6 - traditional cereal fermented foods as sources of...
TRANSCRIPT

Advances in Fermented Foods and Beverages. http://dx.doi.org/10.1016/B978-1-78242-015-6.00006-2Copyright © 2015 Elsevier Ltd. All rights reserved.
Traditional cereal fermented foods as sources of functional microorganismsS.D. Todorov1, W.H. Holzapfel21Universidade de São Paulo, São Paulo, Brazil; 2Handong Global University, Pohang, South Korea
6.1 Introduction
For centuries, human civilisation has used different approaches to preserve and to pre-pare food products. Probably the longest part of this history was based on empirical knowledge, gained by experience without former knowledge either of mechanisms or the scientific basis of fermentation. However, the decisive issue was that fermentation proved to be a successful method of preservation. If we re-evaluate our experience in food biopreservation and traditional medicine, we will be surprised to discover that only since the last century have we tried to find answers to the questions about the scien-tific bases of these phenomena. If we look back in history, we can find the prepara-tion of alcoholic beverages by ancient Egyptians; the preparation of yoghurt, kumis, kefir by the nomadic peoples from central Asia; fermentation of olives by the Greeks and Romans; fermentation of meat by the Germanic tribes and fish by the Eskimos; preparation of boza by the ancient Persians; or fermenting corn (maize) by the native tribes in pre-Columbian America. All of these peoples most probably did not have any knowledge of microbiology from the perspective of nineteenth century scientists, but they were convinced by their personal experience that using specific technical means of preparing these products succeeded in preserving meat, fish, milk, fruits and vegetables.
Based on empiric experience, some of these fermented products have been used in traditional medicine; moreover, traditional medicine quite frequently made use of fermentation processes to prepare medication. Empiric knowledge, frequently trans-ferred from one generation to the next, was the only basis for preparation of these products. Our great-grandmothers knew that, when preparing yoghurt, you need to use the milk at a temperature that cannot burn yourself, and to add a portion of the yoghurt from the previous day’s preparation. Preparation of kefir, differing from yoghurt, was traditionally associated with kefir grains.
6.2 Food fermentation processes
Fermentation of food is a very old technology, and the earliest records may date back to 6000 BC (Fox, Lucey, & Cogan, 1993). These methodologies and knowledge associated
6

124 Advances in Fermented Foods and Beverages
with the manufacturing of fermented products were handed down from generation to generation within local communities (Caplice & Fitzgerald, 1999). Frequently, these communities needed products to be produced in small quantities for distribution in or around the immediate area. However, the population increase in towns and cities, due to the industrial revolution by the middle of the nineteenth century, resulted in a need for these products to be produced in larger quantities, and this led to commercial production of fermented food. Furthermore, developments in science (in general) and particularly in microbiology exponentially increased from the middle of the nineteenth century, and the development of pasteurisation by Louis Pasteur toward the end of the nineteenth century had a major impact on our understanding of the biological basis of fermentation. According to Caplice and Fitzgerald (1999), milk, meat, cucumbers and cabbage are the main substrates used in the production of the most familiar fermented food products.
Even when the scientific basic of fermentation processes and modern microbiol-ogy was established by Louis Pasteur, Ilja Metchnikov and his collaborator, Stamen Grigorov, first suggested the concept basic to modern understanding of functional food products and probiotics. At the end of the nineteenth century, it was still not well known how and why probiotics works; the term “probiotics” was not even used. Only much later, Fuller and Gibson (1997) defined this term as live microorganisms that, when administrated in adequate amounts, confer health benefits to the host such as reduction of gastrointestinal infections and inflammatory bowel disease, and modula-tion of the immune system (Fuller & Gibson, 1997).
Large-scale production required products with consistent quality. Characterisation of microorganisms responsible for the fermentation of various fermented products led to the isolation of starter cultures, which could be produced on a large scale to supply fac-tories involved in the manufacturing of these products. Defined starter cultures replaced undefined starters traditionally used in manufacturing, and ensured reliable fermentation and consistent product quality (Caplice & Fitzgerald, 1999). The digestibility, nutritional value, organoleptic qualities, and shelf-life of industrial food products are increased by controlled fermentation processes (Hancioglu & Karapinar, 1997). However, a number of lactic acid bacteria (LAB) used as starter cultures in fermented food have probiotic properties and may confer potential health benefits to the consumer.
6.2.1 Lactic acid bacteria in cereal-based fermented products
Cereal and cereal-legume–based fermented products are consumed in almost all parts of the world. Cereals are cultivated on more than 73% of agricultural soil and contrib-ute to more than 60% of the world’s food production, providing proteins, vitamins, dietary fibre, energy and minerals (Charalampopoulos, Wang, Pandiella, & Webb, 2002). It is therefore important to study the nutritional value and basic composition of their products. Many cereal-based products are heat treated (boiled or steamed), for example, porridges, rice, pasta and cookies. In many cases, the same product is before or after heat treatment subject to fermentation processes, for example, pancakes and flatbreads in Asia, sourdough bread in Europe, and a variety of fermented dump-lings, porridges, and alcoholic and nonalcoholic beers in Europe, Asia and Africa (Salovaara, 2004).

125Traditional cereal fermented foods as sources of functional microorganisms
By definition, fermentation is the process by which a substrate is subjected to biochemical modification resulting from the activity of microorganisms and their enzymes (Gotcheva, Pandiella, Angelov, Roshkova, & Webb, 2000). Yeasts, LAB, fungi, or mixtures of these are mainly responsible for natural cereal-based fermen-tation. Yeasts mainly perform carbohydrate metabolism, whereas bacteria show pro-teolytic activity (Chavan & Kadam, 1989). Both yeasts and bacteria contribute to the sensory and safety aspects of the final products. Fermentations by yeasts and lac-tobacilli change the biochemical composition of fats, minerals and vitamins in the cereal. Yeasts are predominantly responsible for the production of ethanol (e.g. beers and wines), whereas lactic acid bacteria produce mainly lactic acid (e.g. cereals and fermented milk products). Acetic acid formation, most typically by the aerobic con-version of alcohol to acetic acid in the presence of excess oxygen, is mainly conducted by Acetobacter and Gluconobacter spp. (Blandino, Al-Aseeri, Pandiella, Cantero, & Webb, 2003). Alkaline fermentation is commonly associated with the fermentation of fish and seeds, widely used as condiment (McKay & Baldwin, 1990).
6.2.2 History of boza, a cereal-based fermented beverage
The origin of boza dates back to the ancient peoples who lived in Anatolia and Mesopotamia. The preparation formula were taken by the Ottomans and distributed throughout the countries that they conquered. The Greek historian Xenophon recorded that boza was made in eastern Anatolia in 401 BC and stored in clay jars that were bur-ied beneath the ground. The local speciality remained confined to the region until the arrival of the Turks, who took this nourishing drink and spread it far and wide under the name boza, a word derived from the Persian word “buze”, meaning millet. Boza enjoyed its golden age under the Ottomans, and its preparation became one of the prin-cipal trade items in towns and cities from the early Ottoman period (Todorov, 2010).
Beer is supposed to have originated from boza, a drink, probably dating back 4500 years. Although the alcohol and acid content of boza was not known at that time, boza was described in clay tablets as a stimulant and also as a medicine. From perspective of twentyfirst century science, we have sufficient evidence to relate some of the proper-ties to the probiotic characteristics carried by LAB, part of the boza microbiota (Todorov et al., 2008). Boza consumption was initially widespread in the Islamic countries, but production was prohibited in the eighteenth century because of its high alcohol content. However, it is still produced and consumed widely in Anatolia, southern Russia, eastern European countries, the Middle East and northern Persia (Arici & Daglioglu, 2002).
Boza is a low-alcohol beverage produced by the fermentation of barley, oats, millet, maize, wheat or rice. The cooked cereal is strained to remove most of the solids; sugar is added to taste and inoculated with a starter culture, consisting of either yogurt or sourdough. The sludge is fermented at 30 °C for 24 h, cooled and kept refrigerated for 3–5 days (Hancioglu & Karapinar, 1997; Zorba, Hancioglu, Genc, Karapinar, & Ova, 2003). Boza contains about 0.50–1.61% protein, 12.3% carbohydrate and 75–85% moisture (Yegin & Uren, 2008; Zorba et al., 2003). In general, the pH of the boza sam-ples ranges from 3.16 to 4.02 (Yegin & Uren, 2008), and the average alcohol content is 0.13% (w/v) (Köse & Yücel, 2003).

