STEUD., in the U.K. and Hungary, were compared in terms of plant morphology and anatomy, sediment redox potential and sulphide levels and plant resistance to internal Poiseuille gas flow. In laboratory experiments rhizome cuttings were exposed to acetic acid or dissolved sulphide in unstirred solution cultures in order to determine whether the die-back symptoms found in the field could be induced by these phytotoxins. Most of the die-back symptoms, namely stunting of adventitious roots and laterals, bud death, callus blockages of the gas-pathways, and vascular blockages (both xylem and phloem), were produced by each of the phytotoxin treatments. These symptoms were largely absent from healthy field sites and from the experimental controls.
In a greenhouse experiment, plants were grown in waterlogged sand or loam, with or without a sub-surface organic layer composed of chopped-up rhizomes and roots mixed with the soil base. Especially during the first 70 days, redox levels were considerably lowered, and shoot numbers and shoot growth much reduced by the presence of the organic layers; the effects were most pronounced in the sand plus organic matter treatment.
It is suggested that accumulated phytotoxins, e.g. organic acids and/or sulphide, whether produced from the death and decay of the plant, or from excessive organic loading or as an indirect result of eutrophication, will perpetuate the die-back of Phragmites and prevent the recovery of the plant in the short term.
INTRODUCTION
Direct destruction, mechanical damage, grazing, water and sediment quality, lake level regulation and related effects (OSTENDORP 1989) and eutrophication (e.g. BOAR et al. 1989, DEN HARTOG et al. 1989, 6I2KOVA-KONCALOVA et al. 1-992, KOHL & KOHL 1993) have all been suggested as possible causes of the die-back of Phragmites australis (CANy.) TRIN. ex STEUD., in northern Europe. DEN HARTOG et al. (1989) pointed out that "increased eutrophication in lakes generally leads to phytoplanktonic blooms, and consequently to an increase of the organic matter content and lowered redox potential of the bottom". They suggest that the root systems of the reed will come under more stress and that survival will depend upon (1) the efficiency of the internal gas transport system to provide the roots with sufficient oxygen; (2) the capacity of the plant to deal with poisonous compounds such as ammonia and hydrogen sulphide; (3) the ability of the new sprouts to overcome the handicap of the reduced light levels in the water layer before they reach the surface, and (4) the response of the plants to the increased supply of mineral nutrients, particularly of nitrogen. There is some evidence that increased nitrogen leads to increased branching of the below-ground parts, to a continuation of tillering to the end of the growing season and that the delayed switch
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

128 J. Armstrong et al.
from the vegetative to the generative phase reduces the translocation of reserve materials (KOHL & KOHL 1993).
Various disorders of rice (VAMoS 1959, BABA et al. 1965, TANAKA & YOSHIDA 1970, ALLAM & HOLLIS 1972), and "die-back" of other plant species such as Spartina in waterlogged soils have been attributed to intensely reducing conditions in the soil and the accumulation of phytotoxins such as hydrogen sulphide (GOODMAN & WILLIAMS 1961, PEZESHKI et al. 1988, MENDELSSOHN & MCKEE 1988). In the case of Phragmites australis, KOVACS et al. (1989) have suggested that sulphide and the lower organic acids might be implicated in its decline. Prants are at particular risk in waterlogged soil if they are unable to maintain a level of internal aeration and rhizosphere oxygenation sufficient to prevent the ingress of phytotoxins (ARMSTRONG 1982).
Thi paper is concerned with a comparison of healthy and die-back Phragmites sites in both Hungary and the U.K. in terms of the redox regimes, the concentrations of one of the potential phytotoxins, namely sulphide, and the identification of die-back symptoms, both above and below ground, in terms of morphology, anatomy (especially with regard to callus development), and gas-flow characteristics, namely the resistance to Poiseuille flow into the rhizome system. The paper also describes laboratory experiments in which Phragmites plants were subjected to acetic acid (1.67 mM - 100 ppm; 0.87 mM - 52 ppm) and sulphide (1.4 mM - 45 ppm) for at least 30 days, in order to discover how the plants might be affected, and particularly to see if any of the abnormalities noted at the die-back sites could be induced by these phytotoxins. Root and shoot growth were monitored and the plants were examined anatomically, especially for callus development, blockages in the aeration and vascular systems and for any abnormal lignification/suberisation. The symptoms of laboratory-induced phytotoxin damage are compared with the characteristics of plants from die-back sites. Finally, from the results of a greenhouse experiment, some preliminary observations are made on the effects of decaying root and rhizome material on Phragmites growth and soil redox potentials.
MATERIALS AND METHODS
Sites
U.K. Die-back Site: Saltmarshe Delph Saltmarshe Delph (map ref. SE 775248) near to the town of Goole, is a small triangular
freshwater lake fringed on two sides with reed swamp composed chiefly of Phragmites and Typha. Considerable die-back of the Phragmites occurred during the last ten years up to 1990. Due to falling water tables during 1990-1992 large areas of bare organic sediment were exposed, and the earlier domination by Phragmites was still visible in the form of a persistent stubble. By 1993 there was already some evidence of regeneration from surviving plants but what appeared to be die-back symptoms in the form of aborted buds were common below-ground; in places there has been significant recolonisation from seed.
U.K. Healthy Site: Faxfleet This is a healthy, apparently stable reed bed on the edge of the River Humber a few miles
to the east of Saltmarshe Delph (map ref. SE 852241). The stand was characterised by the uniformity of its appearance. In contrast to the die-back site there was no obvious clumping or bud death and old flower heads were plentiful.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 129
Hungary' Lake Ferto sites Lake Fert6" (Neusiedler See) is a large shallow brackish lake lying across the frontier
between Hungary and Austria. Its southern end, lying within Hungary (16041'E - 47039'N), is bordered by extensive areas of reed marsh (65 km ) much of which until recent times has been regularly harvested by traditional techniques. Also, until recently, water levels in the lake had not been controlled, and substantial seasonal fluctuations in water level were normal. Over the past ten years, however, by an agreement between Austria and Hungary, the water levels in the lake have been stabilised and mowing has become more intensive and is carried out using much larger machinery than previously. Coinciding with these management changes, very extensive areas of die-back have developed within the reed belt. Some of these show the clumping habit (Plate la) which characterised the U.K. site; in some others, more recently mown, the reed appeared to have been completely destroyed (DINKA et al. 1994).
Site measurements These were made either from a boat or from elevated walkways (Plate 1 a) in order to avoid
trampling and damaging the reed.
