8 microbial leaching of metals - wiley-vch · pdf file192 8 microbial leaching of metals 1...
TRANSCRIPT

8 Microbial Leaching of Metals
HELMUT BRANDLZürich, Switzerland
1 Introduction 1922 Terminology 1923 Historical Background 1924 Principles of Microbial Metal Leaching 194
4.1 Leaching Mechanisms 1944.2 Models of Leaching Mechanisms 1944.3 Factors Influencing Bioleaching 1974.4 Bacterial Attachment on Mineral Surfaces 200
5 Microbial Diversity in Bioleaching Environments 2006 Case Studies of Bioleaching Applications 208
6.1 Commercial-Scale Copper Ore Bioleaching 2086.2 Reactor Bioleaching of Fly Ash 2106.3 Shake Flask Bioleaching of Electronic Scrap 212
7 Economics of Metal Bioleaching 2138 Perspectives of Bioleaching Technology 214
8.1 Heterotrophic Leaching 2148.2 Leaching Under Thermophilic Conditions 2158.3 Tapping Microbial Diversity 2158.4 Treatment of Solid Wastes 2158.5 Bioremediation of Metal-Contaminated Sites 215
9 Conclusion 21710 References 217

192 8 Microbial Leaching of Metals
1 Introduction
Future sustainable development requiresmeasures to reduce the dependence on non-renewable raw materials and the demand forprimary resources. New resources for metalsmust be developed with the aid of novel tech-nologies. in addition, improvement of alredyexisting mining techniques can result in metalrecovery from sources that have not been ofeconomical interest until today. Metal-winningprocesses based on the activity of microorgan-isms offer a possibility to obtain metals frommineral resources not accessible by conven-tional mining (BOSECKER, 1997; BRIERLEY,1978; BRYNER et al., 1954; TORMA and BAN-HEGYI, 1984). Microbes such as bacteria andfungi convert metal compounds into theirwater-soluble forms and are biocatalysts ofthese leaching processes. Additionally, apply-ing microbiological solubilization processes, itis possible to recover metal values from in-dustrial wastes which can serve as secondaryraw materials.
2 TerminologyIn general, bioleaching is a process de-
scribed as being “the dissolution of metalsfrom their mineral source by certain naturallyoccurring microorganisms” or “the use of mi-croorganisms to transform elements so thatthe elements can be extracted from a materialwhen water is filtered trough it” (ATLAS andBARTHA, 1997; PARKER, 1992). Additionally,the term “biooxidation” is also used (HANS-FORD and MILLER, 1993). There are, however,some small differences by definition (BRIER-LEY, 1997): Usually, “bioleaching” is referringto the conversion of solid metal values intotheir water soluble forms using microorgan-isms. In the case of copper, copper sulfide ismicrobially oxidized to copper sulfate andmetal values are present in the aqueous phase.Remaining solids are discarded. “Biooxida-tion” describes the microbiological oxidationof host minerals which contain metal com-pounds of interest.As a result, metal values re-main in the solid residues in a more concen-
trated form. In gold mining operations, biooxi-dation is used as a pretreatment process to(partly) remove pyrite or arsenopyrite. Thisprocess is also called “biobeneficiation” wheresolid materials are refined and unwanted im-purities are removed (GROUDEV, 1999; STRAS-SER et al., 1993). The terms “biomining”, “bio-extraction”, or “biorecovery” are also appliedto describe the mobilization of elements fromsolid materials mediated by bacteria and fungi(HOLMES, 1991; MANDL et al., 1996; RAWLINGS,1997; WOODS and RAWLINGS, 1989). “Biomin-ing” concerns mostly applications of microbialmetal mobilization processes in large-scale operations of mining industries for an eco-nomical metal recovery.
The area of “biohydrometallurgy” covers bioleaching or biomining processes (ROSSI,1990). Biohydrometallurgy represents an in-terdisciplinary field where aspects of microbiol-ogy (especially geomicrobiology), geochemis-try, biotechnology, hydrometallurgy, mineralo-gy, geology, chemical engineering, and miningengineering are combined. Hydrometallurgyis defined as the treatment of metals and met-al-containing materials by wet processes anddescribes “the extraction and recovery of met-als from their ores by processes in which aque-ous solutions play a predominant role” (PARK-ER, 1992). Rarely, the term “biogeotechnolo-gy” is also used instead of biohydrometallurgy(FARBISZEWSKA et al., 1994).
3 Historical BackgroundOne of the first reports where leaching
might have been involved in the mobilizationof metals is given by the Roman writer GaiusPlinius Secundus (23–79 A.D.). In his work onnatural sciences, Plinius describes how copperminerals are obtained using a leaching process(KÖNIG, 1989a, b). The translation reads ap-proximately as follows:“Chrysocolla is a liquidin the before mentioned gold mines runningfrom the gold vein. In cold weather during thewinter the sludge freezes to the hardness ofpumice. It is known from experience that themost wanted [chrysocolla] is formed in coppermines, the following in silver mines. The liquid

3 Historical Background 193
is also found in lead mines although it is of mi-nor value. In all these mines chrysocolla is alsoartificially produced by slowly passing waterthrough the mine during the winter until themonth of June; subsequently, the water is evap-orated in June and July. It is clearly demon-strated that chrysocolla is nothing but a de-composed vein.”
The German physician and mineralogistGeorgius Agricola (1494–1555) describes inhis work de re metallica also techniques for therecovery of copper that are based on the leach-ing of copper-containing ores (SCHIFFNER,1977). A woodcut from his book illustrates the (manual) transport of metal-containingleachates from mines and their evaporation inthe sunlight (Fig. 1).
The Rio Tinto mines in south-western Spainare usually considered the cradle of biohydro-metallurgy. These mines have been exploitedsince pre-Roman times for their copper, gold,and silver values. However, with respect tocommercial bioleaching operations on an in-dustrial scale, biohydrometallurgical tech-niques had been introduced to the Tharsis
mine in Spain 10 years earlier (SALKIELD,1987).As a consequence to the ban of open airore roasting and its resulting atmospheric sul-fur emissions in 1878 in Portugal, hydrometal-lurgical metal extraction has been taken intoconsideration in other countries more intense-ly. In addition to the ban, cost savings were an-other incentive for the development: Heapleaching techniques were assumed to reducetransportation costs and to allow the employ-ment of locomotives and wagons for other ser-vices (SALKIELD, 1987). From 1900 on, no openair roasting of low-grade ore was conducted atthe Rio Tinto mines.
Efforts to establish bioleaching at the RioTinto mines had been undertaken in the begin-ning of the 1890s. Heaps (10 m in height) oflow-grade ore (containing 0.75% Cu) werebuilt and left for one to three years for “natu-ral” decomposition (SALKIELD, 1987). 20 to25% of the copper left in the heaps were re-covered annually. It was calculated that ap-proximately 200,000 t of rough ore could betreated in 1896. Although industrial leachingoperations were conducted at the Rio Tintomines for several decades, the contribution ofbacteria to metal solubilization was confirmedonly in 1961, when Thiobacillus ferrooxidanswas identified in the leachates.