126 Advances in Fermented Foods and Beverages
Boza can be classified as either sweet or sour, depending on its acid content. Besides improving organoleptic quality and digestibility by fermentation, it is a nutri-tious food because of its lactic acid, fat, protein, carbohydrate, fibre and vitamin con-tents, and thus is a valuable fermented food that beneficially contributes to human nutrition (Morcos, Hegazi, & El-Damhoughy, 1973). The chemical properties of boza during the fermentation and storage time are significantly affected by the raw material (p < 0.01) (Akpinar-Bayizit, Yilmaz-Ersan, & Ozcan, 2010). The compositional dif-ferences of boza samples may result from the use of different cereals as raw material and their amounts in the recipe (Hammes et al., 2005; Hammes & Gänzle, 1998). The raw material affects the amount and quality of carbohydrates available as primary fermentation substrates, nitrogen sources and growth factors for microbial activity.
6.2.3 Physicochemical characteristics of boza
Total titratable acidity in terms of lactic acid was found to be the lowest in millet, with 0.32 ± 0.04% and the highest in wheat boza (0.61 ± 0.07%) (Akpinar-Bayizit et al., 2010), probably due to the high fermentable carbohydrate content of wheat compared to other raw materials. The pH varied between 3.43 ± 0.08 and 3.86 ± 0.17 (Akpinar-Bayizit et al., 2010). The acidity of the samples increased during storage (being highest after 192 h, with 0.68 ± 0.06%) concomitantly with a decrease in the pH. Furthermore, the alcohol content was lower in wheat boza (0.46 ± 0.04%) (Akpinar-Bayizit et al., 2010) and showed fluctuations during storage, depending on microbial and enzymatic activ-ities. The acidity and alcohol content depended mainly on the fermentation period; it was demonstrated that, with longer fermentation periods, the acidity increases, as does the concentration of alcohol (Akpinar-Bayizit et al., 2010).
The history of boza and similar beverages dates back several thousand years. How-ever, the original boza was probably different from that which is produced nowadays, having high alcohol content (up to 7% by volume). In Egypt, a traditional beverage called bouza is still produced. In the South African region, boza production has become an important section of the beverage industry. Boza and similar beverages are produced with different recipes and methods in various countries (Table 6.1). Boza is called bousa or bouza in Nigeria and some other African countries, and it is similar to beer due to its high alcohol content (Sanni, 1993). In Bulgaria, boza is produced either plain or with cocoa, either in winter or in summer (Enikova, Kozereva, Ivanova, & Yang’ozova, 1985). In the Balkan region of Europe, this beverage is also called boza. In Turkey, boza is mostly produced and consumed in winter, although, because of the refreshing cooling effect of lactic acid, it can also be consumed in summer; however, high tem-peratures in the summer season can lead to the growth of yeasts and acetic acid bacteria (Arici & Daglioglu, 2002). Thus, sensory qualities of the product can change rapidly, causing a dramatic decrease in the shelf-life (Arici & Daglioglu, 2002).
The steps for boza production can be summarised as: (1) preparation of the raw materials, (2) boiling, (3) cooling and straining, (4) addition of sugar and (5) fer-mentation. Boza (2–3%) from a previous batch is usually used as a starter culture. The mixture is left to ferment in wooden barrels. The ratio of the starter culture depends on the season and temperature at which it is produced. The inoculated
127T
raditional cereal fermented foods as sources of functional m
icroorganisms
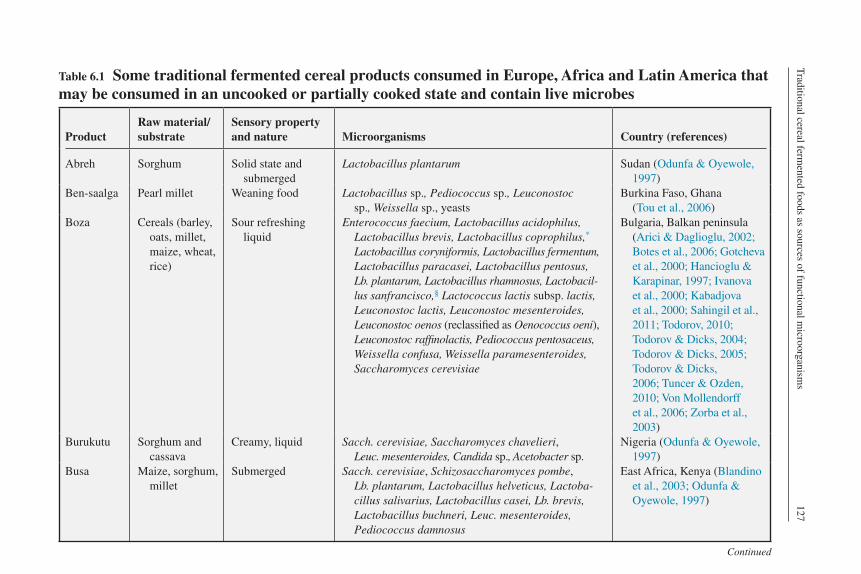
Table 6.1 Some traditional fermented cereal products consumed in Europe, Africa and Latin America that may be consumed in an uncooked or partially cooked state and contain live microbes
ProductRaw material/substrate
Sensory property and nature Microorganisms Country (references)
Abreh Sorghum Solid state and submerged
Lactobacillus plantarum Sudan (Odunfa & Oyewole, 1997)
Ben-saalga Pearl millet Weaning food Lactobacillus sp., Pediococcus sp., Leuconostoc sp., Weissella sp., yeasts
Burkina Faso, Ghana (Tou et al., 2006)
Boza Cereals (barley, oats, millet, maize, wheat, rice)
Sour refreshing liquid
Enterococcus faecium, Lactobacillus acidophilus, Lactobacillus brevis, Lactobacillus coprophilus,* Lactobacillus coryniformis, Lactobacillus fermentum, Lactobacillus paracasei, Lactobacillus pentosus, Lb. plantarum, Lactobacillus rhamnosus, Lactobacil-lus sanfrancisco,§ Lactococcus lactis subsp. lactis, Leuconostoc lactis, Leuconostoc mesenteroides, Leuconostoc oenos (reclassified as Oenococcus oeni), Leuconostoc raffinolactis, Pediococcus pentosaceus, Weissella confusa, Weissella paramesenteroides, Saccharomyces cerevisiae
Bulgaria, Balkan peninsula (Arici & Daglioglu, 2002; Botes et al., 2006; Gotcheva et al., 2000; Hancioglu & Karapinar, 1997; Ivanova et al., 2000; Kabadjova et al., 2000; Sahingil et al., 2011; Todorov, 2010; Todorov & Dicks, 2004; Todorov & Dicks, 2005; Todorov & Dicks, 2006; Tuncer & Ozden, 2010; Von Mollendorff et al., 2006; Zorba et al., 2003)
Burukutu Sorghum and cassava
Creamy, liquid Sacch. cerevisiae, Saccharomyces chavelieri, Leuc. mesenteroides, Candida sp., Acetobacter sp.
Nigeria (Odunfa & Oyewole, 1997)
Busa Maize, sorghum, millet
Submerged Sacch. cerevisiae, Schizosaccharomyces pombe, Lb. plantarum, Lactobacillus helveticus, Lactoba-cillus salivarius, Lactobacillus casei, Lb. brevis, Lactobacillus buchneri, Leuc. mesenteroides, Pediococcus damnosus
East Africa, Kenya (Blandino et al., 2003; Odunfa & Oyewole, 1997)
Continued
128A
dvances in Fermented Foods and B
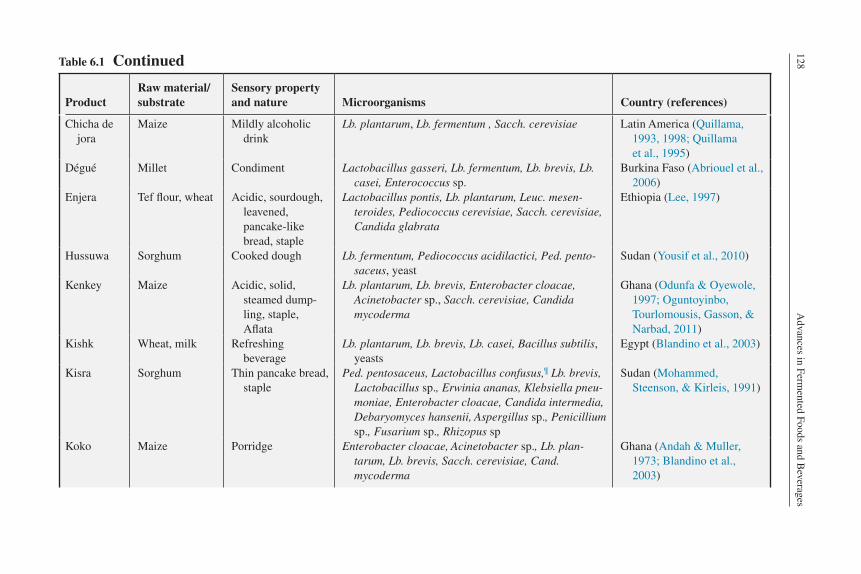
everagesTable 6.1 Continued
Chicha de jora
Maize Mildly alcoholic drink
Lb. plantarum, Lb. fermentum , Sacch. cerevisiae Latin America (Quillama, 1993, 1998; Quillama et al., 1995)
Dégué Millet Condiment Lactobacillus gasseri, Lb. fermentum, Lb. brevis, Lb. casei, Enterococcus sp.
Burkina Faso (Abriouel et al., 2006)
Enjera Tef flour, wheat Acidic, sourdough, leavened, pancake-like bread, staple
Lactobacillus pontis, Lb. plantarum, Leuc. mesen-teroides, Pediococcus cerevisiae, Sacch. cerevisiae, Candida glabrata
Ethiopia (Lee, 1997)
Hussuwa Sorghum Cooked dough Lb. fermentum, Pediococcus acidilactici, Ped. pento-saceus, yeast
Sudan (Yousif et al., 2010)
Kenkey Maize Acidic, solid, steamed dump-ling, staple, Aflata
Lb. plantarum, Lb. brevis, Enterobacter cloacae, Acinetobacter sp., Sacch. cerevisiae, Candida mycoderma
Ghana (Odunfa & Oyewole, 1997; Oguntoyinbo, Tourlomousis, Gasson, & Narbad, 2011)
Kishk Wheat, milk Refreshing beverage
Lb. plantarum, Lb. brevis, Lb. casei, Bacillus subtilis, yeasts
Egypt (Blandino et al., 2003)
Kisra Sorghum Thin pancake bread, staple
Ped. pentosaceus, Lactobacillus confusus,¶ Lb. brevis, Lactobacillus sp., Erwinia ananas, Klebsiella pneu-moniae, Enterobacter cloacae, Candida intermedia, Debaryomyces hansenii, Aspergillus sp., Penicillium sp., Fusarium sp., Rhizopus sp
Sudan (Mohammed, Steenson, & Kirleis, 1991)
Koko Maize Porridge Enterobacter cloacae, Acinetobacter sp., Lb. plan-tarum, Lb. brevis, Sacch. cerevisiae, Cand. mycoderma
Ghana (Andah & Muller, 1973; Blandino et al., 2003)
ProductRaw material/substrate
Sensory property and nature Microorganisms Country (references)
129T
raditional cereal fermented foods as sources of functional m
icroorganisms
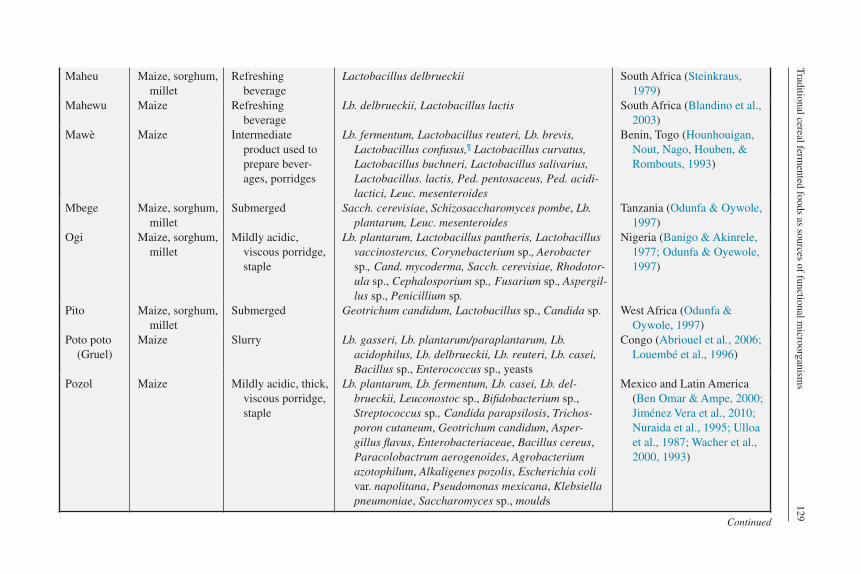
Maheu Maize, sorghum, millet
Refreshing beverage
Lactobacillus delbrueckii South Africa (Steinkraus, 1979)
Mahewu Maize Refreshing beverage
Lb. delbrueckii, Lactobacillus lactis South Africa (Blandino et al., 2003)
Mawè Maize Intermediate product used to prepare bever-ages, porridges
Lb. fermentum, Lactobacillus reuteri, Lb. brevis, Lactobacillus confusus,¶ Lactobacillus curvatus, Lactobacillus buchneri, Lactobacillus salivarius, Lactobacillus. lactis, Ped. pentosaceus, Ped. acidi-lactici, Leuc. mesenteroides
Benin, Togo (Hounhouigan, Nout, Nago, Houben, & Rombouts, 1993)
Mbege Maize, sorghum, millet
Submerged Sacch. cerevisiae, Schizosaccharomyces pombe, Lb. plantarum, Leuc. mesenteroides
Tanzania (Odunfa & Oywole, 1997)
Ogi Maize, sorghum, millet
Mildly acidic, viscous porridge, staple
Lb. plantarum, Lactobacillus pantheris, Lactobacillus vaccinostercus, Corynebacterium sp., Aerobacter sp., Cand. mycoderma, Sacch. cerevisiae, Rhodotor-ula sp., Cephalosporium sp., Fusarium sp., Aspergil-lus sp., Penicillium sp.
Nigeria (Banigo & Akinrele, 1977; Odunfa & Oyewole, 1997)
Pito Maize, sorghum, millet
Submerged Geotrichum candidum, Lactobacillus sp., Candida sp. West Africa (Odunfa & Oywole, 1997)
Poto poto (Gruel)
Maize Slurry Lb. gasseri, Lb. plantarum/paraplantarum, Lb. acidophilus, Lb. delbrueckii, Lb. reuteri, Lb. casei, Bacillus sp., Enterococcus sp., yeasts
Congo (Abriouel et al., 2006; Louembé et al., 1996)
Pozol Maize Mildly acidic, thick, viscous porridge, staple
Lb. plantarum, Lb. fermentum, Lb. casei, Lb. del-brueckii, Leuconostoc sp., Bifidobacterium sp., Streptococcus sp., Candida parapsilosis, Trichos-poron cutaneum, Geotrichum candidum, Asper-gillus flavus, Enterobacteriaceae, Bacillus cereus, Paracolobactrum aerogenoides, Agrobacterium azotophilum, Alkaligenes pozolis, Escherichia coli var. napolitana, Pseudomonas mexicana, Klebsiella pneumoniae, Saccharomyces sp., moulds
Mexico and Latin America (Ben Omar & Ampe, 2000; Jiménez Vera et al., 2010; Nuraida et al., 1995; Ulloa et al., 1987; Wacher et al., 2000, 1993)
Continued
130A
dvances in Fermented Foods and B
everages
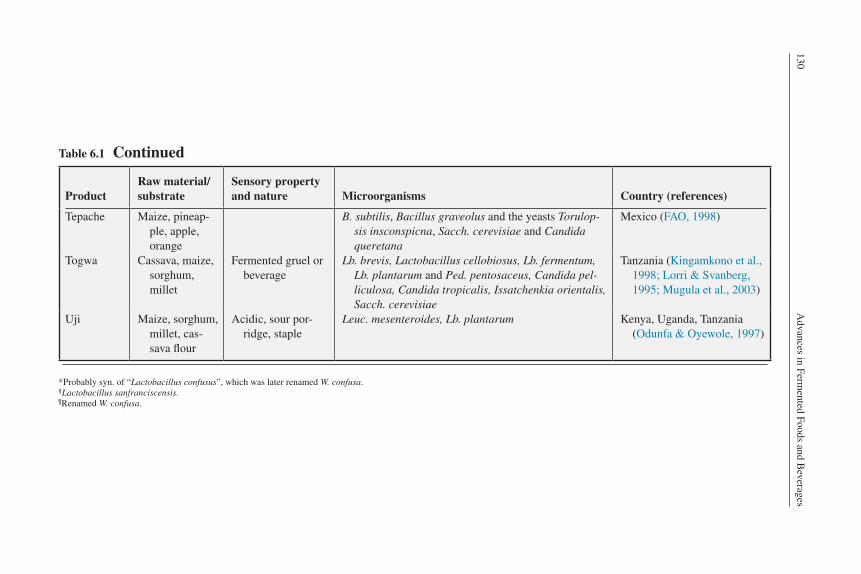
Tepache Maize, pineap-ple, apple, orange
B. subtilis, Bacillus graveolus and the yeasts Torulop-sis insconspicna, Sacch. cerevisiae and Candida queretana
Mexico (FAO, 1998)
Togwa Cassava, maize, sorghum, millet
Fermented gruel or beverage
Lb. brevis, Lactobacillus cellobiosus, Lb. fermentum, Lb. plantarum and Ped. pentosaceus, Candida pel-liculosa, Candida tropicalis, Issatchenkia orientalis, Sacch. cerevisiae
Tanzania (Kingamkono et al., 1998; Lorri & Svanberg, 1995; Mugula et al., 2003)
Uji Maize, sorghum, millet, cas-sava flour
Acidic, sour por-ridge, staple
Leuc. mesenteroides, Lb. plantarum Kenya, Uganda, Tanzania (Odunfa & Oyewole, 1997)
*Probably syn. of “Lactobacillus confusus”, which was later renamed W. confusa.§Lactobacillus sanfranciscensis.¶Renamed W. confusa.
Table 6.1 Continued
ProductRaw material/substrate
Sensory property and nature Microorganisms Country (references)