Soil conditions
Redox profiles These were obtained using Pt-wire electrodes (Pt-tip 10 mm x 1 mm - thermo-pure wire;
shaft 600 mm epoxy-resin filled acrylic tube; o.d. 6 mm; ARMSTRONG & ARMSTRONG 1994) in conjunction with a Ag/AgCl reference electrode (reference potential +I 99mV; ARMSTRONG & WRIGHT 1976), and a high impedance digital electrometer/polarograph. Sediment pH was close to 7 at all sites and hence no pH-based corrections of redox potential have been made in the data presented.
Sulphide measurement (field and laboratory) These were carried out polarographically by means of a dropping mercury electrode (DME)
half-cell in conjunction with a Ag/AgCl half-cell following a previously reported procedure (ARMSTRONG 1967), but with an improved design of DME developed for this study.
Gas flow resistances The flow resistance into the underground rhizome system was determined by measuring
gas-flow into the bases of old culms under an applied pressure delivered from a portable compressed-air cylinder, but taking into account any ambient pressurisation within the stubble. The flows were measured by recording the pressure drops across a specially constructed flow cell using a Furness Instruments (Bexhill) CPFM Portable Pressure Transducer (range 199.9 Pa or 1999 Pa). A separate pressure transducer on the stubble side of the flow cell helped to record the applied and any stubble pressures. Conductivities into the beds were calculated by appropriate substitution in the Poiseuille equation (ARMSTRONG et al. 1988).
Laboratory experiments
Plant material Healthy horizontal rhizomes (length from 270-520 mm) of Phragmites, collected from the
banks of the Humber estuary were placed, with one end emergent, in 1/4 strength Hoagland's
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

130 J. Armstrong et al.
solution in glass cylinders (height: 400 mm; diameter: 50 mm) in a growth room to resume root and shoot production. The growth room was maintained at 18 ?C in continuous light (PAR: 50-80 jmol m-2 s-I). The rhizomes were secured in position by being taped to the neck of the tube to allow free access to the atmosphere for emerging shoots. The tubes were sheathed in black polythene to prevent light from reaching the rhizomes and roots. Care was taken initially to confim that there was no callus blocking the nodal septa of the rhizomes. This was achieved by applying gas-pressures to the ends of the rhizomes and measuring the pressure flow resistances.
Treatments After 1-2 weeks when the roots had begun to emerge, the rhizomes were transferred to a
supporting medium containing single phytotoxins in 0.05% (w/v) agar: water to maintain stagnation. Treatments were as follows with numbers of replicates in brackets: Acetic acid - 1.67 mM: 100 ppm (3), 0.87 mM: 52 ppm (3); Sulphide - 1.4 mM: 45 ppm added as sodium sulphide Na2S . 9 H20 (3); Controls - no phytotoxin (4). No nutrients were added to the medium in order to ascertain the effects of the phytotoxins alone; the rhizomes themselves contained sufficient nutrient for growth during the 30 days of the treatments.
Root and shoot growth were measured every 24 hours, and the plants were harvested after 30 days. At harvesting the nodal gas-flow resistances of the rhizomes were measured. Total lengths of lateral roots were also recorded.
Anatomy: field and laboratory experiments From the field, rhizomes and whole plant clumps were dug up, or in the case of some
die-back sites, simply pulled out of the sediment, and were dissected and examined in the laboratory for signs of bud and root death. Plants from the phytotoxin and control treatments were similarly sampled. Examples were recorded and photographed. From both field specimens and those from the laboratory experiments, sections were made of roots (different axial positions), rhizome nodes, root-rhizome junctions, bud-rhizome junctions and rhizome cortical aerenchyma, and examined for callus, lignification (phloroglucinol and conc. hydrochloric acid, and confirmed using aniline sulphate) and suberisation (Sudan III). Sections were photographed as required using a Zeiss Photomicroscope.
Greenhouse experiment: to investigate effects on growth of rotting rhizomes and roots One year old Phragmites plants, raised from seed, and grown initially in plastic "trainers"
in a waterlogged loam, were placed in plastic boxes (560 mm long x 360 mm tall x 70 mm deep) with hinged fronts, which contained either (a) sand, (b) sand + organic layer, (c) loam, or (d) loam + organic layer. The organic layer in each case consisted of one litre of freshly chopped rhizomes and roots of Phragmites, (volume measured by displacement of water), mixed with a litre of moist sand or loam as appropriate, and placed as a layer 100 mm in depth at a distance of 50 mm from the soil surface in each box. The total depth of soil was approximately 300 mm. Towards one end of each box a break was made in the soil and organic layer to accommodate the rhizome system of one plant - the average fresh weight per plant was 30 g, the average shoot number was seven, and the length of the rhizome system was approximately the same as the soil depth. The soils were waterlogged by inserting the
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 131
boxes in tanks of water and the water surface was maintained at all times at approximately 20 mm. There were six replicates per treatment.
New shoot numbers per plant, and their total lengths, and the redox profiles in the soils. were recorded at approximately 2-week intervals. The results presented are for the first 65 days of the experiment.
RESULTS AND DISCUSSION
Field observations
Plant characteristics Striking features of the plants at die-back sites in both Hungary and the U.K. included (a)
the ckt-mped habit (Plate. la, and also shown by SUKOPP & MARKSTEIN 1989), often with stunted shoots: in the case of total die-back the clumps were evident only as stubble; elsewhere, particularly at the UK site, the clumps often included very large numbers of old culms many of which were rotted at the base, (b) scarcity of new shoots and flowering stems (Tab. 1 and Plate 1 a), and (c) the death of both vertical (Plate lc) and horizontal rhizome apices, and the presence of short adventitious roots with dead apices, and death of laterals (Plate lb). The death of rhizome apices is probably a cause of the clumped habit, and in the case of the vertical rhizomes, of the sustained tillering due to a loss of apical dominance (Plate ic). In some regions of die-back, insect damage was common due to species of Chilo, Archanara, Tomasella and Rhizedra. Fungal damage due to Puccinia phragmitis KORN., was also noted in some locations. None of these symptoms was obvious at the healthy sites. (More details are described in ARMSTRONG & ARMSTRONG 1995).
Gas-flow resistances into reed beds At the die-back sites in both Hungary and the U.K. there was often considerable resistance
to flow when pressures were applied to the bases of old culms: in the U.K. a majority of previous years' die-back culms had infinite resistance (zero conductivity) to gas-flow (Tab. 1). The resistance was located at the bases of culms in the form of callus development. Old culms normally remain open for several years and none of the old culms examined at the healthy site were blocked. In the U.K., culm basal callus was sometimes associated with the holes of rhizome boring insects. The presence of callus in old culms and/or within the rhizome system will increase resistance to flow both by humidity- and Venturi-induced convections (ARMSTRONG et al. 1992, KOHL et al. 1996). However, the prevalence of blockages in old culm bases will not necessarily totally prevent convective gas-flow venting from the rhizome system: rates of flow depend on the ratio of living to non-callused old culms (ARMSTRONG & ARMSTRONG 1990).
