a human apoc3 antibody accelerate apoc-iii clearance and ... clearance with... · apoc-iii is a...
TRANSCRIPT

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
nature medicine advance online publication �
Recent large-scale genetic sequencing efforts have identified rare coding variants in genes in the triglyceride-rich lipoprotein (TRL) clearance pathway that are protective against coronary heart disease (CHD), independently of LDL cholesterol (LDL-C) levels1. Insight into the mechanisms of protection of these variants may facilitate the development of new therapies for lowering TRL levels. The gene APOC3 encodes apoC-III, a critical inhibitor of triglyceride (TG) lipolysis and remnant TRL clearance2. Here we report a detailed interrogation of the mechanism of TRL lowering by the APOC3 Ala43Thr (A43T) variant, the only missense (rather than protein-truncating) variant in APOC3 reported to be TG lowering and protective against CHD3–5. We found that both human APOC3 A43T heterozygotes and mice expressing human APOC3 A43T display markedly reduced circulating apoC-III levels. In mice, this reduction is due to impaired binding of A43T apoC-III to lipoproteins and accelerated renal catabolism of free apoC-III. Moreover, the reduced content of apoC-III in TRLs resulted in accelerated clearance of circulating TRLs. On the basis of this protective mechanism, we developed a monoclonal antibody targeting lipoprotein-bound human apoC-III that promotes circulating apoC-III clearance in mice expressing human APOC3 and enhances TRL catabolism in vivo. These data reveal the molecular mechanism by which a missense variant in APOC3 causes reduced circulating TG levels and, hence, protects from CHD. This protective mechanism has the potential to be exploited as a new therapeutic approach to reduce apoC-III levels and circulating TRL burden.
ApoC-III is a small apolipoprotein (~8.8 kDa) secreted from the liver and small intestine that circulates on TRLs such as VLDL and chylom-icrons, as well as on HDLs. In biochemical studies and experimental animals, apoC-III has been shown to increase plasma TG levels by both direct inhibition of the activity of lipoprotein lipase (LPL) on TRLs and inhibition of the clearance of TRLs by the liver, possibly through competitive interactions with apoE, a critical ligand on TRLs for receptor-mediated clearance of TRLs6–9. In an early study using a mouse model of delayed apoB clearance due to LDL receptor defi-ciency, human apoC-III overexpression increased circulating TRL levels and exacerbated the development of atherosclerosis10, provid-ing initial support for the notion that apoC-III may be a positive mediator of CHD risk.
In humans, several studies of the metabolism of TRLs have impli-cated apoC-III as a critical contributor to hypertriglyceridemia11–14. Dyslipidemia, metabolic syndrome, insulin resistance, visceral adi-posity, chronic renal insufficiency, and several other systemic meta-bolic diseases are associated with elevated apoC-III levels, which may be due to both increased apoC-III production and apoC-III secretion on VLDL15–18, as well as to delayed TRL clearance11–14. ApoC-III exchanges rapidly among TRLs, lipoprotein remnants, and HDLs in humans19,20, which has made accurate assessment of the kinetics and metabolism of apoC-III in humans challenging14,21,22. Some studies have identified apoC-III on LDLs as a marker of a small, dense proath-erogenic phenotype for LDL23,24. ApoC-III has also been identified as a constituent of lipoprotein(a) and may alter the metabolism of this lipoprotein particle through as-yet-undetermined mechanisms25. Analyses of plasma apoC-III levels and lipoprotein-subfraction- associated apoC-III levels have suggested a direct relationship between
A human APOC3 missense variant and monoclonal antibody accelerate apoC-III clearance and lower triglyceride-rich lipoprotein levelsSumeet A Khetarpal1,2, Xuemei Zeng3, John S Millar2, Cecilia Vitali2, Amritha Varshini Hanasoge Somasundara1,2, Paolo Zanoni1,2, James A Landro4, Nicole Barucci4, William J Zavadoski4, Zhiyuan Sun3, Hans de Haard5, Ildikó V Toth6, Gina M Peloso7, Pradeep Natarajan8–10, Marina Cuchel2, Sissel Lund-Katz1,2, Michael C Phillips1,2, Alan R Tall11, Sekar Kathiresan8–10, Paul DaSilva-Jardine4, Nathan A Yates3,12 & Daniel J Rader1,2
1Department of Genetics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, USA. 2Department of Medicine, Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, USA. 3Biomedical Mass Spectrometry Center, Schools of the Health Sciences, University of Pittsburgh, Pittsburgh, Pennsylvania, USA. 4Staten Biotechnology BV, Nijmegen, the Netherlands. 5argenx BVBA, Zwjnaarde, Belgium. 6FairJourney Biologics, Porto, Portugal. 7Department of Biostatistics, Boston University School of Public Health, Boston, Massachusetts, USA. 8Center for Genomic Medicine, Massachusetts General Hospital, Boston, Massachusetts, USA. 9Department of Medicine, Harvard Medical School, Boston, Massachusetts, USA. 10Program in Medical and Population Genetics, Broad Institute, Cambridge, Massachusetts, USA. 11Division of Molecular Medicine, Department of Medicine, Columbia University, New York, New York, USA. 12Department of Cell Biology, School of Medicine, University of Pittsburgh, Pittsburgh, Pennsylvania, USA. Correspondence should be addressed to D.J.R. ([email protected]).
Received 28 November 2016; accepted 25 July 2017; published online 21 August 2017; doi:10.1038/nm.4390

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
� advance online publication nature medicine
apoC-III levels in the circulation and cardiovascular events26,27, although large prospective studies verifying this relationship have yet to be reported. Moreover, multiple lipid-lowering drugs, such as statins, fenofibrates, and pioglitazone, potently reduce circulating apoC-III plasma levels through either reducing apoC-III production or increasing its clearance2,28.
Genetic discoveries of the last decade have highlighted the potential importance of apoC-III to CHD risk in humans. Genome-wide asso-ciation studies (GWAS) have demonstrated that common noncoding polymorphisms in APOC3 are associated with the levels of plasma TG, HDL cholesterol (HDL-C), and non-HDL-C, as well as with CHD risk29. In 2008, the Lancaster Amish were reported to have a relatively high frequency of a truncating variant in APOC3, Arg19Ter (R19*), associated with reduced levels of serum apoC-III, reduced fasting and postprandial TG levels, and reduced coronary artery calcification, a surrogate measure of coronary atherosclerosis5. More recently, two large sequencing efforts3,4 have identified several rare coding variants in APOC3 associated, in aggregate, with a ~40% reduction in TG lev-els and a similar magnitude of reduction in the risk of CHD3,4. These efforts identified four protective APOC3 variants, of which three were classic loss-of-function variants: a nonsense variant (R19*) and two splice-site variants (IVS2+1G>A and IVS3+1G>T). Of these variants, the R19* variant is predicted to disrupt protein expression through premature termination, and the two splice-site variants are predicted to abolish splice-donor sites and cause retention of adjacent introns by altering the RNA thermodynamic properties required for process-ing (Supplementary Table 1). In contrast, the fourth APOC3 variant identified was a missense variant (A43T; also known as A23T on the basis of amino acid sequence numbering for the mature form of the protein). When grouped together, carriers of any of these variants displayed significantly lower apoC-III and TG levels, as compared to non-carriers.
These genetic data suggested that reducing circulating apoC-III lev-els might be a therapeutic approach to reduce circulating TRL levels and CHD risk. Indeed, an antisense oligonucleotide targeting APOC3 mRNA—in essence mimicking the loss-of-function mutations—has been shown to reduce apoC-III and TG levels in preclinical models30 and in humans with hypertriglyceridemia31. On the basis of these results, we hypothesized that intensive investigation of the mechanism of protection by the missense variant A43T might offer insights into alternative therapeutic approaches to reduce apoC-III levels.
To better understand the specific phenotypic consequences of the A43T variant, we searched for A43T carriers using exome-wide genotyping. Because carriers of APOC3 loss-of-function variants have been reported to have high HDL-C levels, we hypothesized that these variants, including A43T, would be enriched in frequency in individuals with extremely high HDL-C levels. We genotyped for the A43T variant in 1,056 participants of the Penn High HDL-C study (HHDL), a cohort of individuals whose HDL-C levels are above the 90th percentile of the general population, and identified 13 A43T carriers (all heterozygous). We also genotyped a cohort of 5,744 indi-viduals with a normal HDL-C distribution, from the Penn Medicine BioBank, and identified an additional six A43T carriers (all hetero-zygous). These results indicate that the A43T variant was present at a significantly higher frequency in the cohort with high HDL-C versus the one with a normal HDL-C distribution (1.2% versus 0.1%, P < 0.0001, Fisher’s exact test; Fig. 1a). Using the 19 A43T carriers and 76 non-carrier controls matched by age, sex, and ancestry (Online Methods), we examined the levels of plasma lipids and apolipopro-teins, including measurements that had not been previously made
in the few earlier studies of this variant, such as measurements of apoA-I, non-HDL-C, and apoB levels. APOC3 A43T heterozygotes displayed decreased fasting TG levels (Fig. 1b) and increased apoA-I and HDL-C levels (Supplementary Fig. 1a,b), but no significant differences in LDL-C, non-HDL-C, or apoB levels (Supplementary Fig. 1c–e). In addition, A43T carriers displayed significantly reduced levels of plasma apoC-III that were approximately 50% of those meas-ured in non-carriers (Fig. 1c). As the A43T variant is not predicted to have an impact on RNA binding, splicing, or expression of APOC3 mRNA (Supplementary Table 1), we next investigated the mecha-nism responsible for the lower circulating apoC-III levels observed in the heterozygous A43T carriers.
To quantify the relative concentrations of mutant and wild-type (WT) apoC-III in the plasma of A43T heterozygotes, we developed a sequence-specific liquid chromatography and tandem mass spectrom-etry (LC–MS/MS) assay (illustrated in Supplementary Fig. 2). The LC–MS/MS assay measures peptides generated by proteolytic diges-tion of plasma with the endoproteinase AspN, followed by oxidation of methionine residues with hydrogen peroxide. Selective reaction mon-itoring was used to measure the relative abundance of peptides (DASLLSFMoxQGYMoxKHATKTAK and DASLLSFMoxQGYMoxKHATKTTK, where the variant residue is underlined), corresponding to the WT and mutant forms of apoC-III, respectively, that contain alanine (Supplementary Fig. 3a) or threonine (Supplementary Fig. 3b) at position 43 of the protein sequence. Internal-standard peptides, synthesized with isotopically labeled leucine, were added to each sam-ple to facilitate quantification of WT and mutant apoC-III protein over a range of concentrations, based on comparison of the product ion intensities for the light versus heavy peptide (Supplementary Fig. 4 and Supplementary Tables 2 and 3). Using this assay, A43T carrier plasma contained clearly detectable levels of A43T mutant apoC-III peptide, whereas non-carrier plasma did not (Fig. 1d). Additionally, the levels of WT apoC-III were substantially lower in the plasma of the A43T carriers than in non-carriers (Fig. 1e). Notably, in A43T carriers, the levels of mutant apoC-III were mark-edly lower than those of WT apoC-III, as assessed by the ratio of peptide abundance, with a mutant:WT ratio of 0.14:1 (as compared with the expected 1:1 ratio anticipated for heterozygotes) (Fig. 1f). This result indicates that a significant imbalance exists in the ratio of circulating mutant to WT apoC-III in the plasma of A43T carriers (P < 0.0001, one-sample t test).
To study the mechanism underlying the reduced levels of circu-lating mutant and total apoC-III in A43T carriers, we developed a mouse model using administration of adeno-associated virus (AAV) vectors encoding either WT or A43T human APOC3. AAV-medi-ated WT APOC3 expression caused a dose-dependent elevation in circulating human apoC-III levels, as well as an increase in plasma TG levels (Supplementary Fig. 5). A dose of 3 × 1011 genome copies (GC) of the AAV encoding WT APOC3 resulted in circulating human apoC-III levels of 10–15 mg/dl, mirroring the range of normal apoC-III concentrations in humans, and also significantly increased fasting TG levels (Supplementary Fig. 5). We therefore used this AAV dose to compare the effects of WT and A43T APOC3 expression in subsequent experiments. Although the effects on TG metabolism of WT and mutant apoC-III expression using this approach may not correspond to the effects of endogenous WT and mutant apoC-III expression, this approach allowed us to com-pare the relative effects of WT versus mutant apoC-III in multiple normolipidemic and hyperlipidemic models, including WT mice, Apoc3-knockout mice, and several ‘humanized’ hyperlipidemic

