activation of internucleosomal dna cleavage in human cem ... · viability were estimated by trypan...
TRANSCRIPT

THE JOURNAL OF BUXOGICAL CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 28, Issue of October 5, pp. 17323-17333.1990 Printed in U.S. A.
Activation of Internucleosomal DNA Cleavage in Human CEM Lymphocytes by Glucocorticoid and Novobiocin EVIDENCE FOR A NON-Ca2’-REQUIRING MECHANISM(S)*
(Received for publication, March 9, 1990)
Emad S. Alnemri and Gerald LitwackS From the FeLs Institute for Cancer Research and Molecular Biology and the Department of Biochemistry, Temple University School of Medicine, Philadelphia, Pennsylvania 19140
Internucleosomal DNA cleavage is the key molecular event of the cytolytic phase of glucocorticoid-induced lymphocytolysis. We find that novobiocin, the topoi- somerase II inhibitor, is a potent inducer of in vivo internucleosomal DNA cleavage in human CEM lym- phocytes. This in vivo effect is very rapid, time- and dose-dependent, requires cellular integrity, and does not require de novo protein synthesis. Recently our data (Alnemri, E. S., and Litwack, G. (1989) J. Biol. Chem. 264, 4104-4111) suggested that activation of DNA cleavage in CEM-C7 lymphocytes by glucocorti- coids is independent of calcium uptake. Similarly, the novobiocin effect is also independent of calcium uptake and does not occur in isolated CEM nuclei or in CEM cells treated previously with the divalent cation iono- phore A23187. Internucleosomal DNA cleavage in- duced by novobiocin or glucocorticoid generates blunt- ended double-stranded DNA fragments possessing 3’- hydroxyls and 5’-phosphates. As demonstrated by gel retardation analysis and DNase I footprinting, novo- biocin causes the disruption and unfolding of an in vitro reconstituted mononucleosome so that it becomes more susceptible to DNase I cleavage. Our data suggest that 1) novobiocin rapid activation of internucleosomal DNA cleavage and chromatin changes in CEM lympho- cytes are molecular features of apoptosis or pro- grammed cell death. 2) CEM lymphocytes apparently do not express a Ca’+-dependent endonuclease. 3) The mechanism(s) of glucocorticoid or novobiocin-induced DNA cleavage in CEM lymphocytes involves activation of a constitutive non Ca’+-dependent endonuclease. We propose that the majority of nuclear chromatin is main- tained in a highly compact and charge-neutralized state and that disruption of this highly ordered struc- ture, directly by novobiocin or indirectly by glucocor- ticoid, may lead to the exposure and unmasking of internucleosomal linker DNA regions which are sub- strates for a constitutive non-Ca’+-dependent endonu- clease.
Chromatin cleavage at internucleosomal sites represents
* This work was supported by Research Grant DK 13531 and by Core Grant CA 12227 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.&C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed: Fels Inst. for Cancer Research and Molecular Biology, Temple University School of Medicine, 3420 North Broad St., Philadelphia, PA 19140. Tel.: 215-221-4340; Fax: 215-221-7849.
the earliest hallmark of the nuclear changes in apoptosis or programmed cell death (1, 2). Apoptosis has been studied most extensively in glucocorticoid-treated immature thymo- cytes (2-5). It has also been observed in glucocorticoid-treated lymphocytes and thymus-derived lymphomas (2-10). This phenomenon is mediated by the glucocorticoid receptor (4, ll), apparently requires protein synthesis (2, 5, 12), and is characterized by a typical internucleosomal DNA cleavage which precedes cell lysis (l-5, 13). In recent years, several studies have focused on understanding the biochemical mech- anism of activation of chromatin cleavage in glucocorticoid- induced apoptosis (14-17). Recently we challenged the idea that glucocorticoid-induced lymphocytolysis is dependent on the induction of nucleases (14, 18). We have shown that glucocorticoid-induced DNA cleavage in rat thymocytes is the result of activation of a constitutive endogenous Ca’+-/Mg*+- dependent endonuclease (18). Support for our earlier conclu- sions comes from many studies which have shown that Ca2+ is involved in the activation of an endogenous endonuclease in isolated nuclei (5, 16, 19) and in several examples of apoptosis (5, 15, 17, 20-23). However, in human CEM lym- phocytes, apoptosis does not seem to involve Ca*+-mediated activation of DNA cleavage (18, 24).
The present study was undertaken to investigate the mech- anism of activation of internucleosomal DNA cleavage in human CEM lymphocytes. Since glucocorticoid-induced DNA cleavage in CEM lymphocytes may not require Ca*+ uptake (18), we propose that the process of activation of internucleo- somal DNA cleavage in these cells involves changes in the chromatin substrate to activate a constitutive endonucle- ase(s). To examine this hypothesis the topoisomerase II in- hibitor, novobiocin, was used to introduce changes in the chromatin structure of CEM cells. Novobiocin was selected because of its reported ability to disrupt chromatin structure through direct interaction with histones (25) and its topoisom- erase II inhibitory effects (26-29). We report here that the in uiuo changes in chromatin structure induced by novobiocin were of sufficient magnitude to allow for internucleosomal DNA cleavage to occur within 1 h of treatment of CEM lymphocytes. This DNA cleavage requires cellular integrity, is similar to that induced by glucocorticoids, and is independ- ent of Ca*+ uptake. Further evidence shows that novobiocin causes the disruption of an in vitro reconstituted mononu- cleosome. Our data suggest that novobiocin kills CEM cells through the mechanism of apoptosis. Moreover, changes in chromatin structure which activate DNA cleavage may be the internal signal initiating the process of apoptosis in T-cell derived lymphocytes.
17323
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

17324 Glucocorticoid- and Novobiocin-induced Apoptosis
EXPERIMENTAL PROCEDURES
Materials-9u-Fluoro-Ilg,l6a,l7,21-tetra-ol-pregna-l,4-diene-3, 20.dione acetonide (triamcinolone acetonide, TA),’ ATP, D-sucrose (grade I), Hepes, Tris, Na,EDTA, EGTA, ethidium bromide, phenyl- methylsulfonyl fluoride (PMSF), agarose, cycloheximide, proteinase K, glutamine, and fetal calf serum were purchased from Sigma. RPM1 1$4b and t&an blue were obtained from GIBCO. DNase i (grade II), NAD+ (grade II), micrococcal nuclease, novobiocin, and divalent cation ionophore A23187 were obtained from Boehringer Mannheim. Low melting point agarose was obtained from Bethesda Research Laboratories. Molecular weight standards (X DNA-B&E11 digest) and BstNI restriction enzyme were obtained from New England Biolabs. [r-““P]ATP (4500 Cijmmol) was obtained from Du P&t-New Eng- land Nuclear. m-AMSA (NSC 249992) and camntothecin (NSC 94600) were obtained from ;he Drug Synthesis and Cdemistry Branch, Division of Cancer Treatment, National Cancer Institute. All other chemicals and enzymes were obtained from Fisher.
Cell Lines and Culture Conditions-The CEM cell line is a cloned human leukemic T-cell line derived from childhood acute lympho- blastic leukemia (30). CEM-C7 is a glucocorticoid-sensitive cell line which is killed by glucocorticoid (8). CEM-Cl is a glucocorticoid- resistant cell line which has an intact glucocorticoid receptor but lacks the lysis function (31, 32). Both cell lines were grown in RPM1 1640 medium supplemented with 5% fetal calf serum and 2 mM glutamine at 37 “C under 5% CO, in a humidified atmosphere.
Exponentially growing cells were used throughout all experiments at a concentration of 1 X 105 to 7 X @/ml. The cells were treated with the glucocorticoid analog TA at a concentration of 1 PM. In other experiments cells were treated with different concentrations of the top&somerase II inhibitor, novobiocin, or divalent cation iono- Dhore A23187. The urotein svnthesis inhibitor, cvcloheximide (10 ;M), was used in some expehments. After steroi&s or other drug treatment, cells were harvested by centrifugation at 800 x g for 5 min, washed with cold RPM1 1640, and recentrifuged again at 800 X g for 5 min.
Determination of Cell Number and Viability-Cell number and viability were estimated by trypan blue exclusion in a hemocytometer. The number and viability of CEM lymphocytes were periodically determined during steroid and drug treatments. Lymphocyte viability was greater than 95% at the beginning of each experiment.
Determination of Znternucleosomal DNA Cleavage-After novobio- tin, steroid, or other drug treatment, cells were lysed by addition of a DNA lysis buffer (0.2 M Tris-HCl, pH 8.0, 0.1 M Na2EDTA, 1% sodium dodecyl sulfate, and 100 pg/ml proteinase K), and the DNA was extracted by standard phenol/chloroform/isoamyl alcohol ex- traction procedures exactly as described previously (18). The isolated DNA was analyzed by electrophoresis on 1.8% agarose slab gels containing 0.2 pg/ml ethidium bromide. A X DNA B&E11 digest was used as molecular weight standards. The percent of chromatin cleav- age was determined from the densitometric scans of the photographic negatives as described previously (18). The percent of chromatin cleavage is defined as the sum of the integrated areas under the scan of DNA with molecular weights lower than 8.5 kilobase pairs (the largest DNA fragment of X DNA-B&E11 digest) expressed as a per- centage of total areas under the whole DNAscan.