Anatomy In all the rhizomes dug or pulled up from the die-back sites, callus, wholly or partly
blocking the nodal diaphragms was a very common feature (Plate 2a), and often associated with rotten internodes and nodes. Regions of the aerenchyma channels close to the node were also frequently blocked by callus. In all cases of bud death examined, callus had completely blocked the diaphragms in the two or more nodes below the decayed apex (Plate lc). It was particularly interesting that callus had developed in the rhizome nodes subtending enlarged
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

132 J. Armstrong et al.
Table 1. Summary of some plant and soil data from Hungarian and U.K. healthy and die-back sites.
Sites Die-back Healthy
Location Some No regeneration regeneration
No. of young culms m-2 (U.K.) 81 0 190
No. of old culms m-2 (U.K.) 227* 0 229 (height > 25 cm) clumped
stubble
No. of old culms m-2 (U.K.) 0 0 119 with flower heads
No. of old culms m-2 (U.K.) 54 0 0 with insect damage
Mean pressurised gas-flow (U.K.) 1.5 0 100.5 conductivity via old culms (10-5 m3 MPa-1 s 1)
Mean Eh (mV) at 40 cm (U.K.) -80 -160 +30 into sediment
Mean Eh (mV) at 40 cm (Hungary) -170 -30 into sediment
Mean conc. (mM) of dissolved (Hungary) 3.2 trace sulphide at 40 cm (108 ppm)
Note: * this number includes many old culms greater than one year old that had rotted at the base.
lateral buds below the decayed apex. Some of these lateral buds were showing a yellowing of the apical meristem. If the formation of this subtending callus preceded or accompanied the enlargement of the lateral bud, the latter would be likely to die also through lack of oxygen. On the other hand it is possible that the buds were affected by a phytotoxin, e.g. sulphide or organic acids, or infected by a pathogen, perhaps fungal, which killed the apex and subsequently induced callus formation in the nodes below.
A striking characteristic of adventitious roots from die-back sites was the presence of callus, sometimes lignified, and partly (Plate 2b), or wholly (Plate 2d) blocking the cortical aerenchyma channels. Callus development in the aerenchyma of roots themselves is a phenomenon which we had never seen before, and we are not aware of it having been previously reported. Similarly, callus was frequently found to be blocking the cortical aerenchyma at root-rhizome junctions. (This latter effect has previously been noted to a limited extent from healthy sites where roots have died, or have been artificially abscised during laboratory experiments.)
The death of adventitious root apices was associated with a band of cortical lignification visible in longitudinal section. Both thick and slender adventitious roots from die-back locations also had abnormal lignifications and suberisations of cell walls of the apical 3-4 mm region in the epidermal layer (including that of the root-cap), the hypodermal layers, and in the non-aerenchymatous cortex. Often, what appeared to be lignin deposits were to be seen within the cortical intercellular spaces of these non-aerenchymatous apical regions. From
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 133
apex to base, the hypodermal and stelar regions of the roots were much more heavily lignified and suberised than those from healthy sites (Plate 2b), with the deposits sometimes extending to the cell walls at the centre of the stele, even within the apical 2 mm.
Lateral roots generally had lignified and, to some extent, suberised epidermises and frequently were found to have died after emergence (Plate 2d) or before emergence. In the latter case the swollen laterals appeared to be imprisoned in the cortex. Possibly their growth had been curtailed because of the aforementioned lignification in the adjoining hypodermal and epidermal layers. Normally the cell walls of these layers are unthickened forming "windows" for lateral root emergence (JUSTIN & ARMSTRONG 1987, ARMSTRONG 1992, VOTRUBOVA & PECHACKOVA 1996). Altemratively, phytotoxins may have entered through these non-lignified "windows" killing the laterals and inducing cortical callus and hypodermal and cortical lignification. Callus was also often associated with those regions of dead emergent laterals embedded in the adventitious root cortex. Both callus and cortical cells were often lignified and the pit fields were prominent, a very unusual feature for cortical cells. In these cases the phytotoxins could have entered via the laterals or their dead remains.
Lignification is well documented as a defence reaction in plants, often in response to microorganism attack (FRIEND 1981, ASADA & MATSUMOTO 1987, MATERN & KNEUSEL 1988). It has also come to be regarded as a root adaptation to waterlogging in flood tolerant species (YAMASAKI 1952). It is envisaged that the development of callus within the aeration pathways of rhizomes, rhizome-root junctions or roots will arrest or reduce the oxygen supply to the roots and the rhizosphere.
Another interesting feature of rhizomes and adventitious roots from die-back sites was the presence of lignified blockages/tyloses within the xylem and phloem (Plate 2b, d). In the apical regions of roots it was frequently the protoxylem rather than metaxylem which was blocked, but in rhizomes and root-rhizome junctions metaxylem elements were also occluded. It would seem that a feature of die-back may be an interference with water and sugar transport. In addition, however, it is almost certain that water and mineral nutrient absorption would be severely curtailed by the stunting and death of lateral and adventitious roots, and by the lignification/suberisation of the surfaces of lateral roots and even the apical regions of adventitious roots (ROBARDs et al. 1979). The effects of phloem blockages, together with early senescence and sustained tillering might be, as suggested by CI2KOVA -KON(ALOvA et al. (1992) and KOHL & KOHL (1993), to reduce the storage of carbohydrates in overwintering rhizomes and buds to levels critically below those normally found in healthy plants (HALDEMANN & BRANDLE 1986). Equally important may be impeded transport of assimilates into the buds.
Lastly, it should be noted that none of the above features was common at healthy sites.
Soil Characteristics
Redox measurements The redox potentials at both the Hungarian and the U.K. die-back sites were generally
lower than those found at the healthy site (Tab. 1). The lowest values, ca. -220 mV, were found where clumps in open water were only of dead stubble with no new shoots. Where there were some signs of regeneration around clumps, potentials were generally a little higher than this: e.g. ca. -80 mV at a depth of 40 cm. In the U.K., and generally in Hungary, potentials
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

134 J. Armstrong et al.
at the healthy sites did not fall below -100 mV and were usually between 140 and 200 mV higher than at the die-back sites.
While the lower potentials of the die-back sites are indicative of conditions which should be potentially more injurious than those at the healthy sites, it must be borne in mind that the higher potentials of the healthy sites probably very much depend upon the greater vigour of the plants and their sediment oxygenating potential. Any disturbance to this equilibrium might create conditions which the plants could no longer ameliorate, and this may be the situation created at the die-back sites. Even a small rise in water table might reduce the oxygenating potential of the roots, or cause anoxia in root and horizontal rhizome apices.