Early reports state that factors affecting bio-leaching operations were the height of theheap, particle size, initial ore washing withacid, and temperature control to about 50 °C(SALKIELD, 1987). Another critical factor wasthe supply of water for the leaching heaps. Al-though usually acidic mine waters were usedfor ore processing, 4 billion liters of freshwaterwere required annually (SALKIELD, 1987).
Although metal leaching from mineral re-sources has a very long historical record (EHR-LICH, 1999; ROSSI, 1990) and although the oxi-dation of reduced sulfur compounds and ele-mental sulfur resulting in the formation of sul-furic acid was demonstrated already in the1880s (WINOGRADSKY, 1887), the oxidation ofmetal sulfides was not described until 1922when mobilization of zinc from zinc sulfidewas investigated (RUDOLFS, 1922; RUDOLFS
and HELBRONNER, 1922). It was found that thetransformation of zinc sulfide to zinc sulfatewas microbially mediated. Based on these re-sults, the economic recovery of zinc from zinc-
Fig. 1. Woodcut from the book de re metallica writ-ten by Georgius Agricola (1494–1555) illustratingthe manual recovery of copper-containing mineeffluents which are collected in wooden basins andconcentrated in the sun.

containing ores by biological methods wasproposed. In 1947, Thiobacillus ferrooxidanswas identified as part of the microbial commu-nity found in acid mine drainage (COLMER andHINKLE, 1947). A first patent was granted in1958 (ZIMMERLEY et al., 1958). The patent de-scribes a cyclic process where a ferric sulfate/sulfuric acid lixiviant solution is used for metalextraction, regenerated by aeration (ferrousiron oxidation by iron-oxidizing organisms),and reused in a next leaching stage.
4 Principles of MicrobialMetal Leaching
4.1 Leaching Mechanisms
Mineralytic effects of bacteria and fungi onminerals are based mainly on three principles,namely acidolysis, complexolysis, and redoxo-lysis. Microorganisms are able to mobilize met-als by (1) the formation of organic or inorgan-ic acids (protons); (2) oxidation and reductionreactions; and (3) the excretion of complexingagents. Sulfuric acid is the main inorganic acidfound in leaching environments. It is formedby sulfur-oxidizing microorganisms such asthiobacilli.A series of organic acids are formedby bacterial (as well as fungal) metabolism resulting in organic acidolysis, complex andchelate formation (BERTHELIN, 1983). A kinet-ic model of the coordination chemistry of min-eral solubilization has been developed whichdescribes the dissolution of oxides by the pro-tonation of the mineral surface as well as thesurface concentration of suitable complex-forming ligands such as oxalate, malonate,citrate, and succinate (FURRER and STUMM,1986). Proton-induced and ligand-inducedmineral solubilization occurs simultaneouslyin the presence of ligands under acidic condi-tions.
4.2 Models of Leaching Mechanisms
Originally, a model with two types of mech-anisms which are involved in the microbialmobilization of metals has been proposed(EWART and HUGHES, 1991; SILVERMAN andEHRLICH, 1964): (1) Microorganisms can oxi-dize metal sulfides by a “direct” mechanismobtaining electrons directly from the reducedminerals. Cells have to be attached to the min-eral surface and a close contact is needed. Theadsorption of cells to suspended mineral parti-cles takes place within minutes or hours. Thishas been demonstrated using either radioac-tively labeled Thiobacillus ferrooxidans cellsgrown on NaH14CO3 or the oxidative capacityof bacteria attached to the mineral surface(ESCOBAR et al., 1996). Cells adhere selectivelyto mineral surfaces occupying preferentially ir-regularities of the surface structure (EDWARDS
et al., 1999; EWART and HUGHES, 1991). In ad-dition, a chemotactic behavior to copper, iron,or nickel ions has been demonstrated for Lep-tospirillum ferrooxidans (ACUNA et al., 1992).Genes involved in the chemotaxis were alsodetected in Thiobacillus ferrooxidans andThiobacillus thiooxidans (ACUNA et al., 1992).(2) The oxidation of reduced metals throughthe “indirect” mechanism is mediated by ferriciron (Fe3c) originating from the microbial oxidation of ferrous iron (Fe2c) compoundspresent in the minerals. Ferric iron is an oxidiz-ing agent and can oxidize, e.g., metal sulfidesand is (chemically) reduced to ferrous ironwhich, in turn, can be microbially oxidizedagain (EWART and HUGHES, 1991). In this case,iron has a role as electron carrier. It was pro-posed that no direct physical contact is neededfor the oxidation of iron.
In many cases it was concluded that the “di-rect” mechanism dominates over the “indi-rect” mostly due to the fact that “direct” wasequated with “direct physical contact”. Thisdomination has been observed for the oxida-tion of covellite or pyrite in studies employingmesophilic T. ferrooxidans and thermophilicAcidianus brierleyi in bioreactors which con-sisted of chambers separated with dialysismembranes to avoid physical contact (LARS-SON et al., 1993; POGLIANI et al., 1990). How-
194 8 Microbial Leaching of Metals

4 Principles of Microbial Metal Leaching 195
ever, the attachment of microorganisms onsurfaces is not an indication per se for the exis-tence of a direct mechanism (EDWARDS et al.,1999). The term “contact leaching” has beenintroduced to indicate the importance of bac-terial attachment to mineral surfaces (TRI-BUTSCH, 1999).
The following equations describe the “di-rect” and “indirect” mechanism for the oxida-tion of pyrite (MURR, 1980; SAND et al., 1999):
direct:
2FeS2c7O2c2H2O ] 2FeSO4
c2H2SO4 (1)
indirect:
4FeSO4cO2
c2H2SO4 ]2Fe2(SO4)3c2H2O (2)
FeS2cFe2(SO4)3 ] 3FeSO4
c2S (3)
2Sc3O2cH2O ] 2H2SO4 (4)
However, the model of “direct” and “indirect”metal leaching is still under discussion. Re-cently, this model has been revised and re-placed by another one which is not dependenton the differentiation between a “direct” andan “indirect” leaching mechanisms (SAND etal., 1995, 1999). All facts have been combinedand a mechanism has been developed which ischaracterized by the following features: (1)cells have to be attached to the minerals and inphysical contact with the surface; (2) cells formand excrete exopolymers; (3) these exopoly-meric cell envelopes contain ferric iron com-pounds which are complexed to glucuronicacid residues. These are part of the primary at-tack mechanism; (4) thiosulfate is formed asintermediate during the oxidation of sulfurcompounds; (5) sulfur or polythionate gran-ules are formed in the periplasmatic space orin the cell envelope.
Thiosulfate and traces of sulfite have beenfound as intermediates during the oxidation ofsulfur (SHRIHARI et al., 1993). Sulfur granules(colloidal sulfur) have been identified as ener-gy reserves in the exopolymeric capsule
T. thiooxidans
chemical oxidation
T. ferrooxidans, L. ferrooxidans
thiobacilli
around cells of T. ferrooxidans during growthon synthetic pyrite films (ROJAS et al., 1995).“Footprints” of organic films containing col-loidal sulfur granules are left on the mineralsurface upon detachment of the bacteria.