131Traditional cereal fermented foods as sources of functional microorganisms
mixture is incubated at 15–25 °C for nearly 24 h before it is ready for use (Arici & Daglioglu, 2002).
Two different types of fermentation occur simultaneously during boza fermenta-tion. The first is the alcoholic fermentation that produces carbon dioxide bubbles and increases the volume; the second, lactic acid fermentation, produces lactic acid and gives the acidic character to boza. Due to the increase in volume during fermentation, the wooden barrels should not be fully filled. After production, boza should be con-sumed within a couple of days to prevent an excessively sour taste. In practice, the fermentation rate is reduced by cold storage to extend the shelf life of boza. In the first boza production of the season, sourdough or yoghurt is used as a starter culture, since fresh boza is not available. When using sourdough, which is less viscous, a more acidic product is obtained when compared to the product that is inoculated with a previous boza batch. If yoghurt is preferred, a viscous but more acidic product is obtained, and the characteristic yoghurt taste is easily detected (Arici & Daglioglu, 2002).
6.2.4 Microbiology of boza
Countries in the Balkan region in Europe are famous for their production of food and beverages fermented with LAB. Boza is one such traditional drink; however, only a few reports have been published on the microbial composition of boza. Most of the LAB that have been isolated belong to the genera Lactobacillus, Lactococcus, Leuco-nostoc, Pediococcus, Enterococcus, Oenococcus and Weissella (Arici & Daglioglu, 2002; Botes, Todorov, von Mollendorff, Botha, & Dicks, 2006; Gotcheva et al., 2000; Hancioglu & Karapinar, 1997; Kabadjova, Gotcheva, Ivanova, & Dousset, 2000; Todorov, 2010; Todorov & Dicks, 2004, 2005, 2006; Von Mollendorff, Todorov, & Dicks, 2006; Zorba et al., 2003). Only one study (Zorba et al., 2003) addressed the selection of boza starter cultures. LAB and presumably yeasts produce a number of vitamins (Leroy & De Vuyst, 2004), and the adequate selection of starter cultures can increase the nutritional value of fermented products (LeBlanc, Taranto, Molina, & Sesma, 2010).
Several studies have reported on the isolation and identification of LAB and yeasts in boza; however, to our knowledge, only the studies of Botes et al. (2006), Todorov and Dicks (2006) and Todorov (2010) used biomolecular approaches to identify these microorganisms. In these studies, the numbers of LAB isolated from three boza sam-ples ranged from 9 × 106 to 5 × 107 CFU/ml. Carbohydrate fermentation reactions and PCR with species-specific primers classified the isolates as Lactobacillus paracasei subsp. paracasei, Lactobacillus pentosus, Lactobacillus plantarum, Lactobacillus brevis, Lactobacillus rhamnosus, Lactobacillus fermentum, Leuconostoc lactis and Enterococcus faecium.
Only a few papers have reported the isolation of yeasts and moulds from boza. Most of the yeasts were identified as strains of Candida glabrata, Candida tropica-lis, Geotrichum candidum, Geotrichum penicilatum, Saccharomyces carlsbergensis, Saccharomyces cerevisiae and Saccharomyces uvarum (Arici & Daglioglu, 2002; Gotcheva et al., 2000). The latter identifications were based on morphological, phys-iological and biochemical characteristics. Boza harbors a diverse population of LAB

132 Advances in Fermented Foods and Beverages
that include strains of Lactobacillus acidophilus, Lb. brevis, Lactobacillus “coprophi-lus” (Syn.: Weissella confusa?), Lactobacillus coryniformis, Lb. fermentum, Lb. para-casei, Lb. pentosus, Lb. plantarum, Lb. rhamnosus, Lactobacillus sanfranciscensis, Lactococcus lactis subsp. lactis, Leuconostoc mesenteroides, Leuconostoc mesen-teroides subsp. dextranicum, Leuconostoc raffinolactis, Leuc. lactis, Ent. faecium, Pediococcus pentosaceus, Leuconostoc oenos (reclassified to Oenococcus oeni), W. confusa and Weissella paramesenteroides (Arici & Daglioglu, 2002; Gotcheva et al., 2000; Todorov, 2010; Todorov & Dicks, 2006; Von Mollendorff et al., 2006).
Yeasts were isolated from two-thirds of the boza samples, with viable numbers ranging from 1.3 × 102 to 1.8 × 103 CFU/ml. Results obtained from sequencing of the D1/D2 rDNA region identified the yeasts as Candida diversa, Candida inconspicua, Candida pararugosa, Issatchenkia orientalis, Pichia fermentans, Pichia guillier-mondii, Pichia norvegensis, Rhodotorula mucilaginosa and Torulaspora delbrueckii. Saccharomyces cerevisiae, commonly associated with fermented beverages, has not been detected in any of the boza samples (Botes et al., 2006). The absence of Sacch. cerevisiae suggests that the species was either not present in the inoculum (at least not in high numbers) or that it was inhibited by the LAB and other yeasts toward the end of fermentation. Candida inconspicua has been isolated from human sputum and tongue and is an opportunistic pathogen. Rhodotorula mucilaginosa is also an opportunistic pathogen implicated in fungaemia, endocarditis and meningitis. Pichia norvegensis has been associated with septicemia in humans (Gomez-Lopez, Mellado, Rodriguez-Tudela, & Cuenca-Estrella, 2005; Maxwell et al., 2003). The presence of potential pathogens emphasises the importance of developing starter cultures with GRAS status for the commercial production of boza and the necessity of introducing novel methods to prevent pathogen propagation.
6.3 Antimicrobial proteins isolated from boza-related lactic acid bacteria
6.3.1 Definition of a bacteriocin
LAB are known for the production of antimicrobial compounds, including bacterio-cins or bacteriocin-like peptides (De Vuyst & Vandamme, 1994). Bacteriocins of LAB are defined as ribosomally synthesised proteins or protein complexes usually antag-onistic to genetically closely related organisms (De Vuyst & Vandamme, 1994), and are generally low-molecular-weight proteins that gain entry into target cells by bind-ing to cell surface receptors. Their bactericidal mechanism varies and may include pore formation, degradation of cellular DNA, disruption through specific cleavage of 16S rDNA and inhibition of peptidoglycan synthesis (De Vuyst & Vandamme, 1994; Heu et al., 2001). In recent studies, specific environmental conditions, including those found in food, have been examined to determine their effect on the production of bac-teriocins (Leroy & De Vuyst, 2004). Bacteriocin production is dramatically affected by changes in environmental conditions, and optimal production may require a specific

133Traditional cereal fermented foods as sources of functional microorganisms
combination of parameters (Leal-Sánchez, Jiménez-Díaz, Maldonado-Barragán, Garrido-Fernández, & Ruiz-Barba, 2002). Little is known about the interactions of these factors with the production of bacteriocins, especially in a complex food environment.
6.3.2 Boza-associated bacteriocin producers
Kabadjova et al. (2000) and Ivanova, Kabadjova, Pantev, Danova, and Dousset (2000) first reported the isolated of bacteriocin-producing strains from boza (produced in Sofia, Bulgaria). Of the 80 isolated strains of LAB, a group of 33 showed antibacterial activity against different test microorganisms (Listeria innocua F, Lb. plantarum 73, Lactococcus cremoris 117, and even against Gram-negative bacteria such as Esche-richia coli). The strain defined as Lact. lactis subsp. lactis 14 (based on biochemical identification tests) was chosen for future tests, and its growth curve and ability to produce bacteriocin under different conditions of cultivation have been studied. An attempt has been made for initial purification of the bacteriocin by means of a classical method (Kabadjova et al., 2000).
Todorov and Dicks (2004) reported on the bacteriocin (mesentericin ST99) pro-duced by Leuc. mesenteroides subsp. dextranicum ST99 isolated from boza originat-ing from Belogratchik (northwest of Bulgaria). The cell-free supernatant of this strain inhibited the growth of Bacillus subtilis, Enterococcus faecalis, several Lactobacillus spp., Lactococcus lactis subsp. cremoris, Li. innocua, Listeria monocytogenes, Ped. pentosaceus, Staphylococcus aureus and Streptococcus thermophilus. However, Clos-tridium spp., Carnobacterium spp., Leuc. mesenteroides and Gram-negative bacteria were not inhibited (Todorov & Dicks, 2004). Maximum antimicrobial activity, that is, 6400 AU/ml, was recorded in MRS broth after 24 h growth at 30 °C. Metabolically active cells of Li. innocua treated with mesentericin ST99 did not undergo lysis, and mesentericin ST99 did not adhere to the cell surface of strain ST99. Mesentericin ST99 were partially purified by precipitation with ammonium sulfate (70% saturation), fol-lowed by Sep-Pack C18 chromatography and reverse-phase high-performance liquid chromatography (HPLC) on a C18 Nucleosil column, and yielded one antimicrobial peptide (Todorov & Dicks, 2004).
In 2005, the same authors (Todorov & Dicks, 2005) reported that 13 of the 52 strains isolated from boza produced in Belogratchik, northwest of Bulgaria, inhibited the growth of Pediococcus spp., Li. innocua and Lb. plantarum. The population of LAB recorded in boza was c. 2 × 108 CFU/ml. One of the strains, identified as Ped. pentosaceus ST18 (based on biochemical identification tests), produced pediocin ST18 at 3200 AU/ml in MRS broth at the end of logarithmic growth (i.e. after 24 h). Pediocin ST18 was active against all tested strains of Pediococcus spp. included in this study, and revealed an important antilisterial activity, including that against Li. monocytogenes. From the 54 bacterial strains tested, 29 were sensitive to pediocin ST18. No activity was recorded against Gram-negative bacteria in this study.
Pediocin ST18 have been partially purified by ammonium sulphate precipitation, followed by separation in a Sep-Pack C18 column and reverse-phase HPLC on a C18 Nucleosil column, and yielded in two active antimicrobial peptides, which suggests that pediocin ST18 may be a two-peptide bacteriocin. The peptide had bacteriostatic

134 Advances in Fermented Foods and Beverages
action toward Li. innocua but did not cause cellular lyses. Pediocin ST18 does not adhere to the cell surface of the producer strain (Todorov & Dicks, 2005).
The study of Todorov and Dicks (2006) is the first report on the isolation of LAB from boza and their identification based on genetic approaches. Boza samples were obtained from the same town in Bulgaria previously reported by Todorov and Dicks (2004, 2005). The population of LAB recorded in Boza was approximately 5 × 107 CFU/ml. Lactobacillus plantarum (strains ST194BZ, ST414BZ and ST664BZ), Lb. pento-sus (strain ST712BZ), Lb. rhamnosus (strains ST461BZ and ST462BZ) and Lb. para-casei (strains ST242BZ and ST284BZ), isolated from boza, produced bacteriocins active against Lactobacillus casei, E. coli, Pseudomonas aeruginosa and Ent. faecalis. Thus far, only a few bacteriocins with activity against Gram-negative bacteria have been reported, but, for the second time, a bacteriocin isolated from LAB originated from boza was reported (Todorov & Dicks, 2006). All eight bacteriocins inhibited Lb. casei, E. coli, Ps. aeruginosa and Ent. faecalis. Bacteriocin ST242BZ inhibited the growth of Acinetobacter baumanii and bacteriocins ST194BZ, ST242BZ, ST284BZ and ST414BZ were active against Enterobacter cloacae (Todorov & Dicks, 2006). All bacteriocins acted bactericidal. The bacteriocins did not adhere to the surface of the producer cells. Production occurred throughout logarithmic growth, with the high-est activity recorded at the end of logarithmic and during stationary growth. Based on tricine SDS-PAGE, the bacteriocins ranged from 2.8 to 14.0 kDa. No plasmids were recorded, suggesting that the genes encoding the bacteriocins are located on the genomes (Todorov & Dicks, 2006).
Von Mollendorff et al. (2006) reported on bacteriocins produced by LAB isolated from boza. Four isolates (JW3BZ, JW6BZ, JW11BZ and JW15BZ) produced bacte-riocins active against a broad spectrum of Gram-positive bacteria. In addition, bac-teriocin JW15BZ was shown to inhibit the growth of Klebsiella pneumoniae. The producer strains were identified as Lb. plantarum (strains JW3BZ and JW6BZ) and Lb. fermentum (strains JW11BZ and JW15BZ) based on biochemical and biomolecu-lar approaches and 16S rDNA sequencing. The spectrum of antimicrobial activity, bio-chemical characteristics, and mode of action of these bacteriocins were compared with bacteriocins previously described for LAB isolated from boza (Von Mollendorff et al., 2006). The highest level of bacteriocin JW3BZ activity (25,600 AU/ml) was recorded after 18 h of growth in MRS broth (30 °C), and it remained at this level for the dura-tion of fermentation. The same level of bacteriocin JW6BZ activity (25,600 AU/ml) was recorded after 15 h of growth, which was followed by a decrease to 12,800 AU/ml during the next 6 h. Bacteriocins JW11BZ and JW15BZ were produced at lower levels (12,800 AU/ml) and only after 15 and 12 h, respectively. Bacteriocin JW11BZ produc-tion decreased to 6400 AU/ml after 18 h of fermentation, and bacteriocin JW15BZ to 3200 AU/ml after 21 h of fermentation (Von Mollendorff et al., 2006). The molecular size of bacteriocins JW3BZ, JW6BZ, JW11BZ and JW15BZ ranged from approxi-mately 2.3 to 3.0 kDa.
Lactococcus lactis subsp. lactis YBD11 was isolated from boza, produced in Turkey (Tuncer & Ozden, 2010). The bacteriocin produced by Lact. lactis subsp. lactis YBD11 inhibited the growth of Lb. plantarum, Lactobacillus sakei, Lact. lactis subsp. lactis, Lact. lactis subsp. cremoris, Micrococcus luteus, Li. innocua, Enterococcus