Sulphide concentrations In Hungary, a strong smell of sulphide emanating from Lake Ferto is often noticeable even
at con,-iderable distances from the lake and, at the die-back sites the sediments, besides being characterised by large quantities of dead and decaying rhizomes and roots, also smelled very strongly of hydrogen sulphide. In Lake Ferto, sulphate concentrations in the waters of between 470 ppm (4.9 mM) and 1000 ppm (10.4 mM) are a normal feature (DINKA et al. 1994), and exceptionally high levels of sulphide were a characteristic feature of the sediment interstitial waters at the die-back sites (Tab. 1). The highest concentrations (136 ppm: 4mM) that we recorded were at a site which had been mown the previous winter; by the end of August there was still no recovery of the Phragmites. At our main site of study (Plate la), except at the surface, sulphide levels were high throughout the profile. In the vigorous stands, however, the highest concentrations (< 30 ppm: 0.88 mM) were often in the surface layer, and at 30-40 cm there was generally only a trace, presumably because of rhizosphere oxidations.
The U.K. site is downwind of several large coal-fired power stations which could have been a source of sulphate in the waters, and is close to a river jetty where sulphate and phosphate fertilizer material had been unloaded in the recent past. However, despite the low redox levels, we were never able to measure more than a trace of sulphide in the interstitial waters, and if sulphide was/is involved in die-back at the site its presence may have been/is transitory or very localised. The U.K. sediment, however, is highly organic in nature, and it is likely that here, organic acids from the decay of the underground parts of the plant were more important at the time of our investigation; also, throughout the summer of 1993 there were intense algal blooms in the waters.
Phytotoxin treatments
Root and shoot growth In both the 1.67 mM acetic acid and the 1.4 mM sulphide treatments, the adventitious roots
remained stunted and growth had virtually ceased or was very slow after 5 to 10 days (Fig. 1). In contrast, the control roots were still growing after 30 days and their mean lengths were at least ca. 5x greater than roots in these two treatments; the total root length per control plant was at least 6.6x greater than in the two treatments (Plate 3a, b, c). Adventitious roots in the 0.87 mM acetic acid treatment were only growing very slowly at 30 days, but on average, individual control roots were 2.7x longer; the total root length of control plants was twice that of those in this acetic acid treatment.
Lateral root growth in 1.67 mM acetic acid and 1.4 mM sulphide was extremely stunted (Tab. 2), average lengths being < 0.5 mm in the former and 3 mm in the latter, compared to
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 135
280 -r ' - ' X . -
260-
240 Cols
220-'AV
200 -
- II
E 180 -
Th 160-
140p 120kT T IIIIiI.08
L0 Q / 4 i ;,;W; T4) 0 0 1.67mM acetc add
20LI+ Nofrt~rHer 14 mM sulphide
0 5 10 15 20 25 30
Time (days)
Fig. 1. Phragmites rhizomes: adventitious root growth in response to dissolved suiphide and acetic acid. Supporting medium was 0.05% agar:water. Data are means ? s.e.: controls, n=22 roots; suiphide, n=18; 50 and 100 ppm acetic acid, n=21.
12 mm for the controls (Plate 3a, b, c). The total length of laterals per control plant was 143x that for plants in 1.67 mM acetic acid and 29x that for plants in the sulphide treatment. The laterals in 0.87 mM acetic acid were less stunted, but looked brown.
Total shoot lengths were shorter and the degrees of senescence were greater in the treatments compared to the controls, especially in 1.67 mM acetic acid and 1.4 mM sulphide (Tab. 2). In these two treatments the grogth of buds and their death before, or shortly after, emergence, were observed. In all cases of bud death, callus was found to be present in at least some of the nodal diaphragms. In some instances, callus formed within the original rhizome, appeared to have occluded the aeration system of the submerged bud (Plate 3d).
Gas-flow features and callus development in rhizomes Higher than normal resistances to pressurised throughflow in rhizomes were correlated
with the development of callus within the nodal diaphragms, e.g. Plate 2c. Where there was infinite resistance, the aerenchyma channels were also similarly blocked. The percentages of nodes totally or partially occluded were much greater in the treatments, especially in 1.67
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

136 J. Armstrong et al.
Table 2. Phytotoxin experiment: lateral root and shoot growth on rhizome cuttings in response to acetic acid or sulphide over 30 days. Mean + s.e. and number of measurements in brackets.
Control Acetic acid Sulphide (0.87 mM) (1.67 mM) (1.4 mM)
Lateral roots Produced from original
adventitious roots mean length (mm) 12 - > 0.5 3
total length/root (mm) 5903 ? 1079 (25) 520 ? 123 (25) 45 ? 20 (18) 215 ? 41 (19) total length/plant (mm) 33948 ? 8163 (4) 4335 ? 901 (3) 271 ? 212 (3) 1365 ? 216 (3)
Total length/plant on original + new roots 40412 ? 8410 (4) 5042 ? 887 (3) 282 ? 205 (3) 1405 ? 248 (3)
Emergent shoots Number/plant 3.0 2.3 2.3 1.5 Mean total length
attained/plant (mm) 1301 ? 145 (4) 1006 ? 212 (3) 443 ? 29 (3) 703 ? 238 (3) Total final non-senesced
shoot length/plant (mm) 1170 506 133 267 laminae failed
to expand
mM acetic acid and sulphide, than in the controls (Tab. 3). Also, the percentage of nodes remaining callus-free was considerably greater in the control plants. We conclude that callus was induced to a greater extent in the treatments probably because of the phytotoxins entering the plants via the roots, where some degree of cortical callus was also observed.
In all plants, including the controls, the lowermost (submerged) node was invariably totally blocked and the emergent node was often partially blocked. Prior to this (ARMSTRONG & ARMSTRONG 1988), we have frequently observed the development of callus at the cut ends of rhizomes in solution culture and conclude that it forms in response to wounding and/or flooding or dehydration. Callus in the rhizomes was also common at die-back sites (Plate lc, 2a) but not at healthy sites. At the Hungarian die-back site where heavy mowing machines had caused wide-spread death, and sulphide levels had become excessively high (ca. 4 mM), we suggest that callus formation and the blockage of aeration pathways in response to mechanical and phytotoxin damage will almost certainly have been major factors contributing to the die-back.