From the existing data two “indirect” leach-ing mechanisms have been proposed whereasno evidence for a “direct” enzymatically medi-ated process has been found (SAND et al.,1999).The mineral structure is the determiningfactor for the prevailing type of leachingmechanism. In the “thiosulfate mechanism”thiosulfate is the main intermediate resultingfrom the oxidation of pyrite, molybdenite, ortungstenite. Polysulfide and elemental sulfurare the main intermediates in the “polysulfidemechanism” during the oxidation of galena,sphalerite, chalcopyrite, hauerite, orpiment, orrealgar. The presence of iron(III) at the begin-ning of mineral degradation is an importantprerequisite (SAND et al., 1999).
The following equations summarize the oxi-dation mechanisms (SAND et al., 1999):
Thiosulfate mechanism (found for FeS2, MoS2,WS2):
FeS2c6Fe3cc3H2O ] S2O32cP7Fe2c
c6Hc (5)
S2O32Pc8Fe3cc5H2O ] 2SO4
2Pc8Fe2c
c10Hc (6)
Polysulfide mechanism (found for PbS, CuFeS2,ZnS, MnS2, As2S3, As3S4):
2MSc2Fe3cc2Hc ] 2M2ccH2Sn
c2Fe2c (7)
H2Snc2Fe3c ] 0.25 S8c2Fe2cc2Hc (8)
0.25 S8c3O2c2H2O ] 2SO42Pc4Hc (9)
Several biomolecules are involved in theaerobic respiration on reduced sulfur and ironcompounds. It has been found that up to 5% ofsoluble proteins of T. ferrooxidans is made ofan acid stable blue copper protein, called rus-ticyanin (BLAKE et al., 1993). Additionally, theiron(II) respiratory system contains a (puta-tive) green copper protein, two types of cyto-chrome c, one or more types of cytochrome a,

a porin, and an iron(II)-sulfate chelate (BLAKE
et al., 1993). The acid stability of rusticyaninsuggests that it is located in the periplasmicspace. Figure 2 shows a scheme of the modelwhich combines the electron transport se-quence proposed earlier with concepts stem-ming from the debate on “direct”/“indirect”leaching mechanisms (BLAKE and SHUTE,1994; BLAKE et al., 1993; HAZRA et al., 1992;SAND et al., 1995).
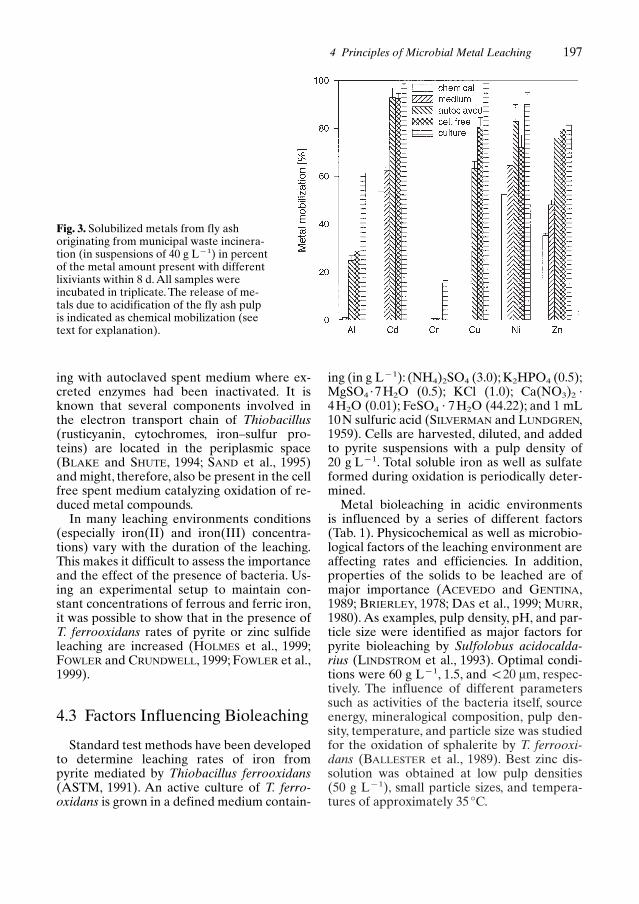
Some details of the metal mobilizationmechanism, the importance of the presenceand attachment of microorganisms and theiractive contribution have been demonstratedfor the leaching of fly ash from municipalwaste incineration (MWI) (BROMBACHER etal., 1998). Generally, several mechanisms ofmetal mobilization can be distinguished: (1)Contact leaching effect on the release of met-als. Stock cultures of Thiobacillus ferrooxidansand Thiobacillus thiooxidans were added toash suspensions and cells were in direct con-tact with the fly ash. Growth of thiobacillimight be stimulated by increased energy avail-ability from oxidation of reduced solid parti-cles. (2) Metal solubilization by metabolicallyactive (enzymatic) compounds in the absenceof bacterial cells. Stock cultures were filteredto obtain the cell free spent medium. This me-dium was used for leaching. (3) Metal solubil-ization by non-enzymatic extracellular meta-bolic products. Cell free spent medium (see 2)
was autoclaved to obtain a sterile leaching so-lution without enzymatic activities and to eval-uate the leaching ability of acid formed. (4)Leaching by fresh medium. Fresh non-inocul-ated and sterile medium was added to the flyash suspension and used as control. (5) Chem-ical leaching due to the preparation of the ashsuspension (acidification to pH 5.4). Certainelements such as, e.g., Cd or Zn might be chem-ically mobilized already during acidification.
MWI fly ash contains reduced copper spe-cies (chalcocite {Cu2S} or cuprite {Cu2O})whereas zinc and others are present in theirfully oxidized forms (BROMBACHER et al.,1998).Therefore, copper release from fly ash isdirectly affected and enhanced by T. ferrooxi-dans, whereas Zn, as well as Al, Cd, Cr, and Ni,are released primarily due to the acidic envi-ronment. Acidification of the fly ash pulp(chemical mobilization) led already to consid-erable extraction yields for Cd, Ni, and Zn andcould slightly be increased using non-inoculat-ed sterile medium as lixiviant (Fig. 3). By com-paring leached amounts of copper by filteredcell free spent medium with autoclaved sterilespent medium, it was concluded that signifi-cant amounts of copper were mobilized – incontrast to other elements – by metabolicproducts of T. ferrooxidans. Leaching with cellfree spent medium indicating a solubilizingmechanism due to extracellular componentswas significantly more effective than a leach-
196 8 Microbial Leaching of Metals
Fig. 2. Schematic mechanistic bio-leaching model (after HAZRA etal., 1992; SAND et al., 1995, 1999;SCHIPPERS et al., 1996; RAWLINGS,1999). C: cytoplasm; CM: cellmembrane; PS: periplasmaticspace; OM: outer membrane;EP: exopolymers; Cyt: cyto-chrome; RC: rusticyanin;MeS: metal sulfide
4 Principles of Microbial Metal Leaching 197
ing with autoclaved spent medium where ex-creted enzymes had been inactivated. It isknown that several components involved inthe electron transport chain of Thiobacillus(rusticyanin, cytochromes, iron–sulfur pro-teins) are located in the periplasmic space(BLAKE and SHUTE, 1994; SAND et al., 1995)and might, therefore, also be present in the cellfree spent medium catalyzing oxidation of re-duced metal compounds.