135Traditional cereal fermented foods as sources of functional microorganisms
feacalis, Staphylococcus aureus, Staphylococcus carnosus, Ped. pentosaceus and Bacillus cereus; however, the nisin-producing strain Lactococcus lactis SIK83 and Gram-negative bacteria were not inhibited. Maximum antimicrobial activity, 10,240 AU/ml, was recorded in glucose-M17 broth after 18 h at 30 °C. Based on cell lysis treatments, bacteriocin of Lact. lactis subsp. lactis YBD11 was determined to have a bactericidal activity against M. luteus. Bacteriocin production occurred through-out the exponential phase, with the highest activity recorded at the end of this phase. Tricine-SDS-PAGE of partially purified bacteriocin gave the same molecular weight as nisin (3.5 kDa). These results indicate that the antimicrobial compound produced by Lact. lactis subsp. lactis YBD11 is a nisin-like bacteriocin (Tuncer & Ozden, 2010).
A different bacterial strain isolated from boza produced in Turkey was also iden-tified, and the physicochemical and microbiological properties of its inhibitory compounds were characterised (Sahingil, Isleroglu, Yildirim, Akcelik, & Yildirim, 2011). The isolate was identified as Lact. lactis subsp. lactis, based on morphology, physiology, carbohydrate fermentation, the fatty acid profile and 16S rDNA gene sequence homology. Lactococcin BZ was active against several Gram-positive and Gram-negative foodborne pathogens and food spoilage bacteria. Lactococcin BZ was produced at the maximum level in MRS broth with an inoculum volume of 0.1%, an initial pH of 7.0, and an incubation temperature of 25 °C. Bacteriocin production began during the logarithmic phase and reached the maximal level during the early stationary phase. Its mode of action against Li. monocytogenes was bactericidal and its molecular weight was ca. 5.5 kDa, as determined using tricine SDS-PAGE. Lacto-coccus lactis subsp. lactis BZ or its bacteriocin, which has a wide inhibitory spectrum, has the potential for use as a biopreservative in food products (Sahingil et al., 2011).
Five bacteriocin-producing LAB strains (Lb. plantarum ST69BZ, Ent. faecium ST62BZ and Leuc. lactis ST63BZ, ST611BZ and ST612BZ) were isolated from boza originating from Belogratchik, Bulgaria (Todorov, 2010). The bacteriocins of all five isolates inhibited the growth of Enterococcus spp., E. coli, K. pneumoniae, Lactobacillus spp., Lact. lactis subsp. lactis, Listeria spp., Ps. aeruginosa, Staphylococcus spp. and Streptococcus caprinus. The mode of activity of bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ and ST612BZ is bactericidal, as determined against Ent. faecium HKLHS and Lb. sakei DSM 20017, respectively. Two of the five studied bacteriocino-genic strains (ST69BZ and ST612BZ) exhibited activity against some fungal cultures. In addition, a synergistic effect between bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ and ST612BZ and ciprofloxacin were registered (Todorov, 2010).
The highest level of bacteriocin ST69BZ activity (12,800 AU/ml) was recorded after 18 h of growth in MRS broth (30 °C), and it remained at this level for the dura-tion of fermentation. The highest level of bacteriocin ST62BZ activity (25,600 AU/ml) was recorded after 21 h of growth, followed by stable production for the next 3 h. Similar results were recorded for bacteriocin ST63BZ. Maximum levels of bacterio-cin ST63BZ (1600 AU/ml) were recorded after 24 h, at the end of stationary growth. Bacteriocin ST611BZ reached 52,600 AU/ml after 15 h, but decreased to 25,600 AU/ml during the following 9 h. Similar results were recorded for strain ST612BZ. Opti-mum levels of bacteriocin ST612BZ was recorded at 15 h (6400 AU/ml), followed by a decrease to 3200 AU/ml during the following 9 h (Todorov, 2010). The size of

136 Advances in Fermented Foods and Beverages
bacteriocins ST69BZ (3.2 kDa), ST62BZ (10.0 kDa), ST63BZ (10.0 kDa), ST611BZ (3.2 kDa) and ST612BZ (6.5 kDa) (Todorov, 2010) was similar to that described for other bacteriocins produced by LAB isolated from boza. Bacteriocins JW3BZ, JW6BZ, JW11BZ and JW15BZ are between 2.3 and 3.3 kDa (Von Mollendorff et al., 2006). Similar results were reported for bacteriocins ST194BZ (3.0 and 14.0 kDa), ST242BZ (10.0 kDa), ST284BZ (3.5 kDa), ST414BZ (3.7 kDa), ST461BZ (2.8 kDa), ST462BZ (8.0 kDa), ST664BZ (6.5 kDa) and ST712BZ (14.0 kDa) (Todorov & Dicks, 2006). The sizes recorded for the five bacteriocins (ST69BZ, ST62BZ, ST63BZ, ST611BZ and ST612BZ) are within the range reported for most bacteriocins produced by Lactobacillus spp. and Enterococcus spp. (De Vuyst & Vandamme, 1994).
Complete inactivation of the bacteriocins was observed after treatment of the cell-free supernatants with proteolytic enzymes, confirming the proteinaceous nature of the antimicrobial compounds (Todorov, 2010). Treatment of cell-free supernatants of the four strains with catalase and α-amylase did not result in activity changes, except for bacteriocin ST63BZ, suggesting that the inhibition factor recorded was not hydro-gen peroxide and that carbohydrate moieties were not required for antimicrobial activ-ity (Todorov, 2010). Inhibition of bacteriocin ST63BZ activity by α-amylase suggests that the bacteriocin is glycosylated and belongs to group IV according to the classifi-cation of Klaenhammer (1988). Stability of the other four bacteriocins in the presence of α-amylase is not unusual, and similar results have also been reported for other bac-teriocins isolated from boza (Todorov & Dicks, 2006; Von Mollendorff et al., 2006). Leuconocin S (Keppler, Geiser, & Holzapfel, 1994) and carnocin 54 (Lewus, Sun, & Montville, 1992) are sensitive to α-amylase, suggesting that their activity is associated with glycosylation of the active peptide.
Thermostability at 100 °C has also been reported for most other bacteriocins (Todorov & Dicks, 2004, 2005, 2006; Von Mollendorff et al., 2006) isolated from boza. The sensitivity of bacteriocins ST63BZ and ST612BZ to 100 °C after 120 min and 121 °C after 20 min may be a result of their molecular mass (10.0 and 6.5 kDa, respectively), although the stability of bacteriocin ST62BZ, a 10.0 kDa peptide was not affected by treatment for 120 min at 100 °C. A difference in the structures of bac-teriocin ST62BZ and ST63BZ may be a reason for these results. Bozacin B14 was inactivated after 10 min at 90–121 °C (Ivanova et al., 2000).
The mode of activity of bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ and ST612BZ is bactericidal, as determined against Ent. faecium HKLHS and Lb. sakei DSM 20017, respectively (Todorov, 2010). The results obtained regarding the leak-age of DNA, RNA, proteins and β-galactosidase confirm that bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ and ST612BZ destabilise the permeability of the cell membrane (Todorov, 2010).
Cell-free supernatants from 24-h-old cultures of Lb. plantarum ST69BZ, Leuc. lac-tis ST63BZ, ST611BZ, ST612BZ and Ent. faecium ST62BZ (pH neutralised) inhib-ited the growth of several bacterial strains such as Li. monocytogenes, Ent. faecalis, and E. coli (Meinken & Todorov, 2009). Early log phase cells of test microorganisms treated with bacteriocins resulted in immediate and complete growth inhibition for at least 10 h (Meinken & Todorov, 2009), suggesting that the mode of action is bac-tericidal. Cells of test microorganisms treated with bacteriocins ST69BZ, ST62BZ,

137Traditional cereal fermented foods as sources of functional microorganisms
ST63BZ, ST611BZ, and ST612BZ were clearly deformed or vesiculated as visual-ised by AFM (Meinken & Todorov, 2009). Sensitive strains treated with bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ, and ST612BZ resulted in leakage of DNA, RNA, proteins, and β-galactosidase. The results obtained by AFM shown by Meinken and Todorov (2009) and leakage of DNA, RNA, proteins, and β-galactosidase confirm that bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ, and ST612BZ destabilise the permeability of the cell membrane. Vesiculation was clearly visible on cells of Lb. sakei DSM20017 after treatment with bacteriocin ST62BZ produced by Ent. faecium ST62BZ, and bacteriocins ST63BZ, ST611BZ, and ST612BZ produced by Leuc. lac-tis ST63BZ, ST611BZ, and ST612BZ, respectively. Changes in morphology, such as collapse and formation of pores of Ent. faecium HKLHS, were observed after treat-ment with bacteriocin ST69BZ produced by Lb. plantarum ST69BZ. The resulting images clearly showed changes in cell morphology, such as collapse of the apical ends or the cell center, signs of cytoplasm leakage or vesiculation. Differences observed between the bacteriocins suggest different modes of action, such as the barrel stave model and the toroidal model, which describe the formation of pores in the cell mem-brane or the carpet model, which leads to a vesiculation of the outer cell membrane (Meinken & Todorov, 2009).
It was interesting to find that two of these five strains exhibited antifungal activ-ity. Lactobacillus plantarum ST69BZ culture showed activity against Absidia spp., Aspergillus niger, Epicoccum nigrum, and Penicillium spp., whereas Leuc. lactis ST612BZ produced an antifungal substance active against Botrytis spp. (Todorov, 2010). As these bacterial species are present in boza, we can presume that they could be an important part of the starter cultures and may therefore contribute to the antifun-gal stability of the product.
Todorov (2010) reported that, in the combined application of the sublethal lev-els of clinical antibiotic (ciprofloxacin) and five bacteriocins (bacteriocins ST69BZ, ST62BZ, ST63BZ, ST611BZ and ST612BZ), antibacterial activity was strongly increased. These results indicate that the mechanism by which the cationic peptide increases the effectiveness of these antibiotics would be through the dissipation of the proton gradient responsible for the extrusion of these compounds. The synergism between antibiotics, particularly ciprofloxacin and bacteriocins, is important to reduce the level of the MIC of the antibiotics. Similar synergetic effects may be important for the development of treatments against multi-drug-resistant strains.
6.4 Fermented cereal-based food from Africa and Latin America
In Africa, numerous traditional fermented cereal-based foodstuffs are produced in small-scale production units in urban areas (Chavan & Kadam, 1989). Some exam-ples of African traditional fermented cereal-based products are summarised in Table 6.1. Cereal grains including sorghum, maize and millet are common substrates for lactic acid–fermented gruels and beverages known by different names such as, for

138 Advances in Fermented Foods and Beverages
example, poto poto in the Congo (Louembé, Brauman, Tchicaya, & Kobawila, 1996), ben-saalga (Tou et al., 2006) and dégué in Burkina Faso, ogi in Nigeria (Banigo & Akinrele, 1977), koko in Ghana (Andah & Muller, 1973) and togwa in Tanzania (Lorri & Svanberg, 1995; Mugula, Nnko, & Sorhaug, 2001). Poto poto is a traditional fer-mented maize dough prepared in Congo homes for direct use as a weaning food. The process includes a soaking step of the grains (first fermentation step), which on average lasts 55 h, followed by milling and settling of the paste in water (second fermentation step, 10–11 h), and cooking to produce a gruel (Ampe & Miambi, 2000; Louembé, Kéléke, Kobawila, & Nzouzi, 2003). The fermented maize paste is dewatered and modelled into poto poto balls, which can be sold as such or diluted and cooked in water with added sugar (poto poto gruel). Dégué is a pearl millet–based fermented food from Burkina Faso. Dégué has a very different processing diagram resulting after dehulling, grinding of the millet grains, modelling with water and steam cooking in the production of gelatinous balls, which are further fermented during storage (24 h) and afterward consumed mixed with dairy products. In both cases, fermentation is spontaneous and uncontrolled, thus resulting in products of variable quality (Abri-uel et al., 2006). In a taxonomic study of poto-poto and dégué, African traditional fermented food products by sequencing of DNA bands from TTGE gels revealed the presence of Lactobacillus gasseri, Enterococcus sp., E. coli, Lb. plantarum/paraplan-tarum, Lb. acidophilus, Lactobacillus delbrueckii, Bacillus sp., Lactobacillus reuteri, and Lb. casei in poto-poto and Lb. gasseri, Enterococcus sp., E. coli, Lb. fermentum, Lb. brevis, and Lb. casei in dégué (Abriuel et al., 2006).
In some regions of Latin America, fermented products are still manufactured traditionally using very simple equipment. Some examples of Latin American tra-ditional fermented cereal-based products are presented in Table 6.1. In specialised markets throughout the world, these fermented products are highly appreciated and are considered to be of premium value because of their flavour characteristics and uniqueness. Indigenous fermented foods have special organoleptic qualities, and some even possess health-promoting properties. This is principally due to the presence of a biologically diverse microbiota present in the raw material or as a contaminant from either the producers or instruments used for preparation. These microorganisms are important genetic reservoirs and hold great biotechno-logical and health-improving potential that should be exploited. Those adapted to harsh environmental conditions were able to survive, whereas others had the ability to produce enzymes and compounds necessary for their growth on raw food materials.
Chicha de jora is one of the oldest fermented beverages in South America. It is believed that its origin dates back to the fifteenth century. Maize was first chewed to pulp, since saliva helps to convert starches to fermentable sugars. This pulp was then sun dried, mixed with warm water and left to ferment for a few days, converting the corn into a mildly alcoholic drink. In ancient Japan, sake (rice wine) was also produced by chewing the grains as a source of amylase to convert the starch to mono-saccharides and disaccharides (Yoshizawa & Ishikawa, 1979). Later it was discovered that rice overgrown with Aspergillus, Rhizopus or Mucor moulds also became sweet and could be fermented to produce sake (Steinkraus, 1979).