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
nature medicine advance online publication �
models, with relatively consistent levels of circulating apoC-III across these models (Supplementary Table 4).
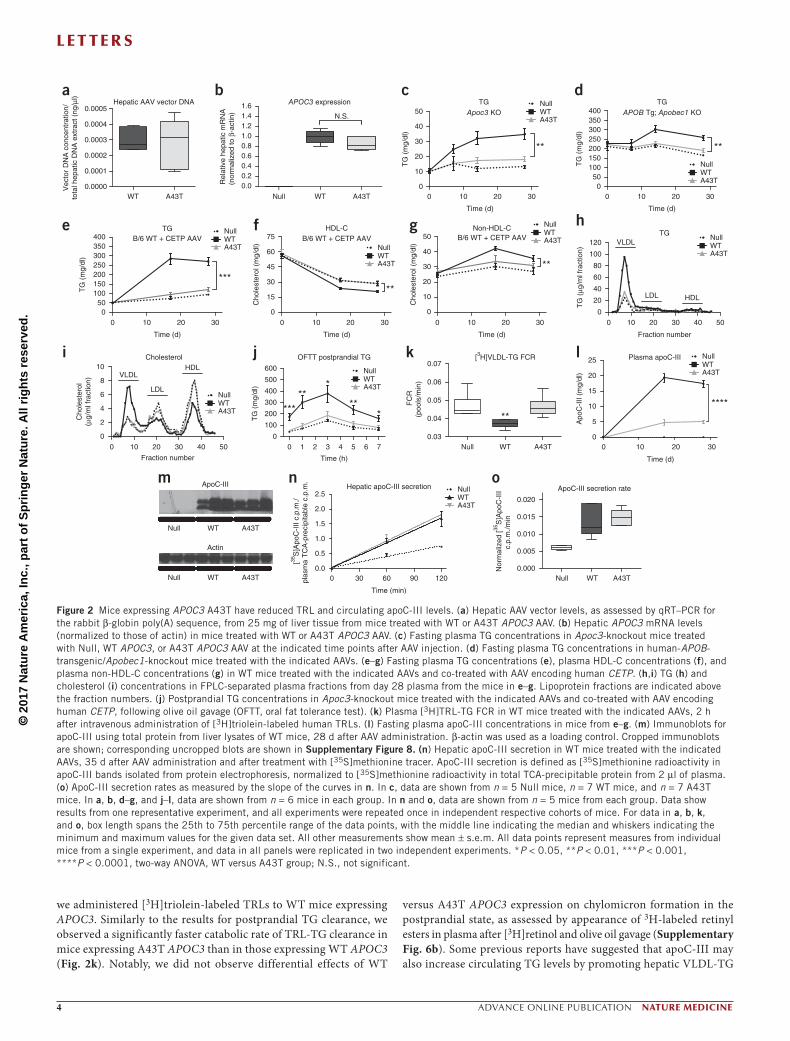
Administration of WT and A43T APOC3 AAV at the selected dose conferred equal hepatic AAV vector DNA levels and APOC3 transcript expression (Fig. 2a,b). Despite equal mRNA levels, WT APOC3 expression raised fasting plasma TG levels, whereas A43T APOC3 expression did not affect fasting plasma TG levels in all mouse models studied (Fig. 2c–e and Supplementary Table 4). For example, in WT mice coexpressing human cholesteryl ester transfer protein (CETP), a key mediator of lipid exchange between TRLs and HDL in humans, which is absent in rodents, WT apoC-III expression raised TG and non-HDL-C levels and reduced HDL-C levels, whereas A43T apoC-III expression had no effect on TG, non-HDL-C, or HDL-C levels (Fig. 2e–g). This difference between the groups expressing WT and A43T apoC-III in WT mice coexpressing CETP was confirmed using lipoprotein fractions isolated from plasma by fast protein liquid chromatography (FPLC) (Fig. 2h,i). In addition to elevated HDL-C levels, A43T-expressing mice displayed reduced VLDL-TG
and VLDL-C levels. The overall atheroprotective phenotype in this humanized model thus partially recapitulates the findings observed in human carriers of the A43T variant, who display reduced TG and increased HDL-C levels.
We next assessed LPL-mediated TG clearance in Apoc3-knockout mice coexpressing CETP and either WT or A43T APOC3 by measuring TG levels after intragastric administration of olive oil. Mice expressing WT APOC3 demonstrated significantly delayed postprandial TG clearance relative to control mice treated with Null virus (Fig. 2j). In contrast, mice expressing A43T APOC3 exhibited no increase in postprandial TG levels relative to control mice. This relative differ-ence between mice expressing WT and A43T APOC3 was abolished when this experiment was repeated in the presence of intravenous Poloxamer P407 (Pluronic), a potent competitive inhibitor of LPL (Supplementary Fig. 6a), suggesting that differences in the ability of A43T and WT apoC-III to inhibit LPL account for their differential effects on postprandial TG clearance. To assess the contribution of post-absorptive TG clearance to the observed plasma TG phenotype,
Non-carriers A43T carriers
–0.2
0.0
0.2
0.4
0.6
0.8
1.0R
atio
of m
utan
t:WT
apo
C-I
II(a
rbitr
ary
units
)
Mutant:WT apoC-III ratioExpected ratio
for no imbalance
****
Non-carriers A43T carriers
0
50
100
150
200
250
300
350
400
TG
(m
g/dl
)
TG
****
a
b c d
e f
Non-carriers A43T carriers
0
5
10
15
20
25
30
Apo
C-I
II (m
g/dl
)
WT apoC-III in plasma
**
Non-carriers A43T carriers
0
5
10
15
20
25
30
Apo
C-I
II (m
g/dl
)
ApoC-III
***
Non-carriers A43T carriers
–1.0
–0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Apo
C-I
II (m
g/dl
)
Mutant apoC-III in plasma
****
HHDL 1,056 13 0.00616 (~1/162) 0.01231 (~1/81)
Penn Medicine BioBank 5,744 6 0.00052 (~1/1,915) 0.00104 (~1/957)P < 0.0001
Total number of participants
APOC3 A43Tcarriers identified
Discovery cohort A43T allelefrequency
A43T carrierfrequency
Association of A43T allele with HHDLvs. BioBank cohort (Fisher's exact test)
Figure 1 Human APOC3 A43T carriers exhibit lower apoC-III levels than non-carriers. (a) APOC3 A43T carriers were identified from exome-wide genotyping in the HHDL and Penn Medicine BioBank cohorts. The significance of the difference in A43T carrier frequency between the two cohorts was assessed by a Fisher’s exact test. (b) TG concentration in overnight-fasted plasma of A43T carriers versus age-, sex-, and ancestry-matched controls (non-carriers) from the two cohorts. (c) Total apoC-III concentration in fasting plasma from A43T carriers and non-carrier controls. (d) A43T apoC-III concentrations in plasma samples of non-carriers and A43T carriers. (e) WT apoC-III concentrations in plasma samples of non-carriers and A43T carriers. (f) The mutant:WT apoC-III ratio of non-carriers and A43T carriers was compared to an expected ratio of 1:1 for no imbalance by a one-sample t test with an expected mean of 1.0. For b and c, n = 19 for A43T carriers and 76 for matched non-carriers. For d–f, n = 19 for A43T carriers and 21 for matched non-carriers. All measurements in b–f were replicated twice in the same plasma samples. All measurements are shown as mean ± s.d., and each data point depicts a single measure from an individual human participant plasma sample. **P < 0.01, ***P < 0.001, ****P < 0.0001, Student’s unpaired two-sided t test. For f, ****P < 0.0001, one-sample t test.
© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
� advance online publication nature medicine
we administered [3H]triolein-labeled TRLs to WT mice expressing APOC3. Similarly to the results for postprandial TG clearance, we observed a significantly faster catabolic rate of TRL-TG clearance in mice expressing A43T APOC3 than in those expressing WT APOC3 (Fig. 2k). Notably, we did not observe differential effects of WT
versus A43T APOC3 expression on chylomicron formation in the postprandial state, as assessed by appearance of 3H-labeled retinyl esters in plasma after [3H]retinol and olive oil gavage (Supplementary Fig. 6b). Some previous reports have suggested that apoC-III may also increase circulating TG levels by promoting hepatic VLDL-TG
0 10 20 30 40 50
0
2
4
6
8
10
Fraction number
Cho
lest
erol
(µg/
ml f
ract
ion)
Cholesterol
VLDL
LDL
HDL
0 10 20 300
50100150200250300350400
Time (d)
TG
(m
g/dl
)
TG
**
APOB Tg; Apobec1 KO
0 10 20 300
50100150200250300350400
Time (d)
0 10 20 30
Time (d)
0 10 20 30
Time (d)
TG
(m
g/dl
)
TG
***
B/6 WT + CETP AAV
0 10 20 300
10
20
30
40
50
0
10
20
30
40
50
Time (d)
TG
(m
g/dl
)
TG
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
NullWTA43T
**
Apoc3 KO
0 10 20 300
5
10
15
20
25
Time (d)
Apo
C-I
II (m
g/dl
)
Plasma apoC-III
****
Null WT A43T
0.00.20.40.60.81.01.21.41.6
Rel
ativ
e he
patic
mR
NA
(nor
mal
ized
to β
-act
in)
APOC3 expression
N.S.
0 30 60 90 1200.0
0.5
1.0
1.5
2.0
2.5
Time (min)
Hepatic apoC-III secretion
Null WT A43T0.000
0.005
0.010
0.015
0.020N
orm
aliz
ed [35
S]A
poC
-III
c.p.
m./m
in
ApoC-III secretion rate
0 1 2 3 4 5 6 70
100
200
300
400
500
600
Time (h)
TG
(m
g/dl
)
OFTT postprandial TG
*****
*
***
Null WT A43T0.03
0.04
0.05
0.06
0.07
[35S
]Apo
C-I
II c.
p.m
./pl
asm
a T
CA
-pre
cipi
tabl
e c.
p.m
.
[3H]VLDL-TG FCR
FC
R(p
ools
/min
) **
WT A43T0.0000
0.0001
0.0002
0.0003
0.0004
0.0005Hepatic AAV vector DNA
Vec
tor
DN
A c
once
ntra
tion/
tota
l hep
atic
DN
A e
xtra
ct (
ng/µ
l)
0
15
30
45
60
75
Cho
lest
erol
(m
g/dl
)
HDL-C
**
**
B/6 WT + CETP AAV
Cho
lest
erol
(m
g/dl
)
Non-HDL-CB/6 WT + CETP AAV
0 10 20 30 40 50
0
20
40
60
80
100
120
Fraction number
TG
(µg
/ml f
ract
ion)
TGVLDL
HDLLDL
Null WT A43T
Null WT A43T
ApoC-III
Actin
a b c d
e f g h
i j
m n o
k l
Figure 2 Mice expressing APOC3 A43T have reduced TRL and circulating apoC-III levels. (a) Hepatic AAV vector levels, as assessed by qRT–PCR for the rabbit β-globin poly(A) sequence, from 25 mg of liver tissue from mice treated with WT or A43T APOC3 AAV. (b) Hepatic APOC3 mRNA levels (normalized to those of actin) in mice treated with WT or A43T APOC3 AAV. (c) Fasting plasma TG concentrations in Apoc3-knockout mice treated with Null, WT APOC3, or A43T APOC3 AAV at the indicated time points after AAV injection. (d) Fasting plasma TG concentrations in human-APOB-transgenic/Apobec1-knockout mice treated with the indicated AAVs. (e–g) Fasting plasma TG concentrations (e), plasma HDL-C concentrations (f), and plasma non-HDL-C concentrations (g) in WT mice treated with the indicated AAVs and co-treated with AAV encoding human CETP. (h,i) TG (h) and cholesterol (i) concentrations in FPLC-separated plasma fractions from day 28 plasma from the mice in e–g. Lipoprotein fractions are indicated above the fraction numbers. (j) Postprandial TG concentrations in Apoc3-knockout mice treated with the indicated AAVs and co-treated with AAV encoding human CETP, following olive oil gavage (OFTT, oral fat tolerance test). (k) Plasma [3H]TRL-TG FCR in WT mice treated with the indicated AAVs, 2 h after intravenous administration of [3H]triolein-labeled human TRLs. (l) Fasting plasma apoC-III concentrations in mice from e–g. (m) Immunoblots for apoC-III using total protein from liver lysates of WT mice, 28 d after AAV administration. β-actin was used as a loading control. Cropped immunoblots are shown; corresponding uncropped blots are shown in Supplementary Figure 8. (n) Hepatic apoC-III secretion in WT mice treated with the indicated AAVs, 35 d after AAV administration and after treatment with [35S]methionine tracer. ApoC-III secretion is defined as [35S]methionine radioactivity in apoC-III bands isolated from protein electrophoresis, normalized to [35S]methionine radioactivity in total TCA-precipitable protein from 2 µl of plasma. (o) ApoC-III secretion rates as measured by the slope of the curves in n. In c, data are shown from n = 5 Null mice, n = 7 WT mice, and n = 7 A43T mice. In a, b, d–g, and j–l, data are shown from n = 6 mice in each group. In n and o, data are shown from n = 5 mice from each group. Data show results from one representative experiment, and all experiments were repeated once in independent respective cohorts of mice. For data in a, b, k, and o, box length spans the 25th to 75th percentile range of the data points, with the middle line indicating the median and whiskers indicating the minimum and maximum values for the given data set. All other measurements show mean ± s.e.m. All data points represent measures from individual mice from a single experiment, and data in all panels were replicated in two independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, two-way ANOVA, WT versus A43T group; N.S., not significant.
© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
nature medicine advance online publication �
secretion by facilitating the fusion of cytoplasmic TG droplets with nascent VLDL particles32. To test this possibility, we measured VLDL-TG secretion into blood in mice after administration of Poloxamer
P407 (Sigma-Aldrich)33, but found no differences in TG, apoB-100, or apoB-48 secretion rates in mice expressing A43T apoC-III ver-sus WT apoC-III or Null virus (Supplementary Fig. 7). Collectively,
0 250 500 750 1,000 1,250 1,500
0.1
1
Time (min)
Fra
ctio
n of
c.p
.m. a
t 1 m
in
Blood [125I]TC apoC-III clearance
A43T
****
WT
WT A43T
0.0
0.2
0.4
0.6
0.8
c.p.
m./3
0 m
g tis
sue
(nor
mal
ized
to p
lasm
a c.
p.m
. at 1
min
)
Liver [125I]TC
**
20 40 60 80
0
2
4
6
8
10
[125I]ApoC-III activity in plasma FPLC fractions
Fraction number
Per
cent
of t
otal
act
ivity
acr
oss
frac
tions
WTA43T
WTA43T
HDL
LDLVLDL
UnboundAPOB Tg; Apobec1 KO mouse plasma
WT A43T
0.0
0.5
1.0
1.5
2.0
2.5
3.0
FC
R (
pool
s/h)
[125I]TC plasma FCR
****
WT A43T
0
5
10
15
c.p.
m./3
0 m
g tis
sue
(nor
mal
ized
to p
lasm
a c.
p.m
. at 1
min
)
Kidney [125I]TC
**
0
20
40
60
80
100
HDL Unbound
Per
cent
of t
otal
act
ivity
HDL binding
*
*
0
20
40
60
80
100
VLDL Unbound
Per
cent
of t
otal
125 I a
ctiv
ity
TRL binding
WT
A43T
WT
A43T
WT
A43T
**
**
WT A43T
0
2
4
6
Kd
(µM
)
Dissociation constant
**
20 40 60 80
0
2
4
6
8
Fraction number
Per
cent
of t
otal
act
ivity
acr
oss
frac
tions
[125I]ApoC-III activity in plasma FPLC fractions
VLDL
LDL
HDL
Unbound
Fractionated human plasma
a cb
d e f
g h i
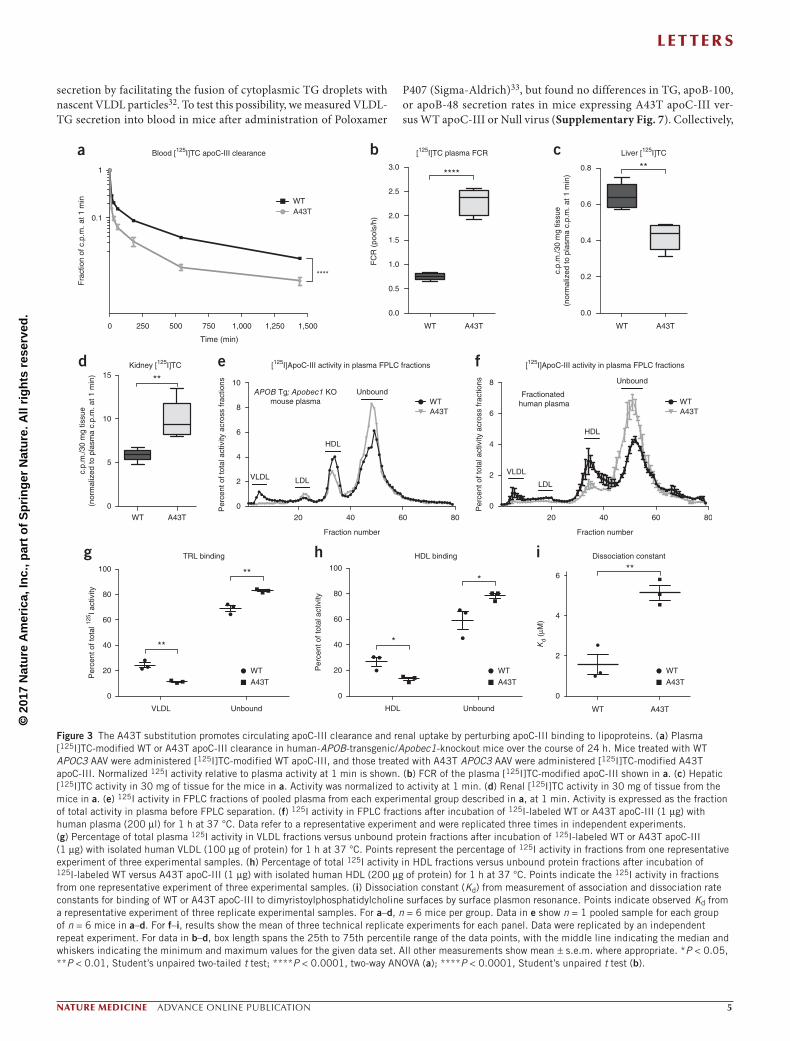
Figure 3 The A43T substitution promotes circulating apoC-III clearance and renal uptake by perturbing apoC-III binding to lipoproteins. (a) Plasma [125I]TC-modified WT or A43T apoC-III clearance in human-APOB-transgenic/Apobec1-knockout mice over the course of 24 h. Mice treated with WT APOC3 AAV were administered [125I]TC-modified WT apoC-III, and those treated with A43T APOC3 AAV were administered [125I]TC-modified A43T apoC-III. Normalized 125I activity relative to plasma activity at 1 min is shown. (b) FCR of the plasma [125I]TC-modified apoC-III shown in a. (c) Hepatic [125I]TC activity in 30 mg of tissue for the mice in a. Activity was normalized to activity at 1 min. (d) Renal [125I]TC activity in 30 mg of tissue from the mice in a. (e) 125I activity in FPLC fractions of pooled plasma from each experimental group described in a, at 1 min. Activity is expressed as the fraction of total activity in plasma before FPLC separation. (f) 125I activity in FPLC fractions after incubation of 125I-labeled WT or A43T apoC-III (1 µg) with human plasma (200 µl) for 1 h at 37 °C. Data refer to a representative experiment and were replicated three times in independent experiments. (g) Percentage of total plasma 125I activity in VLDL fractions versus unbound protein fractions after incubation of 125I-labeled WT or A43T apoC-III (1 µg) with isolated human VLDL (100 µg of protein) for 1 h at 37 °C. Points represent the percentage of 125I activity in fractions from one representative experiment of three experimental samples. (h) Percentage of total 125I activity in HDL fractions versus unbound protein fractions after incubation of 125I-labeled WT versus A43T apoC-III (1 µg) with isolated human HDL (200 µg of protein) for 1 h at 37 °C. Points indicate the 125I activity in fractions from one representative experiment of three experimental samples. (i) Dissociation constant (Kd) from measurement of association and dissociation rate constants for binding of WT or A43T apoC-III to dimyristoylphosphatidylcholine surfaces by surface plasmon resonance. Points indicate observed Kd from a representative experiment of three replicate experimental samples. For a–d, n = 6 mice per group. Data in e show n = 1 pooled sample for each group of n = 6 mice in a–d. For f–i, results show the mean of three technical replicate experiments for each panel. Data were replicated by an independent repeat experiment. For data in b–d, box length spans the 25th to 75th percentile range of the data points, with the middle line indicating the median and whiskers indicating the minimum and maximum values for the given data set. All other measurements show mean ± s.e.m. where appropriate. *P < 0.05, **P < 0.01, Student’s unpaired two-tailed t test; ****P < 0.0001, two-way ANOVA (a); ****P < 0.0001, Student’s unpaired t test (b).

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
� advance online publication nature medicine
0 30 60 90 120 150 1800
255075
100125150
Time (min)
TG
(m
g/dl
)
OFTT postprandial TG
**
Ctrl STT5050
1,000
2,000
3,000
Fasting apoC-III AUC
Apo
C-I
II A
UC
(mg/
dl ×
min
) ***
Ctrl STT5050
500
1,000
1,500
2,000
OFTT apoC-III AUC
**
Ctrl STT505–8,000–6,000–4,000–2,000
02,0004,0006,0008,000
10,000
Apo
C-I
II A
UC
(mg/
dl ×
min
)
Apo
C-I
II A
UC
(mg/
dl ×
min
)
OFTT TG i-AUC*
0 0.5 1.0 4 8 12 16 20 24 280
20406080
100120140
Time (d)
% c
ontr
ol A
b re
spon
seApoC-III
***
FC
R(p
ools
/h)
0 0.5 1.0 4 8 12 16 20 24 280
20406080
100120140
Time (d)
% c
ontr
ol A
b re
spon
se
ApoB
**
0 3 6 9 12 15 18 21 240
20406080
100120140
Time (h)
% c
ontr
olA
b re
spon
se
ApoB
***
0 250 500 750 1,000 1,250 1,500
0.1
1
Time (min)
Fra
ctio
n of
c.p
.m. a
t 1 m
in Blood [125I]TC apoC-III clearance
****
Ctrl STT50580.0
0.1
0.2
0.3
0.4
0.5[125I]TC plasma FCR
**
Ctrl STT50580.0
0.2
0.4
0.6
Spleen [125I]TC
c.p.
m./2
0 m
g tis
sue
(nor
mal
ized
to p
lasm
a c.
p.m
. at 1
min
)
c.p.
m./2
0 m
g tis
sue
(nor
mal
ized
to p
lasm
a c.
p.m
. at 1
min
)
c.p.
m./2
0 m
g tis
sue
(nor
mal
ized
to p
lasm
a c.
p.m
. at 1
min
)***
Ctrl CtrlSTT5058 STT50580.0
0.5
1.0
1.5
Liver [125I]TC
012345678
Kidney [125I]TC
**
0 3 6 9 12 15 18 21 240
20406080
100120140
a
e
i
l
n o p
m q
j k
f g h
b c d
Time (h)
% c
ontr
olA
b re
spon
se
ApoC-III
CtrlSTT505
***
0 30 60 90 120 150 18002468
1012
Time (min)
Apo
C-I
II (m
g/dl
)
Plasma apoC-III during OFTT
*
AAV APOC3 WT3 weeks
B/6 WT
STT505 anti-apoC-IIImAb vs. isotype ctrl
(25 mg/kg i.v.)
STT5058 anti-apoC-IIImAb vs. isotype ctrl(20 mg/kg subcut.)
B/6 WT
ApoC-III WT
Lipolysis andcatabolism
TRL
TRL
TRL
ApoC-III A43T
Lipolysis andcatabolism
Anti-apoC-III antibody
Lipolysis andcatabolism
AAV APOC3 WT3 weeks
CtrlSTT505
CtrlSTT505
CtrlSTT5058
CtrlSTT5058
CtrlSTT5058
CtrlSTT505
Figure 4 Anti-human-apoC-III monoclonal antibodies STT505 and STT5058 lower circulating apoC-III levels and promote TRL clearance. (a) Schematic of the experimental approach, in which the STT505 monoclonal antibody (mAb) or isotype control (ctrl) antibody was tested in C57BL/6 WT (B/6 WT) mice treated with WT APOC3 AAV for 3 weeks. (b) Plasma apoC-III levels over the course of 24 h following antibody administration. Values for the STT505 group are expressed as percentages of those for the control antibody group at the same time point. (c) Plasma apoC-III areas under the curve (AUCs) per mouse for each group in b. (d) Plasma apoB concentrations in mice from b over the course of 24 h. Values for the STT505 group are expressed as percentages of those for the control antibody group at the same time point. (e) Plasma apoC-III concentrations in mice after control or STT505 antibody administration and subsequent intragastric gavage of olive oil. (f) Plasma apoC-III AUCs per mouse for each group in e. (g) Postprandial plasma TG concentrations for the mice in e. (h) Postprandial TG elevation as measured by incremental AUC (i-AUC) per mouse for the groups in e. For i-AUCs, AUCs were calculated relative to a baseline defined as the mean plasma TG at time 0 for all mice in both the control and STT505 groups (72.74 mg/dl). (i) Schematic of the experimental approach, in which the STT5058 monoclonal antibody was tested in WT mice treated with WT APOC3 AAV for 3 weeks. (j,k) Plasma apoC-III (j) and apoB (k) levels over the course of 28 d following antibody administration. Values for the STT5058 group are expressed as percentages of those for the control antibody group at the same time point. (l) Clearance of [125I]TC-modified apoC-III in WT mice that had been treated with APOC3 AAV 3 weeks before administration of STT5058 or control antibody (25 mg/kg subcutaneous dosing), followed 24 h later by intravenous administration of [125I]TC-modified WT apoC-III. Clearance of radiolabeled apoC-III was measured over the course of 21 h. (m) FCR of [125I]TC-modified apoC-III estimated from clearance curves in mice treated with STT5058 versus isotype control antibody. (n–p) Uptake of radiolabeled apoC-III in 20 mg of spleen (n), liver (o), or kidney (p) tissue 21 h after protein administration (values were normalized to plasma activity at 1 min for each mouse). (q) Proposed model of the contribution of TRL-associated apoC-III to TRL clearance. Top, WT apoC-III is bound to TRLs and is capable of inhibiting lipolysis and catabolism of circulating TRLs; a smaller pool of lipoprotein-free apoC-III may be cleared renally. Middle, A43T apoC-III has impaired binding to lipoproteins, augmenting apoC-III clearance by the kidney and promoting TRL lowering. Bottom, the STT505 and STT5058 monoclonal antibodies targeting apoC-III promote clearance of circulating apoC-III partially through an alternative splenic pathway, resulting in TRL lowering. For b and d, n = 7 mice per group. For c and f, n = 8 mice per group. For e, n = 9 mice in the control group and 10 mice in the STT505 group. Each experiment was replicated in an independent group of mice. For g and h, n = 10 mice in the control group and 9 mice in the STT505 group. For j and k, n = 7 mice for each group. Each experiment was replicated once in an independent group of mice. For l–p, n = 10 mice in each group. Each experiment in l–p was performed in one cohort of mice. All data show measures from individual mice. For data in c, f, h, and m–p, box length spans the 25th to 75th percentile range of the data points, with the middle line indicating the median and whiskers indicating the minimum and maximum values for the given data set. All other measurements show mean ± s.e.m. where appropriate. For b, d, e, and j–l, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, two-way ANOVA. For c, f–h, and m–p, *P < 0.05, **P < 0.01, ***P < 0.001, Student’s unpaired two-tailed t test.

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
nature medicine advance online publication �
these data indicate that WT but not A43T apoC-III impairs the in vivo lipolysis of postprandial TRLs.
Despite equal hepatic apoC-III mRNA expression in mice express-ing WT versus A43T apoC-III, WT mice coexpressing CETP that were infected with A43T APOC3 AAV displayed markedly reduced steady-state levels of apoC-III in plasma relative to mice infected with WT APOC3 AAV (Fig. 2l). Because we also observed a reduced amount of A43T apoC-III in the plasma of human A43T carriers as compared to non-carriers, we explored apoC-III production and metabolism in WT mice. Mice expressing WT and A43T apoC-III had equivalent levels of total hepatic apoC-III protein, as assessed by immunoblotting (Fig. 2m and Supplementary Fig. 8). To deter-mine whether the A43T variant affected apoC-III secretion from the liver, we administered [35S]methionine and measured the secretion of newly synthesized, metabolically labeled apoC-III into the circulation. There was no difference in the secretion rate of WT mice expressing A43T as compared to those expressing WT apoC-III (Fig. 2n,o).
Because the secretion rates for WT and A43T apoC-III did not differ, we tested the hypothesis that A43T apoC-III is more rapidly catabolized than WT apoC-III. Apobec1-knockout mice expressing WT or A43T APOC3 were administered [125I]tyramine cellobiose (TC)-modified WT or A43T apoC-III protein, respectively, after pre-incubation with plasma from the corresponding mice. A43T apoC-III exhibited markedly increased catabolism as compared to WT apoC-III (Fig. 3a), with a >3-fold increase in the fractional catabolic rate (FCR) (Fig. 3b). These results were also observed in WT mice admin-istered 125I-labeled apoC-III protein without the TC modification (Supplementary Fig. 9a,b), suggesting that the observed differences in catabolism were not a result of conformational changes caused by the TC moiety.
ApoC-III clearance is thought to occur largely through two path-ways: receptor-mediated remnant lipoprotein clearance by the liver and filtration of unbound apoC-III by the kidney17,34–36. However, the relative contributions of these two clearance pathways and how they affect the functional pool of circulating apoC-III that promotes TRL retention remain unclear. We compared tissue uptake of [125I]TC-modified WT and A43T apoC-III into the liver and kidney 24 h after injection and found that A43T apoC-III was cleared ~40% less by the liver and ~40% more by the kidney as compared to WT apoC-III (Fig. 3c,d). Additional clearance studies with [125I]apoC-III lacking the TC moiety confirmed that the WT and A43T proteins were degraded and that the degradation products of the A43T protein were cleared into the urine at a substantially higher rate than those of the WT protein (Supplementary Fig. 9c–f). Indeed, in mice injected with labeled A43T apoC-III protein, TCA-precipitable 125I counts in the plasma at 1 min, 3 h, and 6 h after injection were significantly lower than those in mice injected with WT apoC-III (Supplementary Fig. 9g,h). These data suggest that the mutant protein is degraded more rapidly than the WT protein, and that [125I]tyrosine-labeled degradation products are filtered by the kidney and excreted.
The predicted helical structure of apoC-III places the A43 residue on the hydrophobic face of an α-helix that mediates binding to lipids32 (Supplementary Fig. 10). We hypothesized that the substitution of a hydrophilic threonine residue for the relatively more hydrophobic alanine residue at this position could impair lipoprotein binding, lead-ing to a relative increase in non-lipoprotein-bound, ‘free’ apoC-III that is more rapidly cleared. To test this, we fractionated plasma from mice administered [125I]TC-modified WT or A43T apoC-III protein and measured the distribution of apoC-III in lipoprotein. In comparison with WT apoC-III, much less A43T apoC-III was bound to TRL and
HDL, and relatively more was found in the lipoprotein-free fraction (Fig. 3e). Consistent with these findings, A43T apoC-III exhibited significantly reduced binding to TRL and HDL and increased presence in the free form after in vitro incubation with human plasma at 37 °C, as com-pared to WT apoC-III (Fig. 3f and Supplementary Fig. 11a–c). We also tested apoC-III binding to isolated human lipoproteins and lipid emul-sions. In comparison to WT apoC-III, A43T apoC-III exhibited signifi-cantly reduced binding to TRLs (Fig. 3g and Supplementary Fig. 11d), HDLs (Fig. 3h and Supplementary Fig. 11e), and apolipoprotein-free lipid emulsions (Supplementary Fig. 11f,g), suggesting that the bind-ing defect exhibited by A43T apoC-III is related to its interaction with lipid surfaces. Consistent with this notion, we found that, as compared to WT apoC-III, A43T apoC-III exhibited a ~4-fold higher dissocia-tion constant for binding to an immobilized surface of phospholipids in vitro, as assessed by surface plasmon resonance (Fig. 3i). The A43T mutant was also less capable than WT apoC-III of inhibition of LPL activity on large TG-rich emulsions in vitro (Supplementary Fig. 12a,b) and an Intralipid TG substrate (2.7-fold increased half-maximal inhibi-tory concentration (IC50) for A43T as compared to WT apoC-III, P < 0.001, Student’s unpaired t test; Supplementary Fig. 12c,d).
On the basis of these data, we postulated that it might be possible to use monoclonal antibodies to lipid-associated human apoC-III to therapeutically target apoC-III by promoting its dissociation from lipoproteins and clearance. We accordingly generated a set of human-ized monoclonal antibodies and screened them for their ability to inhibit apoC-III function, as assessed by abrogation of the inhibitory effect of human apoC-III on DiI-labeled VLDL uptake by HepG2 hepatocytes. One of the antibodies tested, STT505, potently abrogated human apoC-III function in this assay (Supplementary Fig. 13a) and had no cross-reactivity to mouse apoC-III (data not shown). In WT mice expressing human WT APOC3, a single intravenous dose of STT505 (25 mg of IgG per kg body weight) resulted in a robust (~75%) reduction in plasma apoC-III levels at 4 h after administration, relative to saline-administered mice (Fig. 4a–c and Supplementary Fig. 13b,c). This reduction in plasma apoC-III levels was maintained over the course of 7 h after dosing; at 24 h after administration, a reduction of ~34% was still observable. The reduction in plasma apoC-III levels was also associated with a concomitant reduction in circulating apoB levels over the course of 24 h after antibody administration (Fig. 4d and Supplementary Fig. 13d). Human-APOC3-expressing mice gavaged with olive oil immediately after administration of STT505 displayed a robust suppression in postprandial plasma apoC-III (Fig. 4e,f) and TG (Fig. 4g,h) levels, suggesting that the reduced levels of circulating apoC-III following antibody administration result from enhanced postprandial TG clearance.
To extend the duration of the efficacy of STT505 in reducing circu-lating apoC-III levels, we attempted to develop a version of STT505 that has high affinity to apoC-III at pH 7.4 but that dissociates from its antigen at acidic pH, such that it could be recycled from intracellular endosomes. To achieve this, we replaced two residues in the com-plementarity-determining region of STT505 with histidine residues. Administration of this antibody, STT5058, to mice expressing WT APOC3 (25 mg of IgG per kg body weight, subcutaneously) resulted in a sustained ~40–60% lowering of circulating apoC-III levels, as observed over the course of 28 d after a single dose (Fig. 4i,j and Supplementary Fig. 13e). This antibody achieved almost tenfold higher plasma IgG levels than the STT505 antibody, and STT5058 plasma levels were maintained for 20 d after administration of a single dose (Supplementary Fig. 13f). STT5058 administration also resulted in a ~25% reduction in circulating apoB levels over the course of