Anatomy In the treatments, particularly in 1.67 mM acetic acid and sulphide, apical regions of
adventitious roots were abnormally lignified in the surface cell layers, in the cell walls and intercellular spaces of the non-aerenchymatous cortex. In 1.67 mM acetic acid, where the apices had died, there was always a fungal growth which might have been saprophytic. Also in these treatments, thin-walled callus within the basal aerenchyma spaces of adventitious roots was observed. Lateral roots with dead apices and lignified cell walls were present. and there was excess lignification of the adventitious root cortex surrounding the laterals (Plate 4c): phytotoxins entering via the damaged laterals might have stimulated this response in the cortex.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 137
Table 3. Effects of sulphide or acetic acid on the development of nodal callus in Phragmites rhizome cuttings. Total blockage indicates callus occlusion of nodal diaphragm and cortical aerenchyma channels. Partial blockage generally indicates total blockage of diaphragm and some/all of aerenchyma channels free. Mean % per plant t s.e. and numbers in brackets refer to number of plants sampled.
Treatment Number Ave. nodes Per cent nodes per treatment of plants per plant
Totally blocked Partially blocked Free
Controls 4 6 17.2 1.6 (4) 27.9 ? 9.8 (4) 55.0 ? 9.1 (4) 1.4 mM: 45 ppm sulphide 5 5.2 64.6? 16.0 (5) 23.7 11.7 (5) 11.7 ? 4.8 (5)
1.67 mM: 100 ppm acetic acid 3 5.3 66.7 ? 24.0 (3) 6.7 ? 6.7 (3) 26.7 ? 17.6 (3)
0.87 mM 52 ppm acetic acid 3 4.7 33.7 ? 5.2 (3) 49.2 ? 7.9 (3) 27.8 ? 14.7 (3)
In botlh types of treatment, laterals were also found "imprisoned" in, and sometimes growing down through, the adventitious root cortex (Plate 4a). This was probably due to lignification of the hypodermal cell layers of the adventitious roots. Normally, lateral roots emerge through non-thickened "windows" in these layers (see section on field anatomy). Tyloses or lignified blockages were found occluding conducting elements of both xylem and phloem in the adventitious roots and in the root-rhizome junctions, (Plates 3e, 4a, b, d); in the latter, the cortex frequently had become totally blocked by lignified callus (Plate 4b, d). These effects are similar to those found in die-back plants in the field (cf. Plate 2b, d).
It should be noted that control plants and those from healthy sites in the field rarely exhibited any of the above features.
Greenhouse experiment: effects of organic layers After 65 days, both the numbers of new shoots and total lengths were considerably greater
in the control plants than in those growing in the presence of a sub-surface organic layer, (Fig. 2), but these effects were most marked in the sand treatments. Here control shoot lengths per plant were 16x and shoot numbers 8.6x those of plants growing with the organic matter. In both the sand and loam the organic matter had the effect of lowering the redox potential in its vicinity: after 30 days this was ca. -100 mV in both soils compared with +100 mV for the controls. Later, although the values rose somewhat, they remained at least 200 mV lower than the controls (Fig. 2).
When the underground parts of the plants were examined by unhinging one side of each container, in the organic matter treatments the growth of rhizomes and roots was considerably less than that of the controls; again, the effect being more marked in the sand culture, and the roots and rhizomes having tended to avoid growing in the organic layer. However, in none of the treatments did we see the signs of bud and root death that we had observed at die-back sites. This was possibly because the organic layer was confined to only a few centimetres of soil depth, the plants had a large volume of comparatively uncontaminated soil in which to grow, and were initially planted in a gap created in the organic layer. Nevertheless, the experiment demonstrates a clearly inhibitory effect on bud emergence and on growth both above and below-ground. We conclude that this was probably due to the release of phytotoxins, e.g. organic acids and sulphide, from the anaerobic decomposition of the organic layer (KovAcs et al. 1989). We tentatively suggest that the smaller detrimental
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

138 J. Armstrong et al.
600 r f T , 600
500 500
g ,1 T,1
4 00 T _4-0200
- 300 O_ 200 ~~~oo TB X\T~~~~~~~~~Q
100 V 100
0 { D 0
-100~ A- 4~A' -100
-200 - -200
- 300 . 300 0 5 10 15 20 25 0 5 10 15 20 25 30
Depth (cm) Depth (cm)
25 ~~~~~~~~~~~~~~~~~~~~1600 T1400
20~~~~~~~~~~~~~~ T Ct ~~~~~~~~~1200
E K ii~~~~~~~~~~~~~~~~~~~~~
.0 10 ~ ~ ~ ~ ~ ~ 1 0 07
Days Days
Fig. 2. Greenhouse experiment: effects of sub-surface organic layer on the growth of Phragmites, and soil redox potentials, in waterlogged sand and loam-based soils. Top - redox profiles at 57 days: left - (V) loam control, (0) loam plus organic layer; right - (OI) sand control, (A) sand plus organic layer. Bottom - new shoot numbers and total lengths per plant: A, loam plus organic layer; B, loam control; C, sand control; D, sand plus organic layer. Means ? s.e. of six replicates per treatment. Each replicate is represented by one plant. NB: Organic layer where present extends from 5 to 15 cm on the depth axis.
effect of the organic layer in the loam may have been due to some neutralising effect on the phytotoxins; differences in nutrient supply seem an unlikely explanation since growth in the sand control was better than that in the loam.
FINAL COMMENTS
The growth experiments confirm that sulphide and acetic acid can be highly toxic to Phragmites, and the morphological and anatomical investigations reveal that they can induce
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 139
l
RAISE OR STABLE WATER TABLES
EXCESSIVE ORGANIC LOADING INSECT AND FUNGAL DAMAGE
PHYSICAL DAMAGE HIGH TEMPS
EUTROPHICATION
high N lgal blooms IMPEDED AERAION
DEATH OF UNDERGROUND ORGANS
sulphate I
ACCUMULATED ORGANIC
MAIrER
I I ~~~~~ ~~(anaom denoto)
PHYIrOXINS suAphlde and oqIanic acids
r- - - - - - - - - - - - - - -_- -_- - - - -___ __ ___
celus in aeration system POOR ROOT GROWTH I mrot and tizones AND DEATH I
REDUCED WATER, NUTRIENT AND CHO re
I IMPEDED AERATION TRANSPORT SeNESCENCE i
reduced CH 0reserves
ROOT AND RHIZOME, ET SUSANED i DEATH i BUD ID)EATH TILLERING
DIE-BACK
Fig. 3. Tentative scheme showing possible pathways to the die-back of Phragmites, based on the findings of the present study, the work of the EUREED project group and reports in the literature; CHO = carbohydrate.