139Traditional cereal fermented foods as sources of functional microorganisms
Nowadays, Chicha de jora is prepared by first soaking the maize grains in water for 24–48 h, after which the water is drained and the corn allowed to germinate for 8–15 days, covered in dampened plant leaves and finally sun dried. During the germi-nation process, the starches are converted into fermentable sugars (mono- and disac-charides) by the action of amylases, whereas proteins are broken down by proteases, both activities being necessary for yeast growth. The germinated maize is then boiled for 6–24 h, during which different herbs and spices are added such as cinnamon, all-spice and cloves. The liquid is allowed to cool, is strained through a clean cloth, unre-fined sugar is added, and the mixture is allowed to ferment for 1–2 weeks in clay pots. The final fermented product has a final pH of 4.0–4.5 and an alcohol content of 4–6%. However, although Chicha de jora is a product known since ancient times, very few studies have been carried out on the fermentation process. It was shown that Sacch. cerevisiae is the predominant native yeast species present (Quillama, 1993), and that LAB (such as Lb. plantarum and Lb. fermentum) are also present in large quantities (Quillama et al., 1995). It was shown that Lb. plantarum E2, a strain isolated from Chicha de jora, is capable of producing a bacteriocin that limits the growth of Lb. fermentum Chj4C, another strain isolated from the native beverage (Quillama, 1998).
Pozol is a nonalcoholic beverage made from fermented maize that has been pro-duced by the Maya Indians and Mestizo populations in southeastern Mexico and other Central American countries since pre-Hispanic times (Wacher et al., 2000). Today it still forms part of the basic daily nutrition of urban and rural populations of different ethnic groups that reside in this region (Jiménez Vera, González Cortés, Magaña Contreras, & Corona Cruz, 2010). The first step in pozol preparation is called nixtamalisation, where maize kernels (white, yellow or black) are boiled for 1.5 h in a 1% lime solution. These are then dehulled and rinsed using tap water (the discard is called nexayote). The maize is then coarsely ground to make dough (called nixtamal) that is shaped into balls, wrapped in banana leaves, and normally left to ferment at ambient temperatures for 2–7 days but sometimes up to 1 month. The fermented dough is then suspended in water and drunk during meals, at work, or anytime during the day as a refreshing beverage. Traditionally, pozol was produced primarily for family consumption, the drink being consumed by adults, children and infants (Ulloa, Herrera, & Lappe, 1987), although sometimes slightly larger-scale producers have made pozol for sale. Some fibrous components are not completely solubilised by nixtamalisation, and a sediment is present in the beverage when the dough is suspended in water. The Mestizo population has apparently modified the Indian process by adding a second boiling of the grains in water before grinding to reduce sediment formation (Wacher et al., 2000).
In the first complete study of the microbiota of pozol, it was shown that freshly prepared pozol contained 104–106 CFU/g of LAB, l04–105 aerobic mesophiles, 10–103 Enterobacteriaceae, 10–104 yeasts, and <103 mould propagules (Wacher, Cañas, Cook, Barzan, & Owens, 1993). After 30 h at 28 °C, the numbers were 109, 7 × 106, 5 × 105, 106 and 104, respectively. Soaking alkali-treated grains overnight allowed LAB, aerobic mesophiles and Enterobacteriaceae to grow, and these then constituted the primary microbiota of the pozol dough. It was shown that, although the addi-tional cooking stage added in the Mestizo process significantly modified the physical

140 Advances in Fermented Foods and Beverages
properties of the dough, no significant differences were observed in the microbial com-position of Mestizo compared to traditionally prepared pozol (Wacher et al., 2000).
The main soluble sugar of maize is sucrose, which is present at a concentration of 2% (w/w) of the whole kernel on a dry-weight basis. This concentration is reduced to 0.1–0.7% (w/w) of dry dough after alkaline cooking, soaking and washing to pro-duce nixtamal (Diaz-Ruiz, Guyot, Ruiz-Teran, Morlon-Guyot, & Wacher, 2003). This concentration would be insufficient to maintain microbial diversity and the high bac-terial concentration reported in pozol (Diaz-Ruiz et al., 2003). Although LAB are the dominant group during all stages of pozol fermentation, as has been shown previ-ously by classical culture methods (Nuraida, Wacher, & Owens, 1995; Wacher et al., 2000, 1993) and culture-independent methods (Ampe, Ben Omar, Moizan, Wacher, & Guyot, 1999; Escalante, Wacher, & Farres, 2001), significant changes in the popu-lation dynamics occur throughout pozol production. For example, it was shown that high concentrations of amylolytic LAB (ALAB) were detected at the beginning of the fermentation process, and that a relatively high number of nonamylolytic LAB were observed at the end of fermentation (Diaz-Ruiz et al., 2003). These results suggest a symbiosis in that ALAB would first degrade starch at the beginning of the fermenta-tion, decreasing the pH of the dough and liberating sugars that could be used for the growth of nonamylolytic microorganisms.
LAB (such as Lb. plantarum and Lb. fermentum, together with members of the gen-era Leuconostoc and Weissella) accounted for 90–97% of the total active microbiota of pozol; no streptococci were isolated, although members of the genus Streptococcus accounted for 25–50% of the microbiota (Ampe et al., 1999). The presence of Bifidobacterium, Enterococcus and enterobacteria suggests a fecal origin of some important pozol microorganisms (Ben Omar & Ampe, 2000).
A most comprehensive study on microbial community dynamics during the pro-duction of pozol was performed using traditional and culture-independent techniques (Ben Omar & Ampe, 2000). It was shown that Streptococcus species dominated the fermentation and accounted for between 25% and 75% of the total microbiota throughout the process. The initial aerobic microbiota was replaced in the first 2 days by heterofermentative LAB (closely related to Lb. fermentum), and this heterolactic microbiota was then progressively replaced by homofermentative LAB (mainly by genetically close relatives of Lb. plantarum, Lb. casei and Lb. delbrueckii), which continued acidification of the maize dough. At the same time, an extremely diverse community of yeasts and fungi developed, mainly at the periphery of the dough. This study demonstrated that a relatively high number of species, at least six to eight, is needed to perform pozol fermentation. Overall, the results obtained with different cul-ture-dependent or culture-independent techniques clearly confirmed the importance of developing a polyphasic approach to study the ecology of fermented foods.
Since little sanitary measures are taken during pozol preparation, microbial contam-ination of the maize dough is inevitable. Potentially pathogenic fungi such as Candida parapsilosis, Trichosporon cutaneum, Geotrichum candidum and Aspergillus flavus have been recovered from pozol during the first few hours of fermentation (Ulloa et al., 1987). Although most of these pathogens are killed off by the decrease in pH during the fermentation process, serious health problems can still arise, especially if fresh

141Traditional cereal fermented foods as sources of functional microorganisms
underfermented pozol is consumed. It was recently shown that 19% of pozol sam-ples were contaminated with aflatoxin B2 (AFB2) and traces of aflatoxin B1 (AFB1), but only one sample contained aflatoxin concentrations >20 ppb (Mendez-Albores, Arambula-Villa, Preciado-Ortiz, & Moreno-Martinez, 2004). In this study, it was also shown that pozol prepared with white maize and samples prepared with cacao showed the highest ranges of contamination, whereas, when yellow kernels were used, the presence of aflatoxins was not detected. Amerindians consume equal amounts of pozol prepared using white, yellow and black maize, although popular belief has been that the yellow and black varieties contained higher vitamin levels. Mestizos normally use white maize for pozol preparation and also consume chorote, a pozol derivative, in which ground cocoa beans are added to the maize mixture. The former population would thus be less likely to consume aflatoxin containing pozol than the latter; how-ever, in Mestizo preparation, the additional boiling step would be helpful in eliminat-ing aflatoxin-producing microbes.
6.5 Starter cultures and cereal-based fermented food
The use of probiotic LAB, especially Lactobacillus and Bifidobacterium spp., as starter cultures, alone or in combination with traditional starter cultures in various fermentation processes, has been explored for different fermentation food products. Formulated fermented probiotic food may present consumers with a healthy dietary component at a considerabley low cost (Goldin, 1998). Furthermore, it was reported that LAB may contribute to microbiological safety and/or provide one or more techno-logical, nutritional and organoleptic advantages to a fermented product, through pro-duction of ethanol, acetic acid, aroma compounds, exopolysaccharides, bacteriocins and several enzymes (Leroy & De Vuyst, 2004).
Various developments over the years have led to the concept of starter cultures. The earliest fermented food products relied on natural fermentation through microbiota present in the raw material. The load and spectrum of microorganisms populating the raw material have a definite effect on the quality of the end product. Backslopping, that is, inoculation of the raw material with a small quantity of a previously performed successful fermentation, was used to optimise spontaneous fermentation. This is still the basis for the home-scale production of several fermented food products, including, for example, yoghurt, kefir, traditional beer and boza. In this case, the best-adapted strain dominates. The dominant strains can be seen as a starter culture that shortens the fermentation process and reduces the risk of fermentation failure (Leroy & De Vuyst, 2004). Backslopping is still used in developing countries and even in the industrialised countries for production of sauerkraut and sourdough (Holzapfel, 1997). The use of starter cultures in large-scale production of fermented foods has become important for industries in developed countries, as it resulted in improved control over the fermen-tation process and a consistent end product. However, some disadvantages do occur due to the fact that commercial starter cultures were not selected in a rational way but, rather, on the basis of phage resistance and rapid acidification of the raw materials (Leroy & De Vuyst, 2004). With regard to the functionality and desired properties of

142 Advances in Fermented Foods and Beverages
the end product, these starters are not very flexible, and they are frequently difficult to be keep even by modern microbiological procedures. Furthermore, it is believed that commercial starter cultures adapted to the food matrix led to a loss in genetic material (Leroy & De Vuyst, 2004). This may have contributed to the limited biodiversity of commercial starter cultures. Moreover, this leads to a product that lacks the unique-ness and characteristics that made the original product popular (Caplice & Fitzgerald, 1999).
Wild-type LAB that originate from the environment, raw materials and process apparatus serve as natural starter cultures (“inoculation”) for many of the traditionally fermented foods (Böcker, Stolz, & Hammes, 1995; Weerkamp, Klijn, Neeter, & Smit, 1996). Recent studies focused on the use of wild-type strains isolated from traditional products for use as starter cultures (Beukes, Bester, & Mostert, 2001; De Vuyst et al., 2002; Hébert, Raya, Tailliez, & De Giori, 2000) as a resource of improved technolog-ical, beneficial and functional properties. When considering LAB as a starter culture, the following factors have to be taken into account: (1) not all LAB strains have the same practical and technical importance in food fermentations; (2) the genera Lactobacillus (Lb. fermentum, Lb. plantarum, Lb. reuteri, Lb. rhamnosus), Leucono-stoc, and, to a lesser extent, Lactococcus, Enterococcus, Pediococcus and Weisella spp. are usually present in traditional fermented foods; (3) not all strains of the same species are suitable as starter cultures and (4) various industrial scale lactic acid fer-mentation processes are well controlled despite the fact that they are spontaneous (Holzapfel, 2002). Some of these LAB may be classified as functional and beneficial starters, due to their contribution to food safety, sensory properties and other nutri-tional advantages.
LAB are known to produce antimicrobial substances (including bacteriocins, organic acids, diacetyl and low-molecular-weight substances), polymers, sugars, sweeteners, nutraceuticals, aromatic compounds and various enzymes (including pro-teolytic enzymes). This leads to greater flexibility and wider application of LAB as starter cultures. It also represents a way by which chemical additives can be replaced by natural compounds and thus provide the consumer with new, appealing food prod-ucts (Leroy & De Vuyst, 2004). Bacteriocins produced by LAB may prevent food spoilage, for example, late blowing of cheese by clostridia (Thomas, Clarkson, & Delves-Broughton, 2002). Some probiotic strains may also be used as functional start-ers or co-cultures in fermented food (Chandan, 1999; Jahreis et al., 2002; Ross, Stanton, Hill, Fitzgerald, & Coffey, 2000).
However, when considering the use of probiotic strains as functional starters or co-cultures, it is important that they do not enhance acidification and thereby reduce the shelf-life of the product, and that they do not have adverse effects on the aroma or taste of the product (Heller, 2001). Uncertainty still exists as to whether multifunc-tional strains possessing all desirable metabolic features would result from modern isolation techniques and selection procedures. Therefore, recent studies focus on the improvement of selected strains by the application of recombinant DNA technology. By its application, certain beneficial features, such as health-promoting properties, accelerated acid production, wholesomeness and overproduction of specific enzymes or bacteriocins can be improved (Holzapfel, 2002). Gene disruption may be used to