Isolation of Nuclei-Fresh nuclei were prepared from either CEM lymphocytes’or rat thymocytes as described previously (33,34). Cells were suspended at a density of 1 x 10R/ml in N buffer (10 mM Tris- HCl, pH 7.5, 2.5 mM KCl, 2.5 mM MgCl,, 0.5 M sucrose, 0.5 mM PMSF), and incubated on ice for 10 min. Nuclei were partially purified by homogenization (five strokes) and then centrifugation at 800 x g for 10 min. The resulting crude nuclei were washed (twice) in N buffer and then pelleted by centrifugation at 800 x g for 10 min. The pellet was resuspended in DS buffer (50 mM Tris-HCl, pH 7.5, 5 mM MgCl,, 2.1 M sucrose, 0.5 mM PMSF), layered over a cushion of the same buffer, and centrifuged at 30,000 rpm in a Beckman SW-40 rotor for 30 min. The upper layer and the sucrose cushion were removed with an aspirator. The resulting pellet consisting of highly- purified nuclei was resuspended in the assay buffer (25 mM Hepes,
I The abbreviations and trivial names used are: triamcinolone acetonide (TA), 9~-fluoro-ll~,l6a,l7,21-tetra-ol-pregna-1,4-diene- 3.20-dione acetonide; Hepes, 4-(2.hydroxyethyl)-1 piperazineethane- sulfonic acid; EGTA, -[ethylenebis(oxyethylenenitrilo)]tetraacetic acid: PMSF. ohenvlmethvlsulfonvl fluoride: TA, triamcinolone acet- ,. ” ” onide; bp, base pairs; m-AMSA, 4’-(9.acridinylamino)methanesulfon- m-anisidide.
pH 7.0, 2 mM potassium phosphate, pH 7.0, 125 mM KCl, 4 mM WzCM.
In Vitro Nucleosome Reconstitution-The DNA fragments used for this experiment were a 239 base pairs (bp) H, (SmaI/DruI) fragment and a 181-bp H, (SmaI/BstNI) fragment derived from the 5’-flanking sequence of the human histone H1 gene. These fragments were obtained from the plasmid pFNC16 (35) (a gift from Dr. Gary Stein, Department of Cell Biology, University of Massachusetts Medical School) which contains the full-length coding and parts of the 3’ and 5’ noncoding flanking sequences of the human histone HI gene. The plasmid was first cut with SmaI and PuuII restriction enzymes and the resulting 460 bp 5’-flanking sequence was isolated by electropho- resis in a 1% low melting point agarose. The fragment was end labeled using [Y-~*P]ATP and T4 polynucleotide kinase and then cut with either DraI to generate the H, (SmaI/DraI)-labeled fragment or BstNI to generate the H, (SmaI/BstNI)-labeled fragment. These fragments were isolated by electrophoresis in a 1.2% low melting point agarose.
Mononucleosomes were reconstituted by a high salt exchange method as described previously (36, 37) using HI-depleted donor chromatin as a histone source. Donor chromatin was prepared from either human CEM lymphocyte nuclei or rat liver nuclei by the same method described by Lutter (38). Donor chromatin was stored at -20 “C in a buffer containing 5 mM Tris-HCl, pH 7.5, 0.5 mM Na,EDTA, 0.6 M NaCl, 0.25 mM PMSF, and 50% glycerol. Before nucleosome reconstitution the NaCl concentration of the donor chro- matin was made 1.6 M with 5 M NaCl. End-labeled restriction frag- ments in 10 ~1 of water were mixed with 0.1 ml of this donor chromatin (25 OD/ml), and the mixture was incubated at 37 “C for 20 min. The mixture was diluted with a chromatin dilution buffer (15 mM Tris- HCl. DH 7.5. 0.2 mM Na?EDTA. 0.2 mM PMSF) to NaCl concentra- tion of 1 M and incubated again & 37 “C for 20 min. The mixture was then kept at room temperature and stepwise dilution continued for several hours until the NaCl concentration reached a final concentra- tion of 80 mM. The reconstituted mononucleosomes were analyzed by gel retardation analysis in a low ionic strength 5% nondenaturing polyacrylamide gels.
Determination of the Effect of Nouobiocin on the Structure of the Reconsfituted Nucleosome-After novobiocin treatment, the reconsti- tuted nucleosomes were analyzed by gel retardation as described above and by DNase I footprinting analysis. For DNase I footprinting, reconstituted mononucleosome or naked DNA samples (30,000 cpm/ sample) were digested with DNase I (20 rig/sample) in a buffer containing 15 rns Tris-HCl, pH 7.5, 2 mM MgCl*, &d 2 mM CaC12 at 25 “C for 45 s. The DNA was then analvzed on denaturing 6% polyacrylamide-urea sequencing gels.
RESULTS
Novobiocin Activates Internucleosomal DNA Cleavage of Human Lymphocyte Genome in Vivo-To examine the effect of novobiocin on chromatin structure of human lymphocytic cell lines, CEM-C7 cells were treated with O-1200 pg/ml novobiocin for 3 h. DNA was isolated and analyzed by agarose gel electrophoresis. Fig. lA shows that novobiocin was able to activate internucleosomal DNA cleavage in CEM-C7 cell line in a dose-dependent manner. DNA isolated from CEM-C7 cells treated with 200-800 pg/ml (lanes 4-7) of novobiocin showed a characteristic nucleosome ladder pattern. This pat- tern consists of low molecular weight DNA fragments which are multiples of 180 base pairs. The novobiocin-induced DNA cleavage pattern is strongly similar to that obtained after glucocorticoid treatment of rat thymocytes or human CEM- C7 lymphocytes (18). The maximal effect was obtained using 400 pg/ml of novobiocin. Using novobiocin concentrations ranging from 0 to 400 pg/ml resulted in a dose-dependent increase in the amount of DNA cleavage (lanes 2-5), whereas novobiocin concentrations higher than 400 pg/ml showed a dose-dependent decrease in the amount of internucleosomal DNA cleavage (lanes 5-9). DNA isolated from cells incubated for 3 h without novobiocin (lane 2) or with 100 or 1000-1200 Kg/ml of novobiocin (lanes 3, 8 and 9) was virtually unde- graded high molecular weight DNA. The dose-dependent de- crease in the amount of internucleosomal DNA cleavage ob-
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Glucocorticoid- and Novobiocin-induced Apoptosis 17325
A 6
KB 123456789 12 34
8.45- 3.67- 1.37-
0.70-
0.22-
a FIG. 1. A, dose-dependence of novobiocin-induced internucleoso-
ma1 DNA cleavage of CEMC7 lymphocyte DNA. CEM-C7 cells were incubated in complete RPM1 1640 for 3 h with or without novobiocin. The DNA was then isolated and analyzed on a 1.8% agarose gel. The following novobiocin concentrations were used: 0 fig/ml (lane 2), 100 pg/ml (lane 3), 200 pg/ml (lane 4), 400 pg/ml (lane 5), 600 fig/ml (lane 6), 800 rg/ml (lane 7), 1000 pg/ml (lane 8), or 1200 pg/ml (lane 9). A X DNA-BstEII digest was used as molecular weight markers (lane I). KR, kilobase pair. B, effect of cycloheximide on novobiocin-induced internucleosomal DNA cleavage. CEM-C7 cells were incubated in complete RPM1 1640 for 4 h with no addition (lane I), 10 mM cycloheximide (lane 2), 200 pg/ml novobiocin and 10 mM cyclohexi- mide (lane 3), or 200 pg/ml novobiocin (lane 4). The DNA was isolated and analyzed as described above.
tained with high concentrations of novobiocin could be due to the extensive cell lysis and membrane damage observed after treatment with high concentrations of the drug.
To determine whether novobiocin-induced internucleoso- ma1 DNA cleavage is dependent upon de nouo protein biosyn- thesis, CEM-C7 lymphocytes were treated with 200 pg/ml novobiocin, 200 pg/ml novobiocin plus 10 ~.LM cycloheximide, or 10 pM cycloheximide. Untreated CEM-C7 cells were used as a control. DNA was isolated and analyzed by agarose gel electrophoresis (Fig. 1B). DNA isolated from untreated con- trol cells or cycloheximide-treated cells (lanes 1 and 2) had no detectable cleavage, whereas DNA isolated from cells treated with novobiocin in the presence or absence of cyclo- heximide (lanes 3 and 4, respectively) showed a characteristic internucleosomal DNA cleavage. The absence of any effect of cycloheximide on novobiocin-induced DNA cleavage suggests that new protein synthesis is not required for the novobiocin effect.