symptoms recently found to be characteristic of plants from die-back sites (ARMSTRONG & ARMSTRONG 1995), some of which had concentrations of dissolved sulphide as high as 4 mM (140 ppm). These symptoms include restricted adventitious root, lateral root and bud growth, death of root apices and buds, abnormal lignification, callus formation in roots and rhizomes causing blockages in the aeration pathways, and blockages in the xylem and phloem of the vascular system. Although phytotoxins such as sulphide and acetic acid have previously been shown to restrict root growth and cause some death (VAMos 1959, GOODMAN & WILLIAMS 1961, LYNCH 1978, 1982, SANDERSON & ARMSTRONG 1980, KOCH et al. 1990), so far as we are aware, callus production in the gas-transport system, and vascular blockages have not previously been reported as responses to phytotoxins in the rooting medium.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

140 J. Armstrong et al.
The previously known phytotoxic effects of sulphide include the inhibition of respiration (in rice, ALLAM & HOLLIS 1972; in Spartina, MENDELSSOHN & MCKEE 1988; and in Phragmites, FURTIG et al. 1996), and in Spartina both the induction of carbohydrate deficiency (MENDELSSOHN & MCKEE 1988) and photosynthetic inhibition (PEZESHKI et al. 1988). The latter effect was detected at concentrations as low as 0.01 mM (0.34 ppm) in stirred solution culture. Recently FURTIG et al. (1996) have shown that Phragmites has some capacity to detoxify sulphide by incorporating it into glutathione. However, it seems reasonable to conclude that the concentrations of sulphide in the Hungarian die-back sites (4 mM) and that used in the phytotoxin treatments (1.4 mM), albeit in comparatively stagnant media, would be sufficient to inhibit photosynthesis and respiration, and these effects may also help to account for the premature senescence observed in the die-back sites and phytotoxin experiments.
De:,,pite there being many theories about the causes of die-back in Phragmites, very little information can be found in the literature which describes the symptoms of plants affected by the syndrome. This study has identified several morphological, anatomical and physiological abnormalities common to plants from die-back sites and to plants treated with phytotoxins in the laboratory. As a result of discussions about die-back with members of the EUREED Group and others, and after reviewing the literature, we suggest a syndrome in which anthropogenic influences and phytotoxin damage are key factors in the initiation and perpetuation of Phragmites die-back (Fig. 3).
We envisage that firstly some critical or catastrophic event(s) occur(s) which locally cause(s) a greater than normal rate of death of plant parts. Causal factors might be raised or stabilised water-tables (WEISNER 1988, WEISNER & GRANELI 1989, HELLINGS & GALLAGHER 1992, DINKA, pers. comm), mechanical damage (BOORMAN & FULLER 1981, OSTENDORP 1989, DINKA, pers. comm.), organic matter accumulation (KLOTZLI 1971, FIALA 1976, CIZKOVA -KONCALOVA et al. 1992, DEN HARTOG et al. 1989, KovAcs et al. 1989), development of intensely reducing soil conditions, eutrophication (CIZKOVA-KONCALOVA et al. 1992, DEN HARTOG et al. 1989, KOHL & KOHL 1993, OSTENDORP 1989), phytotoxin accumulation e.g. organic acids and sulphide (KovAcs et al. 1989), raised temperatures and insect or fungal damage. These factors can interfere with internal aeration whether driven by humidity-induced or Venturi-convection or by diffusion (ARMSTRONG & ARMSTRONG 1995, and in prep.), and/or carbohydrate balance (HALDEMANN & BRANDLE 1986, CIZKOVA-KONPALOVA et al. 1992), and maybe the water and mineral uptake of the reed, leading locally to stunted growth and an abnormally high rate of death of underground organs.
Consequently, as a result of the decay of the plant, and/or other sources of organic matter, there will be locally an accumulation of high levels of phytotoxins such as hydrogen sulphide (especially in sulphate-enriched waters), and the volatile organic acids (e.g. formic, acetic, butyric, propionic, caproic); this may be especially so in stagnant, warm conditions. These phytotoxins could be expected to induce further root, rhizome, bud and shoot death, and premature shoot senescence, and, ultimately, prevent the recovery of the plant in the short term, thus producing the vicious circle which we know as the die-back syndrome (Fig. 3).
Acknowledgements: For financial support, we thank the European Community (3rd framework Environment Programme - contract EV5V-CT92-0083 - EUREED: co-ordinator, Dr. W.H. van der Putten of the Netherlands Institute of Ecology, Heteren). We also thank Dr. Maria Dinka and Dr. Peter Szeglet of the Institute of Ecology and Botany, Hungarian Academy of Sciences, Vacrat6t, and the Director, Fert6rdkos Research Centre for
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

A role for phytotoxins in the Phragmites die-back syndrome? 141
making possible our visits to Lake Fert6, Mr. Ian Armstrong and Mr. Graeme Pittaway for assistance in the field, Mr. Vic Swetez for help with the greenhouse experiment, Mr. Sayed Md. A. Zobayed for help with the phytotoxin experiments, Mr. Richard Holt for photographic assistance, Mr. M. Bailey and Miss S. Lythe for technical help and Mr. Jack Taylor, Dr. D.J. Boatman and the Yorkshire Naturalist Trust for information and access to Saltmarshe Delph.
REFERENCES
ALLAM A.I. & HOLLIS J.P. (1972): Sulphide inhibition of oxidases in rice roots. Phytopathology 62: 634-639. ARMSTRONG J. (1992): Pathways and mechanisms of aeration in Phragmites australis. PhD Thesis, University
of Hull, England. ARMSTRONG J. & ARMSTRONG W. (1995): U.K. Group Report. In: VAN DER PUrTEN W.H. (ed.), Reed News 3,
Reports of EC Project EUREED - EV5V-C792-0083, Netherlands Institute of Ecology, Heteren, pp. 6-23. ARMSTRONG J. & ARMSTRONG W. (1988): Phragmites australis - A preliminary study of soil oxidizing sites
and internal gas transport pathways. New Phytol. 108: 373-382. ARMSTRONG J. & ARMSTRONG W. (1990): Pathways and mechanisms of oxygen transport in Phragmites
australis. In: COOPER P.F. & FINDLATER B.C. (eds.), The use of constructed wetlands in water pollution control, Pergamon Press, Oxford, pp. 41-45.
ARMSTRONG J., ARMSTRONG W. & BECKETr P.M. (1988): Phragmites australis - A critical appraisal of the ventilating pressure concept and an analysis of resistance to pressurised gasflow and gaseous diffusion in horizontal rhizomes. New Phytol. 110: 383-390.
ARMSTRONG J., ARMSTRONG W. & BECKETr P.M. (1 992): Phragmites australis: Venturi- and huriidity-induced convections enhance rhizome aeration and rhizosphere oxidation. New Phytol. 120: 197-207.
ARMSTRONG W. (1967): A polarographic technique for the measurement of dissolved sulphide in waterlogged soils. J. Soil Sci. 18: 282-288.