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
� advance online publication nature medicine
28 d (Fig. 4k and Supplementary Fig. 13g). To better understand how STT5058 treatment led to a sustained reduction in circulating apoC-III levels, we measured the turnover of [125I]TC-modified WT apoC-III protein over the course of 21 h in APOC3-expressing mice treated with either STT5058 or control antibody 24 h before iodinated apoC-III administration. STT5058-treated mice displayed accelerated clearance of radiolabeled apoC-III, which was most notable at 1 h after radi-olabeled apoC-III administration (Fig. 4l,m). At 21 h after [125I]TC apoC-III administration, we measured the uptake of labeled apoC-III into the liver, kidney, and spleen and found a marked, ~2-fold increase in splenic uptake in mice treated with STT5058 antibody relative to those treated with control antibody (Fig. 4n). In contrast, we found no difference in hepatic clearance of [125I]TC-modified apoC-III between mice treated with STT5058 antibody and control antibody and decreased renal clearance in STT5058-treated mice (Fig. 4o,p). These data suggest that the STT5058 antibody lowers circulating apoC-III through enhancement of apoC-III clearance from the cir-culation that can be partially explained by increased splenic uptake and results in a sustained reduction in steady-state apoC-III levels.
Taken together, our data reveal how the naturally occurring APOC3 A43T missense variant and an anti-apoC-III monoclonal antibody each reduce circulating apoC-III levels and, thereby, TRL levels (Fig. 4q). Our results have several implications regarding the mechanisms by which apoC-III is metabolized that are relevant to the therapeutic targeting of apoC-III. Previous studies of apoC-III kinetics in limited numbers of human subjects who were administered apoC-III labeled with stable isotopes have suggested a complex metabolism of circulat-ing apoC-III (ref. 22). Here we show that the kidney and liver are the predominant mediators of both WT and A43T apoC-III uptake from the circulation in mice. Furthermore, through study of the A43T vari-ant that promotes apoC-III dissociation from lipoproteins in vivo, we show that decreased apoC-III binding to lipoproteins increases the pool of free apoC-III and augments apoC-III catabolism, thus reducing circulating apoC-III levels. In particular, we found that the kidney is a major site of apoC-III degradation, suggesting that glomerular filtra-tion of free apoC-III and subsequent proximal tubular cell degradation are likely critical mediators of apoC-III clearance. In contrast, hepatic apoC-III clearance is postulated to occur via internalization of TRLs and remnants that carry apoC-III (refs. 6,36). TRL-bound apoC-III has previously been demonstrated to inhibit the clearance of these particles by the liver through the LDL receptor and related receptors of the same family. Our studies using monoclonal antibodies target-ing apoC-III support the notion that alternative, non-renal pathways of apoC-III clearance might also be leveraged to lower apoC-III and thereby TRL levels. The physiological fate of the apoC-III cleared by these antibodies is an important area for future study, as are the conse-quences of antibody-mediated apoC-III lowering on TRL production, remodeling, and turnover in vivo.
This work provides a paradigmatic example of how rare genetic variants can inform on protective mechanisms for drug development. Specifically, the discovery that A43T mutant apoC-III impairs lipo-protein binding and promotes plasma apoC-III catabolism led us to the notion that a therapeutic antibody could mimic this phenomenon. This strategy is similar to that taken for antibody targeting of PCSK9 to reduce LDL-C levels, a therapeutic approach that emerged from human genetics studies of nonsynonymous PCSK9 coding variants37. Several other nonsynonymous variants in genes regulating the LPL pathway of TRL clearance such as ANGPTL4 (refs. 38,39) and APOA5 (ref. 40) have recently been identified through similar large sequencing efforts, providing additional potential opportunities for clinical translation
through exploration of the underlying mechanisms by which these variants alter protein function.
MeTHODSMethods, including statements of data availability and any associated accession codes and references, are available in the online version of the paper.
Note: Any Supplementary Information and Source Data files are available in the online version of the paper.
ACKNoWLeDGMeNTSThe authors thank A. Wilson, E. Edouard, J. McParland, M. McCoy, K. Trindade, S. DerOrhannessian, M. Risman, K. Burton, and M. Sun for technical expertise and M. Lazar, M. Bucan, Z. Arany, B. Garcia, N. Hand, D. Marchadier, D. Conlon, and R. Bauer for helpful discussions. This work was supported in part by NIH grants R01HL133502 and R37HL055323 and a grant from the Foundation Leducq CVGeneF(x) Transatlantic Network of Excellence to D.J.R. and by NIH grant F30HL124967 to S.A.K. This project also used the UPCI Cancer Proteomics Facility, which is supported in part by NIH award P30CA047904. Recruitment to the Penn Medicine BioBank was supported by the Penn Cardiovascular Institute, the Perelman School of Medicine of the University of Pennsylvania, and a gift from the Smilow family.
AUTHoR CoNTRIBUTIoNSS.A.K., J.S.M., C.V., A.V.H.S., and P.Z. performed experiments on the A43T variant. S.A.K., X.Z., Z.S., and N.A.Y. performed selective reaction monitoring measurements from human plasma samples. S.A.K., P.D.-J., J.A.L., N.B., W.J.Z., I.T., and H.d.H. designed and performed experiments on anti-apoC-III monoclonal antibodies. G.M.P., P.N., M.C., S.L.-K., M.C.P., A.R.T., and S.K. provided guidance with study design. S.A.K. and D.J.R. secured funding, conceived and designed experiments, interpreted all results, and wrote the manuscript. All authors provided input on the manuscript.
CoMPeTING FINANCIAL INTeReSTSThe authors declare competing financial interests: details are available in the online version of the paper.
Reprints and permissions information is available online at http://www.nature.com/reprints/index.html. Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
1. Musunuru, K. & Kathiresan, S. Surprises from genetic analyses of lipid risk factors for atherosclerosis. Circ. Res. 118, 579–585 (2016).
2. Khetarpal, S.A., Qamar, A., Millar, J.S. & Rader, D.J. Targeting ApoC-III to reduce coronary disease risk. Curr. Atheroscler. Rep. 18, 54 (2016).
3. Jørgensen, A.B., Frikke-Schmidt, R., Nordestgaard, B.G. & Tybjærg-Hansen, A. Loss-of-function mutations in APOC3 and risk of ischemic vascular disease. N. Engl. J. Med. 371, 32–41 (2014).
4. Crosby, J. et al. Loss-of-function mutations in APOC3, triglycerides, and coronary disease. N. Engl. J. Med. 371, 22–31 (2014).
5. Pollin, T.I. et al. A null mutation in human APOC3 confers a favorable plasma lipid profile and apparent cardioprotection. Science 322, 1702–1705 (2008).
6. Aalto-Setälä, K. et al. Mechanism of hypertriglyceridemia in human apolipoprotein (apo) CIII transgenic mice. Diminished very low density lipoprotein fractional catabolic rate associated with increased apo CIII and reduced apo E on the particles. J. Clin. Invest. 90, 1889–1900 (1992).
7. Aalto-Setälä, K. et al. Further characterization of the metabolic properties of triglyceride-rich lipoproteins from human and mouse apoC-III transgenic mice. J. Lipid Res. 37, 1802–1811 (1996).
8. de Silva, H.V. et al. Overexpression of human apolipoprotein C-III in transgenic mice results in an accumulation of apolipoprotein B48 remnants that is corrected by excess apolipoprotein E. J. Biol. Chem. 269, 2324–2335 (1994).
9. Eisenberg, S., Patsch, J.R., Sparrow, J.T., Gotto, A.M. & Olivecrona, T. Very low density lipoprotein. Removal of apolipoproteins C-II and C-III-1 during lipolysis in vitro. J. Biol. Chem. 254, 12603–12608 (1979).
10. Masucci-Magoulas, L. et al. A mouse model with features of familial combined hyperlipidemia. Science 275, 391–394 (1997).
11. Chan, D.C., Watts, G.F., Nguyen, M.N. & Barrett, P.H. Apolipoproteins C-III and A-V as predictors of very-low-density lipoprotein triglyceride and apolipoprotein B-100 kinetics. Arterioscler. Thromb. Vasc. Biol. 26, 590–596 (2006).
12. Chan, D.C., Watts, G.F., Redgrave, T.G., Mori, T.A. & Barrett, P.H. Apolipoprotein B-100 kinetics in visceral obesity: associations with plasma apolipoprotein C-III concentration. Metabolism 51, 1041–1046 (2002).
13. Olivieri, O. et al. Apolipoprotein C-III, metabolic syndrome, and risk of coronary artery disease. J. Lipid Res. 44, 2374–2381 (2003).

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
l e t t e r s
nature medicine advance online publication �
14. Ooi, E.M., Barrett, P.H., Chan, D.C. & Watts, G.F. Apolipoprotein C-III: understanding an emerging cardiovascular risk factor. Clin. Sci. (Lond.) 114, 611–624 (2008).
15. Caron, S. et al. Transcriptional activation of apolipoprotein CIII expression by glucose may contribute to diabetic dyslipidemia. Arterioscler. Thromb. Vasc. Biol. 31, 513–519 (2011).
16. Ginsberg, H.N. & Brown, W.V. Apolipoprotein CIII: 42 years old and even more interesting. Arterioscler. Thromb. Vasc. Biol. 31, 471–473 (2011).
17. Ooi, E.M. et al. Plasma apolipoprotein C-III metabolism in patients with chronic kidney disease. J. Lipid Res. 52, 794–800 (2011).
18. Qamar, A. et al. Plasma apolipoprotein C-III levels, triglycerides, and coronary artery calcification in type 2 diabetics. Arterioscler. Thromb. Vasc. Biol. 35, 1880–1888 (2015).
19. Cohn, J.S. et al. Plasma turnover of HDL apoC-I, apoC-III, and apoE in humans: in vivo evidence for a link between HDL apoC-III and apoA-I metabolism. J. Lipid Res. 44, 1976–1983 (2003).
20. Nguyen, M.N. et al. Use of Intralipid for kinetic analysis of HDL apoC-III: evidence for a homogeneous kinetic pool of apoC-III in plasma. J. Lipid Res. 47, 1274–1280 (2006).
21. Ginsberg, H.N. & Ramakrishnan, R. Kinetic studies of the metabolism of rapidly exchangeable apolipoproteins may leave investigators and readers with exchangeable results. Arterioscler. Thromb. Vasc. Biol. 28, 1685–1686 (2008).
22. Ginsberg, H.N. & Ramakrishnan, R. Investigations of apoC-III metabolism using stable isotopes: what information can you acquire and how can you interpret your results? J. Lipid Res. 52, 1071–1072 (2011).
23. Mauger, J.F., Couture, P., Bergeron, N. & Lamarche, B. Apolipoprotein C-III isoforms: kinetics and relative implication in lipid metabolism. J. Lipid Res. 47, 1212–1218 (2006).
24. Shin, M.J. & Krauss, R.M. Apolipoprotein CIII bound to apoB-containing lipoproteins is associated with small, dense LDL independent of plasma triglyceride levels in healthy men. Atherosclerosis 211, 337–341 (2010).
25. Yang, X. et al. Reduction in lipoprotein-associated apoC-III levels following volanesorsen therapy: phase 2 randomized trial results. J. Lipid Res. 57, 706–713 (2016).
26. Pechlaner, R. et al. Very-low-density lipoprotein-associated apolipoproteins predict cardiovascular events and are lowered by inhibition of APOC-III. J. Am. Coll. Cardiol. 69, 789–800 (2017).
27. Wyler von Ballmoos, M.C., Haring, B. & Sacks, F.M. The risk of cardiovascular events with increased apolipoprotein CIII: a systematic review and meta-analysis. J. Clin. Lipidol. 9, 498–510 (2015).
28. Nagashima, K. et al. Effects of the PPARγ agonist pioglitazone on lipoprotein metabolism in patients with type 2 diabetes mellitus. J. Clin. Invest. 115, 1323–1332 (2005).
29. Willer, C.J. et al. Discovery and refinement of loci associated with lipid levels. Nat. Genet. 45, 1274–1283 (2013).
30. Graham, M.J. et al. Antisense oligonucleotide inhibition of apolipoprotein C-III reduces plasma triglycerides in rodents, nonhuman primates, and humans. Circ. Res. 112, 1479–1490 (2013).
31. Gaudet, D. et al. Antisense inhibition of apolipoprotein C-III in patients with hypertriglyceridemia. N. Engl. J. Med. 373, 438–447 (2015).
32. Sundaram, M. et al. Functional analysis of the missense APOC3 mutation Ala23Thr associated with human hypotriglyceridemia. J. Lipid Res. 51, 1524–1534 (2010).
33. Millar, J.S., Cromley, D.A., McCoy, M.G., Rader, D.J. & Billheimer, J.T. Determining hepatic triglyceride production in mice: comparison of poloxamer 407 with Triton WR-1339. J. Lipid Res. 46, 2023–2028 (2005).
34. Malmendier, C.L., Lontie, J.F., Grutman, G.A. & Delcroix, C. Metabolism of apolipoprotein C-III in normolipemic human subjects. Atherosclerosis 69, 51–59 (1988).
35. Malmendier, C.L. et al. Apolipoproteins C-II and C-III metabolism in hypertriglyceridemic patients. Effect of a drastic triglyceride reduction by combined diet restriction and fenofibrate administration. Atherosclerosis 77, 139–149 (1989).
36. Gordts, P.L. et al. ApoC-III inhibits clearance of triglyceride-rich lipoproteins through LDL family receptors. J. Clin. Invest. 126, 2855–2866 (2016).
37. Kathiresan, S. Developing medicines that mimic the natural successes of the human genome: lessons from NPC1L1, HMGCR, PCSK9, APOC3, and CETP. J. Am. Coll. Cardiol. 65, 1562–1566 (2015).
38. Dewey, F.E. et al. Inactivating variants in ANGPTL4 and risk of coronary artery disease. N. Engl. J. Med. 374, 1123–1133 (2016).
39. Stitziel, N.O. et al. Coding variation in ANGPTL4, LPL, and SVEP1 and the risk of coronary disease. N. Engl. J. Med. 374, 1134–1144 (2016).
40. Do, R. et al. Exome sequencing identifies rare LDLR and APOA5 alleles conferring risk for myocardial infarction. Nature 518, 102–106 (2015).

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
nature medicine doi:10.1038/nm.4390
ONLINe MeTHODSEthics statement. This study was performed in accordance with the principles of the Declaration of Helsinki, and all procedures were approved by the Institutional Review Board (IRB) of the Perelman School of Medicine at the University of Pennsylvania. All participants provided written informed consent.
Human subject identification and ascertainment. Human participants were recruited from two independent cohorts for ascertainment of APOC3 loss-of-function variants and subsequent phenotyping. The first cohort, the Penn High HDL Study (HHDL), is a cross-sectional study of heritable factors con-tributing to elevated levels of HDL-C. Participants were recruited with HDL-C levels greater than the 95th percentile for age and sex by physician referrals or through the Hospital of the University of Pennsylvania clinical lipid labora-tory. Relatives of probands with high HDL-C were also invited to participate in the study. Participants were consented for additional plasma lipid analysis and research genetic studies including targeted sequencing of candidate genes, exome sequencing, and SNP array genotyping under a protocol approved by the IRB of the University of Pennsylvania. Over 2,500 participants have been recruited for the HHDL study, with recruitment ongoing. The second cohort, the Penn Medicine BioBank, is a collection of banked plasma samples from participants who were treated at the University of Pennsylvania Health System and were consented for genetic analysis and biochemical studies of collected plasma for the study of heritable risk factors and novel biomarkers related to CHD risk. Approximately 50,000 participants have been recruited so far, with recruitment of additional participants ongoing through IRB-approved protocols of the University of Pennsylvania. Demographic characteristics for carriers and non-carriers from each identification cohort were as follows: Penn Medicine BioBank cohort: A43T carriers: n = 6, female n = 2, male n = 4, age = 61.7 ± 11.9 years, Caucasian n = 5, African-American n = 1; non-carriers: n = 54, female n = 18, male n = 36, age = 60.2 ± 6.6 years, Caucasian n = 48, African-American n = 6; HHDL cohort: A43T carriers: n = 13, female n = 11, male n = 2, age = 60.4 ± 9.58 years, Caucasian n = 12, African-American n = 1; non-carriers: n = 22, female n = 18, male n = 4, age = 60.9 ± 10.1 years, Caucasian n = 18 African-American n = 4. Age is expressed as mean ± s.d. for each group.
For both studies, genomic DNA was isolated from whole blood from partici-pants and prepared for genotyping using the Exome Chip SNP genotyping array (Human Exome BeadChip v1.0, Illumina). The Exome Chip is a commercially available platform that contains more than 240,000 previously identified SNP markers, mostly corresponding to coding regions across the genome that are derived from variants found two or more times across more than one data set among 23 unique data sets of >12,000 exome or whole-genome sequences41. In particular, the Exome Chip includes the APOC3 A43T (rs147210663) variant. After removing subjects from analysis for whom genotyping quality control measures failed or for whom demographic covariates were not available, a total of 1,056 participants from the HHDL study and 5,744 subjects from the Penn Medicine BioBank cohort were genotyped. APOC3 A43T variant genotypes identified using Exome Chip genotyping were confirmed by Sanger sequencing, as follows. A 674-bp fragment of the APOC3 gene sequence was amplified by PCR from 200 ng of genomic DNA from identified variant carriers using the following oligonucleotide primers: F 5′-CTCCTTCTGGCAGACCCAGCTAAGG-3′, R 5′-CCTAGGACTGCTCCGGGGAGAAAG-3′. PCR products were puri-fied using a QIAquick PCR purification kit (Qiagen) and Sanger sequenced by Genewiz. Sequence chromatograms were aligned to the APOC3 reference sequence in NCBI using Sequencher (Gene Codes).
Lipid phenotyping of human participants. Plasma samples of carriers of the APOC3 A43T coding variant, along with plasma samples of non-carriers from the same identification cohorts matched for age, sex, and ancestry, were selected for lipid analysis. The number of non-carrier samples analyzed was chosen on the basis of a power estimate for identifying a 50% difference in plasma apoC-III levels, with an s.d. equivalent to the difference in means. From this estimate, a minimum of 16 carriers and 16 non-carriers were required for the analysis. Given the identification of 19 A43T carriers, we searched for at least an equivalent number of matched non-carriers, and ultimately identified 76 non-carriers for the lipid and apoC-III analyses described here.
Plasma total cholesterol, HDL-C, TG, apoC-III, apoA-I, apoB, and apoE levels were measured using an Axcel clinical autoanalyzer (Alfa Wassermann Diagnostic Technologies). Non-HDL-C was calculated by the difference in the TC and HDL-C measurements. All measurements were made from overnight-fasted plasma samples.
Determination of apoC-III A43T isoforms in human plasma via LC–MS/MS. Overview of the targeted mass spectrometry assay. There is no commer-cially available immunoassay that is capable of distinguishing WT versus A43T mutant apoC-III. Therefore, we took advantage of the power of targeted mass spectrometry to provide allele-specific quantification of apoC-III in plasma samples. The general workflow of our targeted mass spectrometry assay and data analysis is illustrated in Supplementary Figure 2. In brief, we used AspN protease to digest plasma proteins into proteolytic peptides. Because of the presence of various levels of methionine oxidation in selected apoC-III pep-tides in different samples (data not shown), the resultant AspN peptides were treated with hydrogen peroxide to oxidize all methionine residues. Heavy-iso-tope-labeled synthetic peptides corresponding to both WT (peptide A43) and A43T (peptide T43) AspN peptides containing the A43T residue were added to each digestion to control for variability in sample preparation and mass spec-trometer performance (Supplementary Fig. 3). Mouse plasma samples with known amounts of WT and A43T mutant human apoC-III were processed using the same workflow and used as external calibrators (Supplementary Fig. 4 and Supplementary Table 2). The light/heavy ratio was calculated for each peptide as the peak area of endogenous peptide divided by the peak area of internal isotope-labeled peptide (Supplementary Fig. 3). Linear regression was used to define the targeted assay response (light/heavy ratio) in com-parison to the expected WT and A43T mutant apoC-III concentrations. The derived standard curves were used to determine the concentration of WT or mutant apoC-III in human plasma samples.
Sample preparation for the LC–MS/MS assay. Fasting plasma from A43T carriers (n = 19) and non-carriers (n = 21) was enzymatically digested to pep-tides and analyzed by LC–MS/MS to quantify the abundance of WT and A43T mutant peptides. Plasma samples were diluted 50-fold with 50 mM ammonium bicarbonate, and an aliquot containing the equivalent of 150 nl of plasma was biochemically processed for LC–MS/MS analysis.
Isotopically labeled internal standard peptides and exogenous proteins were added to each plasma sample before reduction with dithiothreitol (DTT), oxi-dation with hydrogen peroxide, and digestion with AspN. Mass-spectrometry-grade AspN (Thermo Fisher Scientific) was used to generate proteolytic peptides for LC–MS/MS analysis. For each measurement, a 150-nl equivalent of mouse or human plasma in 80 µl of 50 mM NH4HCO3 was mixed with heavy-isotope-labeled synthetic peptides covering the apoC-III A43T locus (2.5 pmol for WT A43 peptide and 5 pmol for mutant T43 peptide) (Supplementary Figs. 4 and 5, and Supplementary Table 3). 1.25 pmol of ovalbumin (SERPINB14) was spiked into each sample before digestion to monitor the reproducibility of sample diges-tion. The mixture was then boiled for 10 min in the presence of 10 mM DTT and alkylated by incubating in the presence of 45 mM iodoacetamide for 1 h in the dark at room temperature. 0.4 µg of AspN enzyme was then added, and diges-tion was performed overnight at 37 °C. PepClean C-18 spin columns (Thermo Fisher Scientific) were used for peptide cleanup. To oxidize the methionine residues of the resultant AspN peptides, the dried peptides were resuspended in 20 µl of 20 mM hydrogen peroxide and incubate at 4 °C overnight. Samples were dried again in a SpeedVac and reconstituted in 15 µl of 0.1% formic acid for LC–MS/MS assays.
LC–MS/MS assay for apoC-III. Selective reaction monitoring was performed on a TSQ Quantiva triple-stage quadrupole mass spectrometer (Thermo Fisher Scientific) coupled to an UltiMate 3000 Nano LC Systems (Thermo Fisher Scientific). Mobile phase A consisted of 0.1% formic acid in HPLC water, and mobile phase B consisted of 0.1% formic acid in 100% acetonitrile. A 3-µl injec-tion volume of AspN peptides, equivalent to 30 nl of plasma, was loaded onto a C18 trap column (Dionex), washed with mobile phase A at 6 µl/min for 2 min. Nano-LC separation was carried out on an analytical C18 PicoChip column packed with 10.5 cm of Reprosil C18 3-µm 120-Å chromatography medium with a 75-µm-ID column and a 15-µm tip (New Objective) at a flow rate of 400 nl/min with a gradient comprising 2–8% solvent B for 1 min, 8–35% solvent B