In many leaching environments conditions(especially iron(II) and iron(III) concentra-tions) vary with the duration of the leaching.This makes it difficult to assess the importanceand the effect of the presence of bacteria. Us-ing an experimental setup to maintain con-stant concentrations of ferrous and ferric iron,it was possible to show that in the presence ofT. ferrooxidans rates of pyrite or zinc sulfideleaching are increased (HOLMES et al., 1999;FOWLER and CRUNDWELL, 1999; FOWLER et al.,1999).
4.3 Factors Influencing Bioleaching
Standard test methods have been developedto determine leaching rates of iron from pyrite mediated by Thiobacillus ferrooxidans(ASTM, 1991). An active culture of T. ferro-oxidans is grown in a defined medium contain-
ing (in g LP1): (NH4)2SO4 (3.0); K2HPO4 (0.5);MgSO4 ·7H2O (0.5); KCl (1.0); Ca(NO3)2 ·4H2O (0.01); FeSO4 · 7H2O (44.22); and 1 mL10 N sulfuric acid (SILVERMAN and LUNDGREN,1959). Cells are harvested, diluted, and addedto pyrite suspensions with a pulp density of 20 g LP1. Total soluble iron as well as sulfateformed during oxidation is periodically deter-mined.
Metal bioleaching in acidic environments is influenced by a series of different factors(Tab. 1). Physicochemical as well as microbio-logical factors of the leaching environment areaffecting rates and efficiencies. In addition,properties of the solids to be leached are ofmajor importance (ACEVEDO and GENTINA,1989; BRIERLEY, 1978; DAS et al., 1999; MURR,1980). As examples, pulp density, pH, and par-ticle size were identified as major factors forpyrite bioleaching by Sulfolobus acidocalda-rius (LINDSTROM et al., 1993). Optimal condi-tions were 60 g LP1, 1.5, and ~20 µm, respec-tively. The influence of different parameterssuch as activities of the bacteria itself, sourceenergy, mineralogical composition, pulp den-sity, temperature, and particle size was studiedfor the oxidation of sphalerite by T. ferrooxi-dans (BALLESTER et al., 1989). Best zinc dis-solution was obtained at low pulp densities (50 g LP1), small particle sizes, and tempera-tures of approximately 35 °C.
Fig. 3. Solubilized metals from fly ashoriginating from municipal waste incinera-tion (in suspensions of 40 g LP1) in percentof the metal amount present with differentlixiviants within 8 d. All samples wereincubated in triplicate. The release of me-tals due to acidification of the fly ash pulpis indicated as chemical mobilization (seetext for explanation).

Metal oxidation mediated by acidophilic mi-croorganisms can be inhibited by a variety offactors such as, e.g., organic compounds, sur-face-active agents, solvents, or specific metals:The presence of organic compounds (yeast ex-tract) inhibited pyrite oxidation of T. ferrooxi-dans (BACELAR-NICOLAU and JOHNSON, 1999).Certain metals present in bioleaching environ-ments can inhibit microbial growth, thereforereducing leaching efficiencies. For instance, ar-senic added to cultures inhibited Sulfolobusacidocaldarius grown on pyrite and T. ferro-oxidans grown on arsenopyrite (HALLBERG etal., 1996; LAN et al., 1994).Additions of copper,nickel, uranium, or thorium adversely influ-
enced iron(II) oxidation by T. ferrooxidanswith uranium and thorium showing higher tox-icities than copper and nickel (LEDUC et al.,1997). Silver, mercury, ruthenium, and molyb-denum reduced the growth of Sulfolobusgrown on a copper concentrate (MIER et al.,1996). Industrial biocides such as tetra-n-bu-tyltin, isothiazolinones, N-dimethyl-Nb-phe-nyl-Nb-(fluorodichloro-methylthio)-sulfamide,or 2,2b-dihydroxy-5,5b-dichlorophenylmethane(dichlorophen) reduced the leaching of man-ganese oxides by heterotrophic microorgan-isms (ARIEF and MADGWICK, 1992). Biocideswere externally added as selective inhibitors tosuppress unwanted organisms and to improve
198 8 Microbial Leaching of Metals
Tab. 1. Factors and Parameters Influencing Bacterial Mineral Oxidation and Metal Mobilization
Factor Parameter
Physicochemical parameters of a bioleaching environment temperaturepHredox potentialwater potentialoxygen content and availabilitycarbon dioxide contentmass transfernutrient availabilityiron(III) concentrationlightpressuresurface tensionpresence of inhibitors
Microbiological parameters of a bioleaching environment microbial diversitypopulation densitymicrobial activitiesspatial distribution of microorganismsmetal toleranceadaptation abilities of microorganisms
Properties of the minerals to be leached mineral typemineral compositionmineral disseminationgrain sizesurface areaporosityhydrophobicitygalvanic interactionsformation of secondary minerals
Processing leaching mode (in situ, heap, dump, or tankleaching)pulp densitystirring rate (in case of tank leaching opera-tions)heap geometry (in case of heap leaching)

4 Principles of Microbial Metal Leaching 199
manganese leaching efficiencies. At low con-centrations of ~5 mg LP1, however, mangan-ese mobilization was increased by 20% (BOUS-SIOS and MADGWICK, 1994).
Also gaseous compounds can show inhibito-ry effects on metal leaching: Aqueous-phasecarbon dioxide at concentration `10 mg LP1
was inhibiting growth of T. ferrooxidans on pyrite–arsenopyrite–pyrrothite ore (NAGPAL
et al., 1993). Optimal concentrations of carbondioxide were found to be in the range of 3 to 7 mg LP1.There are reports on the stimulationof bacterial leaching and the increase of leach-ing rates by supplementing leaching fluids withcarbon dioxide (ACEVEDO et al., 1998; BRIER-LEY, 1978; TORMA et al., 1972). Concentrationsof 4% (v/v) carbon dioxide in the inlet gas of afermenter showed maximum growth rates ofT. ferrooxidans, maximum iron(II), copper,and arsenic oxidation (ACEVEDO et al., 1998).