ARMSTRONG W. (1982): Waterlogged soils. In: ETHERINGTON J.R. (ed.), Environment and plant ecology, 2nd ed., John Wiley & Sons, London, pp. 290-330.
ARMSTRONG W. (1994): Polarographic oxygen electrodes and their use in plant aeration studies. Proc. Roy. Soc. Edinburgh, Ser B, 102: 511-528.
ARMSTRONG W. & WRIGHT E.J. (1976): A polarographic assembly for the large scale sampling of soil oxygen in the field. J. Appl. Ecol. 13: 849-856.
ASADA Y. & MATSUMOTO J. (1987): Induction of disease resistance in plants by a lignification-inducing factor. In: NISHIMURA S., VANCE C.P. & DOKE N. (eds.), Molecular determinations of plant diseases, Japan Sci. Soc. Press, Tokyo, pp. 223-231.
BABA I., INADA K. & TAJIMA K. (1965): Mineral nutrition and the occurrence of physiological diseases. In: TANAKA A. (ed.), The mineral nutrition of the rice plant, Johns Hopkins Press, Baltimore, pp. 173-195.
BOAR R.R., CROOK C.E. & Moss B. (1989): Regression of Phragmites australis reedswamps and recent changes of water chemistry in the Norfolk Broadland, England. Aquatic Bot. 35: 41-56.
BOORMAN L.A. & FULLER R.M. (1981): The changing status of reedswamp in the Norfolk Broads. J. AppI. Ecol. 18: 241-269.
CdZKOVA-KONCALOVA H., KVET J. & THOMPSON K. (1992): Carbon starvation: a key to reed decline in eutrophic lakes. Aquatic Bot. 43: 105-113.
DEN HARTOG C., KvErJ. & SUKOPP H. (1989): Reed. A common species in decline. Aquatic Bot. 35: 1-4. DINKA M., SZEGLET P. & SZAB6 I. (1995): Hungarian Group Report. In: VAN DER PUTTEN W.H. (ed.), Reed
News 3, Reports of EC Project EUREED - EV5V-CT92-0083, Netherlands Institute of Ecology, Heteren, pp. 96-107.
FIALA K. (1976): Underground organs of Phragnites communis, their growth, biomass and production. Folia Geobot. Phvtotax. 11: 225-259.
FRIEND J. (1981): Plant phenolics, lignification and plant disease. Progr Phytochem. 7: 197-261. FUR1IG K., RUEGSEGGER A., BRUNOLD C. & BRANDLE R. (1996): Sulphide utilization and injuries in hypoxic
roots and rhizomes of common reed (Phragmites australis). Folia Geobot. Phytotax. 31: 143-151. GOODMAN P.J. & WILLIAMS W.T. (1961): Investigations into "die-back" in Spartina townsendii agg. III.
Physiological correlates of "die-back". J. Ecol. 49: 391-398.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

142 J. Armstrong et al.
HELLINGS S.E. & GALLAGHER J.L. (1992): The effects of salinity and flooding on Phragmites australis. J. Appl. Ecol. 29: 41-49.
HALDEMANN CH. & BRANDLE R. (1986): Seasonal variations of reserves and of fermentation processes in wetland plant rhizomes at the natural site. Flora 178: 223-226.
JUSTIN S.H.F.W. & ARMSTRONG W. (1987): The anatomical characteristics of roots and plant response to soil flooding. New Phytol. 105: 465-495.
KLOTzL F. (1971): Biogenous influence on aquatic macrophytes especially Phragmites communis. Hidrobiologia 12: 107-111.
KOCH M.S., MENDELSSOHN I.A. & MCKEE K.L. (1990): Mechanism for the hydrogen sulphide-induced growth limitation in wetland macrophytes. Limnol. & Oceanogr. 35: 399-408.
KOHL J.-G., HENzE R. & KUHL H. (1996): Evaluation of the ventilation resistance of the rhizomes of natural reed beds against convective through-flow of gases in Phragmites australis (CAV.) TRIN. ex STEUD. Aquatic Bot. (in press).
KovAcs M., TURCSANYI G., TUBA Z., WOLCSANSZKY S.E., VASARHELYI T., DELY-DRASKOVITS A., T6TH S., KOLTAY A., KASZAB L., SZOKE P. & JANKO B. (1989): The decay of reed in Hungarian lakes. Symp. Bio. Hung. 38: 461-471.
KOHL H. & KOHL J.-G. (1993): Seasonal nitrogen dynamics in reed beds (Phragmites australis (CAV.) TRIN. ex STEUDEL) in relation to productivity. Hydrobiologia 251: 1-12.
LYNCH J.M. (1978): Production and phytotoxicity of acetic acid in anaerobic soils containing plant residues. Soil Biol. & Biochem. 10: 131-135.
LYNCH J.M. (1982): The role of water-soluble compounds in phytotoxicity from decomposing straw. Pl. & Soil 65: 11-17.
MATERN U. & KNEUSEL R.E. (1988): Phenolic components in plant disease resistance. Phytoparasitica 16: 153-170.
MENDELSSOHN I.A. & MCKEE K.L. (1988): Spartina alterniflora die-back in Louisiana: Time-course investigation of soil waterlogging effects. J. EcoL 76: 509-521.
OSTENDORP W. (1989): "Die-back" of reeds in Europe - A critical review of literature. Aquatic Bot. 35: 5-26. PEZESHKI S.R., PAN S.Z., DELAUNE R.D. & PATRICK W.H. Jr. (1988): Sulfide-induced toxicity: Inhibition of
carbon assimilation in Spartina alterniflora. Photosynthetica 22: 437-442. ROBARDS A.W., CLARKSON D.T. & SANDERSON J. (1979): Structure and permeability of the
epidermal/hypodermal layers of the sand sedge (Carex arenaria L). Protoplasma 101: 331-347. SANDERSON P.L. & ARMSTRONG W. (1980): The responses of conifers to some of the adverse factors associated
with waterlogged soils. New Phytol. 85: 351-362. SUKOPP H. & MARKSTEIN B. (1989): Changes of the reed beds along the Berlin Havel. Aquatic Bot. 35: 27-40. TANAKA A. & YOSHIDA S. (1970): Nutritional disorders of the rice plant in Asia. Int. Rice Res. Inst. (Manila)
Techn. Bull. 10. VAMOS R. (1959): "Brusone" disease of rice in Hungary. P1. & Soil 11: 103-109. VOTRUBOVA 0. & PECHAIPKOVA A. (1996): Effect of nitrogen over-supply on root structure of common reed.