Comparison of the Effects of TA and Novobiocin on Human Lymphocyte Genome-Glucocorticoids have been shown to inhibit proliferation and cause lysis of the human lympho- blastic leukemia cell line CEM-C7 (8). In contrast, the human lymphoblastic leukemia cell line CEM-Cl is resistant to the lethal action of glucocorticoids. Interestingly, both cell lines have functional glucocorticoid receptors (31,32); however, the basis of the glucocorticoid resistance of CEM-Cl cell line is unknown.
Fig. 2 shows time-course studies of the growth of CEM-C7 (panel A) and CEM-Cl (panel B) after treatment with the glucocorticoid agonist TA or novobiocin. It is evident from the growth curves that CEM-C7 is sensitive to TA. The growth rate of TA-treated CEM-C7 cells was slightly sup- pressed up to 32-46 h. After this time, cell viability decreased sharply, whereas CEM-Cl cells continued to grow in the presence of 1 PM TA without any effect on cell viability. Both cell lines were sensitive to the lethal action of novobiocin. Treatment of CEM-C7 and CEM-Cl (Fig. 2, A and B) with 200 rg/ml of novobiocin caused a time course-dependent decrease in cell viability. Surprisingly, CEM-C7 was found to be more sensitive than CEM-Cl to the lethal action of novo-
10’
104
Time (hr) Time (hr)
FIG. 2. Effect of TA and novobiocin on the growth and viabilitv of CEM-C7 and CEM-Cl human lvmuhocvtes. CEM- C7 (panel A) and CEM-Cl (B) cells were seeded in complete RPM1 1640 at the indicated concentrations. The cells were then treated with 1 pM TA or 200 rg/ml novobiocin. At the indicated times, viable cell counts were determined by the trypan blue exclusion technique.
biocin. By 46 h, novobiocin caused about 8.3-fold decrease in cell viability of CEM-C7 compared with approximately 1.3- fold decrease in CEM-Cl viability over the same period of time. Electron microscopic examination of cells treated with TA or novobiocin showed ultrastructural changes of chroma- tin condensation characteristic of apoptosis (data not shown). The observed chromatin condensation is the result of inter- nucleosomal DNA cleavage.
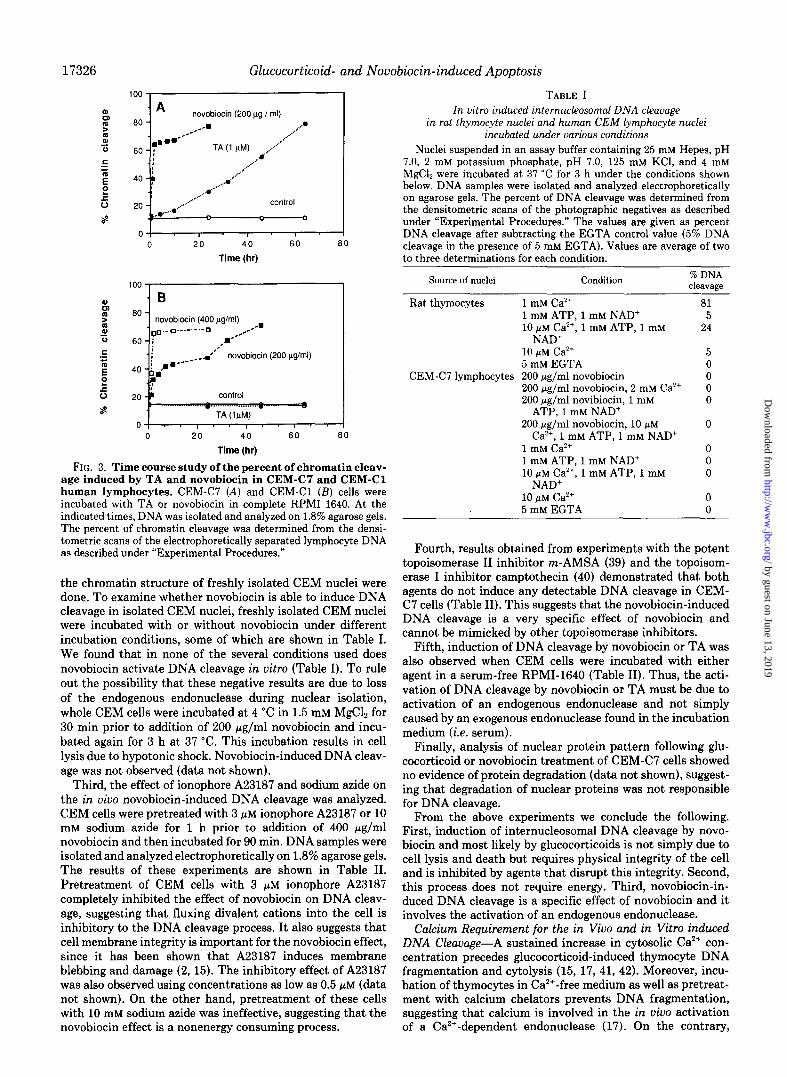
To determine whether the activation of internucleosomal DNA cleavage by novobiocin or TA requires the physical integrity of the cell or simply is a result of cellular death and disintegration, several experiments were performed. First, Fig. 3 shows a time course analysis of the in vivo effect of TA and novobiocin on DNA fragmentation of both CEMC7 and CEM-Cl cell lines. DNA samples from TA- and novobiocin- treated lymphocytes were analyzed by agarose gel electropho- resis. The percent of chromatin cleavage was determined from the densitometric scan of the electrophoretically separated lymphocyte DNA. As shown in Fig. 3, A and B, the percent of chromatin cleavage indicates that 30% of the DNA was degraded 24 h after TA treatment of CEM-C7. A time course increase in the percent of chromatin cleavage was observed with longer incubation of CEM-C7 with TA. At 32-46 h almost 37-55% of the lymphocyte DNA was degraded to internucleosomal size fragments; however, the viability of the lymphocytes during this time was relatively high, 95% at 32 h and 86% at 46 h (see also Fig. 2). Novobiocin-induced internucleosomal DNA cleavage is a rapid event occurring within 1 h of novobiocin treatment of human CEM lympho- cytes. More than 65% DNA cleavage was observed in CEM- C7 in less than 5 h (Fig. 3A), yet the cell viability exceeded 90% during this time (see also Fig. 2). CEM-Cl, as mentioned earlier, was more resistant to the drug than CEM-C7. How- ever, within less than 10 h, more than 40% of CEM-Cl DNA was cleaved (Fig. 3B), and the viability was greater than 90% during this time. Treatment of CEM-Cl with 400 pg/ml novobiocin produced a similar time course increase in percent of DNA cleavage as that observed when CEM-C7 was treated with 200 pg/ml novobiocin. Since cellular viability was deter- mined by trypan blue exclusion technique, the viability values mentioned above may not reflect the actual capacity of cells to proliferate. However, these values definitely reflect the cellular membrane integrity of those cells under investigation.
Second, in vitro experiments on the effect of novobiocin on
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from
Glucocorticoid- and Novobiocin-induced Apoptosis
100 , I .A novobiocin (200 pg / ml)
80 - /-- . . . ..a
,.a*-- .A ,./
60 - ; TA (1 NM) /” . ..d”
,...”
40 -c ,*..”
_..’
.A-..‘* 8
,,,,/.. 20 - /... control
&.A 0
0, . I . I . I I 0 20 40 60 80
Time (hr)
I " "
. B 80 -
novobiocin (400 pg/ml) ‘oo.-g ._.-.-.- q /--
.
60 - i /
4o -6i=‘-----
*“’ novobiocin (200 pg/ml) -I
0 20 40 60 80
Time (hr)
FIG. 3. Time course study of the percent of chromatin cleav- age induced by TA and novobiocin in CEM-C7 and CEM-Cl human lymphocytes. CEM-C7 (A) and CEM-Cl (B) cells were incubated with TA or novobiocin in complete RPM1 1640. At the indicated times, DNA was isolated and analyzed on 1.8% agarose gels. The percent of chromatin cleavage was determined from the densi- tometric scans of the electrophoretically separated lymphocyte DNA as described under “Experimental Procedures.”
the chromatin structure of freshly isolated CEM nuclei were done. To examine whether novohiocin is able to induce DNA cleavage in isolated CEM nuclei, freshly isolated CEM nuclei were incubated with or without novobiocin under different incubation conditions, some of which are shown in Table I. We found that in none of the several conditions used does novobiocin activate DNA cleavage in vitro (Table I). To rule out the possibility that these negative results are due to loss of the endogenous endonuclease during nuclear isolation, whole CEM cells were incubated at 4 “C in 1.5 mM MgCl, for 30 min prior to addition of 200 pg/ml novobiocin and incu- bated again for 3 h at 37 “C. This incubation results in cell lysis due to hypotonic shock. Novobiocin-induced DNA cleav- age was not observed (data not shown).