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
nature medicinedoi:10.1038/nm.4390
for 42 min, 35–95% solvent B for 3 min, and 95% solvent B for 10 min. Collision energies were calculated using the linear equation CE = 0.034 × m/z + 3.314. The full width at half maximum was set to be 0.7 Da for Q1 and Q3. The instrument was operated using unscheduled SRM mode with 1.5-s cycling. The majority of peptides had base peak widths of ~30 sec/base, and ~20 data points were acquired per chromatogram peak.
An open-source data analysis package (University of Washington, Skyline) was used to facilitate targeted SRM assay method development and data analy-ses42. All selected apoC-III peptides and their corresponding SRM assay param-eters are listed in Supplementary Table 3. The peak area of each analyte was the sum of the peak areas of all transitions. Total mutant protein concentrations were measured by fitting LC–MS/MS selective-reaction-monitoring-derived relative amounts of mutant to WT protein to the total plasma apoC-III concentrations measured by chemical autoanalyzer.
In silico analysis of the splicing effects of the APOC3 A43T variant. The impact of the APOC3 A43T coding variant on APOC3 exon splicing was assessed using the Automated Splice Site and Exon Definition Analyses (ASSEDA) server (MutationForcaster; https://www.mutationforecaster.com/learn.php#asseda; ref. 43).
Cloning of human APOC3 cDNA and adeno-associated virus genera-tion. The cDNA of the human APOC3 gene (NM_000040) was obtained from OriGene and cloned in the pcDNA3.1/V5-His TOPO expression plasmid (Thermo Fisher Scientific). The A43T variant was introduced into the pcDNA3.1/V5-His plasmid containing WT APOC3 cDNA using the QuikChange II Site-Directed Mutagenesis kit (Agilent Technologies) with the following primers: F 5′-GCACGCCACCAAGACCACCAAGGATGCACTGAGCAG-3′, R 5′-CTGCTCAGTGCATCCTTGGTGGTCTTGGTGGCGTGC-3′, where the underlined nucleotides indicate the position of the variant in the primer sequences. The WT and A43T variant APOC3 cDNA sequences were further amplified by PCR to introduce KpnI and SalI restriction sites using the following primers: F 5′-ACGCGTGGTACCATGCAGCCCCGGGTACTCCTTG-3′, R 5′-CCGCCCGGGTCGACTCAGGCAGCCACGGCTGAAG-3′. Amplified PCR products corresponding to the WT and A43T variant cDNA sequences of APOC3 were digested with KpnI and SalI and ligated with an AAV serotype 8 vector plasmid containing the liver-specific thyroxine-binding globulin (TBG) promoter, which was provided by the Vector Core of the University of Pennsylvania44–47. The DNA sequence after ligation of the AAV plasmid was confirmed by Sanger sequencing and by restriction digestion and agarose gel electrophoresis analysis. AAV production, purification, and titer-ing from the cloned WT and A43T APOC3 vector plasmids were performed by the Vector Core.
125I radiolabeling of apoC-III. Full-length forms of WT and A43T variant apoC-III were produced by solid-state peptide synthesis by Pierce Custom Peptides (Thermo Fisher Scientific). Purified peptides were confirmed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry to be of >95% purity. Proteins were solubilized in 10 mM ammonium bicarbo-nate buffer, pH 7.4, to a concentration of approximately 0.2–0.4 mg/ml.
Solubilized proteins were iodinated with 125I directly using the iodine mono-chloride method48 or through incorporation with [125I]TC49. For direct 125I labeling of apoC-III by the ICl method, approximately 0.5 ml of apoC-III (Pierce Custom Peptides, Thermo Fisher Scientific) in 10 mM ammonium bicarbonate buffer (0.2–0.4 mg/ml) was iodinated with 1 mCi of 125I (PerkinElmer), 300 µl of 1 M glycine, and 150 µl of 1.84 M NaCl/2.84 µM ICl solution, vortexed, and applied to a PG-10 desalting column (Amersham Biosciences) that was pre-equilibrated with 0.15 M NaCl/1 mM EDTA solution. Iodinated proteins were eluted in a final volume of 2.5 ml in NaCl/EDTA solution and dialyzed against PBS before measurement of protein concentration by BCA assay and 125I activity by gamma counting.
For iodination of TC, 180 µl of 0.4 M sodium phosphate, Pierce iodination beads (Thermo Fisher Scientific), and 1 mCi of 125I were combined, and 20 µl of 0.2 µM TC (a kind gift from D. Usher and W. Cain, University of Delaware) was then added. This mixture was incubated at room temperature for 30 min with shaking every 5 min, and 10 µl of 0.1 M NaI and 20 µl of 0.1 M NaHSO3 were
added to stop the reaction. Next, 40 µl of 0.2 µM cyanuric chloride was added to activate the [125I]TC. The activated [125I]TC was then added to the apoC-III protein samples (solubilized in ammonium bicarbonate buffer at a concentra-tion of 0.2–0.4 mg/ml), and reactions were incubated at room temperature for 3 h. Finally, the reaction was fractionated over a PD-10 desalting column to remove unbound iodine, and fractions containing iodinated protein were pooled together and dialyzed against PBS for further use.
AAV WT versus A43T APOC3 expression studies in mice. C57BL/6 WT mice were purchased from The Jackson Laboratory (000664). Additionally, Apoc3-knockout mice on a C57BL/6 background (002057) were recov-ered from cryopreservation from The Jackson Laboratory and bred at the University of Pennsylvania. Apoc3-knockout mice were bred with Apobec1-deficient, human-APOB-transgenic mice that either were WT (LahB WT), heterozygous (LahB Het), or deficient (LahB KO) for the mouse Ldlr gene. Mice were maintained in a monitored small animal facility at the University of Pennsylvania under IACUC-approved protocols and were fed ad libitum with a standard chow diet or Western diet containing 0.21% cholesterol from OpenSource Diets (D12079B, Research Diets) for the indicated periods of time. All mice were provided access to water ad libitum and were maintained with a 12-h on/12-h off light cycle with lights off from 7:00 p.m. to 7:00 a.m. daily. All blood samples from mice were collected by retro-orbital bleeding from mice anesthetized with isoflurane using EDTA-coated glass tubes under approved protocols. For all studies, male mice approximately 10–12 weeks of age were used. Mice were not randomized for any measure before use in AAV experiments, and experimenters were not blinded to group allocation for the described experiments. Six mice per group were used for all studies, on the basis of prior requirements for group sample sizes for measuring 50% mean differences in plasma TGs from previous studies in our laboratory.
For APOC3 AAV expression experiments in mouse models, mice were ini-tially fasted for 4 h, bled, and administered a control AAV serotype 8 vector lacking a transgene (Null), APOC3 WT AAV (WT), or APOC3 A43T (A43T), all at a dose of 3 × 1011 GC of virus per mouse as determined by digital-PCR-based titering by the Vector Core. AAVs were diluted in sterile PBS in a sterile chemical hood and administered by intraperitoneal injection using insulin syringes. For some experiments, CETP AAV, which was generated as described previously 50, was co-administered with Null or APOC3 AAVs at a dose of 3 × 1010 GC per mouse; for these experiments, appropriate amounts of CETP AAV vector were mixed with Null or APOC3 AAV vectors and PBS for administration by intra-peritoneal injection. Subsequent blood collection was performed after fasting the mice for 4 h and under isoflurane anesthesia at the indicated time points. Blood was separated to obtain plasma by centrifugation in a microcentrifuge at 4 °C at 10,600g for 7 min. All mouse experiments were performed with six mice per group unless otherwise noted.
Lipid and human apolipoprotein measurements were performed on mouse plasma using an Axcel autoanalyzer. In addition, for some experiments, plasma samples were pooled by experimental group freshly after collection and 150 µl of plasma was separated by FPLC on a Superose 6 gel-filtration column (GE Healthcare Life Sciences) into fractions each 0.5 ml in volume. total cholesterol and TG were measured from FPLC-separated fractions using Infinity Liquid Stable cholesterol and triglyceride reagents (Thermo Scientific) in 96-well micro-plates with a Synergy Multi-Mode Microplate Reader (BioTek).
Oral fat tolerance tests. OFTTs were performed in APOC3 WT, A43T, and Null AAV treated mice 4 weeks following AAV administration. For these experiments, mice were fasted for 12 h and weighed. Olive oil (Sigma-Aldrich) was prepared in 1.0-ml syringes at a volume of 10 µl × fasting body weight (in grams) for each mouse. Mice were bled before olive oil administration to measure pre-gavage plasma TGs after fasting and were then gavaged orally with olive oil and bled subsequently at 1, 3, 5, and 7 h after gavage. Plasma TGs at each time point were measured by colorimetric assay in 96-well microplates, as described above. For measurement of [3H]retinyl ester accumulation and clearance in the mouse groups described above, gavage with olive oil was per-formed as described in mice after overnight fast, but instead of volumes of 10 µl olive oil × fasting body weight (in grams), 250 µl was used for each mouse and was premixed with 10 µCi of [11,12-3H]retinol (PerkinElmer). Plasma was

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
nature medicine doi:10.1038/nm.4390
collected at 0, 1, 3, 5, and 7 h after gavage, and 3H activity in 10 µl of each plasma sample was used to assess [3H]retinyl ester accumulation. For the measurement of OFTTs in the presence of Poloxamer P407 (Pluronic, Sigma-Aldrich) treatment, the same procedure for treating the mice as described above was used, but, just before olive oil gavage with 10 µl olive oil × fasting body weight (grams), mice were administered a solution of Pluronic in PBS at a dose of 1 mg/kg body weight, administered intravenously by tail vein injec-tion. The time points and subsequent sample processing were otherwise the same as for the above experiments.
TRL clearance studies. For [3H]TRL clearance studies in mice, human TRLs were obtained from pooled human non-fasted plasma from the Hospital of the University of Pennsylvania by density-gradient ultracentrifugation in a Beckman XL-90 Ultracentrifuge (Beckman Coulter) using a 70.1-Ti rotor (Beckman Coulter) at a speed of 40,000 r.p.m. for 18 h. Ultracentrifugation of the plasma resulted in the formation of an upper white, cream-like TRL layer (d < 1.006), which was collected and subjected to another ultracentrifugation step, in the same conditions. The upper cream-like layer (d < 1.006) was col-lected, and the total protein concentration was measured using a bicinchoninic assay (Thermo Fisher Scientific). Approximately 3 mg of TRL protein was used for labeling with 0.5 mCi of [3H]triolein (PerkinElmer). 0.5 mCi of [3H]triolein in toluene was dried under a nitrogen evaporator and resolubilized in 150–200 µl of ethanol and then added dropwise to a mixture of TRLs (3 mg of protein) and lipoprotein-deficient human serum (d > 1.220, 100 mg of protein per 3 mg of TRL protein) in a glass tube. The mixture was then incubated at 37 °C with gentle shaking for 12–14 h. After addition of KBr solution (d = 1.006 g/ml), the mixture was subjected to an additional ultracentrifugation step, according to the protocol detailed above. Following ultracentrifugation, the cream-like top layer (approximately 2.5–3.0 ml) was collected, dialyzed against PBS, and fractionated by FPLC to measure TG and 3H activity in the TRL frac-tions (5–12) relative to unincorporated 3H activity. Dialyzed [3H]TRLs were then administered by intravenous tail vein injection into mice 5 weeks after administration of Null, WT, or A43T AAVs. Mice were bled at 1, 8, 15, 30, 60, and 120 min after [3H]TRL administration. 3H activity in 10 µl of plasma from each time point was measured by scintillation counting, and the relative 3H activity remaining in the circulation was calculated by normalizing the activity from each time point by that at 1 min for each mouse.
VLDL-TG, apoC-III, and apoB secretion studies. Hepatic VLDL secretion was measured in mice treated with Null or APOC3 AAVs using [35S]methionine metabolic labeling as previously described51. Briefly, mice injected with Null or APOC3 AAVs at 4 weeks after AAV administration were fasted for 4 h and then administered the surfactant Poloxamer P407 (Sigma-Aldrich) (25 mg in 0.4 ml of PBS) by intraperitoneal injection to inhibit peripheral lipolysis. 15 min after P407 administration, each mouse was administered 35S-labeled methionine (0.5 mCi in 0.2 ml of PBS, PerkinElmer) by intravenous tail vein injection. Mice were bled at 0, 30, 60, 90, and 120 min after radioisotope administration for plasma collection. Plasma TGs were measured at each time point with Infinity Triglycerides reagent (Thermo Fisher Scientific) in 96-well microplates. The plasma samples at 60 and 120 min were also used to measure the secretion rate of newly synthesized 35S-labeled apoB and apoC-III. For apoB measurements, 2 µl of each plasma sample from these time points was subjected to SDS–PAGE under reducing conditions using NuPAGE 3–8% Tris-acetate gels (Thermo Fisher Scientific). Gels were fixed in 25% methanol/20% acetic acid solution for 30 min and then with AutoFluor intensifier solution (National Diagnostics) for 30 min. Gels were dried using a vacuum pump for 4 h at 65 °C and exposed to autoradiography film for 1 week at −80 °C, and films were then developed. Visible bands on the film corresponding to apoB-100 and apoB-48 were marked and positions correlated on the dried gels, and those bands were excised from the gels, solubilized with Solvable reagent (0.5 ml; PerkinElmer) at 55 °C for 3 h, and 35S activity was measured by liquid scintillation counting. The activity of each band was normalized to the activ-ity measured after trichloroacetic acid precipitation of proteins from 5 µl of plasma from the same plasma sample used for apoB SDS–PAGE. Normalized 35S activity from the time points at 60 and 120 min was used to calculate the relative apoB-100 and apoB-48 secretion rates. For apoC-III secretion
measurements, the procedure followed was the same as described above for measurement of apoB secretion, except NuPAGE 4–12% Bis-Tris gels were used for SDS–PAGE instead of 3–8% Tris-acetate gels.
ApoC-III kinetic studies. For [125I]apoC-III clearance studies, iodinated apoC-III (specific activity of approximately 300,000–500,000 counts per minute (c.p.m.)/µg protein) was mixed with plasma from recipient mice at a ratio of 1 µg of 125I-labeled protein/200 µl of plasma. Iodinated apoC-III and mouse plasma mixtures were incubated at 37 °C for 1 h and were then admin-istered by intravenous tail vein injection into mice treated with either WT or A43T variant APOC3 4 weeks after they were administered APOC3 AAVs. Mice expressing WT APOC3 received 125I-labeled WT apoC-III protein, and those expressing A43T APOC3 received 125I-labeled A43T apoC-III. Mice were bled 1 min, 15 min, 30 min, 1 h, 3 h, 5 h, 9 h, and 24 h after radioisotope admin-istration and sacrificed at 24 h. 125I activity was measured from 10 µl of each plasma sample by counting on a Packard Cobra II Auto-Gamma counter. The relative activity at each time point was determined as the fraction of activity at 1 min for each mouse respectively. For experiments with [125I]TC-modified apoC-III, the above procedure was followed, and in addition liver and kidney were collected from mice at 24 h after perfusion with 10 ml of cold PBS. 30 mg of each tissue was homogenized in PBS and measured for 125I activity. Tissue 125I activity was normalized to the plasma 125I activity at 1 min for each mouse respectively. For experiments measuring urinary clearance of [125I]apoC-III, mice were administered 125I-labeled proteins in the manner above and mice were bled at 1 min, 3 h, 6 h, and 24 h after administration. For these experi-ments, mice were housed singly in rodent metabolic cages (Tecniplast) and urine was collected from 0–6 h, 6–12 h, and 12–24 h during the course of the study. Plasma and urine samples (20 µl of each sample) were counted for total 125I activity by gamma counting. In addition, proteins were separated by TCA precipitation from 20 µl of each sample, and the TCA-soluble (degraded) and TCA pellet (intact protein) fractions were counted for 125I activity.
Hepatic gene expression. For gene expression measurements, approximately 20 ng of liver tissues from each mouse was snap-frozen at −80 °C. Total RNA was extracted from tissues using TRIzol reagent (Thermo Fisher Scientific) according to the manufacturer’s protocol. Synthesis of cDNA was completed using 1 µg of RNA following extraction using the High-Capacity cDNA Reverse Transcription kit (Thermo Fisher Scientific). Quantitative real-time PCR was performed on the synthesized cDNA using a QuantStudio 7 Real-Time PCR System with the following TaqMan primer sets (Thermo Fisher Scientific): human APOC3, Hs00163644_m1; mouse Apoc3, Mm00445670_m1; mouse Actb (actin), Mm00607939_s1. The relative expression of human APOC3 and mouse Apoc3 in the liver was analyzed for each mouse by normalizing to actin gene expression from the same sample.
Hepatic AAV vector DNA content in each mouse was also assessed by quan-titative real-time PCR. Total hepatic DNA was isolated from 25 mg of liver for each mouse using the QIAamp DNA Mini kit (Qiagen). Isolated liver DNA was used for quantitative real-time PCR with TaqMan primers targeting the rabbit beta-globin poly(A) sequence of the AAV vector backbone. Cycle numbers from this assay were compared to those from a standard curve of serial dilutions of a linearized AAV plasmid of known quantity.
Immunoblotting. For immunoblotting of apoC-III from mouse liver sam-ples, 30 µg of liver was separated on 10% Bis-Tris NuPAGE gels using MES buffer by one-dimensional SDS–PAGE. Separated proteins were transferred to nitrocellulose membranes, and membranes were blocked with 5% fat-free milk in PBS (0.05% Tween-20) for 3 h. Membranes were then incubated for 2 h at room temperature with a rabbit polyclonal antibody against human apoC-III (33A-R1a, Academy Biomedical) at a dilution of 1:2,000, followed by three 15-min washes with PBS/Tween solution and incubation with goat anti-rabbit IgG HRP conjugate (sc-2030, Santa Cruz Biotechnology) for 30 min at room temperature. Membranes were again washed 3 × 15 min, and proteins were visualized with Luminata Crescendo chemiluminescent rea-gent (Millipore). Mouse β-actin, used as a loading control, was detected using mouse anti-actin primary antibody (sc-81178, Santa Cruz Biotechnology) at a dilution of 1:2,000; membranes were incubated with the antibody for 2 h