143Traditional cereal fermented foods as sources of functional microorganisms
eliminate undesirable properties such as antibiotic and mycotoxin production by food-grade moulds (Geisen & Holzapfel, 1996; Hammes & Vogel, 1990). A large array of these optimised cultures is available but is not used because of legal regulations (Holzapfel, 2002).
6.6 Cereal-based probiotic foods
The concept of probiotic foods and feeds has been developed to quite an extent since its introduction to clinical nutrition and food science during the 1980s (Fuller, 1989; Shortt, 1999). The majority of probiotic food products available today are milk-based, although a few attempts have been made to use cereals. Cereal grains have a high nutritive value and are distributed worldwide, making cereal a very suitable raw mate-rial for the development of various fermented functional foods (Angelov, Gotcheva, Kuncheva, & Hristozova, 2006). Togwa, a lactic acid–fermented maize and sorghum gruel, inhibits the growth of some enterotoxin-producing bacteria in children <5 years of age. This suggests that togwa may possess probiotic properties (Kingamkono, Sjögren, & Svanberg, 1998). Vogel et al. (1999) found that the LAB present in various LAB-fermented foods, such as sourdough, is similar or, in some cases, identical to species found in the gastrointestinal tract of humans and animals. Lactobacillus plan-tarum indigenous to a variety of cereal-based fermented food is also associated with the gastrointestinal tract of humans (Ahrné et al., 1998; Molin, 2001). Colonisation of the intestinal mucosa with strains of Lb. plantarum isolated from sourdough has also been reported (Johansson, Molin, Jeppsson, Nobaek, & Ahrné, 1993).
The early Mayans used pozol as a source of nutrients and in ceremonies promoting the growth and harvest of maize. They also used this fermented beverage as a med-icine to control diarrhoea, to reduce fever and to cure intestinal infections (Phister, O’Sullivan, & McKay, 2004). Microbiological studies on pozol provided evidence in support of these popular beliefs. It has been stated earlier that LAB are well known for their health-promoting capabilities, and, since LAB are the dominant microbial species in pozol, it is not surprising that this refreshing beverage possesses health-pro-moting properties. Antimicrobial compounds produced by Bacillus sp. strain CS93 isolated from pozol were identified and exhibited activities against several Gram-pos-itive and Gram-negative bacteria, yeasts and moulds (Phister et al., 2004). It was also shown that Agrobacterium azotophilum, present in pozol, possesses bacterio-cidal, bacteriolytic, bacteriostatic and fungistatic activities against a wide range of pathogenic microorganisms (Ulloa & Herrera, 1972). All of these findings, along with future studies, will surely help in promoting the consumption of this interesting Mayan drink.
Barley and oats contain β-glucan (Angelov et al., 2006), a prebiotic that may contribute to the reduction of LDL-cholesterol levels by 20–30%, and in this way may also contribute to a decrease in the risk of cardiovascular diseases (Gallaher, 2000; Stark & Madar, 1994; Wrick, 1994). For a polysaccharide or oligosaccharide to be characterised as a prebiotic, it should withstand digestion in the upper part of the gastrointestinal tract, be hydrolysable, be soluble, and stimulate the growth

144 Advances in Fermented Foods and Beverages
and activity of beneficial microbiota in the gut (Gibson & Roberfroid, 1995). The low glycemic index of oats and barley is quite beneficial to diabetes in the gastro-intestinal tract after ingestion, as it alters the level of fat emulsification and reduces lipase activity (Angelov et al., 2006). Furthermore, β-glucan stimulates the growth of beneficial bacteria associated with the colons of animals and humans (Jaskari et al., 1998; Wood & Beer, 1998).
To increase the number of beneficial bacteria in the gut, large numbers of probiotic bacteria have to be consumed by means of capsules or by using food as vector. Incor-porating suitable dietary polysaccharides or oligosaccharides to the capsules may be even more effective. The latter is referred to as the prebiotic concept. Arabinoxylan is another prebiotic compound commonly found in wheat and rye (Crittenden et al., 2002; Jaskari et al., 1998; Karppinen, 2003).
6.6.1 Beneficial properties of lactic acid bacteria isolated from boza
According to the definition of the World Health Organisation, probiotics are live microorganisms that, when administrated in adequate amounts, confer a health ben-efit on the host (Fuller & Gibson, 1997). This includes reduction of gastrointestinal infections and inflammatory bowel disease, and modulation of the immune system (Fuller & Gibson, 1997). Probiotic strains should survive the harsh conditions of the gastrointestinal tract and adhere to intestinal epithelial cells. They form a defense against the colonisation of pathogenic microorganisms by competing for adher-ence to mucus and epithelial cells and, in certain cases, production of hydrogen peroxide and bacteriocins (Boris &and Barbes, 2000; Lepargneur &and Rousseau, 2002; Reid &and Burton, 2002; Velraeds, Van De Belt-Gritter, Van Der Mei, Reid, & Busscher, 1998). Changes in diet, stress, contraceptives, medication and micro-bial infection disturb the microbial balance, which often leads to a decrease in the number of viable LAB (Fuller & Gibson, 1997). The subsequent uncontrolled pro-liferation of pathogenic bacteria may lead to diarrhoea and other clinical disorders such as cancer, inflammatory bowel disease and ulcerative colitis (Fooks, Fuller, & Gibson, 1999). A variety of Lb. plantarum strains are presently marketed as pro-biotics (De Vries, Vaughan, Kleerebezem, & De Vos, 2006). The best-studied pro-biotic strains are from the species Lb. acidophilus, Lb. fermentum, Lb. plantarum, Lb. brevis, Lactobacillus jensenii, Lb. casei, Lb. delbrueckii, Lactobacillus vagina-lis and Lactobacillus salivarius. A number of surface-anchored proteins have been described for Lb. plantarum (Kleerebezem et al., 2003), suggesting that the species has the potential to associate with many different surfaces and substrates, and to adapt to changing environmental conditions.
Criteria for the selection of probiotic strains have been formulated by the Food and Agriculture Organisation of the United Nations and the World Health Organi-zation (FAO/WHO, 2001). Some of the most important criteria are gastric and bile acid resistance, adhesion to mucus and human epithelial cells, competition with pathogens for adhesion sites, growth inhibition of potentially pathogenic bacteria, bile salt hydrolase activity and, in the case of vaginal applications, resistance to

145Traditional cereal fermented foods as sources of functional microorganisms
contraceptives (FAO/WHO, 2001). The concept of probiotic foods has been devel-oped extensively since its introduction to clinical nutrition and food science during the 1980s (Fuller, 1989; Shortt, 1999). Most probiotic foods available today are milk based, although a few attempts have been made to use cereals. Cereal grains are very suitable raw materials for the development of various fermented functional foods (Angelov et al., 2006). Examples of beneficial cereal-based fermented prod-ucts are togwa, a LAB-fermented maize and sorghum gruel (Kingamkono et al., 1998). Vogel et al. (1999) reported on LAB present in various fermented foods, such as sourdough, that are similar or, in some cases, identical to species found in the gastrointestinal tracts of humans and animals. Lactobacillus plantarum indigenous to a variety of cereal-based fermented foods is also associated with the gastrointes-tinal tract of humans (Ahrné et al., 1998; Molin, 2001). Colonisation of the intes-tinal mucosa with strains of Lb. plantarum isolated from sourdough has also been reported (Johansson et al., 1993).
A number of LAB with probiotic properties have been isolated from boza, a tradi-tional beverage produced from the fermentation of cereals. Lactobacillus plantarum (strains ST194BZ, ST414BZ and ST664BZ), Lb. pentosus (strain ST712BZ), Lb. rhamnosus (strains ST461BZ and ST462BZ) and Lb. paracasei (strains ST242BZ and ST284BZ) have been described previously as bacteriocin producers with activ-ities against several Gram-positive and Gram-negative test organisms (Todorov & Dicks, 2006). Strains ST194BZ, ST242BZ, ST284BZ, ST414BZ, ST461BZ, ST462BZ, ST664BZ and ST712BZ were shown to have potential probiotic proper-ties (Todorov et al., 2008), as all of them survived low pH conditions (pH 3.0), grew well at pH 9.0 and were not affected by the presence of 0.3% (w/v) oxbile. Cyto-toxicity levels of the bacteriocins, expressed as CC50, ranged from 38 μg/ml for bac-teriocin ST194BZ to 3776 μg/ml for bacteriocin ST284BZ. Bacteriocin ST284BZ, bacteriocin ST461BZ and bacteriocin ST462BZ were the least cytotoxic. Bacterio-cin ST284BZ revealed high activity (EC50 = 735 μg/ml) against the HSV-1 virus that causes encephalitis and oro-facial and genital lesions. Growth of Mycobacterium tuberculosis was repressed by 69% after 5 days of incubation in the presence of the bacteriocin ST194BZ. Various levels of auto- (self-) aggregation between the probiotic bacteria and co-aggregation with Li. innocua LMG 13568 were observed. Adhesion of strains ST194BZ, ST284BZ, ST414BZ, ST461BZ, ST462BZ and ST664BZ to HT-29 cells ranged from 18% to 22%, which is similar to that reported for Lb. rhamnosus GG. Adherence of strains ST194BZ, ST242BZ and ST712BZ to Caco-2 cells ranged between 7.0% and 9.0%, similar to values reported for Lb. rhamnosus GG. High hydrophobicity readings were recorded for most of the pro-biotic strains. Strain ST712BZ revealed only 38% hydrophobicity, but 63% of the cells adhered to HT-29 cells, compared to 32% adherence recorded for Lb. rhamno-sus GG. Growth of strains ST194BZ, ST242BZ, ST284BZ, ST414BZ, ST461BZ, ST462BZ, ST664BZ and ST712BZ were inhibited by only seven of 24 medicaments tested (Todorov et al., 2008).
Enterococcus mundtii ST4V isolated from soya beans produces a broad-spectrum bacteriocin active against Gram-positive and Gram-negative bacteria and has anti-viral activity (Todorov et al., 2009). The aim of the study of Todorov et al. (2009)

146 Advances in Fermented Foods and Beverages
was to determine the probiotic properties of strain Ent. mundtii ST4V, to evaluate its survival in boza, and to study the antimicrobial activity of the strain in situ. For this purpose, boza was prepared based on the traditional recipe (Todorov, 2010; Todorov et al., 2009). Cereals have been cooked and inoculated with Ent. mundtii ST4V (a potential probiotic and bacteriocin-producing strain). Control batches have been prepared with commercial boza (as deliver vector of starter cultures). All samples received also Sacch. cerevisiae. Fermentation was carried out at 37 °C for 3 h. The organoleptic properties of fermented products were evalu-ated by a qualified taste panel. No significant differences in rheological properties were observed, suggesting that Ent. mundtii ST4V had no negative effect on the sensory qualities of the final product. Microbial cell numbers remained relatively unchanged during 1 week of storage. Sensory analysis of the six boza preparations was reported in the study of Todorov et al. (2009). A slightly greater bitterness was recorded when boza was produced by the starter culture of commercial boza. Similar results were obtained for this sample with regard to a stronger yeasty flavour of the product when commercial boza was used as starter culture. These differences, albeit statistically significant, were so small that they would probably not be detected by the consumers. The acid taste of all samples was relatively low, with the lowest recorded for boza prepared with Ent. mundtii ST4V. No correla-tion was found between the different sensory attributes tested. Fermentation of the product contributes to the acidity of the product (Todorov et al., 2009). The preser-vative properties of bacteriocin ST4V were evaluated by contaminating boza with Lb. sakei DSM 20017. Changes in microbial populations were monitored by using classical microbiological methods, PCR with species-specific primers and dena-turing gradient gel electrophoresis (DGGE). Enterococcus mundtii ST4V survived 7 days in boza that was stored at 4 °C, and produced bacteriocin ST4V at levels high enough to prevent the growth of the target strain Lb. sakei DSM 20017T. No off-flavours or abnormal textural changes were recorded in boza during storage. Boza could thus be developed as a vector to deliver strain ST4V (Todorov et al., 2009).
Incorporation of probiotic strains in cereal-based fermented foods is feasible and holds great promise for the future. Cereals are high in nutrition and confer specific health benefits. In addition, cooked bran provides an excellent substrate for several strains of probiotic bacteria. A snack exhibiting the combined postulated beneficial effects of bran and probiotic bacteria may serve as an alternative to soy-based and milk-based yogurts (Salovaara, 1996; Salovaara & Simonson, 2003). In general, oats are a suitable substrate for fermentation with probiotic LAB after appropriate processing (Johansson et al., 1993; Marklinder & Lonner, 1992; Salovaara, 1996;
Salovaara & Simonson, 2003).
Acknowledgements
The author would like to thank CAPES, CNPq and FAPESP (Brasil) for their financial support.