Third, the effect of ionophore A23187 and sodium azide on the in vivo novobiocin-induced DNA cleavage was analyzed. CEM cells were pretreated with 3 nM ionophore A23187 or 10 mM sodium azide for 1 h prior to addition of 400 @g/ml novobiocin and then incubated for 90 min. DNA samples were isolated and analyzed electrophoretically on 1.8% agarose gels. The results of these experiments are shown in Table II. Pretreatment of CEM cells with 3 PM ionophore A23187 completely inhibited the effect of novobiocin on DNA cleav- age, suggesting that fluxing divalent cations into the cell is inhibitory to the DNA cleavage process. It also suggests that cell membrane integrity is important for the novobiocin effect, since it has been shown that A23187 induces membrane blebbing and damage (2, 15). The inhibitory effect of A23187 was also observed using concentrations as low as 0.5 pM (data not shown). On the other hand, pretreatment of these cells with 10 mM sodium azide was ineffective, suggesting that the novobiocin effect is a nonenergy consuming process.
TABLE I In uitro induced internucleosomal DNA cleavage
in rat thymocyte nuclei and human CEM lymphocyte nuclei incubated under various conditions
Nuclei suspended in an assay buffer containing 25 mM Hepes, pH 7.0, 2 mM potassium phosphate, pH 7.0, 125 mM KCl, and 4 mM MgCl, were incubated at 37 “C for 3 h under the conditions shown below. DNA samples were isolated and analyzed electrophoretically on agarose gels. The percent of DNA cleavage was determined from the densitometric scans of the photographic negatives as described under “Experimental Procedures.” The values are given as percent DNA cleavage after subtracting the EGTA control value (5% DNA cleavage in the presence of 5 mM EGTA). Values are average of two to three determinations for each condition.
Source of nuclei Condition % DNA ClawdEe
Rat thymocytes
CEMC7 lymphocytes
1 mM Ca’+ 1 mM ATP, 1 mM NAD+ 10 FM Ca*+, 1 mM ATP, 1 DIM
NAD+ 10 yM Ca*+ 5 mM EGTA 200 rg/ml novobiocin 200 rg/ml novobiocin, 2 mM ca*+ 200 pg/ml novibiocin, 1 mM
ATP, 1 mM NAD’ 200 fig/ml novobiocin, 10 pM
Ca’+, 1 mM ATP, 1 mM NAD+ 1 mM Ca*+ 1 UIM ATP, 1 mM NAD+ 10 pM Ca”, 1 mM ATP, 1 mM
NAD+ 10 fiM Ca2+ 5 mM EGTA
81 5
24
Fourth, results obtained from experiments with the potent topoisomerase II inhibitor m-AMSA (39) and the topoisom- erase I inhibitor camptothecin (40) demonstrated that both agents do not induce any detectable DNA cleavage in CEM- C7 cells (Table II). This suggests that the novobiocin-induced DNA cleavage is a very specific effect of novobiocin and cannot be mimicked by other topoisomerase inhibitors.
Fifth, induction of DNA cleavage by novobiocin or TA was also observed when CEM cells were incubated with either agent in a serum-free RPMI-1640 (Table II). Thus, the acti- vation of DNA cleavage by novobiocin or TA must be due to activation of an endogenous endonuclease and not simply caused by an exogenous endonuclease found in the incubation medium (i.e. serum).
Finally, analysis of nuclear protein pattern following glu- cocorticoid or novobiocin treatment of CEM-C7 cells showed no evidence of protein degradation (data not shown), suggest- ing that degradation of nuclear proteins was not responsible for DNA cleavage.
From the above experiments we conclude the following. First, induction of internucleosomal DNA cleavage by novo- biocin and most likely by glucocorticoids is not simply due to cell lysis and death but requires physical integrity of the cell and is inhibited by agents that disrupt this integrity. Second, this process does not require energy. Third, novobiocin-in- duced DNA cleavage is a specific effect of novobiocin and it involves the activation of an endogenous endonuclease.
Calcium Requirement for the in viva and in Vitro induced DNA Cleavage-A sustained increase in cytosolic Ca2+ con- centration precedes glucocorticoid-induced thymocyte DNA fragmentation and cytolysis (15, 17, 41, 42). Moreover, incu- bation of thymocytes in Ca’+-free medium as well as pretreat- ment with calcium chelators prevents DNA fragmentation, suggesting that calcium is involved in the in uiuo activation of a Ca’+-dependent endonuclease (17). On the contrary,
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Glucocorticoid- and Nouobiocin-induced Apoptosis 17327
TABLE II In vivo induced internucleosomal DNA cleavage in human CEM lymphocytes incubated under various conditions
CEM-C7 lymphocytes were incubated under the conditions shown below. DNA samples were isolated and analyzed on 1.8% agarose gels. All incubations were in RPM1 1640 containing 5% fetal calf serum unless otherwise indicated.
Pretreatment condition Incubation condition DNA cleavage None No treatment None 400 pg/ml novobiocin + 10 mM NaN.I (1 h)” 10 mM NaN:, (1.5 h) - 3 /IM A23187 (1 h) 3 MM A23187 (1.5 h) - 10 mM NaN, (1 h) 10 mM NaNzj, 400 rg/ml novobiocin (1.5 h) + 3 PM A23187 (1 h) 3 FM A23187, 400 pg/ml novobiocin (1.5 h) - None 100 rg/ml mAMSA (4 h) - None 20 PM camptothecin (4 h) - None Serum-free medium (48 h) - None Serum-free medium, 1 PM TA (48 h) + None Serum-free medium, 400 fig/ml novobiocin (4 h) + 5 mM EGTA (0.5 h) 5 mM EGTA (5 h) - 5 mM EGTA (0.5 h) 5 mM EGTA, 200 pg/ml novobiocin (5 h) +
’ The length of incubation period in hours.
glucocorticoid-induced DNA cleavage and cytolysis in CEM- C7 lymphocytes are not dependent on calcium uptake (18). To examine whether the effect of novobiocin on CEM lym- phocytes is dependent on the presence of calcium in the incubation medium, CEM lymphocytes were incubated with novobiocin in the presence or absence of 5 mM EGTA. DNA was isolated and analyzed as described under “Experimental Procedures.” Table II shows that regardless of the presence or absence of EGTA, novobiocin was able to cause internu- cleosomal DNA cleavage in CEM-C7 lymphocytes. Similar results were obtained with CEM-Cl lymphocytes (data not shown). No DNA cleavage was observed in untreated or EGTA-treated control cells. Thus, like the glucocorticoid effect, the novobiocin effect on DNA cleavage is not depend- ent on calcium uptake.
In vitro studies with isolated nuclei from different tissues have shown that rapid autodigestion of DNA to internucleo- somal fragments can occur when nuclei are incubated in a buffer containing millimolar concentrations of Ca*+ and Mg’ (5, 16 18, 19). Incubation of isolated rat liver nuclei with submicromolar Ca*+ concentration, in the presence of ATP and NAD’, can also result in extensive DNA fragmentation (16). Thus, we decided to compare the Ca’+ dependence of DNA fragmentation in rat thymocyte nuclei and human lym- phocyte nuclei incubated under various in vitro conditions. Fresh rat thymocyte nuclei and CEM-C7 nuclei isolated and treated under identical conditions were incubated with EGTA or various concentrations of Ca’+. DNA was isolated and analyzed as described under “Experimental Procedures.” Fig. 4, A and B, show that incubation of rat thymocyte nuclei with various Ca2+ concentrations resulted in a dose-dependent activation of internucleosomal DNA cleavage. Maximum ac- tivation was observed at 1 mM Ca2+ concentration. Interest- ingly, nuclei incubated with 10 mM Ca*+ routinely showed 15% less DNA cleavage than those incubated with 1 mM Ca”‘, reflecting a requirement for an optimal Ca*+ concentration. Calcium concentrations higher than 1 mM have been shown to cause a dose-dependent inhibition of endogenous DNA autodigestion (19). Unlike thymocyte nuclei, incubation of CEM-C7 nuclei with various Ca*+ concentrations did not activate internucleosomal DNA cleavage (Fig. 4). Incubation of thymocyte nuclei with 10 PM Ca*+ resulted in a very small amount of DNA cleavage (Table I). However, addition of 1 mM NAD’ and 1 mM ATP to thymocyte nuclei incubated with 10 PM Ca2+ caused substantial activation of DNA cleav- age above that seen with either NAD+ plus ATP or 10 PM
KB 123456789lo1112
Thymocyte nuclei
‘:\,_,,,,,,,, / FFrfiYcyte;cIei . . . . . . . . . . . ..__ b __....__._,._... * .,._........... I.. . ...““““..