Pulp densities of 20 g LP1 delayed the onsetof bioleaching of pyrite derived from coal(BALDI et al., 1992). Increasing pulp densitiesfrom 30 to 100 g LP1 decreased rates of pyriteoxidation in Sulfolobus cultures (NGUBANE
and BAECKER, 1990). For fungi such as Asper-gillus niger, optimal pulp densities for maxi-mum metal leaching efficiencies were found tobe in the range of 30 to 40 g LP1 (BOSSHARD
et al., 1996). Quartz particles at pulp densitiesof 80 g LP1 almost completely inhibited theoxidation of covellite by T. ferrooxidans espe-cially in the absence of iron(II) (CURUTCHET
et al., 1990).During bioleaching processes, coprecipita-
tion of metals with mineral phases such as ja-rosites can reduce leaching efficiencies (HI-ROYOSHI et al., 1999). In addition, the precipita-tion of compounds present in the leachates onthe minerals to be leached can make the solidmaterial inaccessible for bacterial leaching.
Organic solvents such as flotation or solventextraction agents, which are added for thedownstream processing of leachates from bio-leaching, might also lead to inhibition prob-lems (ACEVEDO and GENTINA, 1989). Isopro-pylxanthate and LIX 984 (used as flotationagent and solvent extraction agent, respective-ly) prevented the oxidation of pyrite and chal-copyrite by T. ferrooxidans (HUERTA et al.,1995). This fact is of special importance whenspent leaching liquors are recycled for a reuse.
It has been demonstrated recently that theaddition of small amounts of amino acids (cys-teine in this case) resulted in an increased pyrite corrosion by T. ferrooxidans as com-pared to controls without additions (ROJAS-CHAPANA and TRIBUTSCH, 2000). It is suggest-ed that the microorganisms may profit fromweakening and break up of chemical bondsmediated by the formation of the cysteine–py-rite complex.This might also be the case undernatural conditions by the excretion of cys-teine-containing metabolites. An inexpensivealternative to increase metal recovery fromore heaps by the addition of sulfur-containingamino acids such as cysteine has been suggest-ed (TRIBUTSCH and ROJAS-CHAPANA, 1999).
Other metabolites excreted by Thiobacillusmight also enhance metal leaching efficiencies:Wetting agents such as mixtures of phospho-lipids and neutral lipids are formed by Thioba-cillus thiooxidans (BEEBE and UMBREIT, 1971).As a consequence, growth of T. thiooxidans onsulfur particles is supported by the excretionof metabolites acting as biosurfactants whichfacilitate the oxidation of elemental sulfur. Itwas also hypothesized that Thiobacillus caldusis stimulating the growth of heterotrophic or-ganisms in leaching environments by the ex-cretion of organic compounds and is support-ing the solubilization of solid sulfur by the for-mation of surface-active agents (DOPSON andLINDSTROM, 1999). Metal solubilization mightalso be facilitated by microbial metabolites ex-creted by organisms other than Thiobacilluswhich are part of microbial consortia found inbioleaching operations. Microbial surfactants,which show large differences in their chemicalnature, are formed by a wide variety of micro-organisms. In the presence of biosurfactantswhich lead to changes in the surface tension,metal desorption from solids might be en-hanced resulting in an increased metal mobil-ity in porous media. It has been suggested thatthis metabolic potential can be practically usedin the bioremediation of metal-contaminatedsoils (MILLER, 1995). However, there is someevidence that surface-active compounds aswell as organic solvents are inhibitory to bio-leaching reactions and prevent bacterial at-tachment (MURR, 1980). The external additionof Tween reduced the oxidation of chalcopyr-ite by T. ferrooxidans (TORMA et al., 1976). It

was concluded that the need of the microor-ganisms for surfactants is met by their own for-mation. In contrast, it was reported that the ad-dition of Tween 80 increased the attachment ofT. ferrooxidans on molybdenite and the oxida-tion of molybdenum in the absence of iron(II)(PISTACCIO et al., 1994).
4.4 Bacterial Attachment on Mineral Surfaces
It is known that the formation of extracellu-lar polymeric substances plays an importantrole in the attachment of thiobacilli to mineralsurfaces such as, e.g., sulfur, pyrite, or covellite.Extraction or loss of these exopolymers pre-vent cell attachment resulting in decreasedmetal leaching efficiencies (ESCOBAR et al.,1997; GEHRKE et al., 1998; POGLIANI and DO-NATI, 1992). It was concluded that a direct con-tact between bacterial cells and solid surfacesis needed and represents an important prere-quisite for an effective metal mobilization(OSTROWSKI and SKLODOWSKA, 1993). Interac-tions between microorganisms and the miner-al surface occur on two levels (BARRETT et al.,1993). The first level is a physical sorption be-cause of electrostatic forces. Due to the lowpH usually occurring in leaching environ-ments, microbial cell envelopes are positivelycharged leading to electrostatic interactionswith the mineral phase. The second level ischaracterized by chemical sorption wherechemical bonds between cells and mineralsmight be established (e.g., disulfide bridges).In addition, extracellular metabolites areformed and excreted during this phase in thenear vicinity of the attachment site (EWART
and HUGHES, 1991). Low-molecular weightmetabolites excreted by sulfur oxidizers in-clude acids originating from the TCA cycle,amino acids, or ethanolamine, whereas com-pounds with relatively high molecular weightsinclude lipids and phospholipids (BARRETT etal., 1993). In the presence of elemental sulfur,sulfur-oxidizing microorganisms from sewagesludge form a filamentous matrix similar to abacterial glycocalyx suggesting the relative im-portance of these extracellular substances inthe colonization of solid particles (BLAIS et al.,1994).