147Traditional cereal fermented foods as sources of functional microorganisms
References
Abriouel, H., Ben Omar, N., López, R. L., Martínez-Cañamero, M., Keleke, S., & Gálvez, A. (2006). Culture-independent analysis of the microbial composition of the African tradi-tional fermented foods poto poto and dégué by using three different DNA extraction meth-ods. International Journal of Food Microbiology, 111, 228–233.
Ahrné, S., Nobaek, S., Jeppsson, B., Adlerberth, I., Wold, E. E., & Molin, G. (1998). The normal Lactobacillus flora of healthy human rectal and oral mucosa. Journal of Applied Microbiology, 85, 88–94.
Akpinar-Bayizit, A., Yilmaz-Ersan, L., & Ozcan, T. (2010). Determination of boza’s organic acid composition as it is affected by raw material and fermentation. International Journal of Food Properties, 13, 648–656.
Ampe, F., Ben Omar, N., Moizan, C., Wacher, C., & Guyot, J. P. (1999). Polyphasic study of the spatial distribution of microorganisms in Mexican pozol, a fermented maize dough, demonstrates the need for cultivation-independent methods to investigate traditional fer-mentations. Applied and Environmental Microbiology, 65, 5464–5473.
Ampe, F., & Miambi, E. (2000). Cluster analysis, richness and biodiversity indexes derived from denaturing gradient gel electrophoresis fingerprints of bacterial communities demonstrate that traditional maize fermentations are driven by the transformation process. International Journal of Food Microbiology, 60, 91–97.
Andah, A., & Muller, H. G. (1973). Studies on koko a Ghanian fermented maize porridge. Ghana Journal of Agricultural Science, 6, 93–102.
Angelov, A., Gotcheva, V., Kuncheva, R., & Hristozova, T. (2006). Development of a new oat – based probiotic drink. International Journal of Food Microbiology, 112, 75–80.
Arici, M., & Daglioglu, O. (2002). Boza: a lactic acid fermented cereal beverage as a traditional Turkish food. Food Reviews International, 18, 39–48.
Banigo, O., & Akinrele, I. A. (1977). Nigerian ogi. In K. Steinkraus (Ed.), Handbook (Vol. 9, pp. 189–198). New York and Basel: Dekker, M. Inc.
Ben Omar, N., & Ampe, F. (2000). Microbial community dynamics during production of the Mexican fermented maize dough pozol. Applied and Environmental Microbiology, 66, 3664–3673.
Beukes, E. M., Bester, B. H., & Mostert, J. F. (2001). The microbiology of South African tradi-tional fermented milks. International Journal of Food Microbiology, 63, 189–197.
Blandino, A., Al-Aseeri, M. E., Pandiella, S. S., Cantero, D., & Webb, C. (2003). Cereal-based fermented foods and beverages. Food Research International, 36, 527–543.
Böcker, G., Stolz, P., & Hammes, W. P. (1995). Neue Erkenntnisse zum Ökosystem Sauerteig und qur Physiologie der sauerteigtypischen Stämme Lactobacillus sanfrancisco und Lactobacillus pontis. Getreide, Mehl und Brot, 49, 370–374.
Boris, S., & Barbes, C. (2000). Role played by lactobacilli in controlling the population of vaginal pathogens. Microbes and Infection, 4, 543–546.
Botes, A., Todorov, S. D., von Mollendorff, J. W., Botha, A., & Dicks, L. M. T. (2006). Identi-fication of lactic acid bacteria and yeast from boza. Process Biochemistry, 42, 267–270.
Caplice, E., & Fitzgerald, G. F. (1999). Food fermentations: role of microorganisms in food production and preservation. International Journal of Food Microbiology, 50, 131–149.
Chandan, R. C. (1999). Enhancing market value of milk by adding cultures. Journal of Dairy Science, 82, 2245–2256.
Charalampopoulos, D., Wang, R., Pandiella, S. S., & Webb, C. (2002). Application of cereals and cereal components in functional foods: a review. International Journal of Food Micro-biology, 79, 131–141.

148 Advances in Fermented Foods and Beverages
Chavan, J. K., & Kadam, S. S. (1989). Nutritional improvement of cereals by fermentation. Critical Reviews in Food Science and Nutrition, 28, 349–393.
Crittenden, R., Karppinen, S., Ojanen, S., Tenkanen, M., Fagerström, R., Mättö, J., et al. (2002). In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bac-teria. Journal of the Science of Food and Agriculture, 82, 781–789.
De Vries, M. C., Vaughan, E. E., Kleerebezem, M., & De Vos, W. M. (2006). Lactobacillus plantarum – survivor, functional and potential probiotic properties in the human intestinal tract. International Dairy Journal, 16, 1018–1028.
De Vuyst, L., Schrijvers, V., Paramithiotis, S., Hoste, B., Vancanneyt, M., Swings, J., et al. (2002). The biodiversity of lactic acid bacteria in Greek traditional wheat sourdoughs is reflected in both composition and metabolite formation. Applied and Environmental Microbiology, 68, 6059–6069.
De Vuyst, L., & Vandamme, E. J. (1994). Bacteriocins of lactic acid bacteria: Microbiology, genetics and applications. London: Blackie Academic and Professional.
Diaz-Ruiz, G., Guyot, J. P., Ruiz-Teran, F., Morlon-Guyot, J., & Wacher, C. (2003). Microbial and physiological characterization of weakly amylolytic but fast-growing lactic acid bac-teria: a functional role in supporting microbial diversity in pozol, a Mexican fermented maize beverage. Applied and Environmental Microbiology, 69, 4367–4374.
Enikova, R., Kozereva, M., Ivanova, T., & Yang’ozova, Z. (1985). Sanitary microbiological analysis of boza. Khranitelnopromishlena Nauka, 1, 73–81.
Escalante, A., Wacher, C., & Farres, A. (2001). Lactic acid bacterial diversity in the traditional mexican fermented dough pozol as determined by 16S rDNA sequence analysis. Interna-tional Journal of Food Microbiology, 64, 21–31.
FAO. (1998). Fermented fruits and vegetables. A global perspective. FAO Agricultural Services Bulletin 134. Rome Italy: FAO.
FAO/WHO. (October 2001). Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Report of a Joint FAO/WHO Expert Consul-tation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. Rome, Italy: FAO.
Fooks, L. J., Fuller, R., & Gibson, G. R. (1999). Prebiotics, probiotics and human gut microbi-ology. International Dairy Journal, 9, 53–61.
Fox, P. F., Lucey, J. A., & Cogan, T. M. (1993). Cheese: an overview. In P. F. Fox (Ed.), Cheese: chemistry, physics and microbiology (2nd ed.). (pp. 1–36). London: Chapman and Hall.
Fuller, R. (1989). Probiotics in man and animals. A review. Journal of Applied Bacteriology, 66, 365–378.
Fuller, R., & Gibson, G. R. (1997). Modification of the intestinal microflora using probiotics and prebiotics. Scandinavian Journal of Gastroenterology, 32, 28–31.
Gallaher, D. D. (2000). Dietary fiber and its physiological effects. In M. Schmidt, & T. P. Labuza (Eds.), Essentials of functional foods. Gaithersburg, Maryland: Aspen Publishers Inc.
Geisen, R., & Holzapfel, H. W. (1996). Genetically modified starter and protective cultures. International Journal of Food Microbiology, 30, 315–324.
Gibson, G. R., & Roberfroid, M. B. (1995). Dietary modulation of the human colonic microbio-tia: introducing the concept of prebiotics. Journal of Nutrition, 125, 1401–1412.
Goldin, B. R. (1998). Health benefits of probiotics. British Journal of Nutrition, 80, 203–207.Gomez-Lopez, A., Mellado, E., Rodriguez-Tudela, J. L., & Cuenca-Estrella, M. (2005). Sus-
ceptibility profile of 29 clinical isolates of Rhototorula spp. and literature review. Journal of Antimicrobial Chemotherapy, 55, 312–318.
Gotcheva, V., Pandiella, S. S., Angelov, A., Roshkova, Z. G., & Webb, C. (2000). Microflora identification of the Bulgarian cereal – based fermented beverage boza. Process Biochem-istry, 36, 127–130.

149Traditional cereal fermented foods as sources of functional microorganisms
Hammes, W. P., Brandt, M. J., Francis, K. L., Rosenheim, J., Seitter, M. F. H., & Vogelmann, S. A. (2005). Microbial ecology of cereal fermentations. Trends in Food Science & Tech-nology, 16, 4–11.
Hammes, W. P., & Gänzle, M. G. (1998). Sourdough breads and related products. (2nd ed.). In B. J. B. Wood (Ed.), Microbiology of fermented foods (Vol. 1, pp. 199–216). London: Blackie Academic and Professional.
Hammes, W. P., & Vogel, R. F. (1990). Gentechnik zur Modifizierung von Starterorganismen. Lebensmitteltechnik, 1-2, 24–32.
Hancioglu, O., & Karapinar, M. (1997). Microflora of Boza, a traditional fermented Turkish beverage. International Journal of Food Microbiology, 35, 271–274.
Hébert, E. M., Raya, R. R., Tailliez, P., & De Giori, G. S. (2000). Characterization of natural isolates of Lactobacillus strains to be used as starter cultures in dairy fermentation. Inter-national Journal of Food Microbiology, 59, 19–27.
Heller, K. (2001). Prebiotic bacteria in fermented foods: product characteristics and starter organisms. American Journal of Clinical Nutrition, 73, 374–379.
Heu, S., Oh, J., Kang, Y., Ryu, S., Cho, S. K., Cho, Y., et al. (2001). Gly gene cloning and expression and purification of Glycinecin A, a bacteriocin produced by Xanthomonas campestris pv. Glycines 8ra. Applied and Environmental Microbiology, 67, 4105–4110.
Holzapfel, W. (1997). Use of starter cultures in fermentation on a household scale. Food Con-trol, 8, 241–258.
Holzapfel, H. W. (2002). Appropriate starter culture technologies for small-scale fermentation in developing countries. International Journal of Food Microbiology, 75, 197–212.
Hounhouigan, D., Nout, M., Nago, C., Houben, J., & Rombouts, F. (1993). Characterization and frequency distribution of species of lactic acid bacteria involved in the processing of mawe, a fermented maize dough from Benin. International Journal of Food Microbiology, 18, 279.
Ivanova, I., Kabadjova, P., Pantev, A., Danova, S., & Dousset, X. (2000). Detection, purification and partial characterization of a novel bacteriocin substance produced by Lactococcus lac-tis subsp. lactis B14 isolated from boza – Bulgarian traditional cereal beverage. Biocatal – Vestnik Moskov Univ Kimia, 41, 47–53.
Jahreis, G., Vogelsang, H., Kiessling, G., Schubert, R., Bunte, C., & Hammes, W. P. (2002). Influence of probiotic sausage (Lactobacillus paracasei) on blood lipids and immunologi-cal parameters of healthy volunteers. Food Research International, 35, 133–138.
Jaskari, J., Kontula, P., Siitonen, A., Jousimies-Somer, H., Mattila-Sandholm, T., & Pou-tanen, K. (1998). Oat-beta-glucan and xylan hydrolysates as selective substrates for Bifidobacterium and Lactobacillus strains. Applied Microbiology and Biotechnology, 49, 175–181.
Jiménez Vera, R., González, Cortés, N., Magaña Contreras, A., & Corona Cruz, A. (2010). Evaluación microbiológica y sensorial de fermentados de pozol blanco, con cacao (Theo-broma cacao) y coco (Cocos nucifera). Revista Venezolana de Ciencia y Tecnología de Alimentos, 1, 70–80.
Johansson, M., Molin, G., Jeppsson, B., Nobaek, S., & Ahrné, S. (1993). Administration of different Lactobacillus strains in fermented oatmeal soup: in vivo colonisation of human intestinal mucosa and effect on the indigenous flora. Applied and Environmental Microbi-ology, 59, 15–20.
Kabadjova, P., Gotcheva, I., Ivanova, I., & Dousset, X. (2000). Investigation of bacteriocin activity of lactic acid bacteria isolated from boza. Biotechnology & Biotechnological Equipment, 14, 56–59.
Karppinen, S. (2003). ‘Dietary fibre components of Rye bran and their fermentation’ in vitro (Ph.D. thesis). University of Helsinki.