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
nature medicinedoi:10.1038/nm.4390
at room temperature, washed, and then incubated with goat anti-mouse IgG HRP conjugate (sc-2302, Santa Cruz Biotechnology) at a dilution of 1:5,000 for 30 min at room temperature.
Expression and purification of recombinant monoclonal antibodies to apoC-III (STT505 and STT5058). After selection from phage Fab antibody libraries, genes encoding the variable regions of antibodies scored positive for binding a central epitope of human apoC-III were sequenced and compared with each other (using Clone manager software, DeLano Scientific). Codon-optimized synthetic fragments encoding the different variable domains (VH and Vκ or Vλ) were ordered from Geneart. Recombinant antibodies were produced by U-Protein Express BV as follows: the synthetic DNA fragments encoding the variable domains were ligated into in-house-constructed expres-sion vectors, resulting in heavy chain–, Igκ- and Igλ-encoding expression con-structs. Equal amounts of the heavy chain–encoding vector were mixed with the light chain–encoding vector for cotransfection of HEK293E cells (ATCC). Cell lines were authenticated by ATCC by short-tandem-repeat profiling and karyotyping and were tested by both ATCC and our facility for mycoplasma contamination. Full-length recombinant antibodies were produced by tran-sient transfection of HEK293E cells grown in suspension at 0.1–15 l scale. Six days after transfection, conditioned medium was collected by centrifuga-tion and concentrated. The recombinant antibodies were purified via affinity chromatography using rmp Protein A Sepharose Fast Flow (GE Healthcare) and gel filtration chromatography using HiLoad 26/60 Superdex 200 pg (GE Healthcare), according to the manufacturer’s recommendations. Purity was determined using SDS–PAGE, and the concentration was determined by meas-urement of the optical density at 280 nm. Endotoxin levels were determined using a Limulus Amebocyte Lysate (LAL) assay (Thermo Fisher Scientific). Typical yields of the recombinant antibodies varied between 20 and 80 mg/l with a purity of at least 98% and with an endotoxin level below the detection limit. The STT5058 antibody is a modified version of the STT505 antibody, in which a serine-to-histidine substitution in the third residue of the heavy chain complementarity-determining region 2 (HCDR2) and a glutamine-to-histidine substitution in the second residue of the light chain complementa-rity-determining region 3 (LCDR3) were introduced. These modifications were incorporated to allow differential affinity of the antibody for the apoC-III epitope at pH 7.4 versus pH 5.5.
In vivo studies with STT505 and STT5058. All animal studies using STT505 and STT5058 were carried out in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All procedures were approved by the Institutional Animal Care and Use Committee of Vascumab, LLC, and the University of Pennsylvania. For STT505 antibody studies, 24 male C57Bl/6 mice (Charles River) weighing 20–25 g were maintained on a constant 12-h light/12-h dark cycle with free access to water and ad libitum access to a standard chow diet (Lab Diet, 5001). For overexpression of human WT APOC3, mice were injected intraperitoneally with an AAV vector expressing human APOC3 as described above. Twelve days after administration of 3 × 1011 GC/mouse, mice were bled via retro-orbital sinus and assigned to groups to achieve equivalent starting mean plasma apoC-III levels between groups. On day 14, mice were fasted for 6 h and retro-orbital sinus bled to measure lipid levels before antibody administration. The experi-ment was initiated by an intravenous tail injection of test antibody followed immediately by a 10 ml/kg oral bolus of olive oil (Sigma-Aldrich, 01514). Mice were bled via retro-orbital sinus to determine levels of plasma TGs, apoC-III, and test antibody at 15, 30, 60, 120, and 240 min following olive oil challenge. Measured values were plotted as a function of time and the AUC for plasma TGs was calculated using GraphPad Prism 6. For STT5058 antibody studies, 20 male C57Bl/6 mice were treated with WT APOC3 AAV and maintained as described above for the mice in the STT505 antibody studies. [125I]TC-modi-fied apoC-III was prepared as described above for the WT versus A43T studies, and the experiment was performed as follows. Three weeks after injection with WT APOC3 AAV, mice were administered STT5058 antibody or iso-type control antibody (anti–hen egg lysozyme antibody; HyHEL-5, U-Protein Express) (25 mg/kg body weight at time of dosing, in PBS) by injection into the subcutaneous layer of the back. 24 h later, the mice were administered
[125I]TC-modified WT apoC-III via tail vein injection as described above. Plasma was collected from these mice at 1 min, 15 min, 30 min, 60 min, 3 h, 5 h, 9 h, and 21 h after radioiodinated apoC-III administration. At 21 h after apoC-III administration, mice were sacrificed, perfused with 10 ml of cold PBS, and the liver, kidneys, and spleen were dissected for 125I activity measure-ments from 20 mg of each tissue in the manner described above for the A43T experiments. Tissue 125I activity was normalized to the plasma 125I activity at 1 min for each mouse respectively.
TGs were analyzed by incubating 5 µl of serum with 150 µl of Thermo Fisher Scientific Triglycerides Reagent (TR22421) supplemented with 200 µM Amplex Red (AAT Bioquest) in a black 96-well plate (Costar, 3915). After incubation for 10 min at 30 °C, the plate was read (excitation 560 nm, emission 585 nm), and the TG concentration was calculated from a four-parameter fit (Molecular Devices) of a glycerol standard curve.
A 96-well plate (Greiner, 655061) was coated overnight at 4 °C with 50 µl of primary antibody to apoC-III (Abcam rabbit polyclonal anti–human apoC-III, ab21032) diluted in PBS. The plate was washed four times with 200 µl of TBS-T and blocked with 200 µl of blocking buffer (Pierce Clear Milk Blocker, 37587; in PBS) for 90 min at 30 °C. The blocking buffer was removed, and 50 µl of test sample diluted in blocking buffer was added and allowed to incubate for 2 h at room temperature with mixing at 300 r.p.m. The plate was washed four times with 200 µl of TBS-T, and 50 µl of secondary antibody (Abcam goat polyclonal biotin-conjugate apoC-III, ab21024) diluted in blocking buffer was added and allowed to incubate for 1 h at room temperature with rotation at 300 r.p.m. The plate was washed once with TBS-T, and 50 µl of SA-HRP (Abcam, 34028) diluted 100-fold in PBS was added and allowed to incubate for 30 min at room temperature with mixing at 300 r.p.m. The plate was washed four times with 200 µl of TBS-T and developed with 80 µl of TMB (Thermo Ultra-TMB ELISA, 34028) followed by 50 µl of 0.5 N HCl. Plates were read at 450 nm. The amount of apoC-III in test wells was calculated from a four-parameter fit of a standard curve (Molecular Devices) constructed using purified apoC-III (Athens Research and Technology).
For in vivo studies of anti-apoC-III monoclonal antibodies, apoB levels were determined with an ELISA assay. A 96-well plate (Greiner, 655061) was coated overnight at 4 °C with 50 µl of primary antibody to apoB (Meridian Life Sciences goat polyclonal anti–human ApoB, K45253G) diluted in PBS. The plate was washed four times with 200 µl of TBS-T and blocked with 200 µl of blocking buffer (Pierce Clear Milk Blocker, 37587; in PBS) for 90 min at 30 °C. The block-ing buffer was removed, and 50 µl of test sample diluted in blocking buffer was added and allowed to incubate for 2 h at room temperature with rotation at 300 r.p.m. The plate was washed four times with 200 µl of TBS-T, and 50 µl of secondary antibody (Meridian Life Sciences goat polyclonal biotin-conjugate ApoB48/100, 34003G) diluted in blocking buffer was added and allowed to incubate for 1 h at room temperature with rotation at 300 r.p.m. The plate was washed once with TBS-T, and 50 µl of SA-HRP (Abcam, 64269) diluted 100-fold in PBS was added and allowed to incubate for 30 min at room temperature with rotation at 300 r.p.m. The plate was washed four times with 200 µl of TBS-T and developed with 80 µl of TMB (Thermo Ultra-TMB ELISA, 34028) followed by 50 µl of 0.5 N HCl. Absorbance was read at 450 nm. The amount of apoB in test wells was calculated from a four-parameter fit of a standard curve (Molecular Devices) constructed using mouse VLDL isolated by centrifugation (apoB con-tent is assumed to be 20% of the total protein content).
The plasma levels of the STT505 and STT5058 antibodies were determined with an ELISA assay. A 96-well plate (Greiner, 655061) was coated overnight at 4 °C with 50 µl of primary IgG antibody (Fitzgerald 41-XG57 goat anti–human IgG Fc polyclonal) diluted in PBS. The plate was washed four times with 200 µl of TBS-T and blocked with 200 µl of blocking buffer consisting of 3% BSA (Roche BSA Fraction V Protease Free, 03 117 332 001) plus clear milk (Pierce Clear Milk Blocker, 37587) in PBS for 90 min at 30 °C. The blocking buffer was removed, and 50 µl of test sample diluted in blocking buffer was added and allowed to incubate for 2 h at room temperature with rotation at 300 r.p.m. The plate was washed four times with 200 µl of TBS-T, and 50 µl of secondary anti-body (Abcam, goat anti-human IgG Fc (biotin) polyclonal, ab97223) diluted in blocking buffer was added and allowed to incubate for 1 h at room temperature with rotation at 300 r.p.m. The plate was washed once with TBS-T, and 50 µl of SA-HRP (Abcam, 64269) diluted 100-fold in PBS was added and allowed to

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
nature medicine doi:10.1038/nm.4390
incubate for 30 min at room temperature with rotation at 300 r.p.m. The plate was washed four times with 200 µl of TBS-T and developed with 80 µl of TMB (Thermo Ultra-TMB ELISA, 34028) followed by 50 µl of 0.5 N HCl. The absorb-ance was read at 450 nm. The amount of IgG in test wells was calculated from a four-parameter fit of a standard curve (Molecular Devices) constructed using the purified test antibody.
ApoC-III lipoprotein-binding measurements. Human TRLs and HDL3 were isolated from pooled human plasma by density-gradient ultracentrifugation. Isolated lipoproteins were dialyzed against PBS, and protein concentration was measured by BCA assay. Radioiodinated WT or A43T apoC-III, generated as described above, was solubilized in 10 mM ammonium bicarbonate buffer and dialyzed against PBS. For binding studies of apoC-III with plasma lipoproteins, 1 µg of [125I]apoC-III was combined with 200 µl of pooled donor plasma and incubated at 37 °C for 1, 5, or 24 h with gentle shaking. Immediately after incubation, mixtures were fractionated on a Superose 6 gel-filtration column. TG and cholesterol concentrations were measured in 0.1 ml of each FPLC fraction by colorimetric assay in a 96-well microplate, and [125I]apoC-III abun-dance was measured by gamma counting. The activity in each FPLC fraction was expressed as the percentage of the total activity (the sum of the activity in all the fractions). The relative activity in lipoprotein fractions versus free protein fractions was assessed by comparing the sum of the total area for the lipoprotein fractions versus the free protein fractions. Results were compared across replicate experiments. For binding of apoC-III to TRLs or HDL3, the procedure above was followed with the exception that 200 µg of TRL protein or 100 µg of HDL3 protein was used for each experiment in combination with 1 µg of [125I]apoC-III.
Binding of [125I]apoC-III to apolipoprotein-free lipid emulsions was also measured. Large lipid emulsions were prepared as described previously52. Briefly, a mixture of 300 mg of triolein (Sigma-Aldrich) and 18 mg of egg-phosphatidylcholine (Sigma-Aldrich) was dried under a nitrogen evaporator and then sonicated after addition of 5 ml of glycerol (Thermo Fisher Scientific) using a Branson 450 microtip sonicator for 5 min. Concentrated emulsions were allowed to clear overnight. 15 µl of this concentrated emulsion was combined with 90 µl of distilled water, 15 µl of 1.0 M Tris pH 8.0, 15 µl of 15% BSA solu-tion, and 15 µl of 3.0 M NaCl to give a final volume of 150 µl of working emul-sion. 1 µg of [125I]apoC-III was combined with 150 µl of the working emulsion and incubated at 37 °C for 1 h, and mixtures were then fractionated by FPLC and analyzed for 125I activity in the emulsion-containing fractions versus the unbound protein fractions as detailed above. Preliminary FPLC fractionation experiments showed that emulsions alone eluted in fractions 5–15 of the 79 fractions obtained from gel-filtration analysis, which mirrored the fractiona-tion pattern of human TRLs on the same Superose 6 column under the same conditions. For binding of apoC-III to TRLs in the presence of STT5058 anti-body versus isotype control antibody, the procedure above was followed with the exception that 200 µg of human TRL protein was first incubated with 1 µg of [125I]TC-modified apoC-III for 1 h at 37 °C and either of the two antibodies was then added to the reaction mixture. 0.2 mg of antibody was added to the reactions to reflect the approximate concentration of antibody present in the in vivo experiment (0.54 mg of antibody per mouse). Next, the reactions were incubated for another 8 h before fractionation by FPLC and measurement of [125I]TC-modified apoC-III activity in each fraction, normalized to the total activity in the reaction mixtures as described above.
Surface plasmon resonance assay. WT or A43T human apoC-III (Thermo Fisher Scientific) were captured on a chip coated with dimyristoyl phosphati-dylcholine (DMPC) (Avanti Polar Lipids), and the binding kinetics of these apoC-III forms to the coated chip were measured at pH 7.4 and pH 5.5. 60 µl of each of these proteins was diluted in HBS-EP buffer (GE, BR-1008-26; 0.010 M HEPES, 0.150 M NaCl, 3 mM EDTA, 0.05% (v/v) surfactant P20, pH 7.4) and was injected at a concentration of 1–100 nM. The proteins were passed through the flow cells at a flow rate of 30 µl/min, followed by an off-rate wash at pH 7.4 or pH 5.5 for 5 min. The resulting sensorgrams were analyzed using BIAevaluation 4.1 software applying the Langmuir 1:1 binding model to derive binding kinetics. Data were zero adjusted, and the reference cell sensorgrams were subtracted and used to calculate the on rate and off rate of apoC-III chip
association over a range of apoC-III concentrations of 10, 25, and 50 µg/ml. The ratio of the off rate to the on rate at these concentrations was used as the dissociation constant (Kd) for WT versus mutant apoC-III.
LPL activity assays. In vitro LPL activity in the presence of WT versus A43T variant apoC-III was measured by two independent methods. First, activity was measured against a large lipid emulsion made as described above but with the addition of [3H]triolein (PerkinElmer; 99 µg per 300 mg of nonra-dioactive triolein) before glycerol addition and sonication as described previ-ously52. Working emulsions were made by the method above except that 75 µl of water was used and 15 µl of heat-inactivated human serum was added to provide a source of apoC-II. The working emulsion (150 µl) was combined with conditioned medium (100 µl) from COS-7 cells expressing human LPL through adenoviral transduction, as described previously52. 50 µl of WT or A43T apoC-III (final reaction concentration of 20 µM for each) or culture medium without apoC-III was added to give a final reaction volume of 300 µl. Reactions were incubated at 37 °C for 30 min with gentle shaking before addition of 3.25 ml of methanol:chloroform:heptane solvent (1.41:1.25:1.00) to stop the reactions and subsequent addition of 1.05 ml of pH 10.0 Buffer Potassium Carbonate, Potassium Tetraborate, Potassium Hydroxide, Disodium EDTA Dihydrate (Thermo Fisher Scientific). Stopped reactions were then centrifuged at 688g for 20 min to complete the extraction, and the upper phase containing liberated fatty acids was used for scintillation counting (0.5 ml per sample). The relative amount of hydrolysis of [3H]triolein to [3H]oleic acid was calculated for each sample and compared across groups. A second LPL activity assay was performed using an Intralipid TG substrate in the presence or absence of WT versus A43T apoC-III, as described previously53. Briefly, 10% Intralipid substrate (Santa Cruz Biotechnology; 1.2 nM per reaction) and WT or A43T apoC-III (range of 0–58 µM per reaction) were combined in 96-well microplates to a volume of 40 µl and incubated at 30 °C for 15 min, followed by the addition of purified LPL (Sigma-Aldrich; 40 nM per reaction) to give a final reaction volume of 60 µl. Reactions were incubated at 30 °C for 30 min, and 100 µl of Wako NEFA-HR(2) R1 reagent (Wako Chemicals) was added along with Orlistat (Santa Cruz Biotechnology; 20 µM final reaction concentration) to inhibit further lipolysis and the reactions were incubated for another 15 min at 30 °C. Then, 50 µl of Wako NEFA-HR(2) R2 reagent was added, and the reactions were incubated for another 5 min at 30 °C and the absorbance was measured at 550 nm for each sample. All LPL assays were performed using three independent replicate experiments.
ApoC-III-depleted VLDL and DiI labeling. To float chylomicrons, pooled human plasma (Innovative Research) was centrifuged at 100,000g for 10 min at 4 °C. Chylomicrons were removed by pipetting, and the resulting plasma was gently mixed with goat anti-human apoC-III Sepharose 4b (Academy Biomedical) overnight at 4 °C. The unbound fraction was collected, mixed with OptiPrep (12% iodixanol final concentration), and centrifuged for 3.5 h at 350,000g at 4 °C. VLDL particles were removed from the top of the gradi-ent. For DiIC12(3) labeling, a 6 mM stock of DiIC12(3) (Molecular Probes) in DMSO was slowly added to a 1.5 µg/µl sample of VLDL to give a final concen-tration of 100 µM DiIC12(3). The sample was kept in the dark for 7 h at 37 °C, and unincorporated DiIC12(3) was removed through an 8-µm PSE filter.
DiI-VLDL uptake assays. HepG2 cells (ATCC, HB-8065) were seeded on poly(d-lysine)-coated 96-well tissue culture plates (Greiner Bio-One, 655940) in 100 µl of complete MEM (Life Technologies) plus with 10% FCS (Seradigm) and grown at 37 °C in 5% CO2. After 24 h, the medium was removed and the cell monolayer was washed once with 200 µl of complete MEM plus with 0.0125% BSA (Roche) and replaced with 100 µl of complete MEM plus with 0.0125% BSA. After 24 h at 37 °C and 5% CO2, the medium was removed and replaced with 50 µl of a test component mixture assembled as follows: apoC-III (Athens Research and Technology) and test antibody were mixed in MEM plus 0.0125% BSA and incubated for 15 min at 37 °C, followed by 30 µg/µl apoC-III-depleted DiI-VLDL (see below) for 20 min at 37 °C. After 3.5 h of incuba-tion at 37 °C and 5% CO2, the test mixture was removed and the cells were incubated with 100 µl of 1% intralipid diluted in complete MEM for 20 min at 37 °C and 5% CO2. The medium was removed, and cells were washed three