5 Microbial Diversity inBioleaching Environments
A variety of microorganisms is found inleaching environments and has been isolatedfrom leachates and acidic mine drainage. Al-though environmental conditions are usuallydescribed (from an anthropocentric view!) asbeing extreme and harsh due to pH values (aslow as P3.6; NORDSTROM et al., 2000) and highmetal concentrations (as high as 200 g LP1;NORDSTROM et al., 2000), these systems canshow high levels of microbial biodiversity in-cluding bacteria, fungi, and algae (LOPEZ-AR-CHILLA et al., 1993). It has long been knownthat bacteria (Thiobacillus sp.), yeasts (Rho-dotorula sp., Trichosporon sp.), flagellates (Eu-trepia sp.), amoebes and protozoa are part ofthe microbial biocenosis found in acidic watersof a copper mine (EHRLICH, 1963). Recent de-tailed investigations based on molecular meth-ods such as DNA–DNA hybridization, 16SrRNA sequencing, RCR-based methods withprimers derived from rRNA sequencing, fluor-escence in situ hybridization (FISH), or im-munological techniques revealed that micro-bial bioleaching communities are composed ofa vast variety of microorganisms resulting incomplex microbial interactions and nutrientflows (such as synergism, mutualism, competi-tion, predation) (AMARO et al., 1992; DE
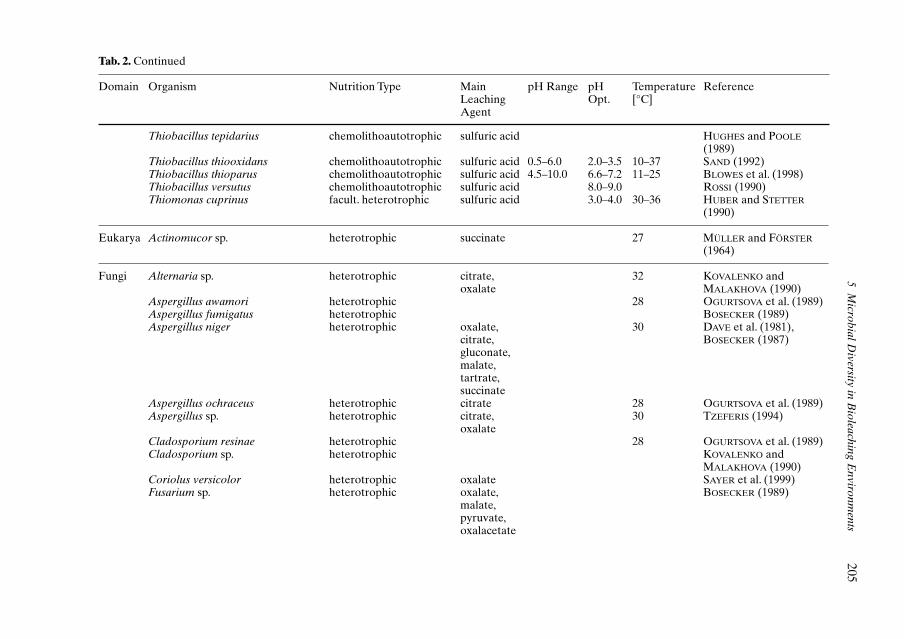
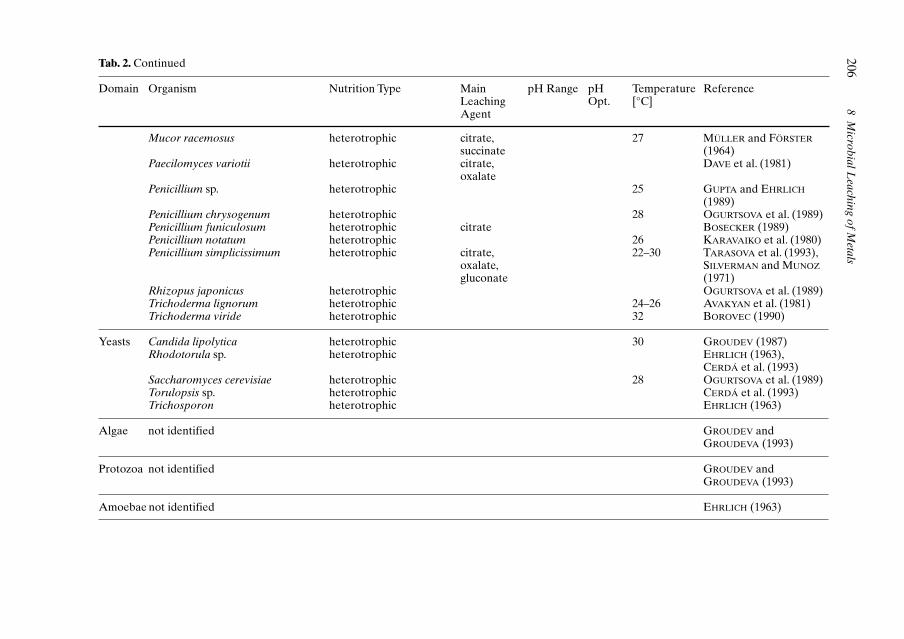
WULF-DURAND et al., 1997; EHRLICH, 1997;JOHNSON, 1998; EDWARDS et al., 1999). Selectedorganisms of these communities are given inTable 2.The composition of these communitiesis usually subjected to seasonal fluctuationsand may vary between different mining loca-tions (EDWARDS et al., 1999; GROUDEV andGROUDEVA, 1993). In addition, organisms arenot homogeneously distributed over the wholeleaching environment (CERDÁ et al., 1993).
The organism studied most is Thiobacillusferrooxidans. Although this is the best knownorganism from acidic habitats, one may notconclude that this organism is dominant inthese ecosystems. It has been found that underspecific environmental conditions Leptospiril-lum sp. is even more abundant than T. ferro-oxidans suggesting an important ecologicalrole in the microbial community structure of
200 8 Microbial Leaching of Metals

5 Microbial Diversity in Bioleaching Environments 201
bioleaching habitats (SAND, 1992; SCHRENK etal., 1998). Thiobacilli are members of the divi-sion of Proteobacteria close to the junctionbetween the � and � subdivision whereas leptospirilli are placed in the Nitrospira divi-sion (RAWLINGS, 1999). Genetic studies re-vealed that the role of T. ferrooxidans in leach-ing operations has probably been overestimat-ed. Excellent reviews on the genetics of Thio-bacilli and leptospirilli have been publishedrecently (RAWLINGS, 1999; RAWLINGS and KU-SANO, 1994).
Thiobacillus ferrooxidans belongs to thegroup of chemolithotrophic organisms.The or-ganism is rod-shaped (usually single or inpairs), non-spore forming, gram-negative, mo-tile, and single-pole flagellated (HORAN, 1999;KELLY and HARRISON, 1984; LEDUC and FER-RONI, 1994; MURR, 1980). As carbon source,carbon dioxide is utilized. Ferrous iron is oxi-dized. Ammonium is used as nitrogen source.Although T. ferrooxidans has been character-ized as being a strictly aerobic organism, it canalso grow on elemental sulfur or metal sulfidesunder anoxic conditions using ferric iron aselectron acceptor (DONATI et al., 1997; PRONK
et al., 1992).The genus Thiobacillus represents a versa-
tile group of chemolithoautotrophic organ-isms. Optimum pH values for growth varybetween 2 and 8 (Fig. 4). It has been demon-strated that sulfur-oxidizing bacteria are ca-pable of reducing the pH of highly alkaline flyash suspensions amended with elemental sul-fur from approximately 9 to 0.5 (KREBS et al.,1999) (Fig. 5). It is likely that thiobacilli con-tribute to increasing acidification of leachingecosystems in a successive mode: In the initialstages the growth of less acidophilic strains(e.g., Thiobacillus thioparus) is stimulatedwhereas during prolonged leaching the pH de-creases gradually supporting growth of moreacidophilic strains. This has already been ob-served in metal leaching from wastewater sew-age sludge (BLAIS et al., 1993).
A variety of thermophilic microorganisms(especially Sulfolobus species) has been en-riched and isolated from bioleaching environ-ments (BRIERLEY, 1990; NEMATI et al., 2000;NORRIS and OWEN, 1993).Temperature optimafor growth and metal leaching were in therange between 65 and 85°C. Although copper
extraction from mine tailings is more efficientusing thermophilic instead of mesophilic or-ganisms, extremely thermophilic microorgan-isms show a higher sensitivity to copper and tohigh pulp densities in agitated systems limit-ing, therefore, some practical applications(DUARTE et al., 1993; NORRIS and OWEN,1993).