Folia Geobot. Phytotax. 31: 119-125. WEISNER S.E.B. (1988): Factors affecting the internal oxygen supply of Phragmites australis (CAV.) TRIN. ex
STEUDEL. Aquatic Bot. 31: 329-335. WEISNER S.E.B. & GRANtLI W. (1989): Influence of substrate conditions on the growth of Phragmites australis
after a reduction in oxygen transport to below-ground parts. Aquatic Bot. 35: 71-80. YAMASAKI T. (1952): Studies on the excess moisture injury of upland crops in overmoist soil from the viewpoint
of soil chemistry and plant physiology. Bull. Nat. Inst. Agric. Sci. B 1: 1-92.
Encl. Plate 1, 2, 3, 4, pp. III-VI.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions
J. Armstrong et al.: Phytotoxins in the Phragmites die-back 111
a
MR. _
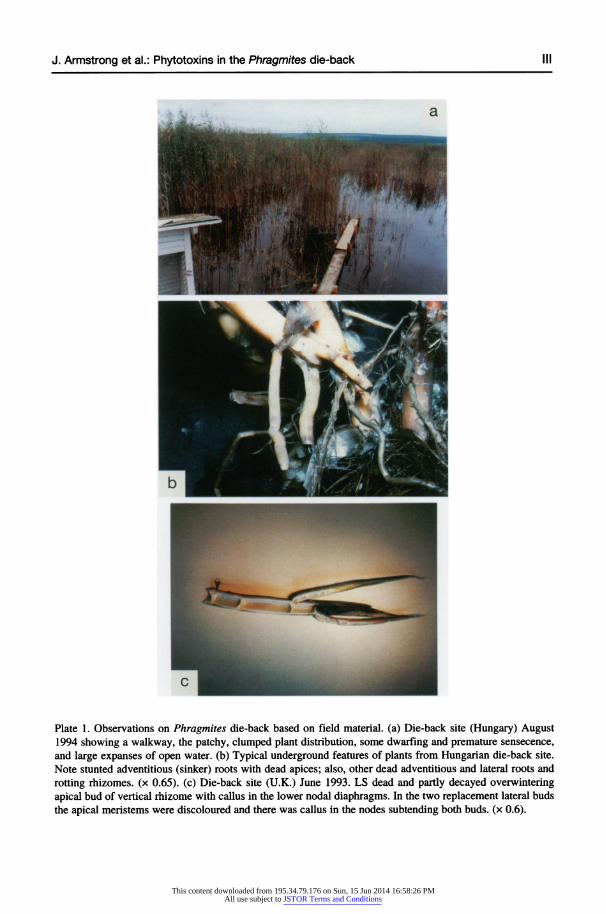
Plate 1. Observations on Phragmites die-back based on field material. (a) Die-back site (Hungary) August 1994 showing a walkway, the patchy, clumped plant distribution, some dwarfing and premature sensecence, and large expanses of open water. (b) Typical underground features of plants from Hungarian die-back site. Note stunted adventitious (sinker) roots with dead-apices; also, other dead adventitious and lateral roots and rotting rhizomes. (x 0.65). (c) Die-back site (U.K.) June 1993. LS dead and partly decayed overwintering apical bud of vertical rhizome with callus in the lower nodal diaphragms. In the two replacement lateral buds the apical meristems were discoloured and there was callus in the nodes subtending both buds. (x 0.6).
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

IV J. Armstrong et al.: Phytotoxins in the Phragmites die-back
a A
Plate 2. Observations on Phragmites die-back based on field material. (a) LS part of nodal diaphragm from U.K. die-back site with typical callus, developed here from parenchyma between vascular bundles. Note brown "impenetrable" layer on upper surface - composition unknown. (x 67). (b) TS basal region of adventitious root with intensely lignified and thickened outer cortex and lignified callus developed from outer cortical cells and partly blocking aerenchyma spaces; also, tyloses/blockages in protoxylem. (x 45). (c) As above (a) but from laboratory sulphide treatment. (x 60). (d) TS mid-length of adventitious root with dead lateral, and callus completely blocking cortical aerenchyma spaces; also, tyloses/blockages in protoxylem. (x 45). (Hand-cut sections stained with phloroglucinol and concentrated hydrochloric acid.) TS = transverse section; LS = longitudinal section.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

J. Armstrong et al.: Phytotoxins in the Phragmites die-back V
5eB _~~~~~~~~~~~~~~
Plate 3. Phytotoxin treatments involving Phragmites rhizomes. (a) controls after 15 days showing long adventitious roots and normal formation of lateral roots. (x 0.75). (b) sulphide (1.4 mM) treatmnent after 15 days showing extreme stunting of adventitious roots, and black sulphide staining of scale leaves and rhizome; laterals < 0.1 mm-in length; in other examples lateral roots grew to ca. 3 mm (x 1). (c) acetic acid (1_67 mM) treatment after 15 days showing stunted adventitious roots; tips of most adventitious and laterals roots were dead; buds growing at this stage but subsequently died. In other examples lateral root growth was even more stunted, < 0.5 mm. (x 0.75). (d) sulphide (1.4 mM) treatmnent after 28 days: L.S. rhizome nod e and base of dead enlarged bud with callus blocking the rhizome nodal diaphragm and the basal diaphragm of the bud; note also, callus in cortex at root-shoot junction and rhizome-shoot junction. (x 3.2). (e) sulphide (1.4 mM) treatment after 30 days: LS fully blocked metaxylem vessels in nodal region of rhizome. (x 70).
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions

VI J. Armstrong et al.: Phytotoxins in the Phragmites die-back
Plate 4. Phytotoxin treatments involving Phragmites rhizomes. (a) acetic acid (1.67 mM) treatment after 30 days: TS mid-length of adventitious root with lateral root "imprisoned" in, and growing down through, the cortex. (x 80). (b) acetic acid (1.67 mM) treatment after 30 days: TS root at root-rhizome junction with lig- nified strands of cell walls and callus totally blocking the cortical aerenchyma, and lignified tyloses/blockages within the protoxylem and metaxylem elements. (x 52). (c) acetic acid (1.67 mM) treatment after 30 days: TS mid region of adventitious root showing lateral (emergent part having rotted) with heavy lignification in outer layers, and in adjoining adventitious root cortex. (x 56). (d) sulphide (1.4 mM) treatment after 30 days: TS root-rhizome junction with cortical aerenchyma totally occluded by partially lignified callus; vascular ele- ments also blocked as in (b). (x 200). Hand-cut sections stained with phloroglucinol and concentrated hydrochloric acid to show lignification (red.). TS = transverse section.
This content downloaded from 195.34.79.176 on Sun, 15 Jun 2014 16:58:26 PMAll use subject to JSTOR Terms and Conditions