© 2
017
Nat
ure
Am
eric
a, In
c., p
art
of
Sp
rin
ger
Nat
ure
. All
rig
hts
res
erve
d.
nature medicinedoi:10.1038/nm.4390
times with 200 µl of 37 °C DPBS (Life Technologies, 14190-144). After wash-ing, 100 µl of isopropanol was added to each well and the plate was incubated at room temperature for 15 min with gentle shaking. A 75-µl aliquot was transferred to a black 96-well plate (VWR, 89089-582), and fluorescence was measured (excitation 520 nm, emission 580 nm). The amount of DiI-VLDL extracted from cells per unit volume of isopropanol was determined from a DiI-VLDL standard curve. The remaining isopropanol was removed from the cell plate, 100 µl of lysis buffer (0.1 N NaOH, 0.1% SDS) was added to each well, and the plate was incubated for a minimum of 30 min. A 25-µl aliquot was used to measure protein concentration (Pierce BCA Protein Assay; Thermo Scientific, 23225), and the quantity of VLDL taken into the cell was calculated from the DiI-VLDL/protein ratio. Data were graphed using GraphPad Prism 6 and are reported as average ± s.e.m. One-way ANOVA with multiple com-parisons was calculated using GraphPad Prism 6.
Statistical analysis. Data from human samples shown are represented as mean ± s.d., where error bars show s.d. All other data are shown as mean ± s.e.m., where error bars show s.e.m. Statistical comparisons between two groups were performed using a two-tailed Student’s t test or one-way ANOVA as appropri-ate and when assumptions of distribution of the data were valid. Comparisons between three experimental groups were performed using two-way ANOVA. For all measurements, variance was estimated to be similar between groups compared. For measurement of mutant:WT apoC-III ratios in A43T hetero-zygous carriers, the mean of the ratios among carriers was compared to an expected theoretical mean of 1.0 using a one-sample t test. Statistical signifi-cance was defined as P < 0.05 for all analyses.
Life sciences reporting summary. Additional information regarding human participants and animals used, reagent validation and availability, statistics, software for data analysis, and reproducibility of results can be found in the Life Sciences Reporting Summary accompanying this manuscript.
Data availability. The authors declare that all data relevant to supporting the conclusions of this study are present within the manuscript and its sup-plementary information. Exome Chip SNP genotyping data from human par-
ticipants from the Penn Medicine BioBank and HHDL cohorts were deposited in the Gene Expression Omnibus (GEO) database (accession GSE100271). Additional requests for correspondence regarding data in this study may be made to the corresponding author (D.J.R.). Source data files for Figures 1–4 are available online.
41. Peloso, G.M. et al. Association of low-frequency and rare coding-sequence variants with blood lipids and coronary heart disease in 56,000 whites and blacks. Am. J. Hum. Genet. 94, 223–232 (2014).
42. MacLean, B. et al. Skyline: an open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 26, 966–968 (2010).
43. Mucaki, E.J., Shirley, B.C. & Rogan, P.K. Prediction of mutant mRNA splice isoforms by information theory–based exon definition. Hum. Mutat. 34, 557–565 (2013).
44. Ibrahim, S., Somanathan, S., Billheimer, J., Wilson, J.M. & Rader, D.J. Stable liver-specific expression of human IDOL in humanized mice raises plasma cholesterol. Cardiovasc. Res. 110, 23–29 (2016).
45. Somanathan, S. et al. AAV vectors expressing LDLR gain-of-function variants demonstrate increased efficacy in mouse models of familial hypercholesterolemia. Circ. Res. 115, 591–599 (2014).
46. Chen, S.J. et al. Biodistribution of AAV8 vectors expressing human low-density lipoprotein receptor in a mouse model of homozygous familial hypercholesterolemia. Hum. Gene Ther. Clin. Dev. 24, 154–160 (2013).
47. Kassim, S.H. et al. Adeno-associated virus serotype 8 gene therapy leads to significant lowering of plasma cholesterol levels in humanized mouse models of homozygous and heterozygous familial hypercholesterolemia. Hum. Gene Ther. 24, 19–26 (2013).
48. Rader, D.J., Castro, G., Zech, L.A., Fruchart, J.C. & Brewer, H.B. Jr. In vivo metabolism of apolipoprotein A-I on high density lipoprotein particles LpA-I and LpA-I,A-II. J. Lipid Res. 32, 1849–1859 (1991).
49. Cain, W.J. et al. Lipoprotein [a] is cleared from the plasma primarily by the liver in a process mediated by apolipoprotein [a]. J. Lipid Res. 46, 2681–2691 (2005).
50. Tanigawa, H. et al. Expression of cholesteryl ester transfer protein in mice promotes macrophage reverse cholesterol transport. Circulation 116, 1267–1273 (2007).
51. Ota, T., Gayet, C. & Ginsberg, H.N. Inhibition of apolipoprotein B100 secretion by lipid-induced hepatic endoplasmic reticulum stress in rodents. J. Clin. Invest. 118, 316–332 (2008).
52. McCoy, M.G. et al. Characterization of the lipolytic activity of endothelial lipase. J. Lipid Res. 43, 921–929 (2002).
53. Larsson, M., Vorrsjö, E., Talmud, P., Lookene, A. & Olivecrona, G. Apolipoproteins C-I and C-III inhibit lipoprotein lipase activity by displacement of the enzyme from lipid droplets. J. Biol. Chem. 288, 33997–34008 (2013).
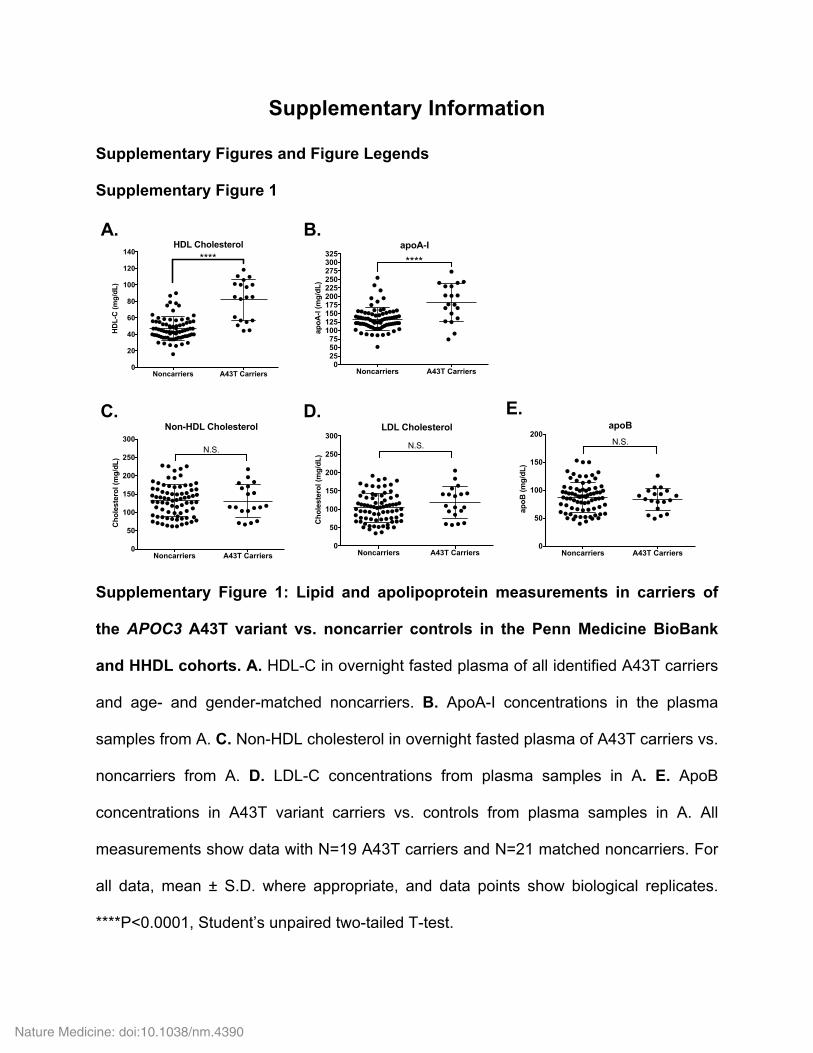
Supplementary Information
Supplementary Figures and Figure Legends
Supplementary Figure 1
Supplementary Figure 1: Lipid and apolipoprotein measurements in carriers of
the APOC3 A43T variant vs. noncarrier controls in the Penn Medicine BioBank
and HHDL cohorts. A. HDL-C in overnight fasted plasma of all identified A43T carriers
and age- and gender-matched noncarriers. B. ApoA-I concentrations in the plasma
samples from A. C. Non-HDL cholesterol in overnight fasted plasma of A43T carriers vs.
noncarriers from A. D. LDL-C concentrations from plasma samples in A. E. ApoB
concentrations in A43T variant carriers vs. controls from plasma samples in A. All
measurements show data with N=19 A43T carriers and N=21 matched noncarriers. For
all data, mean ± S.D. where appropriate, and data points show biological replicates.
****P<0.0001, Student’s unpaired two-tailed T-test.
E.
A.
Figure S1
Noncarriers A43T Carriers0
50
100
150
200
250
300
Cho
lest
erol
(mg/
dL)
Non-HDL Cholesterol
N.S.
Noncarriers A43T Carriers0
50
100
150
200
250
300
Cho
lest
erol
(mg/
dL)
LDL Cholesterol
N.S.
Noncarriers A43T Carriers0
50
100
150
200
apoB
(mg/
dL)
apoB
N.S.
B.
C. D.
Noncarriers A43T Carriers0
20
40
60
80
100
120
140
HD
L-C
(mg/
dL)
HDL Cholesterol****
Noncarriers A43T Carriers0
255075
100125150175200225250275300325
apoA
-I (m
g/dL
)
apoA-I
****
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 2
Supplementary Figure 2: Schematic illustration of the targeted mass
spectrometry assay workflow. Approach for development and implementation of
human-specific WT vs. A43T apoC-III isoform targeted multiple reaction monitoring
assays is shown.
Sample preparation • AspN digestion of plasma proteins • H2O2 treatment to oxidize
methionine residues in AspN peptides
Data Acquisition • Selection of peptides and transitions • Liquid Chromatography • Targeted mass spectrometry
Data Analysis • Fit standard curve using mouse
samples • Calculate concentration for human
samples
Heavy isotope-labeled peptides
ApoC-III Concentration
Figure S2
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 3
Supplementary Figure 3: Example LC-MS/MS data showing the extracted ion
chromatograms for the native peptide (blue) and isotopically labeled peptide
standards (red). The peptide sequence and product ion spectra for the AspN-
generated 20 amino acid peptide corresponding to WT (A) and mutant apoC-III (B) are
included above each chromatogram. The position of the variant amino acid between
WT and mutant peptides is highlighted in red.
13.5 14.0 14.5 15.0 15.5 16.0 16.5 Time (min)
0 20 40 60 80 100 120 140 160
400 600 800 1000 m/z
0 20 40 60 80
100
Rel
ativ
e A
bund
ance
y18
y16 y14
y15
WT ApoC-III: Ala43 peptide
Inte
nsity
(103
)
13.5 14.0 14.5 15.0 15.5 16.0 16.5 Time (min)
0 20 40 60 80 100 120
400 600 800 1000 m/z
0 20 40 60 80
100
Rel
ativ
e A
bund
ance
Mutant ApoC-III: Thr43 peptide y18
y16 y14
y15
Inte
nsity
(103
)
A.
B.
Figure S3
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 4
Supplementary Figure 4: external LC-MS/MS calibration curve. Five mouse plasma
samples from mice expressing either human WT or A43T APOC3 via AAV expression
with known WT and A43T mutant apoC-III concentration were used as external
calibrators to determine WT (A) and A43T mutant (B) apoC-III concentration in human
samples. These samples were processed and analyzed in parallel with other human
plasma samples. Linear regression was used to fit standard curves.
0 5 10 150.0
0.5
1.0
1.5
WT apoC-III Concentration (mg/dL)
Ligh
t/Hea
vy
0.0 0.5 1.0 1.50.00
0.05
0.10
0.15
0.20
0.25
Mutant apoCIII Concentration (mg/dL)
Ligh
t/Hea
vy
A. B.
Figure S4
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 5
Supplementary Figure 5: Dose-optimization studies of WT APOC3 AAV in mice. A.
Plasma human ApoC-III concentration in C57BL/6 WT mice administered the indicated
doses (genome copies of vector per mouse) of WT APOC3 AAV by intraperitoneal
injection (N=3 mice per group). Measurements were made eight weeks after AAV
administration. B. Plasma TGs from four hour fasted plasma from mice in A over eight
weeks. C. Plasma HDL-C from mice in A. D. Plasma Non-HDL-C in mice from A. E.
Plasma phospholipids from mice in A. F. Plasma alanine aminotransferase (ALT) levels
in mice from A. For all panels, N=3 mice per group. All data was replicated once by an
independent repeat experiment. All measurements show mean ± S.E.M. where
appropriate and data show biological replicates. *P<0.05, ***P<0.001, Student’s
unpaired two-tailed T-test compared to Null group.
0 20 40 600
30
60
90
120
150
Time (days)
Trig
lyce
rides
(mg/
dL)
Triglycerides
Null3E101E113E11
0 20 40 600
50
100
150
200
250
Time (days)
Phos
phol
ipid
s (m
g/dL
)Phospholipids
Null3E101E113E11
0 20 40 600
20
40
60
80
100
Time (days)
Cho
lest
erol
(mg/
dL)
HDL Cholesterol
Null3E101E113E11
0 20 40 600
10
20
30
40
50
Time (days)
Cho
lest
erol
(mg/
dL)
Non-HDL Cholesterol
Null3E101E113E11
0 20 40 600
50
100
150
200
250
300
Time (days)
IU/L
ALT
Null3E101E113E11
Null 3E10 1E11 3E110
5
10
15apoC-III Concentration
AAV Dose
apoC
-III (
mg/
dL)
***
*
A. B. C.
D. E. F.
Figure S5
Nature Medicine: doi:10.1038/nm.4390
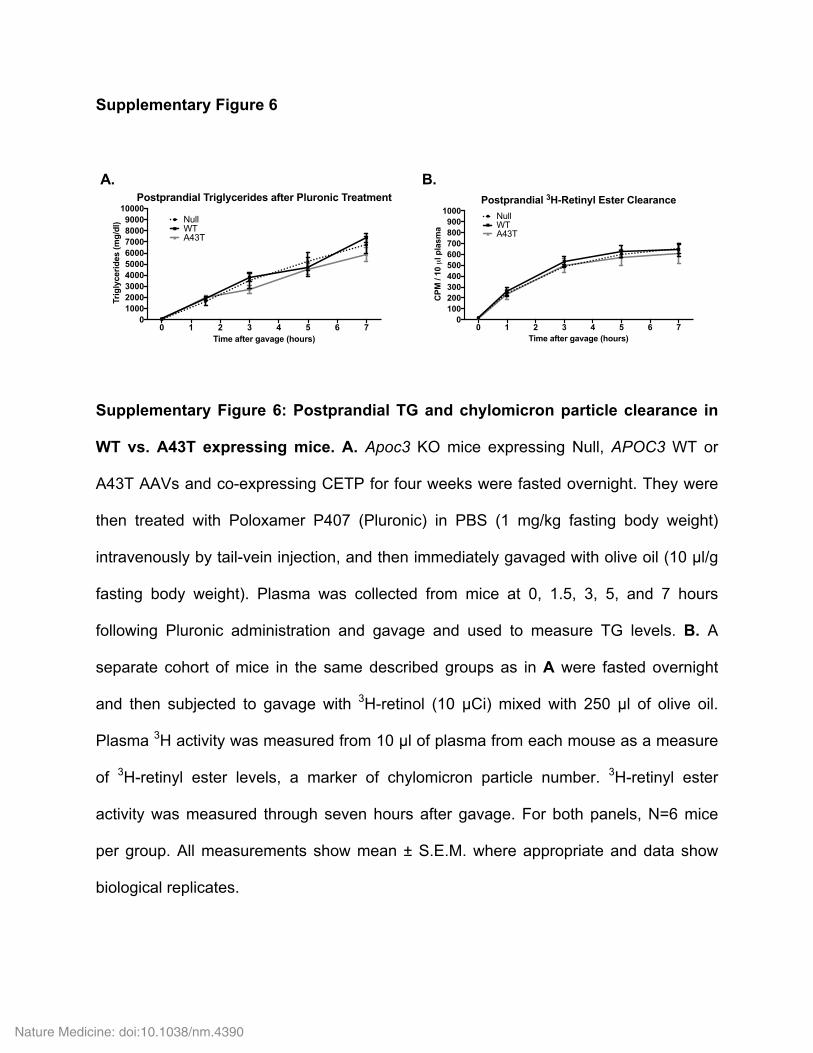
Supplementary Figure 6
Supplementary Figure 6: Postprandial TG and chylomicron particle clearance in
WT vs. A43T expressing mice. A. Apoc3 KO mice expressing Null, APOC3 WT or
A43T AAVs and co-expressing CETP for four weeks were fasted overnight. They were
then treated with Poloxamer P407 (Pluronic) in PBS (1 mg/kg fasting body weight)
intravenously by tail-vein injection, and then immediately gavaged with olive oil (10 µl/g
fasting body weight). Plasma was collected from mice at 0, 1.5, 3, 5, and 7 hours
following Pluronic administration and gavage and used to measure TG levels. B. A
separate cohort of mice in the same described groups as in A were fasted overnight
and then subjected to gavage with 3H-retinol (10 µCi) mixed with 250 µl of olive oil.
Plasma 3H activity was measured from 10 µl of plasma from each mouse as a measure
of 3H-retinyl ester levels, a marker of chylomicron particle number. 3H-retinyl ester
activity was measured through seven hours after gavage. For both panels, N=6 mice
per group. All measurements show mean ± S.E.M. where appropriate and data show
biological replicates.
0 1 2 3 4 5 6 70
100200300400500600700800900
1000
Time after gavage (hours)
CPM
/ 10
µl p
lasm
a
Postprandial 3H-Retinyl Ester ClearanceNullWTA43T
0 1 2 3 4 5 6 70
100020003000400050006000700080009000
10000
Time after gavage (hours)
Trig
lyce
rides
(mg/
dl)
Postprandial Triglycerides after Pluronic Treatment
NullWTA43T
Figure S6A. B.
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 7
Supplementary Figure 7: VLDL TG and apoB secretion in mice expressing WT vs.
A43T APOC3. A. (top) Plasma TG levels over time after Poloxamer P407 and 35S
administration in WT mice expressing Null or APOC3 AAVs for four weeks. Curves were
fitted to a line in GraphPad Prism. (bottom) Secretion rates as measure by the slope of
the curves at the top. B. (top) Plasma 35S-apoB-100 activity (normalized to activity in 2
µl of plasma after TCA precipitation) for mice expressing the indicated transgenes four
weeks after AAV administration. ApoB-100 bands were excised from gels after SDS-
PAGE and activity measured by scintillation counting and normalized to the TCA
precipitable activity from plasma from the same plasma samples. (bottom) Secretion
rates as measure by the slope of the curves at the top. C. Plasma 35S-apoB-48 activity
from mice in A and B. ApoB-48 bands were excised after SDS-PAGE, measured for
activity and normalized to TCA precipitable activity from the same plasma samples.
(bottom) Secretion rates as measured by the slope of the curves at the top. For all data,
0 30 60 90 1200
200
400
600
800
1000
1200
Time (min)
Trig
lyce
rides
(mg/
dL)
VLDL-TG Secretion
NullWTA43T
Null WT A43T6
7
8
9
10
11
TG s
ecre
tion
rate
[m
g/(d
L x
hr)]
VLDL-TG Secretion Rate
0 30 60 90 1200.0
0.5
1.0
1.5
2.0
2.5
Time (min)
35S-
Apo
B10
0 C
PM /
plas
ma
TCA
-pre
cipi
tabl
e C
PM
Hepatic apoB-100 Secretion
NullWTA43T
Null WT A43T0.010
0.012
0.014
0.016
0.018
0.020N
orm
aliz
ed 35
S-ap
oB-1
00 C
PM/m
in
VLDL-apoB-100 Secretion Rate
0 30 60 90 1200.0
0.5
1.0
1.5
2.0
2.5
Time (min)
35S-
Apo
B48
CPM
/ pl
asm
a TC
A-p
reci
pita
ble
CPM
Hepatic apoB-48 Secretion
NullWTA43T
Null WT A43T0.008
0.010
0.012
0.014
0.016
0.018
Nor
mal
ized
35S-
apoB
-48
CPM
/min
VLDL-apoB-48 Secretion Rate
A. B. C.Figure S7
Nature Medicine: doi:10.1038/nm.4390

N=6 mice per group, and data was replicated twice by two repeat experiments in
independent cohorts of mice. For data in bottom sections of panels A-C, box length
spans 25th to 75th percentile range of the data points, with the middle line indicating the
median, and whiskers indicating the minimum and maximum values for the given data
set. Data in top sections of panels A-C show mean ± S.E.M. where appropriate and
data shows biological replicates.
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 8
Supplementary Figure 8: Original images from immunoblots shown in Figure 2.
Uncropped Hyblot CL films of the images in Figure 2M are shown. A. Uncropped film for
anti-apoC-III immunoblot in Figure 2M. B. Uncropped film for anti-actin immunoblot in
Figure 2M.
MW ladder
Null AAV mice (N=3)
WT APOC3 AAV mice (N=3)
A43T APOC3 AAV mice (N=3)
Human plasma apoC-III
standard
~8.8 kDa (2 glycosylated
isoforms of apoC-III)
20 kDa
30 kDa
40 kDa
50 kDa
60 kDa80 kDa
Molecular Weights of Ladder Standard
Nonspecific Bands
Sample Loaded:
20 kDa
30 kDa
40 kDa
50 kDa
60 kDa80 kDa
Molecular Weights of Ladder Standard
~8.8 kDa (2 glycosylated
isoforms of apoC-III)
Sample Loaded:
MW ladder
Null AAV mice (N=3)
WT APOC3 AAV mice (N=3)
A43T APOC3 AAV mice (N=3)
Human plasma apoC-III
standard
Mouse β-actin
Nonspecific Bands
Residual signal from α-apoC-III immunoblot of
same membrane
α-humanapoC-III
α-mouse β-actin
A.
B.
Figure S8
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 9
Supplementary Figure 9: Urinary clearance of 125I-labeled WT vs. A43T apoC-III in
mice. A. Plasma 125I clearance curves over 24 hours for radiolabeled WT vs. A43T
apoC-III in WT mice expressing either WT or A43T APOC3 AAVs (N=4 mice per group).
Mice expressing WT APOC3 AAV received WT iodinated ApoC-III by intravenous
administration and A43T APOC3-expressing mice received iodinated A43T apoC-III
protein. B. Fractional catabolic rate of plasma 125I activity, as measured by WinSAAM
multi-exponential modeling program. Catabolic rates were calculated for each mouse
individually. C. Total 125I activity measured in urine collected from WT vs. A43T
expressing mice at the end of 24 hours of collection following intravenous 125I ApoC-III
administration. D. Total urinary 125I activity at the indicated time intervals over the
0 6 12 18 240.0
0.1
0.2
0.3
0.4
0.5
0.6
Time (min)
125 I
Act
ivity
in to
tal v
olum
e co
llect
ed /
Initi
al a
ctiv
ity a
dmin
iste
red
Total Urinary 125I Activity
WT*
A43T
0 6 12 18 240.0
0.1
0.2
0.3
0.4
0.5
Time (hours)
125 I
Act
ivity
in to
tal v
olum
e co
llect
ed /
Initi
al a
ctiv
ity a
dmin
iste
red
TCA Supernatant Urinary 125I Activity
WTA43T
*
0 6 12 18 240.00
0.02
0.04
0.06
0.08
0.10
0.12
Time (hours)
125 I
Act
ivity
in to
tal v
olum
e co
llect
ed /
Initi
al a
ctiv
ity a
dmin
iste
red
TCA Precipitable Urinary 125I Activity
WTA43T
*
0 250 500 750 1000 12500.01
0.1
1
Time (min)
Frac
tion
of a
dmin
iste
red
activ
ity
Plasma 125I apoC-III Clearance
WTA43T
**
WT A43T0.004
0.005
0.006
0.007
0.008
Frac
tiona
l Cat
bolic
Rat
e (p
ools
/min
.)
125I Plasma FCR
*
1 min
180 m
in
360 m
in
1440
min
1 min
180 m
in
360 m
in
1440
min
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Frac
tion
of to
tal 12
5 I ac
tivity
Distribution of Plasma 125I Activity after TCA Precipitation
TCA SupernatantTCA Pellet
WT A43T
**** *
0-6 hrs
6-12 h
rs
12-24
hrs
0-6 hrs
6-12 h
rs
12-24
hrs0.0
0.2
0.4
0.6
0.8
1.0
1.2Fr
actio
n of
tota
l 125 I
activ
ity
Distribution of Urine 125I Activity after TCA Precipitation
TCA SupernatantTCA Pellet
WT A43T
A. B.
D. E. F.
G. H.
Figure S9
WT A43T0.0
0.2
0.4
0.6
0.8
Frac
tion
of a
dmin
iste
red
activ
ity
Total Urinary 125I Activity *
C.
Nature Medicine: doi:10.1038/nm.4390

course of the 125I apoC-III clearance study. Urine was collected individually at the
indicated timepoints for each mouse and 125I activity was measured and removed from
metabolic cages prior to the next collection period up through 24 hours. E. 125I activity in
TCA soluble fraction of urine collected from each mouse at the indicated time intervals.
20 µl of urine was subjected to TCA precipitation and TCA soluble fractions were
collected and re-measured for 125I activity. The fraction of the 20 µl of urinary 125I activity
appearing in the TCA soluble fraction was used to calculate the proportion of TCA
soluble 125I activity in the total urine collected for each mouse for the indicated time
intervals. F. 125I activity in the TCA precipitable fraction of the urine collected from each
mouse at the indicated time intervals from samples used in D. G. Fraction of plasma 125I
activity in TCA supernatant vs. pellet for each timepoint from 20 µl of plasma from mice
administered 125I-labeled WT vs. A43T apoC-III. Total plasma 125I activity was measured
for each mouse and subsequently TCA precipitation and measurement of activity in
soluble vs. precipitable fractions was performed. H. Fraction of urinary 125I activity in
TCA supernatant vs. pellet for each time interval of urine collection for mice
administered 125I labeled WT vs. A43T apoC-III. TCA soluble vs. precipitable 125I activity
was measured from 20 µl of urine for each mouse for each time interval as indicated in
D-E. For all panels, N=4 mice per group. For data in panels B-C, box length spans 25th
to 75th percentile range of the data points, with the middle line indicating the median,
and whiskers indicating the minimum and maximum values for the given data set. All
other measurements show mean ± S.E.M. where appropriate, and all data shows
biological replicates. For A, **P<0.01, two-way ANOVA of WT vs. A43T clearance
curves. For B-F, *P<0.05, ***P<0.001, Student’s unpaired T-test. For F, comparisons
Nature Medicine: doi:10.1038/nm.4390

are of the fraction of total activity in TCA precipitable fraction for WT vs. A43T for a
given time interval.
Nature Medicine: doi:10.1038/nm.4390