Although environmental conditions inleaching operations favor the growth and de-velopment of mesophilic, moderately thermo-philic, and extremely thermophilic microbialcommunities, metal leaching at low tempera-tures has also been observed. Copper andnickel were leached from pyritic ore samplesin significant amounts at 4 °C (AHONEN andTUOVINEN, 1992). However, leaching rateswere lower by a factor of 30 to 50 as comparedto experiments conducted at 37°C. T. ferrooxi-dans recovered from mine waters was able togrow at 2 °C with a generation time of approx-imately 250 h suggesting a psychrotrophic na-ture of the organism (FERRONI et al., 1986).Bacterial iron mobilization has also been ob-served at 0 °C in ore samples obtained fromGreenland (LANGDAHL and INGVORSEN, 1997).Solubilization rates at these low temperatureswere still approximately 25 to 30% of the max-imum values observed at 21 °C. All these find-ings may have a potential for practical applica-tions in geographical areas where field opera-tions are subjected to low temperature regimes.
A series of heterotrophic microorganisms(bacteria, fungi) is also part of microbial bio-leaching communities (Tab. 2). This group oforganisms uses extracellular metabolites andcell lysates from autotrophs as carbon sourceresulting in the removal of an inhibitory excessof carbon and stimulating, therefore, growthand iron oxidation of thiobacilli (BUTLER andKEMPTON, 1987; FOURNIER et al., 1998). In ad-dition, several heterotrophs can also con-tribute to metal solubilization by the excretionof organic acids such as citrate, gluconate, oxa-late, or succinate.
2028
Microbial L
eaching of Metals
Tab. 2. Microbial Diversity of Acidic Bioleaching Environments and Acidic Mine Drainage: Selection of Microorganisms known to Mediate Metal Bio-leaching Reactions from Ores and Minerals or known to be Part of the Microbial Consortia Found in Bioleaching Habitats
Domain Organism Nutrition Type Main pH Range pH Temperature ReferenceLeaching Opt. [°C]Agent
Archaea Acidianus ambivalens facult. heterotrophic sulfuric acid JOHNSON (1998)Acidianus brierleyi facult. heterotrophic sulfuric acid acidophilic 1.5–3.0 45–75 MUÑOZ et al. (1995)Acidianus infernus facult. heterotrophic sulfuric acid JOHNSON (1998)Ferroplasma acidiphilum chemolithoautotrophic ferric iron 1.3–2.2 1.7 15– 45 GOLYSHINA et al. (2000)Metallosphaera prunae chemolithoautotrophic ferric iron, JOHNSON (1998)
sulfuric acidMetallosphaera sedula chemolithoautotrophic ferric iron, acidophilic extr. JOHNSON (1998)
sulfuric acid thermophilicPicrophilus oshimae JOHNSON (1998)Picrophilus torridus JOHNSON (1998)Sulfolobus acidocaldarius chemolithoautotrophic ferric iron, 0.9–5.8 2.0–3.0 55–85 AMARO et al. (1992)
sulfuric acidSulfolobus ambivalens chemolithoautotrophic ferric iron, extr. ROSSI (1990)
sulfuric acid thermophilicSulfolobus brierleyi chemolithoautotrophic ferric iron, extr. BRIERLEY (1977)
sulfuric acid thermophilicSulfolobus hakonensis chemolithoautotrophic ROSSI (1990)Sulfolobus metallicus chemolithoautotrophic ROSSI (1990)Sulfolobus solfataricus chemolithoautotrophic ferric iron, extr. JOHNSON (1998)
sulfuric acid thermophilicSulfolobus thermosulfidooxidans chemolithoautotrophic ferric iron, extr. JOHNSON (1998)
sulfuric acid thermophilicSulfolobus yellowstonii chemolithoautotrophic ferric iron, extr. JOHNSON (1998)
sulfuric acid thermophilicSulfurococcus mirabilis mixotrophic ferric iron, acidophilic extr. BARRETT et al. (1993),
sulfuric acid thermophilic JOHNSON (1998)Sulfurococcus yellowstonii mixotrophic ferric iron, JOHNSON (1998)
sulfuric acidThermoplasma acidophilum JOHNSON (1998)Thermoplasma volcanicum JOHNSON (1998)
Bacteria Acetobacter methanolicus heterotrophic gluconate acidophilic GLOMBITZA et al. (1988)Acidimicrobium ferrooxidans JOHNSON (1998),
EDWARDS et al. (1999)
5M
icrobial Diversity in B
ioleaching Environm
ents203
Acidiphilium angustum EDWARDS et al. (1999)Acidiphilium cryptum heterotrophic organic acids 2.0–6.0 mesophilic GOEBEL and
STACKEBRANDT (1994)Acidiphilium symbioticum heterotrophic organic acids 3.0 mesophilic BHATTACHARYYA et al.
(1991)Acidobacterium capsulatum chemoorganotrophic 3.0–6.0 mesophilic KISHIMOTO et al. (1991)Acidocella sp. JOHNSON (1998)Acidomonas methanolica heterotrophic JOHNSON (1998)Arthrobacter sp. heterotrophic BOSECKER (1993)Aureobacterium liquifaciens heterotrophic EDWARDS et al. (1999)Bacillus sp. heterotrophic CERDÁ et al. (1993),
GROUDEV and GROUDEVA (1993)
Bacillus coagulans heterotrophic 5.4–6.0 22 BAGLIN et al. (1992)Bacillus licheniformis heterotrophic 37 MOHANTY and MISHRA
(1993)Bacillus megaterium heterotrophic citrate KREBS et al. (1997)Bacillus polymyxa heterotrophicChromobacterium violaceum heterotrophic cyanide LAWSON et al. (1999)Comamonas testosteroni heterotrophic EDWARDS et al. (1999)Crenothrix sp. facult. autotrophic ferric iron 5.5–6.2 18–24 ROSSI (1990)Enterobacter agglomerans heterotrophic 5.4–6.0 22 BAGLIN et al. (1992)Enterobacter cloacae heterotrophic 22 BAGLIN et al. (1992)Gallionella sp. autotrophic ferric iron 6.4–6.8 6–25 ROSSI (1990)Kingella kingae EDWARDS et al. (1999)Lactobacillus acidophilus heterotrophic 37 ACHARYA et al. (1998)Leptospirillum ferrooxidans chemolithoautotrophic ferric iron 2.5–3.0 30 SAND (1992),
RAWLINGS et al. (1999)Leptospirillum thermoferrooxidans chemolithoautotrophic ferric iron 1.7–1.9 45–50 BARRETT et al. (1993)Leptothrix discophora facult. autotrophic ferric iron, 5.8–7.8 5–40 EDWARDS et al. (1999)