150 Advances in Fermented Foods and Beverages
Keppler, K., Geiser, R., & Holzapfel, W. H. (1994). An a-amylase sensitive bacteriocin of Leuconostoc carnosum. Food Microbiology, 11, 39–45.
Kingamkono, R. R., Sjögren, E., & Svanberg, U. (1998). Inhibition of enterotoxin production by, and growth of enteropathogens in a lactic acid-fermenting cereal gruel. World Journal of Microbiology & Biotechnology, 14, 661–667.
Klaenhammer, T. (1988). Bacteriocins of lactic acid bacteria. Biochimie, 70, 337–349.Kleerebezem, M., Boekhorst, J., van Kranenburg, R., Molenaar, D., Kuipers, O. P., Leer, R.,
et al. (2003). Competitive genome sequence of Lactobacillus plantarum WCFS1. Proceed-ings of the National Academy of Sciences of the United States of America, 100, 1990–1995.
Köse, E., & Yücel, U. (2003). Chemical composition of Boza. Journal of Food Technology, 1, 191–193.
Leal-Sánchez, M. V., Jiménez-Díaz, R., Maldonado-Barragán, A., Garrido-Fernández, A., & Ruiz-Barba, J. L. (2002). Optimization of bacteriocin production by batch fermentation of Lactobacillus plantarum LPCO10. Applied and Environmental Microbiology, 68, 4465–4471.
LeBlanc, J. G., Taranto, M. P., Molina, V., & Sesma, F. (2010). B-group vitamins production by probiotic lactic acid bacteria. In F. Mozzi, R. Raya, & G. Vignolo (Eds.), Biotechnology of lactic acid bacteria: Novel applications (pp. 211–232). Ames, IA, USA: Wiley-Blackwell.
Lee, C. H. (1997). Lactic acid fermented foods and their benefits in Asia. Food Control, 8, 259–269.
Lepargneur, J. P., & Rousseau, V. (2002). Protective role of the Doderlein flora. Journal de Gynecologie Obstetrique et Biologie de la Reproduction, 31, 485–494.
Leroy, F., & De Vuyst, L. (2004). Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends in Food Science & Technology, 15, 67–78.
Lewus, C. B., Sun, S., & Montville, J. T. (1992). Production of an a-amylase sensitive bacte-riocin by an atypical Leuconostoc paramesenteroides strain. Applied and Environmental Microbiology, 58, 143–149.
Lorri, W., & Svanberg, U. (1995). Lower prevalence of diarrhoea in young children fed lactic acid-fermented gruels. Food and Nutrition Bulletin, 15, 57–63.
Louembé, D., Brauman, A., Tchicaya, F., & Kobawila, S. C. (1996). Etude microbiologique et biochimique de la bouillie de maïs “poto-poto”. Microbiologie, Aliments, Nutrition, 14, 245–253.
Louembé, D., Kéléke, S., Kobawila, S. C., & Nzouzi, J. P. (2003). Bactéries lactiques de la pâte fermentée de maïs au Congo. Tropicultura, 21, 3–9.
Marklinder, I., & Lonner, C. (1992). Fermentation properties of intestinal strains of Lactoba-cillus, of a sour dough and of a yoghurt starter culture in an oat-based nutritive solution. Food Microbiology, 9, 197–205.
Maxwell, M. J., Messer, S. A., Hollis, R. J., Boyken, L., Tendolkar, S., Diekema, D. J., et al. (2003). Evaluation for E test method for determining fluconazole and voriconazole MICs for 279 clinical isolates of Candida species infrequently isolated from the blood. Journal of Clinical Microbiology, 41, 1087–1090.
McKay, L. L., & Baldwin, K. A. (1990). Applications for biotechnology: present and future improvements in lactic acid bacteria. FEMS Microbiology Reviews, 87, 3–14.
Meinken, M., & Todorov, S. D. (2009). Atomic force microscopy on the effect of bacteriocins on target cells: a new method for visualising its mode of action. Trakia Journal of Sciences, 7, 28–32.
Mendez-Albores, J. A., Arambula-Villa, G., Preciado-Ortiz, R. E., & Moreno-Martinez, E. (2004). Aflatoxins in pozol, a nixtamalized, maize-based food. International Journal of Food Microbiology, 94, 211–215.

151Traditional cereal fermented foods as sources of functional microorganisms
Mohammed, S. I., Steenson, L. R., & Kirleis, A. W. (1991). Isolation and characterization of microorganisms associated with the traditional sorghum fermentation for production of Sudanese kisra. Applied and Environmental Microbiology, 57, 2529–2533.
Molin, G. (2001). Probiotics in foods not containing milk or milk constituents, with special reference to Lactobacillus plantarum 299v. American Journal of Clinical Nutrition, 73, 380–385.
Morcos, S. R., Hegazi, S. M., & El-Damhoughy, S. T. (1973). Fermented foods of common use in Egypt II. The chemical composition of bouza and its ingredients. Journal of the Science of Food and Agriculture, 24, 1157–1161.
Mugula, J., Narvhus, J., & Sørhaug, T. (2003). Use of starter cultures of lactic acid bacteria and yeasts in the preparation of togwa, a Tanzanian fermented food. International Journal of Food Microbiology, 83, 307–318.
Mugula, J. K., Nnko, S. A. M., & Sorhaug, T. (2001). Changes in quality attributes during stor-age of togwa, a lactic acid fermented gruel. Journal of Food Safety, 21, 181–194.
Nuraida, L., Wacher, M. C., & Owens, J. D. (1995). Microbiology of pozol, a Mexican fer-mented maize dough. World Journal of Microbiology & Biotechnology, 82, 783–790.
Odunfa, S. A., & Oyewole, O. B. (1997). African fermented foods. London: Blackie Academic and Professional, ISBN: 0751402168.
Oguntoyinbo, F. A., Tourlomousis, P., Gasson, M. J., & Narbad, A. (2011). Analysis of bacterial communities of traditional fermented West African cereal foods using culture independent methods. International Journal of Food Microbiology, 145, 205–210.
Phister, T. G., O’Sullivan, D. J., & McKay, L. L. (2004). Identification of bacilysin, chlorote-taine, and iturin a produced by Bacillus sp. strain CS93 isolated from pozol, a Mexican fermented maize dough. Applied and Environmental Microbiology, 70, 631–634.
Quillama, E. (1993). Estudio de la microflora de chicha de jora y su implicancia en la ali-mentación humana. Theorema Año 1, N° 2. Lima – Perú: UNMSM.
Quillama, E. (1998). Producción de bacteriocinas por cepas de Lactobacillus aisladas de chi-cha de jora. Lima, Peru: Facultad de Farmacia y Bioquímica. Universidad Nacional Mayor de San Marcos.
Quillama, E., Coha, J. M., García, R., Ramírez, P., Huamaní, D., Huisa, D., et al. (1995). Cinética de crecimiento y fermentación de levaduras nativas (Parte II). Theorema Año 4, N° 6. Lima – Perú: UNMSM.
Reid, G., & Burton, J. (2002). Use of Lactobacillus to prevent infections by pathogenic bacteria. Microbes and Infection, 4, 319–324.
Ross, P., Stanton, C., Hill, C., Fitzgerald, G. F., & Coffey, A. (2000). Novel cultures for cheese improvement. Trends in Food Science & Technology, 11, 96–104.
Sahingil, D., Isleroglu, H., Yildirim, Z., Akcelik, M., & Yildirim, M. (2011). Characterization of lactococcin BZ produced by Lactococcus lactis subsp. lactis BZ isolated from boza. Turkish Journal of Biology, 35, 21–33.
Salovaara, H. (1996). The time of cereal-based functional foods is here: introducing Yosa®, a vellie. In Proceedings of the 26th Nordic Cereal Congress, 12–15 May, Haugesund, Norway.
Salovaara, H. (2004). Lactic acid bacteria in cereal-based products. In S. Salminen, A. von Wright, & A. Ouwehand (Eds.), Lactic acid bacteria, microbiological and functional aspects (3rd ed.). (pp. 431–452). New York: Marcel Dekker.
Salovaara, H., & Simonson, L. (2003). Fermented cereal-based functional foods. Handbook of Fermented Food and Beverages. New York: Marcel Dekker.
Sanni, A. I. (1993). The need for process optimization of African fermented foods and bever-ages. International Journal of Food Microbiology, 18, 85–95.

152 Advances in Fermented Foods and Beverages
Shortt, C. (1999). The probiotic century: historical and current perspectives. Trends in Food Science & Technology, 10, 411–417.
Stark, A., & Madar, Z. (1994). Dietary fiber. In I. Goldberg (Ed.), Functional foods: Designer foods, pharmafoods, nutraceuticals (pp. 183–201). New York: Chapman and Hall.
Steinkraus, K. H. (1979). Nutritionally significant indigenous fermented foods involving an alcoholic fermentation. New York: Academic Press.
Thomas, L. V., Clarkson, M. R., & Delves-Broughton, M. (2002). Nisin. In A. S. Naidu (Ed.), Natural food antimicrobial systems (pp. 463–524). Boca Raton, Florida: CRC Press.
Todorov, S. D. (2010). Diversity of bacteriocinogenic lactic acid bacteria isolated from boza, a cereal-based fermented beverage from Bulgaria. Food Control, 21, 1011–1021.
Todorov, S. D., Botes, M., Guigas, C., Schillinger, U., Wiid, I., Wachsman, M. B., et al. (2008). Boza, a natural source of probiotic lactic acid bacteria. Journal of Applied Microbiology, 104, 465–477.
Todorov, S. D., & Dicks, L. M. T. (2004). Characterization of mesentericin ST99, a bacteriocin produced by Leuconostoc mesenteroides subsp. dextranicum ST99 isolated from boza. Journal of Industrial Microbiology & Biotechnology, 31, 323–329.
Todorov, S. D., & Dicks, L. M. T. (2005). Pediocin ST18, an anti-listerial bacteriocin produced by Pediococcus pentosaceus ST18 isolated from boza, a traditional cereal beverage from Bulgaria. Process Biochemistry, 40, 365–370.
Todorov, S. D., & Dicks, L. M. T. (2006). Screening for bacteriocin-producing lactic acid bacte-ria from boza, a traditional cereal beverage from Bulgaria. Comparison of the bacteriocins. Process Biochemistry, 41, 11–19.
Todorov, S. D., von Mollendorff, J. W., Moelich, E., Muller, N., Witthuhn, R. C., & Dicks, L. M. T. (2009). Evaluation of potential probiotic properties of Enterococcus mundtii, its survival in boza and in situ bacteriocin production. Food Technology and Biotechnology, 47, 178–191.
Tou, E. H., Guyot, J. P., Mouquet-Rivier, C., Rochette, I., Counil, E., Traoré, A. S., et al. (2006). Study through surveys and fermentation kinetics of the traditional processing of pearl mil-let (Pennisetum glaucum) into ben-saalga, a fermented gruel from Burkina Faso. Interna-tional Journal of Food Microbiology, 106, 52–60.
Tuncer, Y., & Ozden, B. (2010). Partial biochemical characterization of nisin – like bacteriocin produced by Lactococcus lactis subsp. lactis YBD11 isolated from boza, a traditional fer-mented Turkish beverage. Romanian Biotechnological Letters, 15, 4940–4948.
Ulloa, M., & Herrera, T. (1972). Description of 2 new species of bacteria isolated from pozol: Agrobacterium azotophilum and Achromobacter pozolis. Revista Latinoamericana de Microbiología, 14, 15–24.
Ulloa, M., Herrera, T., & Lappe, P. (1987). Fermentaciones tradicionales indigenas de Mexico. Serie de Investigaciones Sociales Mexico Instituto Nacional Indigenista, 16, 13–20.
Velraeds, M., Van De Belt-Gritter, B., Van Der Mei, H., Reid, G., & Busscher, H. (1998). Interference in initial adhesion of uropathogenic bacteria and yeast and silicone rub-ber by Lactobacillus acidophilus biosurfactant. Journal of Medical Microbiology, 47, 1081–1085.
Vogel, R. F., Knorr, R., Muller, M. R. A., Steudel, U., Gänzle, M. G., & Ehrmann, M. A. (1999). Non-diary lactic fermentations: the cereal world. Antonie van Leeuwenhoek, 76, 403–411.
Von Mollendorff, J. W., Todorov, S. D., & Dicks, L. M. T. (2006). Comparison of bacteriocins produced by lactic acid bacteria isolated from boza, a cereal-based fermented beverage from the Balkan Peninsula. Current Microbiology, 53, 209–216.
Wacher, C., Cañas, A., Bárzana, E., Lappe, P., Ulloa, M., & Owens, J. D. (2000). Microbiology of Indian and Mestizo pozol fermentations. Food Microbiology, 17, 251–256.

153Traditional cereal fermented foods as sources of functional microorganisms
Wacher, C., Cañas, A., Cook, P. E., Barzan, E., & Owens, J. D. (1993). Sources of microorgan-isms in pozol, a traditional Mexican fermented maize dough. World Journal of Microbiol-ogy & Biotechnology, 9, 269–274.
Weerkamp, A. H., Klijn, N., Neeter, R., & Smit, G. (1996). Properties of mesophilic lactic acid bacteria from raw milk and naturally fermented raw milk products. Netherlands Milk and Dairy Journal, 50, 319–332.
Wood, P. J., & Beer, M. U. (1998). Functional oat products. In G. Mazza (Ed.), Functional foods: Biochemical and processing aspect (pp. 1–29). Lankaster: Technomic Publishing Co., Inc.
Wrick, K. L. (1994). The potential role of functional foods in medicine and public health. In I. Goldberg (Ed.), Functional foods: Designer foods, pharmafoods, nutraceuticals (pp. 480–494). New York: Chapman and Hall.
Yegin, S., & Uren, A. (2008). Biogenic amine content of boza: a traditional cereal-based, fer-mented Turkish beverage. Food Chemistry, 111, 983–987.
Yoshizawa, K., & Ishikawa, T. (1979). Industrialization of sake fermentation. New York: Marcel Dekker Inc.
Yousif, N. M. K., Huch, M., Schuster, T., Cho, G. S., Dirar, H. A., Holzapfel, W. H., et al. (2010). Diversity of lactic acid bacteria from Hussuwa, a traditional African fermented sorghum food. Food Microbiology, 27, 757–768.
Zorba, M., Hancioglu, O., Genc, M., Karapinar, M., & Ova, G. (2003). The use of starter cul-ture in the fermentation of boza, a traditional Turkish beverage. Process Biochemistry, 38, 1405–1411.

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具