Supplementary Figure 10
Supplementary Figure 10: Helical wheel diagrams of the 18-residue human apoC-III
peptide 34-51 of the full-length apoC-III sequence (peptide 14-31 of the mature form
which lacks the first 20 residues, which are the signal peptide). A. Helical wheel for the wild
type peptide which contains A43 (A23 of the mature form) - the N-terminal residue G and C-
terminal residue Q are marked. The vector of the helical hydrophobic moment is perpendicular
(black arrow) with the hydrophobic face of the amphipathic α-helix, which contains the 7 amino
acids LMAVAAY oriented towards the bottom of the picture (A43 is located between residues M
and V). B. equivalent helical wheel for the A43T variant showing that the hydrophobic face is
now LMTVAAY. The helical wheels were constructed using the program Heliquest (www.
heliquest.ipmc.cnrs.fr).
Figure S10A. B.
Nature Medicine: doi:10.1038/nm.4390
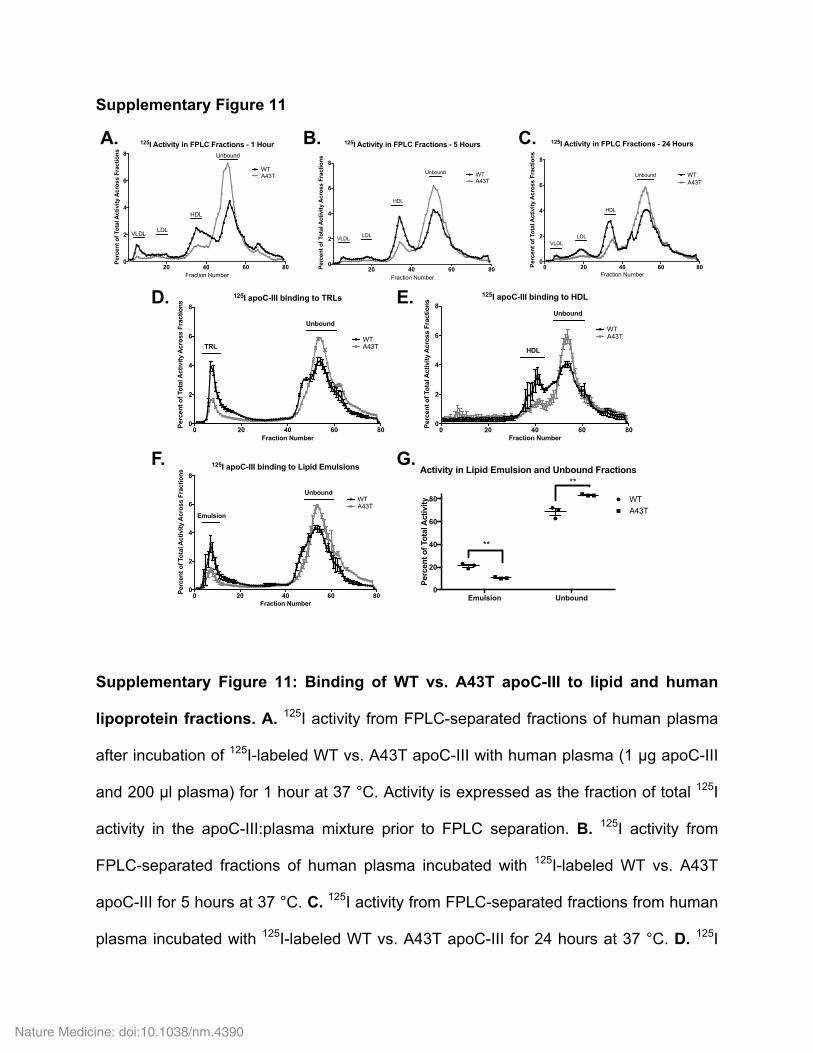
Supplementary Figure 11
Supplementary Figure 11: Binding of WT vs. A43T apoC-III to lipid and human
lipoprotein fractions. A. 125I activity from FPLC-separated fractions of human plasma
after incubation of 125I-labeled WT vs. A43T apoC-III with human plasma (1 µg apoC-III
and 200 µl plasma) for 1 hour at 37 °C. Activity is expressed as the fraction of total 125I
activity in the apoC-III:plasma mixture prior to FPLC separation. B. 125I activity from
FPLC-separated fractions of human plasma incubated with 125I-labeled WT vs. A43T
apoC-III for 5 hours at 37 °C. C. 125I activity from FPLC-separated fractions from human
plasma incubated with 125I-labeled WT vs. A43T apoC-III for 24 hours at 37 °C. D. 125I
0 20 40 60 800
2
4
6
8
Fraction Number
Perc
ent o
f Tot
al A
ctiv
ity A
cros
s Fr
actio
ns
125I apoC-III binding to TRLs
WTA43TTRL
Unbound
20 40 60 800
2
4
6
8
125I Activity in FPLC Fractions - 1 Hour
Fraction Number
Perc
ent o
f Tot
al A
ctiv
ity A
cros
s Fr
actio
ns
WTA43T
VLDL LDL
HDL
Unbound
0 20 40 60 800
2
4
6
8
Fraction Number
Perc
ent o
f Tot
al A
ctiv
ity A
cros
s Fr
actio
ns
125I apoC-III binding to Lipid Emulsions
WTA43T
Emulsion
Unbound
20 40 60 800
2
4
6
8
125I Activity in FPLC Fractions - 5 Hours
Fraction Number
Perc
ent o
f Tot
al A
ctiv
ity A
cros
s Fr
actio
ns
WTA43T
HDL
LDLVLDL
Unbound
0 20 40 60 800
2
4
6
8
Fraction Number
Perc
ent o
f Tot
al A
ctiv
ity A
cros
s Fr
actio
ns
125I apoC-III binding to HDL
WTA43T
HDL
Unbound
0
20
40
60
80
Emulsion Unbound
Perc
ent o
f Tot
al A
ctiv
ity
Activity in Lipid Emulsion and Unbound Fractions
WTA43T
**
**
0 20 40 60 800
2
4
6
8
Fraction Number
Perc
ent o
f Tot
al A
ctiv
ity A
cros
s Fr
actio
ns
125I Activity in FPLC Fractions - 24 Hours
WTA43T
VLDLLDL
HDL
Unbound
D. E.
F. G.
A. B. C.Figure S11
Nature Medicine: doi:10.1038/nm.4390

activity from FPLC-separated fractions of human TRLs incubated with 125I-labeled WT
vs. A43T apoC-III (1 µg apoC-III and 100 µg TRL protein) for 1 hour at 37 °C. Graphs
show mean ± S.D. of three replicate experiments. E. 125I activity from FPLC-separated
fractions of human HDL3 incubated with 125I-labeled WT vs. A43T apoC-III (1 µg apoC-
III and 200 µg HDL3 protein) for 1 hour at 37 °C. Graphs show ± S.D. of three replicate
experiments. F. 125I activity from FPLC-separated fractions of large lipid emulsions
incubated with 125I-labeled WT vs. A43T apoC-III for 1 hour at 37 °C. Graphs show ±
S.E.M. of three replicate experiments. G. Relative 125I activity in lipid emulsion fractions
vs. unbound protein fractions from experiments in F. Bars indicate sum of relative
amount of activity in fractions from three replicate experiments. All measurements show
mean ± S.D. where appropriate and data shows biological replicate values. All data was
replicated once by an independent repeat experiment. For A-C and G, **P<0.01,
Student’s unpaired two-tailed T-test.
Nature Medicine: doi:10.1038/nm.4390
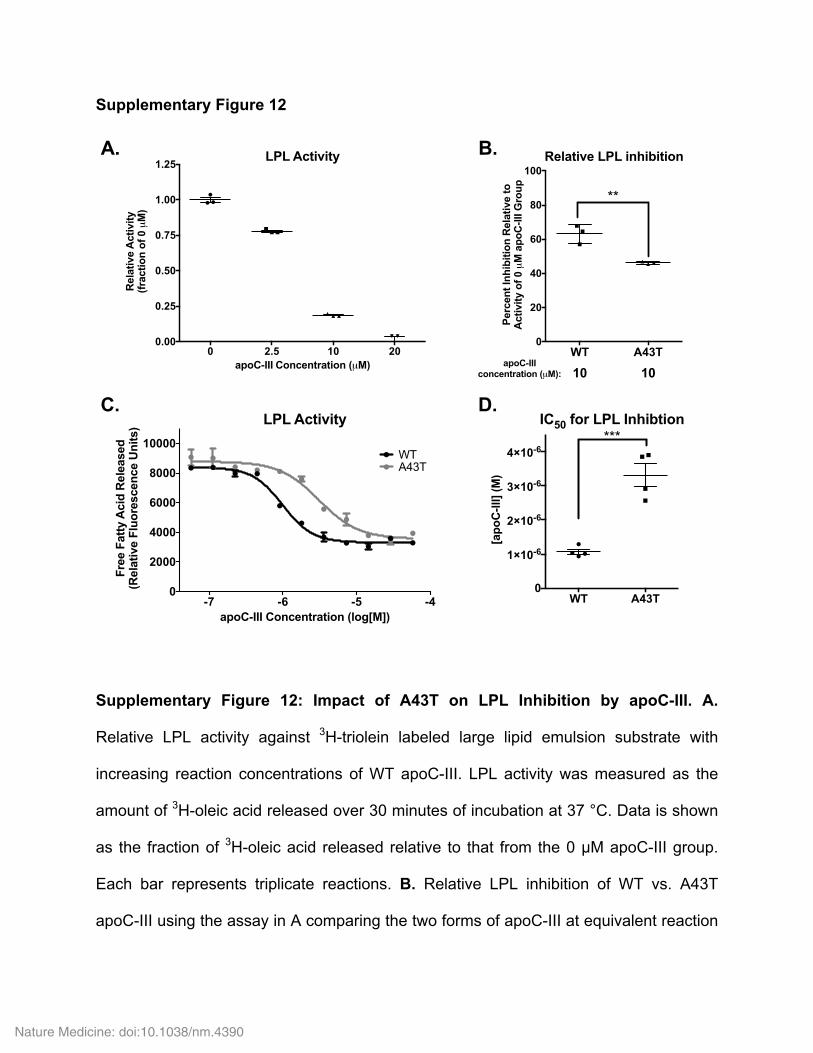
Supplementary Figure 12
Supplementary Figure 12: Impact of A43T on LPL Inhibition by apoC-III. A.
Relative LPL activity against 3H-triolein labeled large lipid emulsion substrate with
increasing reaction concentrations of WT apoC-III. LPL activity was measured as the
amount of 3H-oleic acid released over 30 minutes of incubation at 37 °C. Data is shown
as the fraction of 3H-oleic acid released relative to that from the 0 µM apoC-III group.
Each bar represents triplicate reactions. B. Relative LPL inhibition of WT vs. A43T
apoC-III using the assay in A comparing the two forms of apoC-III at equivalent reaction
0 2.5 10 200.00
0.25
0.50
0.75
1.00
1.25
apoC-III Concentration (µM)
Rel
ativ
e A
ctiv
ity
(fra
ctio
n of
0 µ
M)
LPL Activity
WT A43T0
1×10-6
2×10-6
3×10-6
4×10-6
[apo
C-II
I] (M
)
IC50 for LPL Inhibtion***
WT A43T0
20
40
60
80
100
Per
cent
Inhi
bitio
n R
elat
ive
to
Act
ivity
of 0
µM
apo
C-II
I Gro
up
Relative LPL inhibition
**
apoC-III concentration (µM): 10 10
-7 -6 -5 -40
2000
4000
6000
8000
10000
apoC-III Concentration (log[M])
Free
Fat
ty A
cid
Rel
ease
d (R
elat
ive
Fluo
resc
ence
Uni
ts) LPL Activity
WTA43T
A. B.
C. D.
Figure S12
Nature Medicine: doi:10.1038/nm.4390

concentrations. LPL inhibition is graphed as the relative difference in LPL activity
between a 0 µM apoC-III group and each of the apoC-III groups (WT and A43T). 10 µM
of WT or A43T apoC-III was used for these reactions. All reactions took place for 30
minutes at 37 °C and were performed in triplicate. C. Relative LPL activity against an
Intralipid emulsion substrate with concentrations of WT or A43T apoC-III as indicated.
LPL activity was measured as the amount of free fatty acids released for each reaction
as measured by relative light units after addition of colorimetric reagent to reaction
incubations. Each reaction was performed in triplicate. D. IC50 concentration of apoC-III
for WT vs. A43T from the activity curves in C. Each bar represents triplicate reactions.
All data was replicated twice by two independent repeat experiments. For all data
presented here, 1 µM apoC-III = 0.088 mg/dL apoC-III. All measurements show mean ±
S.E.M. where appropriate, and data and spread show technical replicate values.
**P<0.01, ***P<0.001, Student’s unpaired two-tailed T-test.
Nature Medicine: doi:10.1038/nm.4390
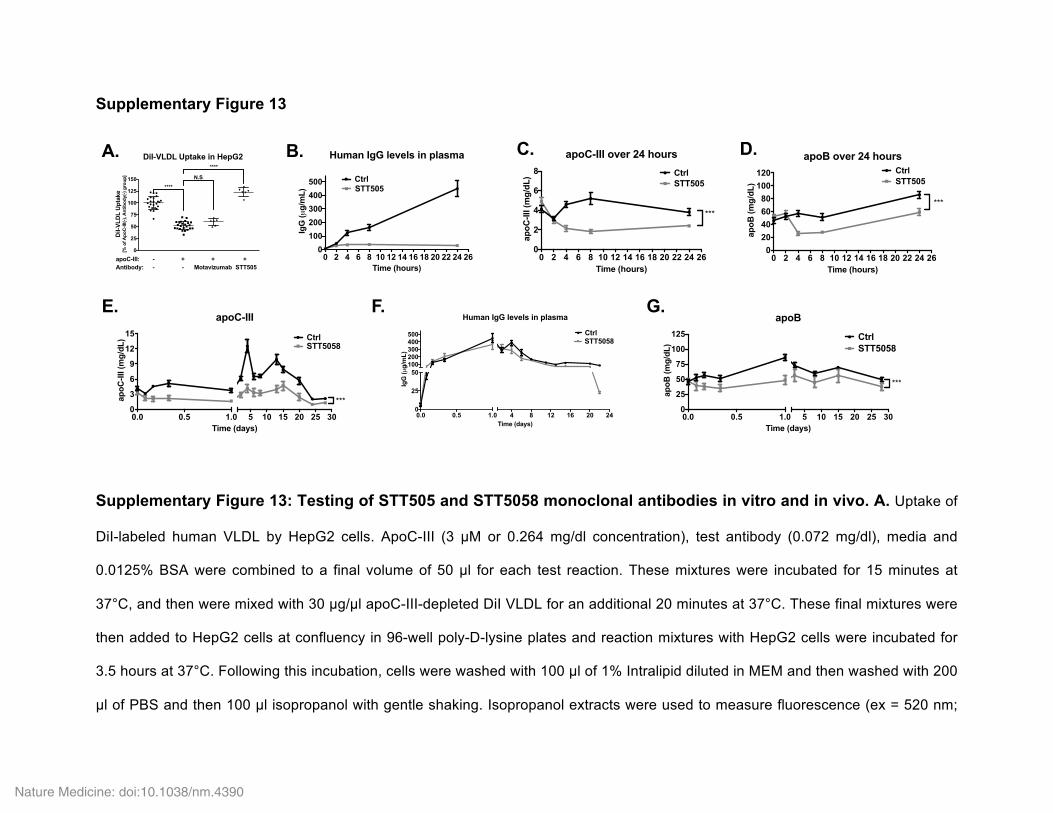
Supplementary Figure 13
Supplementary Figure 13: Testing of STT505 and STT5058 monoclonal antibodies in vitro and in vivo. A. Uptake of
DiI-labeled human VLDL by HepG2 cells. ApoC-III (3 µM or 0.264 mg/dl concentration), test antibody (0.072 mg/dl), media and
0.0125% BSA were combined to a final volume of 50 µl for each test reaction. These mixtures were incubated for 15 minutes at
37°C, and then were mixed with 30 µg/µl apoC-III-depleted DiI VLDL for an additional 20 minutes at 37°C. These final mixtures were
then added to HepG2 cells at confluency in 96-well poly-D-lysine plates and reaction mixtures with HepG2 cells were incubated for
3.5 hours at 37°C. Following this incubation, cells were washed with 100 µl of 1% Intralipid diluted in MEM and then washed with 200
µl of PBS and then 100 µl isopropanol with gentle shaking. Isopropanol extracts were used to measure fluorescence (ex = 520 nm;
0
25
50
75
100
125
150
DiI-
VLD
L U
ptak
e [%
of A
poC
-III(-
), A
ntib
ody(
-) gr
oup]
apoC-III: Antibody:
--
+-
+Motavizumab
+STT505
N.S.
****
****
DiI-VLDL Uptake in HepG2
0 2 4 6 8 10 12 14 16 18 20 22 24 260
100200300400500
Time (hours)
IgG
(µg/
mL)
Human IgG levels in plasma
STT505Ctrl
0.0 0.5 1.0 4 8 12 16 20 240
25
50100200300400500
Time (days)
IgG
(µg/
mL)
Human IgG levels in plasma
STT5058Ctrl
Figure S13A. B. C. D.
E. F. G.
0 2 4 6 8 10 12 14 16 18 20 22 24 260
2
4
6
8
Time (hours)
apoC
-III (
mg/
dL)
apoC-III over 24 hours
***
CtrlSTT505
0 2 4 6 8 10 12 14 16 18 20 22 24 260
20406080
100120
Time (hours)
apoB
(mg/
dL)
apoB over 24 hours
***
CtrlSTT505
0.0 0.5 1.0 5 10 15 20 25 300
3
6
9
12
15
Time (days)
apoC
-III (
mg/
dL)
apoC-III
***
CtrlSTT5058
0.0 0.5 1.0 5 10 15 20 25 300
25
50
75
100
125
Time (days)
apoB
(mg/
dL)
apoB
***
CtrlSTT5058
Nature Medicine: doi:10.1038/nm.4390

em = 580 nm), and this was compared to the fluorescence from a DiI VLDL standard curve to determine the amount of DiI VLDL per
volume in the cell reactions. The remaining reactions on the cell plates were lysed with 0.1 N NaOH + 0.1% SDS and protein
measured by BCA assay. The quantity of VLDL internalized was calculated as the DiI VLDL/protein ratio. For this experiment,
motavizumab was used as a control antibody and compared to STT505 prior to uptake assays for indicated groups. N=3 technical
replicates per group. Data was replicated by a repeat experiment once. B. Plasma human IgG levels following treatment of APOC3
WT AAV-expressing WT mice with control or STT505 antibody at the zero time point for up to 24 hours as measured by
autoanalyzer. C. Plasma apoC-III levels in mice from B over 24 hours as measured by autoanalyzer (correlating with the values
normalized to the control apoC-III levels shown in Figure 4B). D. Plasma apoB levels in mice from B over 24 hours as measured by
autoanalyzer (correlating with the values normalized to the control apoC-III levels shown in Figure 4D). For B-D, all data show
experiments with N=8 mice treated with STT505 antibody and N=10 control-treated mice. Each experiment was replicated once by
an independent repeat experiment. E. Plasma apoC-III levels following treatment of APOC3 WT AAV-expressing WT mice with
control or STT5058 antibody (25 mg/kg subcutaneously dosed) for 28 days as measured by autoanalyzer (correlating with the values
normalized to the control apoC-III levels shown in Figure 4J). F. Plasma human IgG levels in mice from E as measured by
autoanalyzer. G. Plasma apoB levels in mice from E as measured by autoanalyzer (correlating with the values normalized to the
control apoB levels shown in Figure 4K). For E-G, all data show experiments with N=7 mice treated with STT5058 antibody and N=7
isotype control-treated mice. Each experiment was replicated once by an independent repeat experiment. All measurements show
mean ± S.E.M. where appropriate. For A, ****P<0.0001, Student’s unpaired T-test of comparison of the indicated groups. For all
other panels, ***P<0.001, 2-way ANOVA.
Nature Medicine: doi:10.1038/nm.4390
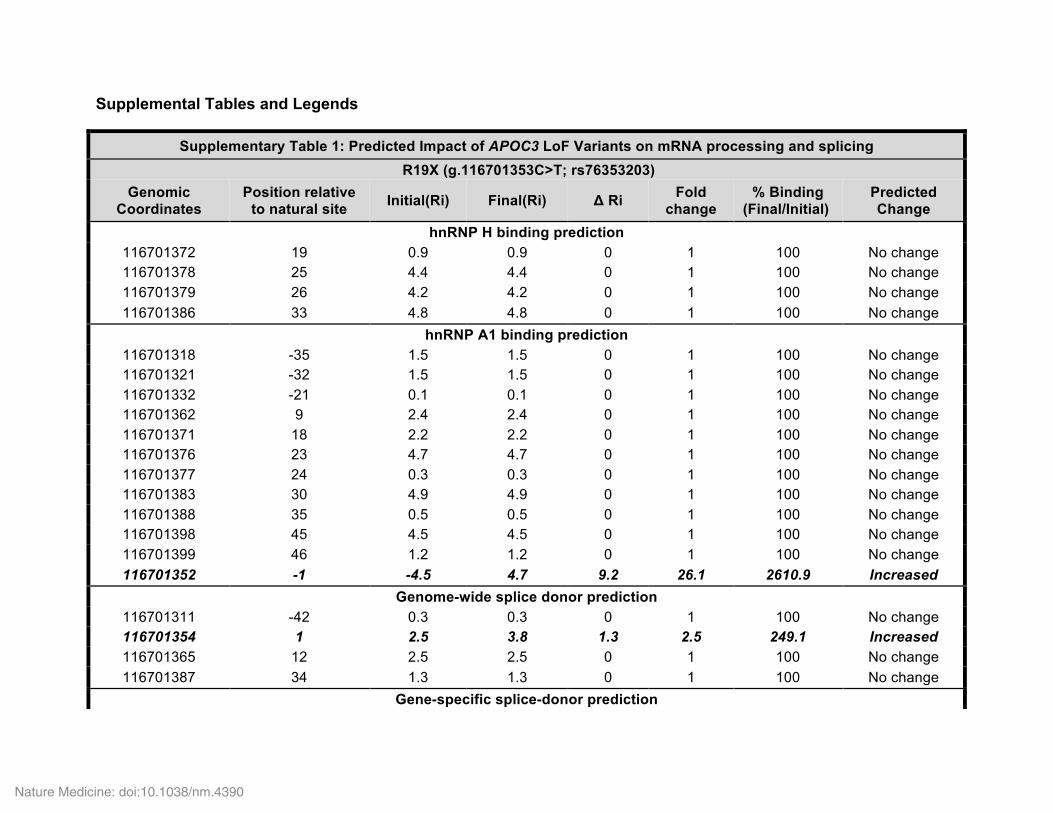
Supplemental Tables and Legends
Supplementary Table 1: Predicted Impact of APOC3 LoF Variants on mRNA processing and splicing R19X (g.116701353C>T; rs76353203)
Genomic Coordinates
Position relative to natural site Initial(Ri) Final(Ri) Δ Ri Fold
change % Binding
(Final/Initial) Predicted Change
hnRNP H binding prediction 116701372 19 0.9 0.9 0 1 100 No change 116701378 25 4.4 4.4 0 1 100 No change 116701379 26 4.2 4.2 0 1 100 No change 116701386 33 4.8 4.8 0 1 100 No change
hnRNP A1 binding prediction 116701318 -35 1.5 1.5 0 1 100 No change 116701321 -32 1.5 1.5 0 1 100 No change 116701332 -21 0.1 0.1 0 1 100 No change 116701362 9 2.4 2.4 0 1 100 No change 116701371 18 2.2 2.2 0 1 100 No change 116701376 23 4.7 4.7 0 1 100 No change 116701377 24 0.3 0.3 0 1 100 No change 116701383 30 4.9 4.9 0 1 100 No change 116701388 35 0.5 0.5 0 1 100 No change 116701398 45 4.5 4.5 0 1 100 No change 116701399 46 1.2 1.2 0 1 100 No change 116701352 -1 -4.5 4.7 9.2 26.1 2610.9 Increased
Genome-wide splice donor prediction 116701311 -42 0.3 0.3 0 1 100 No change 116701354 1 2.5 3.8 1.3 2.5 249.1 Increased 116701365 12 2.5 2.5 0 1 100 No change 116701387 34 1.3 1.3 0 1 100 No change
Gene-specific splice-donor prediction
Nature Medicine: doi:10.1038/nm.4390