sulfuric acidMetallogenium sp. heterotrophic ferric iron 3.5–6.8 4.1 ROSSI (1990)Ochrobacterium anthropi heterotrophic EDWARDS et al. (1999)Propionibacterium acnes heterotrophic 37 ACHARYA et al. (1998)Pseudomonas cepacia heterotrophic 5.4–6.0 22 BAGLIN et al. (1992)
Tab. 2. Continued
Domain Organism Nutrition Type Main pH Range pH Temperature ReferenceLeaching Opt. [°C]Agent
2048
Microbial L
eaching of Metals
Pseudomonas putida heterotrophic citrate, KREBS et al. (1997)gluconate
Psychrobacter glacincola heterotrophic EDWARDS et al. (1999)Serratia ficaria heterotrophic EDWARDS et al. (1999)Siderocapsa sp. heterotrophic ferric iron ROSSI (1990)Staphylococcus lactis heterotrophic 37 ACHARYA et al. (1998)Stenotrophomonas maltophila heterotrophic EDWARDS et al. (1999)Sulfobacillus thermosulfidooxidans chemolithoautotrophic ferric iron, extr. 50 JOHNSON (1998)
sulfuric acid acidoph.Thermothrix thiopara chemolithoautotrophic sulfuric acid neutral 60–75 BRIERLEY (1977)Thiobacillus acidophilus mixotrophic sulfuric acid 1.5–6.0 3.0 25–30 CERDÁ et al. (1993),
JOHNSON (1998)Thiobacillus albertis chemolithoautotrophic sulfuric acid 2.0–4.5 3.5–4.0 28–30 JOHNSON (1998)Thiobacillus caldus chemolithoautotrophic sulfuric acid 45 AMARO et al. (1992),
DOPSON and LINDSTROM
(1999)Thiobacillus capsulatus chemolithoautotrophic sulfuric acid EWART and HUGHES
(1991)Thiobacillus concretivorus chemolithoautotrophic sulfuric acid 0.5–6.0 ROSSI (1990)Thiobacillus delicatus mixotrophic sulfuric acid 5.0–7.0 25–30 ROSSI (1990)Thiobacillus denitrificans chemolithoautotrophic sulfuric acid 5.0–7.0 30 GROUDEV and
GROUDEVA (1993)Thiobacillus ferrooxidans chemolithoautotrophic ferric iron, 1.4–6.0 2.4 28–35 SAND (1992)
sulfuric acidThiobacillus intermedius facult. heterotrophic sulfuric acid 1.9–7.0 6.8 30 ROSSI (1990)Thiobacillus kabobis mixotrophic sulfuric acid 1.8–6.0 3.0 28 ROSSI (1990)Thiobacillus neapolitanus chemolithoautotrophic sulfuric acid 3.0–8.5 6.2–7.0 28 GROUDEV and
GROUDEVA (1993)Thiobacillus novellus chemolithoautotrophic sulfuric acid 5.0–9.0 7.8–9.0 30 ROSSI (1990)Thiobacillus organoparus mixotrophic sulfuric acid 1.5–5.0 2.5–3.0 27–30 ROSSI (1990)Thiobacillus perometabolis chemolithoheterotrophic sulfuric acid 2.6–6.8 6.9 30 ROSSI (1990)Thiobacillus prosperus chemolithoautotrophic sulfuric acid 1.0–4.5 23–41 HUBER and STETTER
(1989)Thiobacillus pumbophilus chemolithoautotrophic sulfuric acid 4.0–6.5 27 DROBNER et al. (1992)Thiobacillus rubellus chemolithoautotrophic sulfuric acid 5.0–7.0 25–30 BARRETT et al. (1993)
Tab. 2. Continued
Domain Organism Nutrition Type Main pH Range pH Temperature ReferenceLeaching Opt. [°C]Agent
5M
icrobial Diversity in B
ioleaching Environm
ents205
Thiobacillus tepidarius chemolithoautotrophic sulfuric acid HUGHES and POOLE
(1989)Thiobacillus thiooxidans chemolithoautotrophic sulfuric acid 0.5–6.0 2.0–3.5 10–37 SAND (1992)Thiobacillus thioparus chemolithoautotrophic sulfuric acid 4.5–10.0 6.6–7.2 11–25 BLOWES et al. (1998)Thiobacillus versutus chemolithoautotrophic sulfuric acid 8.0–9.0 ROSSI (1990)Thiomonas cuprinus facult. heterotrophic sulfuric acid 3.0–4.0 30–36 HUBER and STETTER
(1990)
Eukarya Actinomucor sp. heterotrophic succinate 27 MÜLLER and FÖRSTER
(1964)
Fungi Alternaria sp. heterotrophic citrate, 32 KOVALENKO and oxalate MALAKHOVA (1990)
Aspergillus awamori heterotrophic 28 OGURTSOVA et al. (1989)Aspergillus fumigatus heterotrophic BOSECKER (1989)Aspergillus niger heterotrophic oxalate, 30 DAVE et al. (1981),
citrate, BOSECKER (1987) gluconate,malate,tartrate,succinate
Aspergillus ochraceus heterotrophic citrate 28 OGURTSOVA et al. (1989)Aspergillus sp. heterotrophic citrate, 30 TZEFERIS (1994)
oxalateCladosporium resinae heterotrophic 28 OGURTSOVA et al. (1989)Cladosporium sp. heterotrophic KOVALENKO and
MALAKHOVA (1990)Coriolus versicolor heterotrophic oxalate SAYER et al. (1999)Fusarium sp. heterotrophic oxalate, BOSECKER (1989)
malate,pyruvate,oxalacetate
Tab. 2. Continued
Domain Organism Nutrition Type Main pH Range pH Temperature ReferenceLeaching Opt. [°C]Agent
2068
Microbial L
eaching of Metals
Tab. 2. Continued
Domain Organism Nutrition Type Main pH Range pH Temperature ReferenceLeaching Opt. [°C]Agent
Mucor racemosus heterotrophic citrate, 27 MÜLLER and FÖRSTER
succinate (1964)Paecilomyces variotii heterotrophic citrate, DAVE et al. (1981)
oxalatePenicillium sp. heterotrophic 25 GUPTA and EHRLICH
(1989)Penicillium chrysogenum heterotrophic 28 OGURTSOVA et al. (1989)Penicillium funiculosum heterotrophic citrate BOSECKER (1989)Penicillium notatum heterotrophic 26 KARAVAIKO et al. (1980)Penicillium simplicissimum heterotrophic citrate, 22–30 TARASOVA et al. (1993),
oxalate, SILVERMAN and MUNOZ
gluconate (1971)Rhizopus japonicus heterotrophic OGURTSOVA et al. (1989)Trichoderma lignorum heterotrophic 24–26 AVAKYAN et al. (1981)Trichoderma viride heterotrophic 32 BOROVEC (1990)
Yeasts Candida lipolytica heterotrophic 30 GROUDEV (1987)Rhodotorula sp. heterotrophic EHRLICH (1963),
CERDÁ et al. (1993)Saccharomyces cerevisiae heterotrophic 28 OGURTSOVA et al. (1989)Torulopsis sp. heterotrophic CERDÁ et al. (1993)Trichosporon heterotrophic EHRLICH (1963)
Algae not identified GROUDEV and GROUDEVA (1993)
Protozoa not identified GROUDEV and GROUDEVA (1993)
Amoebae not identified EHRLICH (1963)