EGTA f3 -5 -4 -3 2 -‘l
log [M] Calcium
FIG. 4. Dose-dependence of in vitro calcium-induced inter- nucleosomal DNA cleavage in isolated nuclei. Nuclei from rat thymocytes or human CEM-C7 lymphocytes were incubated at 37 “C with EGTA or various concentrations of Ca’+ for 3 h in the same assay buffer mentioned in Table I legend. In A, DNA was isolated and analyzed on 1.8% agarose gel. Lanes 1-6, DNA isolated from rat thymocyte nuclei; lanes 7-12, DNA isolated from human CEM-C7 lymphocyte nuclei. Lanes 1 and 7, 5 mM EGTA; lanes 2 and 8, 1 PM Ca’+; lanes 3 and 9, 10 PM Ca’+; lanes 4 and 10, 100 pM Cay+; lanes 5 and 11, 1 mM Ca*+; lanes 6 and 12, 10 mM Ca’+. KB, kilobase pair. In B, the percent of chromatin cleavage was determined from the den- sitometric scan of the photographic negative of the gel shown in A as described under “Experimental Procedures.”
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

17328 Glucocorticoid- and Novobiocin-induced Apoptosis
Ca’+ alone. These results are consistent with the recent tind- ings that rat liver and thymocyte nuclei possess a calmodulin- dependent nuclear Ca2+ uptake system which is probably involved in endogenous endonuclease activation (16). How- ever, no detectable DNA cleavage was observed in CEM-C7 nuclei incubated with different concentrations of Ca*+ and Ca’+ plus ATP and NAD’. These data indicate that CEM lymphocytes do not depend on a Ca’+-dependent endonucle- ase to activate the in vivo DNA cleavage process observed after glucocorticoid or novobiocin treatment. Instead, we sug- gest that CEM lymphocytes express a non-Ca’+-dependent endonuclease which is capable of cleaving chromatin at inter- nucleosomal sites in vivo but not in vitro.
Glucocorticoid and Novobiocin Induce Blunt-ended Double- stranded DNA Cleavage-Villeponteau et al. (43) demon- strated that novobiocin induces blunt-ended double-stranded DNA cleavage in immature chicken red blood cells. To further characterize the endogenous endonuclease(s) responsible for the glucocorticoid or novobiocin-induced internucleosomal DNA cleavage in human CEM lymphocytes or rat thymocytes, we performed a DNA end analysis similar to that described by Villeponteau et al. (43). Mononucleosomal DNA fragments (140-200 bp) were isolated from glucocorticoid-treated CEM- C7 lymphocytes or rat thymocytes (Fig. 5A, lanes 2 and 3) and from novobiocin-treated CEM-C7 cells (lane 1). As a control, mononucleosomal DNA was also isolated from CEM- C7 nuclei digested with micrococcal nuclease (lane 4). Inter- estingly, we observed that the mononucleosomes generated by glucocorticoid in CEM-C7 cells were further cleaved by the endogenous endonuclease to generate DNA fragments shorter than 140 bp (he 2). No further cleavage was observed in novobiocin-treated CEM-C7 cells or TA-treated rat thy- mocytes (lanes 1 and 3).
The isolated mononucleosomal DNA fragments were ana- lyzed for the presence of 5’- or 3’-protruding ends. The DNA fragments were first 5’-32P-end-labeled with [T-~*P]ATP and polynucleotide kinase and then incubated with or without T4 DNA polymerase which fills the 5’-protruding ends and trims the 3’-protruding ends. Plasmid DNA restriction fragments (pFNC16, see “Experimental Procedures”) generated by digestion with either HpaII restriction endonuclease which generates 2-base 5’-protruding ends or HaeIII restriction en- zyme which generates blunt ends were used as additional controls for the Tq DNA polymerase reaction. After Tq DNA polymerase treatment, all samples were analyzed on a urea- polyacrylamide sequencing gel.
Tq DNA polymerase treatment of mononucleosomal DNA fragments isolated from glucocorticoid-treated CEM-C7 lym- phocytes or rat thymocytes (Fig. 5B, lanes 7-10 and lanes 13 and 14) did not cause any detectable shift-up or shift-down in their mobility. Similarly, no detectable change in the mo- bility of novobiocin-induced DNA fragments was observed (Fig. 5 lanes 5 and 6). As expected, a slight shift-up in the mobility of micrococcal nuclease-induced DNA fragments was observed (lanes 11 and 12). This mobility shift-up was small but reproducible; nevertheless, it was expected since micro- coccal nuclease generates only 2-base 5’-protruding ends. HpaII restriction fragments showed a clear shift-up in their mobility following Tq DNA polymerase treatment (lanes 1 and 2). No detectable shift-up was observed with HaeIII restriction fragments (lanes 3 and 4). Thus, glucocorticoid and novobiocin induce blunt-ended DNA cleavage as judged by the absence of any shift observed with their induced DNA fragments.
To analyze the 3’-ends for the presence of 3’-phosphate or hydroxyl groups, the mononucleosomal DNA fragments de-
scribed in Fig. 5A as well as the plasmid restriction fragments were incubated with dCTP in the presence (+) or absence (-) of terminal deoxynucleotidyl transferase. The DNA samples were 32P-end-labeled and then analyzed on urea-polyacryl- amide sequencing gel. Fig. 5C demonstrates that, except for the micrococcal nuclease-generated DNA fragments (which possess 3’-phosphate groups), all mononucleosomal DNA fragments and the restriction fragments were efficiently tailed. These results demonstrate clearly that the glucocorti- coid- and the novobiocin-induced DNA fragments possess 3’- hydroxyl groups.
For 5’-phosphate or hydroxyl end analysis, the DNA frag- ments described above were incubated with or without calf intestinal alkaline phosphatase. The DNA samples were 32P- end-labeled and then analyzed on a urea-polyacrylamide se- quencing gel. Fig. 50 shows that without prior treatment with alkaline phosphatase (-) the glucocorticoid- and novobiocin- generated DNA fragments as well as the plasmid restriction fragments were not efficiently labeled (lanes 1, 3, 5, 7, and 9). On the other hand, micrococcal nuclease-generated DNA fragments (which possess 5’-hydroxyl groups) were highly labeled regardless of prior treatment with or without alkaline phosphatase (lanes 11 and 12). From the above data we conclude that glucocorticoid and novobiocin induce double- stranded blunt-ended DNA fragments possessing 5’-phos- phates and 3’-hydroxyls.
Novobiocin Disrupts in Vitro Reconstituted Mononucleo- some-To elucidate the mechanism of novobiocin-induced DNA cleavage in human CEM lymphocytes, the effect of novobiocin on the structure and nuclease sensitivity of an in vitro reconstituted mononucleosome was investigated. Two “‘P-end-labeled DNA fragments, 239- and 181-bp histone H1 DNA fragments, were used to reconstitute mononucleosomes by a high salt exchange method (see “Experimental Proce- dures”). R-depleted donor chromatin was mixed with the 32P- labeled DNA fragments at 1.6 M NaCl followed by stepwise dilution to 80 mM NaCl. Electrophoretic analysis of the reconstituted material on native polyacrylamide gels shows that more than 95% of the naked DNA was converted to mononucleosomes by this procedure (Fig. 6, lanes 2 and 3). As expected, more than one species of nucleosomes were obtained with the 239-bp fragment. This is because DNA fragments longer than 180 bp could direct the histone octo- mers to occupy multiple specific positions along the DNA fragment (36). However, reconstitution of the 181-bp frag- ment with core histones generated only one particle (data not shown). The migration of the reconstituted mononucleosomes was also compared with that of native mononucleosomes derived by extensive digestion of chromatin with micrococcal endonuclease on native gels. The reconstituted mononucleo- somes comigrate with the native mononucleosomes detected by ethidium bromide staining (data not shown).