116701311 -42 1.8 1.8 0 1 100 No change 116701354 1 5.2 6 0.7 1.7 165.8 Increased 116701365 12 4.2 4.2 0 1 100 No change 116701387 34 3 3 0 1 100 No change
Splice branch-point prediction 116701396 43 0.4 0.4 0 1 100 No change
Genome-wide splice-acceptor prediction 116701334 -19 0 0 0 1 100 No change
Gene-specific splice-acceptor prediction 116701334 -19 3.3 3.3 0 1 100 No change 116701335 -18 0 0 0 1 100 No change 116701343 -10 1.2 1.2 0 1 100 No change 116701358 5 1.3 1.5 0.2 1.1 114 Increased 116701357 4 -0.5 0.1 0.6 1.1 107.8 Increased
IVS2+1 G>A (g.116701354 G>A; rs138326449) Genomic
Coordinates Position relative
to natural site Initial(Ri) Final(Ri) Δ Ri Fold change
% Binding (Final/Initial)
Predicted Change
hnRNP H binding prediction 116701372 18 0.9 0.9 0 1 100 No change 116701378 24 4.4 4.4 0 1 100 No change 116701379 25 4.2 4.2 0 1 100 No change 116701386 32 4.8 4.8 0 1 100 No change
hnRNP A1 binding prediction 116701318 -36 1.5 1.5 0 1 100 No change 116701321 -33 1.5 1.5 0 1 100 No change 116701332 -22 0.1 0.1 0 1 100 No change 116701362 8 2.4 2.4 0 1 100 No change 116701371 17 2.2 2.2 0 1 100 No change 116701376 22 4.7 4.7 0 1 100 No change 116701377 23 0.3 0.3 0 1 100 No change
Nature Medicine: doi:10.1038/nm.4390

116701383 29 4.9 4.9 0 1 100 No change 116701388 34 0.5 0.5 0 1 100 No change 116701398 44 4.5 4.5 0 1 100 No change 116701399 45 1.2 1.2 0 1 100 No change
Genome-wide splice donor prediction 116701311 -43 0.3 0.3 0 1 100 No change 116701365 11 2.5 2.5 0 1 100 No change 116701387 33 1.3 1.3 0 1 100 No change 116701354 0 2.5 -16.1 -18.6 -5.8 NA Decreased
Gene-specific splice-donor prediction 116701311 -43 1.8 1.8 0 1 100 No change 116701365 11 4.2 4.2 0 1 100 No change 116701387 33 3 3 0 1 100 No change 116701354 0 5.2 -7.6 -12.8 -38 NA Decreased
Splice branch-point prediction 116701396 42 0.4 0.4 0 1 100 No change
Genome-wide splice-acceptor prediction 116701334 -20 0 0 0 1 100 No change
Gene-specific splice-acceptor prediction 116701334 -20 3.3 3.3 0 1 100 No change 116701335 -19 0 0 0 1 100 No change 116701343 -11 1.2 1.2 0 1 100 No change 116701358 4 1.3 1.6 0.3 1.2 123.3 Increased
A43T (g.116701560 G>A; rs147210663) Genomic
Coordinates Position relative
to natural site Initial(Ri) Final(Ri) Δ Ri Fold change
% Binding (Final/Initial)
Predicted Change
hnRNP H binding prediction 116701531 -29 1 1 0 1 100 No change 116701580 20 1.2 1.2 0 1 100 No change
hnRNP A1 binding prediction
Nature Medicine: doi:10.1038/nm.4390

116701530 -30 4.9 4.9 0 1 100 No change 116701539 -21 4.6 4.6 0 1 100 No change 116701563 3 4.6 4.6 0 1 100 No change 116701572 12 2.4 2.4 0 1 100 No change 116701584 24 5.2 5.2 0 1 100 No change 116701585 25 0.1 0.1 0 1 100 No change 116701593 33 3 3 0 1 100 No change
Genome-wide splice donor prediction 116701596 36 2.2 2.2 0 1 100 No change
Gene-specific splice-donor prediction 116701596 36 4.2 4.2 0 1 100 No change
Splice branch-point prediction 116701521 -39 1.3 1.3 0 1 100 No change 116701555 -5 0.9 0.9 0 1 100 No change 116701564 4 0.4 0.9 0.5 1.4 140 Increased 116701575 15 1.1 1.1 0 1 100 No change
Genome-wide splice-acceptor prediction 116701532 -28 6 6 0 1 100 No change
Gene-specific splice-acceptor prediction 116701532 -28 9.7 9.7 0 1 100 No change 116701607 47 0.3 0.3 0 1 100 No change
IVS3+1 G>T (g.116701613G>T; rs140621530) Genomic
Coordinates Position relative
to natural site Initial(Ri) Final(Ri) Δ Ri Fold change
% Binding (Final/Initial)
Predicted Change
hnRNP H binding prediction 116701580 -33 1.2 1.2 0 1 100 No change 116701606 -7 3 -1.9 -4.9 -8.2 NA Decreased
hnRNP A1 binding prediction 116701563 -50 4.6 4.6 0 1 100 No change 116701572 -41 2.4 2.4 0 1 100 No change
Nature Medicine: doi:10.1038/nm.4390
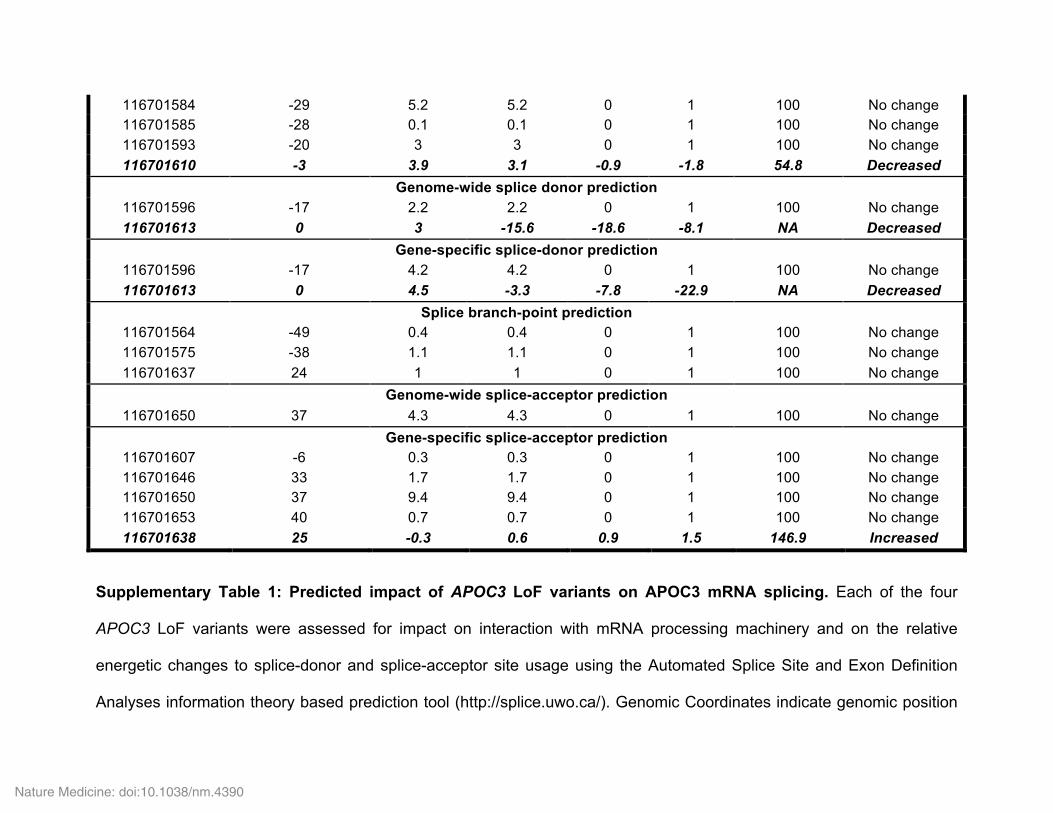
116701584 -29 5.2 5.2 0 1 100 No change 116701585 -28 0.1 0.1 0 1 100 No change 116701593 -20 3 3 0 1 100 No change 116701610 -3 3.9 3.1 -0.9 -1.8 54.8 Decreased
Genome-wide splice donor prediction 116701596 -17 2.2 2.2 0 1 100 No change 116701613 0 3 -15.6 -18.6 -8.1 NA Decreased
Gene-specific splice-donor prediction 116701596 -17 4.2 4.2 0 1 100 No change 116701613 0 4.5 -3.3 -7.8 -22.9 NA Decreased
Splice branch-point prediction 116701564 -49 0.4 0.4 0 1 100 No change 116701575 -38 1.1 1.1 0 1 100 No change 116701637 24 1 1 0 1 100 No change
Genome-wide splice-acceptor prediction 116701650 37 4.3 4.3 0 1 100 No change
Gene-specific splice-acceptor prediction 116701607 -6 0.3 0.3 0 1 100 No change 116701646 33 1.7 1.7 0 1 100 No change 116701650 37 9.4 9.4 0 1 100 No change 116701653 40 0.7 0.7 0 1 100 No change 116701638 25 -0.3 0.6 0.9 1.5 146.9 Increased
Supplementary Table 1: Predicted impact of APOC3 LoF variants on APOC3 mRNA splicing. Each of the four
APOC3 LoF variants were assessed for impact on interaction with mRNA processing machinery and on the relative
energetic changes to splice-donor and splice-acceptor site usage using the Automated Splice Site and Exon Definition
Analyses information theory based prediction tool (http://splice.uwo.ca/). Genomic Coordinates indicate genomic position
Nature Medicine: doi:10.1038/nm.4390

on chromosome 11 for which a particular prediction is made. Position relative to the natural site indicates the position on
the (+) strand of the variant assessed relative to the Genomic Coordinates. Initial and Final Ri indicate the information
content of the base in the WT or variant form, respectively, and ∆Ri indicates the difference between Final Ri and Initial
Ri. Fold change indicates the difference in binding affinity for a site in the variant vs. WT form based on the ∆RI. %
Binding indicates the percentage increase or decrease in binding propensity for a given binding factor. Predicted change
indicates the overall predicted effect for the given measure based on the ∆Ri and predicted fold change for a given
position.
Nature Medicine: doi:10.1038/nm.4390
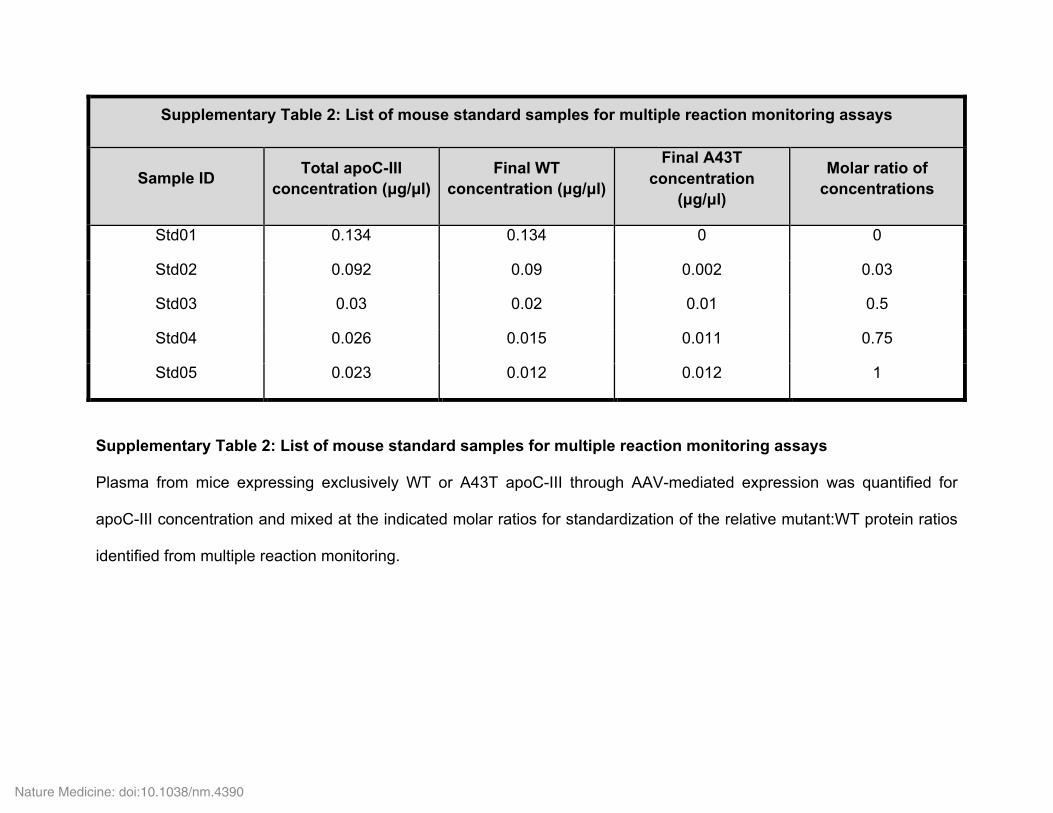
Supplementary Table 2: List of mouse standard samples for multiple reaction monitoring assays
Sample ID Total apoC-III concentration (µg/µl)
Final WT concentration (µg/µl)
Final A43T concentration
(µg/µl)
Molar ratio of concentrations
Std01 0.134 0.134 0 0
Std02 0.092 0.09 0.002 0.03
Std03 0.03 0.02 0.01 0.5
Std04 0.026 0.015 0.011 0.75
Std05 0.023 0.012 0.012 1
Supplementary Table 2: List of mouse standard samples for multiple reaction monitoring assays
Plasma from mice expressing exclusively WT or A43T apoC-III through AAV-mediated expression was quantified for
apoC-III concentration and mixed at the indicated molar ratios for standardization of the relative mutant:WT protein ratios
identified from multiple reaction monitoring.
Nature Medicine: doi:10.1038/nm.4390

Supplementary Table 3: List of selected apoC-III peptides and transitions for multiple reaction monitoring assays
Protein Peptide name Peptide sequence CE Light SRM (m/z) Heavy SRM (m/Z)
apoC-III Thr43 Oxi
DASLLSFM(oxi)QGYM(oxi)KHATKTTK 24.7 573.3 → 702.0 635.3 597.6
568.1 575.0 →704.4 637.6 597.6
568.6
apoC-III Ala43 Oxi
DASLLSFM(oxi)QGYM(oxi)KHATKTAK 24.5 565.8 → 692.0 625.3 587.6
558.6 567.5 → 694.3 627.7 587.6
558.6
apoC-III DGF DGFSSLK 14.5 377.2 → 581.3 434.3 347.2
apoC-III DYW DYWSTVK 17.1 449.7 → 620.3 434.3 347.3
SERPINB14 DIL DILNQITKPN 21.8 578.3 → 459.3 358.2 230.1
Supplementary Table 3: List of selected ApoC-III peptides and transitions for multiple reaction monitoring assays
Peptides spanning the 23rd residue of apoC-III were analyzed for fragmentation and transition ion mass to charge (m/Z)
ratios were monitored in the multiple reaction monitoring assays. Heavy amino acids are underlined (13C, 15N). CE,
collision energy.
Nature Medicine: doi:10.1038/nm.4390
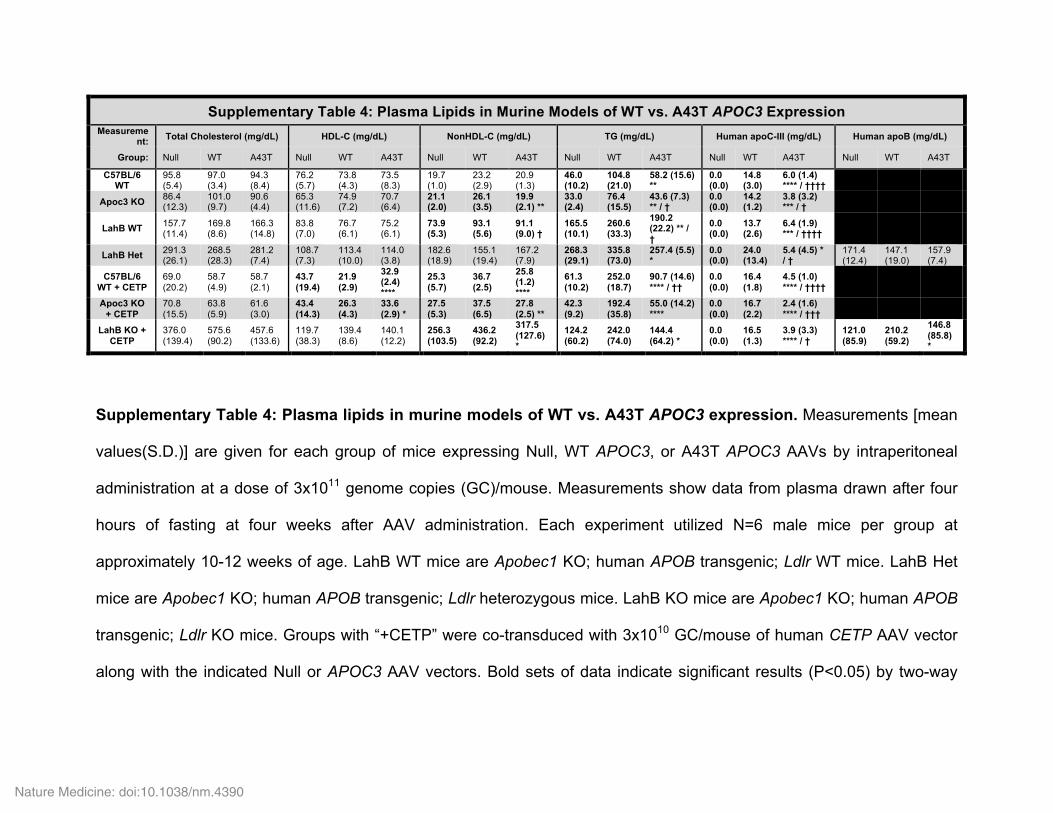
Supplementary Table 4: Plasma Lipids in Murine Models of WT vs. A43T APOC3 Expression Measureme
nt: Total Cholesterol (mg/dL) HDL-C (mg/dL) NonHDL-C (mg/dL) TG (mg/dL) Human apoC-III (mg/dL) Human apoB (mg/dL)
Group: Null WT A43T Null WT A43T Null WT A43T Null WT A43T Null WT A43T Null WT A43T
C57BL/6 WT
95.8 (5.4)
97.0 (3.4)
94.3 (8.4)
76.2 (5.7)
73.8 (4.3)
73.5 (8.3)
19.7 (1.0)
23.2 (2.9)
20.9 (1.3)
46.0 (10.2)
104.8 (21.0)
58.2 (15.6) **
0.0 (0.0)
14.8 (3.0)
6.0 (1.4) **** / ††††
Apoc3 KO 86.4 (12.3)
101.0 (9.7)
90.6 (4.4)
65.3 (11.6)
74.9 (7.2)
70.7 (6.4)
21.1 (2.0)
26.1 (3.5)
19.9 (2.1) **
33.0 (2.4)
76.4 (15.5)
43.6 (7.3) ** / †
0.0 (0.0)
14.2 (1.2)
3.8 (3.2) *** / †
LahB WT 157.7 (11.4)
169.8 (8.6)
166.3 (14.8)
83.8 (7.0)
76.7 (6.1)
75.2 (6.1)
73.9 (5.3)
93.1 (5.6)
91.1 (9.0) †
165.5 (10.1)
260.6 (33.3)
190.2 (22.2) ** / †
0.0 (0.0)
13.7 (2.6)
6.4 (1.9) *** / ††††
LahB Het 291.3 (26.1)
268.5 (28.3)
281.2 (7.4)
108.7 (7.3)
113.4 (10.0)
114.0 (3.8)
182.6 (18.9)
155.1 (19.4)
167.2 (7.9)
268.3 (29.1)
335.8 (73.0)
257.4 (5.5) *
0.0 (0.0)
24.0 (13.4)
5.4 (4.5) * / †
171.4 (12.4)
147.1 (19.0)
157.9 (7.4)
C57BL/6 WT + CETP
69.0 (20.2)
58.7 (4.9)
58.7 (2.1)
43.7 (19.4)
21.9 (2.9)
32.9 (2.4) ****
25.3 (5.7)
36.7 (2.5)
25.8 (1.2) ****
61.3 (10.2)
252.0 (18.7)
90.7 (14.6) **** / ††
0.0 (0.0)
16.4 (1.8)
4.5 (1.0) **** / ††††
Apoc3 KO + CETP
70.8 (15.5)
63.8 (5.9)
61.6 (3.0)
43.4 (14.3)
26.3 (4.3)
33.6 (2.9) *
27.5 (5.3)
37.5 (6.5)
27.8 (2.5) **
42.3 (9.2)
192.4 (35.8)
55.0 (14.2) ****
0.0 (0.0)
16.7 (2.2)
2.4 (1.6) **** / †††
LahB KO + CETP
376.0 (139.4)
575.6 (90.2)
457.6 (133.6)
119.7 (38.3)
139.4 (8.6)
140.1 (12.2)
256.3 (103.5)
436.2 (92.2)
317.5 (127.6) *
124.2 (60.2)
242.0 (74.0)
144.4 (64.2) *
0.0 (0.0)
16.5 (1.3)
3.9 (3.3) **** / †
121.0 (85.9)
210.2 (59.2)
146.8 (85.8) *
Supplementary Table 4: Plasma lipids in murine models of WT vs. A43T APOC3 expression. Measurements [mean
values(S.D.)] are given for each group of mice expressing Null, WT APOC3, or A43T APOC3 AAVs by intraperitoneal
administration at a dose of 3x1011 genome copies (GC)/mouse. Measurements show data from plasma drawn after four
hours of fasting at four weeks after AAV administration. Each experiment utilized N=6 male mice per group at
approximately 10-12 weeks of age. LahB WT mice are Apobec1 KO; human APOB transgenic; Ldlr WT mice. LahB Het
mice are Apobec1 KO; human APOB transgenic; Ldlr heterozygous mice. LahB KO mice are Apobec1 KO; human APOB
transgenic; Ldlr KO mice. Groups with “+CETP” were co-transduced with 3x1010 GC/mouse of human CETP AAV vector
along with the indicated Null or APOC3 AAV vectors. Bold sets of data indicate significant results (P<0.05) by two-way
Nature Medicine: doi:10.1038/nm.4390

ANOVA. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001, comparison of WT and A43T groups by Tukey’s post-test after
two-way ANOVA. † P<0.05, †† P<0.01, ††† P<0.001, †††† P<0.0001, comparison of Null and A43T groups by Tukey’s post-
test after two-way ANOVA.
Nature Medicine: doi:10.1038/nm.4390