The effect of novobiocin on the structure of the reconsti- tuted mononucleosome was examined. Mononucleosomes re- constituted with the 239-bp DNA fragment and donor chro- matin from either human CEM nuclei (Fig. 6A) or rat liver nuclei (Fig. 6B) were incubated with various concentrations of novobiocin. Following incubation, the reconstituted mate- rial was analyzed by gel retardation analysis on native poly- acrylamide gels. As shown in Fig. 6 novobiocin has a substan- tial effect on the structure of the mononucleosome. Incubation of mononucleosomes with novobiocin resulted in disruption and unfolding of the mononucleosomes in a dose-dependent fashion. This can be seen as more of the slow migrating mononucleosome band is converted to the fast migrating naked DNA band with increasing novobiocin concentrations
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Glucocorticoid- and Nouobiocin-induced Apoptosis 17329
FIG. 5. Glucocorticoid and novo- biocin induce blunt-end cleavage. A, low melting point agarose gel electropho- resis of DNA samples isolated from no- vobiocin-treated CEM-C7 lymphocytes (lane I), TA-treated CEM-C7 lympho- cytes (lane 2), TA-treated rat thymo- cytes (lane 3), and micrococcal nuclease- treated CEM-C7 nuclei (lane 4). B, pro- truding ends analysis. Mononucleosomal DNA was isolated from the low melting point agarose described in A and then .“P-end-labeled. The labeled DNA sam- ples were incubated with (+) or without (-) T, DNA polymerase (T,-DP). The DNA samples were analyzed on a 6% polyacrylamide-urea sequencing gel. Lanes 1 and 2, &a11 restriction digest of the plasmid pFNC1G; lanes 3 and 4, Hue111 digest of the plasmid pFNC16; lanes 5 and 6, mononucleosomal DNA from novobiocin-treated CEM-C7 cells; lanes 7 and 8, mononucleosomal DNA from TA-treated CEM-C7 cells; lanes 9 and 10, mononucleosomal DNA from TA-treated rat thymocytes; lanes 11 and 12, mononucleosomal DNA from micro- coccal nuclease-digested CEM-C7 nu- clei; lanes 13 and 14 are an overexposure of lanes 7 and 8. Canalysis of 3’-ends. The DNA samples described in A and B were incubated with dCTP in the pres- ence (+) or absence (-) of terminal deox- ynucleotidyl transferase (TDT) and then “‘P-end-labeled. The labeled DNA was analyzed on a 6% polyacrylamide-urea sequencing gel. Lane 1, “‘P-end-labeled X DNA-B&E11 digest; lanes 2 and 3, Hue111 digest of pFNC1G; lanes 4 and 5, HpaII digest of pFNC1G; lanes 6-13 are, respectively, the same as lanes 5-12 in B. D, analysis of the 5’-ends. The DNA samples described in A and B were in- cubated in the presence (+) or absence (-) of calf intestinal alkaline phospha- tase (CAP) and then “*P-end-labeled with [y-32P]ATP and polynucleotide ki- nase using the forward kinase reaction. Labeled DNA was then analyzed on 6% polyacrylamide-urea sequencing gel. Lanes l-12 are the same as lanes l-12 in B.
T4-DP -+-+-+ -+-+-+ -+
NUCLEOSOME DNA-
224.
TDT -+-+-+-+-+-+
224- -
CAP-+-+ -+-+-+-+
224-
D
(Fig. 6, A and B). Densitometric scanning of the autoradi- ographic x-ray film (Fig. 6C) shows that maximal disruption of mononucleosomes occurred at novobiocin concentrations of 1500 pg/ml. At this concentration of novobiocin about 80% of the mononucleosomes were converted to naked DNA. At concentrations even lower than those used to induce in vivo internucleosomal DNA cleavage, novobiocin was very effec- tive in causing nucleosome disruption. For example, at 50 pg/ ml novobiocin concentration, about 30% of mononucleosomes were converted to naked DNA. Identical results were obtained with the mononucleosomes reconstituted with the 181-bp DNA fragment (data not shown).
We next examined whether the novobiocin-induced unfold- ing of the mononucleosome would cause the DNA on the core
particle to become more sensitive to DNase I. Mononucleo- somes reconstituted with the 181-bp DNA fragment and donor chromatin from human CEM nuclei were incubated with various concentrations of novobiocin. Following the incuba- tion, the mononucleosomes were digested with DNase I and then analyzed on denaturing polyacrylamide gels. As can be seen in Fig. 7, the DNase I footprinting analysis reveals a distinct pattern of alternating nuclease sensitivity and pro- tection in untreated mononucleosomes (lanes 3 and 4) com- pared with naked DNA (lanes 1 and 2). Four distinct protec- tion areas labeled as Z, ZZ, ZZZ, and IV can be seen along the untreated mononucleosome DNA length. Novobiocin-induced unfolding of the nucleosome structure caused the DNA on the core particle to become more sensitive to cleavage by DNase
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Glucocorticoid- and Novobiocin-induced Apoptosis
123456789
MONONUCLEOSOME-
m t-4UICn c4cu
NAKED DNA-
B 123456789
MONONUCLEOSOME- &Ht?t!krl
NAKED DNA- c ,4rCllliYYYrn
C
80
60
mononucleosome
0 500 1000
Novobiocin [yg/ml] 1500
FIG. 6. Gel retardation analysis of the effect of novobiocin on the structure of reconstituted mononucleosomes. In A, mon- onucleosomes reconstituted with the 239-bp 32P-end-labeled DNA fragment and donor chromatin isolated from human CEM lympho- cyte nuclei were incubated at 4 “C with various concentrations of novobiocin for 5 h. The mononucleosomes were then analyzed by gel retardation analysis as described under “Experimental Procedures.” The following novobiocin concentrations were used: 0 rg/ml (lanes 2 and 3), 50 rg/ml (lane 4), 100 rg/ml (lane 5), 250 pg/ml (lane 6), 500 rg/ml (lane 7), 1000 pg/ml (lane 8), or 1500 pg/ml (lane 9). The 239- bp naked DNA fragment is included in lane 1. In B, same as in A except that the mononucleosomes were reconstituted with donor chromatin isolated from rat liver nuclei. In C, the x-ray autoradiogram shown in A was scanned with a soft laser densitometer equipped with an integrator. The percent relative intensity of the mononucleosomes or the naked DNA bands in each lane were calculated and plotted against the novobiocin concentration.
I (lanes 5-10). This is reflected by the loss of the protection pattern seen in the untreated mononucleosome. In addition, a prominent dose-dependent increase in hypersensitivity at positions located between the four distinct protection regions can be seen after novobiocin treatment. In conclusion, novo- biocin induces disruption and unfolding of nucleosome struc- ture and as a result it increases the sensitivity of the mon- onucleosome DNA to cleavage by DNase I.
DISCUSSION
Despite the critical role of internucleosomal DNA cleavage in programmed cell death or apoptosis, few studies have been
Novobiocin - Mononucleosome -
Naked DNA +
1
-- -++++++ -++++t+++ +---------
2 3 4 56 7 6 910
FIG. 7. DNase I footprinting analysis of the effect of novo- biocin on the nuclease sensitivity of the reconstituted mon- onucleosomes. Mononucleosomes reconstituted with the 181-bp 32P- end-labeled DNA fragment and donor chromatin isolated from human CEM lymphocyte nuclei were incubated with various novobiocin concentrations as described in Fig. 6 legend. The mononucleosomes were digested with DNase I and then analyzed on denaturing 6% polyacrylamide gels. The following novobiocin concentrations were used: 0 rg/ml (lanes 3 and 4), 50 pg/ml (lane 5), 100 pg/ml (lane 6), 250 rg/ml (/one 7), 500 pg/ml (lane 8), 1000 pg/ml (lane 9), or 1500 rg/ml (lane 10). For comparison, the 181-hp 32P-end-labeled naked DNA fragment was digested with DNase I and included in lanes 1 and 2.
devoted to elucidating the molecular mechanism(s) control- ling it. Treatment of thymocytes with glucocorticoids triggers the process of apoptosis (2-5). Characteristics of this process are several recognizable morphological and biochemical changes, including cytoplasmic and nuclear membrane bleb- bing, aggregation of organelles, and most dramatic of all is chromatin condensation which reflects internucleosomal DNA cleavage (1, 2). The glucocorticoid induction of inter- nucleosomal DNA fragmentation does not necessarily require expression of a new nuclease(s), but is the result of the activation of a constitutive endogenous endonuclease(s) (15- 18). In rat thymocytes, it appears that activation of DNA cleavage by glucocorticoids is achieved by induction of a Ca*+ uptake system which mediates activation of a Ca*+-dependent endonuclease by providing Ca2+ as a cofactor. However, this may not be true of all cell types. We have shown recently that incubation of CEM-C7 lymphocytes in a Ca*+-depleted me- dium did not prevent glucocorticoid from activating DNA cleavage and cytolysis (18). In addition, in Fig. 4 and Table I of this report we have shown that, unlike thymocyte nuclei, in uitro incubation of CEM-C7 lymphocyte nuclei with differ- ent concentrations of Ca*+ under various incubation condi- tions was not sufficient to cause activation of internucleoso- ma1 DNA fragmentation. These observations indicate that
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Glucocorticoid- and Novobiocin-induced Apoptosis 17331
FIG. 8. A speculative model of the mechanism of internucleosomal DNA cleavage induced by novobio- tin and glucocorticoids. For discus- sion, see text.
ENDONUCLEASE
CEM-C7 lymphocytes employ a different system than the Ca*+ uptake system of thymocytes to activate their DNA cleavage during the process of apoptosis.
We found that novobiocin could be used as a useful probe to investigate the mechanism of activation of DNA cleavage in human CEM lymphocytes. We have demonstrated that novobiocin has the capacity to set into motion the process of apoptosis in CEM lymphocytes. In addition to the morpho- logical features of apoptosis exhibited by novobiocin-treated cells (data not shown), chromatin isolated from these cells revealed the most characteristic molecular feature of apopto- sis which is chromatin cleavage at internucleosomal sites. The apoptosis process can occur only in intact viable cells but not in dead cells. Several observations support this notion. (a) In both novobiocin- and glucocorticoid-induced apoptosis, inter- nucleosomal DNA cleavage preceded by several hours the cellular uptake of trypan blue vital dye; (b) incubation of CEM nuclei with novobiocin does not activate DNA cleavage; (c) pretreatment of CEM cells with ionophore A23187 which caused calcium influx and subsequently membrane injury prevented the novobiocin-induced DNA cleavage; and, finally, (d) killing of CEM cells with sodium azide, cycloheximide, ionophore A23187, and other toxic agents did not cause acti- vation of DNA cleavage. Thus, the activation of internucleo- somal DNA cleavage by novobiocin is a finely regulated process which requires physical integrity of the cell and is not merely a result of cell damage or death. Our observations are consistent with previous conclusions that activation of DNA cleavage in responsive cells by glucocorticoids (4, 5, 13, 15), cytotoxic T-lymphocytes (13, 20), or 2,3,7,8-tetrachlorodi- benzo-p-dioxin (22) precedes cell death and is not a trivial consequence of cell death.
Although protein synthesis is apparently required for the glucocorticoid-induced DNA cleavage in thymocytes (5, 17), it has been shown that protein synthesis inhibitors have no effect on activation of DNA cleavage in target cells by cyto- toxic T-lymphocytes (20). In this report, we provide evidence that the protein synthesis inhibitor, cycloheximide, also has no effect on novobiocin-induced DNA cleavage in CEM lym- phocytes. This suggests that the novobiocin activation of DNA cleavage in CEM cells is not mediated by transcription- ally induced endonuclease(s), rather there are three possible mechanisms by which novobiocin could activate internucleo- somal DNA cleavage in intact CEM lymphocytes. First, in-
hibition of topoisomerase II; second, allowing the entrance of an exogenous endonuclease(s) from the culture medium (i.e. serum) into the cells; and third, activation of a constitutive endogenous endonuclease(s).
The involvement of topoisomerase II as the cleaving en- zyme in DNA cleavage, however, is unlikely, since it was shown that sodium dodecyl sulfate-induced cleavage by to- poisomerase II produces DNA ends with 4-base 5’-phos- phorylated protruding ends (44-46). We have demonstrated that novobiocin- or glucocorticoid-induced DNA cleavage gen- erates double-stranded blunt-ended DNA fragments, which is inconsistent with the properties of the topoisomerase II cleav- age. Furthermore, we have shown that other topoisomerase inhibitors (m-AMSA, inhibitor of topoisomerase II, and camp- tothecin, inhibitor of topoisomerase I) were unable to induce DNA cleavage. The second alternative is also unlikely, since we have shown clearly that in CEM lymphocytes internucleo- somal DNA cleavage can be induced by novobiocin or gluco- corticoid in a serum-free medium. These observations and the fact that novobiocin and glucocorticoid can induce DNA cleavage in CEM lymphocytes in a calcium-free medium strongly suggest that activation of a constitutive non-Ca*+- dependent endogenous endonuclease appears to be the most favored mechanism by which novobiocin or glucocorticoid could induce DNA cleavage.
In this context we would like to propose a speculative model (Fig. 8) to describe some possible events that may precede and subsequently culminate in novobiocin or glucocorticoid- induced internucleosomal DNA fragmentation in CEM lym- phocytes. The chromatin in the nucleus of mammalian cells is normally organized and packaged into tight higher order structures and maintained in a charge neutralized state. Each nucleosome consists of about 166 base pairs of DNA wrapped in two superhelical turns around an octamer of two molecules each of histones H2A, H2B, HS, and Hq to form the core particle (47, 48). Histone H1 is located in the linker DNA regions where DNA enters or exits the core particle (49), thus protecting these regions from nuclease digestion. In the for- mation of higher order structures, histone H1 may interact with adjacent H1 molecules to form a polymer that brings together neighboring nucleosomes to form a structure called the solenoid (50). There are approximately six nucleosomes/ turn of the solenoidal fiber (48). Additional levels of organi- zation of DNA condensation exist to attain the highly con-
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

17332 Glucocorticoid- and Nouobiocin-induced Apoptosis
densed state of DNA in metaphase chromosomes (50, 51). The highly condensed state of chromatin may be expected to render the DNA inaccessible to many enzymes, including endonucleases. Only a small fraction of mammalian nuclear DNA is actively transcribed. In vitro treatment of mammalian nuclei with nucleases, such as staphylococcal nuclease or activation of auto-digestive endogenous endonucleases, pref- erentially causes cleavage of these transcriptionally active sequences (19, 52). It is likely that the accessibility of those transcriptionally active sequences to endonucleases reflects a more open conformation which could be due to local reduction in histone-DNA interactions. Thus, if we hypothesize that the system of activation of DNA cleavage in CEM lympho- cytes employs an endonuclease which requires for its activa- tion some type of change in the chromatin substrate like a state of decondensation or local reduction in histone-DNA interactions, this nuclease would be expected to be inactive in intact cells in which chromatin is compact, charge-neu- tralized, and highly inaccessible. This endonuclease would become active to cleave the linker DNA region only when this region is made accessible. The ability of novobiocin to interact directly with histones and to interfere with histone-histone and/or histone-DNA interactions which maintain the higher order chromatin structure may cause a transition state of chromatin disruption and decondensation. This transition state of disruption might lead to the exposure and unmasking of internucleosomal linker DNA regions which are substrates for a constitutive endonuclease (Fig. 8). The constitutive endonuclease would attack these regions causing internucleo- somal DNA cleavage. It is, therefore, not surprising that novobiocin cannot induce DNA cleavage in ionophore-treated cells or isolated nuclei. We suspect that the transition state of chromatin disruption and decondensation may be counter- acted in ionophore-treated cells or isolated nuclei by chro- matin condensation and/or neutralization of the negatively charged phosophodiester groups of DNA by cation influx. Therefore the endogenous endonuclease may fail to bind to the condensed or charge neutralized chromatin complex.
Evidence supporting this model was obtained from our studies with an in vitro reconstituted mononucleosome. We have demonstrated that novobiocin has the capacity to disrupt and increase the nuclease hypersensitivity of mononucleo- somes reconstituted in vitro. In addition, it has been shown that novobiocin can interact with and precipitate histones from an in vitro chromatin assembly system causing inhibi- tion of passive chromatin assembly (25, 53). Moreover, no- vobiocin can remove histones from previously assembled static chromatin thus making it more accessible to the action of topoisomerase I (25). Since ATP-dependent topoisomerase II activity is not involved in our in uitro reconstitution system, the effect of novobiocin on the structure of mononucleosomes is not, therefore, mediated through inhibitions of topoisom- erase II activity, but may be due to the ability of novobiocin to interact with and precipitate histones (25, 53). However, we cannot ignore the fact that novobiocin could alter chro- matin structure in viva not only by interacting directly with histones but also through its inhibitory effect on topoisom- erase II activity.
The molecular mechanism(s) responsible for glucocorti- coid-induced changes in chromatin structure is not yet known. However, the growth inhibitory and antiproliferative action of glucocorticoids in lymphoid tissues which is probably due to inhibition of synthesis of growth factors and protoonco- genes (54-58) may be sufficient to induce conformational changes in the chromatin structure. On the other hand, acti- vation of ADP-ribosylation, which is probably a secondary
effect of glucocorticoid treatment of lymphoid cells (59), might also serve as a candidate to alter chromatin structure. Recent studies demonstrated that ADP-ribosylation of histone H1 and core histones induces chromatin decondensation (60).
CEM-Cl cells are resistant to glucocorticoid, even though these cells contain functional glucocorticoid receptors (31, 32). We have shown that CEM-Cl is sensitive to the action of novobiocin, which implies that CEM-Cl possesses the endogenous endonuclease and the means to activate it. How- ever, CEM-Cl resistance to glucocorticoid indicates that glu- cocorticoid does not have the capacity to activate DNA cleav- age in these cells. Perhaps this could be due to the lack of glucocorticoid inhibitory effects on the growth and prolifera- tion of these cells.
Although novobiocin and glucocorticoid induce apoptosis through independent mechanisms, both agents share a com- mon final step which is the activation of an endogenous endonuclease. It is interesting that in chicken erythrocytes (43), human CEM lymphocytes, or rat thymocytes, induction of internucleosomal DNA cleavage by novobiocin or glucocor- ticoids is achieved by activation of endogenous endonucleases which generate double-stranded blunt-ended DNA cleavage. This mode of DNA cleavage has not been described for any known eukaryotic endonuclease. Further work is under way in our laboratory to explain the mechanism by which novo- biocin and glucocorticoid induce DNA cleavage.
Acknowledgments-We are grateful to Dr. G. Stein for a gift of histone H1 gene clone and Dr. A. Maksymowych for help in the art work. We thank Denise Valentine and Maria Randall for excellent secretarial assistance.
1.
2.
3. 4.
5. 6.
7.
8.
9.
10. 11. 12.
13. 14.
15.
16.
17.
18.
19.
20.
21.
22.
REFERENCES
Wyllie, A. H., Kerr, J. F. R., and Currie, A. R. (1980) Znt. Reu. Cytol. 68, 251-306
Wyllie, A. H., Morris, R. G., Smith, A. L., and Dunlop, D. (1984) J. Puthol. 142, 67-77
Wyllie, A. H. (1980) Nature 284, 555-556 Compton, M. M., and Cidlowski, J. A. (1986) Endocrinology 118,
38-45 Cohen, J. J., and Duke, R. C. (1984) J. Zmmunol. 132,38-42 Munck, A., and Crabtree, G. R. (1981) in Cell Death in Biology
and Pathology (Bowen, I. D., and Lockshin, R. A., eds) pp. 329- 359, Chapman and Hall, New York
Harmon, J. M., Norman, M. R., Fowles, B. J., and Thompson, E. B. (1979) J. Cell. Physiol. 98, 267-278
Norman, M. R., and Thompson, E. B. (1977) Cancer Res. 37, 3785-3791
Harris, A. W., Bankhurst, A. D., Mason, S., and Warner, N. L. (1973) J. Zmmunol. 110,431-438
Horibata, K., and Harris, A. W. (1970) Exp. Cell Res. 60, 61-77 Yamamoto, K. R. (1985) Anna Reu. Genei. 19, 209-252 Voris. B. P.. andYoune. D.A. (1981) J. Biol. Chem. 256.11319-
11329 Yl
Uker, D. S. (1987) Nature 327,62-64 Compton, M. M., and Cidlowski, J. A. (1987) J. Biol. Chem. 262,
8288-8292 McConkey, D. J., Hartzell, P., Nicotera, P., and Orrenius, S.
(1989) FASEB J. 3, 1843-1849 Jones, D. P., McConkey, D. J., Nicotera, P., and Orrenius, S.
(1989) J. Biol. Chem. 264.6398-6403 McConkey, D. J., Nicotera, P., Hartzell, P., Bellomo, G., Wyllie,
A. H.. and Orrenius. S. (1989) Arch. Biochem. Biohys. 269. 365-370
_ _
Alnemri, E. S., and Litwack, G. (1989) J. Biol. Chem. 264,4104- 4111
Vanderbilt, J. N., Bloom, K. S., and Anderson, J. N. (1982) J. Biol. Chem. 257, 13009-13017
Duke, R. C., Chervenak, R., and Cohen, J. J. (1983) Proc. N&l. Acad. Sci. U. S. A. 80,6361-6365
Kyprianou, N., and Issacs, J. T. (1988) Endocrinology 122, 552- 562
McConkey, D. J., Hartzell, P., Duddy, S. K., HBkansson, H., and
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Glucocorticoid- and Novobiocin-induced Apoptosis
43.
44. 45.
46.
47.
48.
49.
50.
51.
52. 53.
54.
55.
56.
17333
Orrenius, S. (1988) Science 242, 256-259 Smith, C. A., Williams, G. T., Kingston, R., Jenkinson, E. J., and
Owen, J. J. T. (1989) Nature 337, 181-184 Trauth, B. C., Klas, C., Peters, A. M. J., Matzku, S., Moller, P.,
Falk, W., Dehatin, K.-M., and Krammer, P. H. (1989) Science 245,301-305
Villeponteau, B., Pribyl, T. M., Grant, M. H., and Martinson, H. G. (1986) J. Biol. Chem. 261, 10359-10365
Gellert, M. (1981) Annu. Reu. Biochem. 50,879-910 Sander, M.. and Hsieh, T.-S. (1983) J. Biol. Chem. 258. 8421-
8428
Sealy, L., Cotten, M., and Chalkley, R. (1986) EMBO J. 5, 3305- 3310
Lui, L. F., Rowe, T. C., Yang, L., Tewey, K. M., and Chen, G. L. (1983) J. Biol. Chem. 258, 15365-15370
26. Glikin, G. C., Ruberti, I., and Worcel, A. (1984) Cell 37, 33-41 21. Ryoji, M., and Worcel, A. (1984) Cell 37, 21-32 28. Kmiec, E. B., and Worcel, A. (1985) Cell 41,945-953 29. Han, S., Udvardy, A., and Schedl, P. (1985) J. Mol. Biol. 183,
McGhee, J. D., and Felsenfeld, G. (1980) Annu. Reu. Biochem. 49,1115-1156
Igo-Kemenes, T., Horz, W., and Zachau, H. G. (1982) Annu. Reu. Biochem. 5 1,89-121
Foley, G. E., Lararus, H., Farber, S., Uzman, B. G., Boone, B. A., and McCarthv, R. E. (1965) Cancer 18.522-529
Moyne, G., Freeman, R., Savayosti, S., and Yaniv, M. (1981) J. Mol. Biol. 149, 735-744
Thoma, F., Koller, T. H., and Klug, A. (1980) J. Cell Biol. 83, 403-427
Zawydiwski, R.; Harmon, J. M., and Thompson, E. B. (1983) Cancer Res. 43,3865-3873
Thomas, J. O., and Butler, P. J. G. (1980) J. Mol. Biol. 144, 89- 93
Yuh, Y.-S., and Thompson, E. B. (1987) Somatic Cell Mol. Gen. 13,33-46
Buttyan, R., Olsson, C. A., Sheard, B., and Kallos, J. (1983) J. Biol. Chen. 258,14366-14370
Kaufmann, S. H., Okret, S., Wikstrom, A.-C., Gustafsson, J.-A., and Shaper, J. H. (1986) J. Biol. Chem. 261, 11962-11967
van Wijnen, A. J., Wright, K. L., Massung, R. F., Gerretsen, M., Stein, J. L., and Stein, G. S. (1988) Nucleic Acids Res. 16, 571- 592
Bloom, R. S., and Anderson, J. N. (1978) Cell 15, 141-150 Cotten, M., Bresnahan, D., Thompson, S., Sealy, L., and Chalk-
ley, R. (1986) Nucleic Acids Res. 14, 3671-3686 Eastman-Reks, S. B., and Vedeckis, W. V. (1986) Cancer Res.
46,2457-2462 Forsthoefel, A. M., and Thompson, E. A. (1987) Mol. Endocrinol.
1,899-907 Vedeckis, W. V., Eastman-Reks, S. B., Lapointe, M. C., and
Reker, C. E. (1987) in Steroid and Sterol Hormone Action (Spelsberg, T. C., and Kumar, R., eds) pp. 213-216, M. Nijhoot, Boston
23.
24.
25.
13-29 30.
31.
32.
33.
34.
35.
36. Linxweiler W., and Horz, W. (1985) Cell 42, 281-290 37. Perlmann, T., and Wrange, 6. (1988) EMBO J. 7, 3073-3079 38. Lutter. L. C. (1978) J. Mol. Biol. 124. 391-420 39. Tewey; K. M., Chen, G. L., Nelson, E. M., and Liu, L. F. (1984)
40.
41.
42.
J. Biol. Chem. 259,9182-9187 Hsiang, Y.-H., Hertzberg, R., Hecht, S., and Liu, L. F. (1985) J.
Biol. Chem. 260,14873-14878 Kaiser. N.. and Edelman. I. S. (1977) hoc. N&l. Acad. Sci. U. S.
A. 74,638-642 Kaiser, N., and Edelman, I. S. (1978) Endocrinology 103, 936-
942
57.
58.
59.
60.
Bittner, S., and Wielckens, K. (1988) Endocrinology 123, 991- 1000
Yuh, Y.-S., and Thompson, E. B. (1989) J. Biol. Chem. 264, 10904-10910
Wielckens, K., and Delfs, T. (1986) Endocrinology 119, 2383- 2392
Huletsky, A., de Murcia, G., Muller S., Hengartner, M., Menard, L., Lamarre, D., and Poirier, G. G. (1989) J. Biol. Chem. 264, 8878-8886
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from

E S Alnemri and G Litwackmechanism(s).
glucocorticoid and novobiocin. Evidence for a non-Ca2(+)-requiring Activation of internucleosomal DNA cleavage in human CEM lymphocytes by
1990, 265:17323-17333.J. Biol. Chem.
http://www.jbc.org/content/265/28/17323Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/28/17323.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 13, 2019http://w
ww
.jbc.org/D
ownloaded from